Introduction
Rates of prostate cancer incidence vary dramatically
in different geographic locations (1,2).
Dietary fat intake, which is sometimes used to help explain this
variance, is among the most widely studied dietary risk factors for
prostate cancer, yet its role in influencing cancer risk remains
controversial (3). We do know that
dietary fat composition may greatly influence the risk of prostate
cancer (4) as it has been observed
in prostate cancer cell lines that n-6 fatty acids, such as
linoleic acid and arachidonic acid (AA), promote cell
proliferation, whereas long-chain polyunsaturated n-3 fatty acids,
which are abundant, for example, in fish oil, inhibit cell
proliferation (5–8). These promotional and inhibitory
effects of n-6 and n-3 fatty acids, respectively, have also been
demonstrated in prostate carcinogenesis and progression in
vivo (9). Evidence is also
available from specimens obtained during radical prostatectomy that
AA turnover is 10 times greater in tumor tissue than it is in
normal prostate tissue (7).
Of the possible mechanisms responsible for
AA-induced cell proliferation in prostate cancer, one of the most
likely is the generation of certain eicosanoids, key mediators of
the inflammatory response (10).
They are produced by both tissue cells and tumor-infiltrating
leukocytes (10), and they may act
as autocrine and/or paracrine factors. Synthesized from
polyunsaturated fatty acids, eicosanoids play important biologic
roles in cell proliferation and tissue repair, blood clotting,
blood vessel permeability, inflammation, and immune cell behavior
(10). They fall into three
general groups: prostaglandins (PGs), including PGE2;
leukotrienes (LTs); and thromboxanes. They also are associated with
the enzymatic pathways of the cyclooxygenases (COX-1 and COX-2),
lipoxygenases (LOXs), and cytochrome P450.
The PGs are synthesized by the action of COX
enzymes: the constitutively expressed COX-1 and the inducible
COX-2. COX-2 can be stimulated by inflammatory mediators,
cytokines, growth factors, and tumor promoters and can be inhibited
by steroids and certain nonsteroidal anti-inflammatory drugs
(11). Higher than usual COX-2 and
PGE2 levels have been observed in numerous malignancies,
including those of the colon, lung, head and neck, breast,
pancreas, bladder, and prostate (11). Both AA and PGE2
stimulate cell proliferation and tumor growth in vitro in
PC-3 human prostate cancer cells (12). In PC-3, LNCaP, and DU145 prostate
cancer cell lines, upregulation of COX-2 and PGE2 has
been inversely correlated with apoptosis (13).
Relative to the effects of the PGs, those of the LOX
products, such as LTs and hydroxyeicosatetraenoic acids (HETEs),
are more diverse and may be cell-type specific. An AA metabolite
derived from 12-LOX, 12-HETE, promotes the proliferation of human
colon, pancreatic, and breast cancer cell lines and plays an
important role in cell adhesion and promotion of metastasis
(14). Expression of 12-LOX also
appears to stimulate angiogenesis in human prostate carcinoma cells
(14). More recently, study
results have suggested that 12-LOX functions as a potential
biomarker and therapeutic target for prostate cancer stem cells
(15). Further, the 5-LOX product
5-HETE has been suggested as playing a role as a potent pro-growth
survival factor for human prostate cancer cells (16).
By contrast, we and others have reported that
15-LOX-1, 15-LOX-2, and their related products
13-hydroxyoctadecadienoic acid (13-HODE) and 15-HETE actually
function as tumor suppressors in, respectively, colorectal and
prostate cancer cells (17,18).
In addition, 15-deoxy-Δ12,14-PGJ2, the
metabolite of PGJ2 and a peroxisome
proliferator-activated receptor γ (PPARγ) ligand, has been reported
to induce cell death in three human prostate cancer cell lines,
PC-3, LNCaP, and DU145 (19).
Thromboxanes, additional metabolites of COX enzymes,
have also been studied in cancer. For example, higher thromboxane
synthase mRNA levels have been observed in prostate and uterine
cancers relative to levels in matched normal tissues (20). In prostate cancer, higher
expression of thromboxane synthase was associated with
advanced-stage and high-grade disease (21). Thromboxane synthase expressed in
prostate cancer cells was enzymatically active and might play a
contributory role in tumor progression, especially in tumor cell
motility (22,23).
In vitro studies have demonstrated that
eicosanoids appear to have key roles in the biology of human
prostate cancer, but the role of endogenous eicosanoids in
vivo in prostate cancer development and progression remains
unclear. We and others have investigated the alteration of AA
metabolites in the TRAMP mouse model and found that 12-HETE and
COX-2 levels were significantly higher in prostate tissues of these
mice than they were in those of control or wild-type mice (24,25).
However, endogenous PGE2 was almost five times lower in
the prostate of TRAMP mice than it was in wild-type mice (24).
Shappell et al (26) found in human samples from radical
prostatectomy specimens that expression of COX-2 was elevated only
in high-grade tumors. By comparison, the reduction of 15-LOX-2 and
15-HETE formation is the most characteristic alteration of AA
metabolism in prostate cancer (26). We also reported that 15-LOX-2
expression was lost in all the prostate cancer cell lines we tested
(including PC-3, LNCaP, and DU145 cells), yet easily detectable
levels of 15-LOX-2 were expressed in normal human prostate cells
(17,27). Not surprisingly, eicosanoid
profiles differ between cancer and normal prostate cells and
between mouse and human prostate tissues.
Changes in eicosanoid content appear to be important
for a more complete understanding of prostate cancer progression.
We therefore undertook this study in androgen-dependent and
androgen-independent cell lines from bone, brain, and lymph node
metastases; in rodent xenografts of human tumors with bone
microenvironment influence; and in human tumor tissues. Our
findings from these studies show that eicosanoids may play an
important role in prostate cancer progression in a cell type- and
microenvironment-dependent manner.
Materials and methods
Eicosanoids and antibodies
The eicosanoids PGE2, 5-HETE, 12-HETE,
and 15-HETE, and the corresponding deuterated eicosanoid standards
were purchased from Cayman Chemical. AA, butylated hydroxytoluene
(BHT), citric acid, and EDTA were obtained from Sigma Chemical. All
high-pressure liquid chromatography (HPLC) solvents used for
analyses of eicosanoids were purchased from Fisher Scientific.
Anti-COX-2 and anti-15-LOX antibodies were obtained from Cayman
Chemical. Anti-5-LOX antibody was purchased from Research
Diagnostics and anti-β-actin antibody was purchased from Sigma
Chemical. Anti-platelet-type 12-LOX antibody was obtained from
Oxford Biomedical Research.
Cell lines
Human prostate cancer cell lines LNCaP, PC-3, and
DU145 were obtained from the American Type Culture Collection and
maintained in a humidified atmosphere containing 5% CO2
at 37°C. LNCaP and PC3 cells were routinely cultured in RPMI-1640
medium (Invitrogen), supplemented with 10% heat-inactivated fetal
bovine serum (FBS) (Hyclone Laboratories) containing 50 IU/ml
penicillin, 50 μg/ml streptomycin, and 2 mM L-glutamine [all from
Gibco (Invitrogen)]. DU145 cells were cultured under similar
conditions except that cells were grown in DMEM medium
(Mediatech).
The human prostate cancer cell lines MDA PCa 2a and
MDA PCa 2b were gifts from Dr Nora Navone (28). These cell lines were grown in
BRFF-HPC1 medium (AthenaES) supplemented with 20% FBS in tissue
culture flasks coated with FNC coating mix (AthenaES). All cell
lines were authenticated via microscopic morphology check and DNA
analysis.
Human prostate xenografts
All animal experiments were approved by the
Institutional Animal Care and Use Committee at MD Anderson.
Xenograft models of PC-3, LNCaP, DU145, and MDA PCa 2b cells were
developed as previously described (28). In brief, PC-3 and DU145
(1×106), LNCaP (5×106), and MDA PCa 2b
(1×107) cells were suspended in 50% Matrigel (BD
Biosciences) and subcutaneously implanted into the flanks of
8-week-old male BALB/c athymic (Nu/Nu) mice. Mice were euthanized
with CO2 approximately 1 month later, when the tumor
volume reached 1 cm3. A portion of each tumor was snap
frozen and stored at −80°C until measurements of eicosanoid
profiles and protein expression of eicosanoid enzymes were
obtained.
Human core biopsy specimens from
prostatectomy specimens
Ex vivo biopsy specimens were obtained from
eight patients undergoing prostatectomy who consented to the use of
their tissues in research, according to the Institutional Tissue
Banking protocol at MD Anderson. Whenever possible, multiple core
biopsy specimens were obtained from each prostatectomy specimen.
The presence of tumor in each core was determined by making a touch
prep of the cores before the specimens were snap frozen in liquid
nitrogen, placed in cryovials, and archived at −80°C in the
Prostate Tissue Bank.
Technicians ensured conformance with the
prostatectomy specimen map delineating individual tumor foci. Seven
core biopsy specimens containing tumor and three without tumor were
used to test for the expression of COXs and LOXs; seven core biopsy
specimens without tumor and 19 with tumor were used for evaluating
eicosanoid profiles. In this proof-of-principle analysis, the
limited amount of tissue prevented the testing of matching
specimens across all two-sample types and evaluating intrapatient
as well as interpatient variabilities.
RNA extraction and real-time polymerase
chain reaction
Total RNA was extracted from the five prostate
cancer cell lines using TRI reagent (Molecular Research Center).
The RNA from each sample was reverse transcribed and then measured
quantitatively using real-time polymerase chain reaction (PCR) and
a comparative threshold cycle method, as previously described
(29,30).
Western blot analysis
Cells growing in log phase were washed with cold PBS
and scraped free in the presence of a lysis buffer (Invitrogen)
with protease inhibitor cocktail (Sigma). Cell lysates were then
sonicated on ice for 3 min, incubated for 10 min on ice, and
centrifuged at 14,000 × g for 10 min at 4°C.
Protein levels were quantified via the
detergent-compatible protein assay (Bio-Rad). Equal levels of
protein (60 μg) were fractionated on precast gels (Bio-Rad) and
then transferred onto polyvinylidene diflouride membranes,
according to standard methods. After a 1- to 2-h incubation in 5%
nonfat dry milk blocking buffer prepared in Tris-buffered saline
with 0.1% Tween 20, the membranes were probed with primary
antibodies diluted 1:2,000 in blocking buffer. Protein bands were
visualized via chemiluminescence, using the ECL+ detection kit
(Amersham Biosciences) and hyper-film (Amersham Biosciences). Equal
loading of samples was illustrated by Western blotting for the
presence of β-actin.
Eicosanoid analyses
The levels of eicosanoids in the prostate cancer
cells, xenograft tissues, and human biopsy samples were determined
according to the methods of Kempen et al and Yang et
al (31–33). In brief, the human prostate cancer
cells (5×106) were harvested by trypsinization, washed
with PBS, and then resuspended in 0.5 ml of PBS containing 1 mM
CaCl2. For exogenous eicosanoid analysis, samples were
incubated with 2.5 μl of calcium ionophore A23187 (1 mM; Sigma) for
2 min at 37°C, followed by addition of an aliquot of 2.5 μl of 10
mM AA. Samples were then incubated for a further 10 min under
similar conditions. The reaction was terminated by the addition of
aliquots of 40 μl of 1 N citric acid and 5 μl of 10% BHT. An
aliquot of 10 μl of the relevant deuterated eicosanoids
(PGE2-d4, 15-HETE-d8,
12-HETE-d8, and 5-HETE-d8) per 100 ng/ml) was
added to the reaction mixtures as internal standards. The
eicosanoids were extracted three times with 2 ml of hexane-ethyl
acetate (1:1, v/v). The upper organic phases were then pooled and
evaporated to dryness under a stream of nitrogen at room
temperature. All extraction procedures were performed under
conditions of minimal light. Samples were then reconstituted in 100
μl of methanol with 10 mM ammonium acetate buffer (70:30, v/v; pH,
8.5) before analysis by liquid chromatography/tandem mass
spectrometry (LC/MS/MS). For the measurement of endogenous
eicosanoids, the resuspended cells were acidified by the addition
of an aliquot of 40 μl of 1 N citric acid followed by the addition
of the deuterated internal standards and extraction of eicosanoids
as previously described (33).
For analysis of eicosanoids in the xenograft
tissues, each frozen tissue specimen (25–50 mg) was ground to a
fine powder in a liquid nitrogen-cooled mortar (Fisher). Samples
were then transferred to microcentrifuge tubes. Ice-cold PBS
buffer, triple each sample’s volume and containing 0.1% BHT and 1
mM EDTA, was added, and the tubes were sealed. The mixtures were
then homogenized using an ultrasonic processor (Misonix) at 0°C for
3 min. A 100-μl aliquot of each homogenate was transferred to a
glass tube (13×100 mm), and the eicosanoids were extracted using
the method of Yang et al (33).
We next measured the AA metabolites in the human
prostate core biopsy samples using a modification of the method of
Yang et al (33). In brief,
the frozen specimens were mixed with 150 μl of homogenization
buffer [500 mM Tris-HCl (pH 7.2), 0.5 M sucrose, 200 M EDTA, 100 mM
EGTA, 0.4 M NaF, 10% Triton X-100, and 10 mM sodium orthovanadate
(Sigma)] and incubated at 0°C for 30 min. The samples were then
homogenized and processed for eicosanoid extraction as described
above.
Reverse-phase HPLC electrospray ionization MS was
used to measure eicosanoid (PGE2, 5-HETE, 12-HETE)
levels in cells using our previously published method (31,34).
A Micromass Quattro Ultima tandem mass spectrometer (Waters) was
equipped with an Agilent 1100-HP binary pump HPLC inlet for use in
these studies. Eicosanoids were separated by a Luna 3-μ
phenyl-hexyl (2×150 mm) LC column (Phenomenex). The mobile phase
consisted of 10 mM ammonium acetate (pH 8.5) and methanol; the flow
rate was 250 μl/min, the column temperature was 50°C, and the
sample injection volume was 25 μl. Samples were kept at 4°C in an
autosampler before their injection into the analytical column. The
mass spectrometer was operated in the electrospray negative-ion
mode with a cone voltage of 100 V. Fragmentation of all compounds
was performed using argon as the inert collision gas at a cell
pressure of 2.1×10−3 torr. The collision energy was 19
V. All eicosanoids were detected using negative ionization and
multiple-reaction monitoring of the transition ions for eicosanoid
products and their internal standards (33).
Statistical analyses
Descriptive statistics were calculated and
exploratory data analyses were performed. Categorical data were
summarized by frequency counts, proportions, or percentages.
Continuously scaled measures were summarized as means and standard
deviations or as medians with ranges. Gene expression image plots,
bar graphs, and scatter plots were used to graphically present the
data. Student’s t-test was used for analyzing continuous variables.
A mixed-effects model was used in the analysis of repeated
measurements from the same patients. All statistical tests were two
sided, and p-values of <0.05 were considered to indicate
statistically significant differences. Statistical analyses were
performed with S-PLUS software (TIBCO Software).
Results
AA metabolism enzyme expression in human
prostate cancer cell lines
The data from the quantitative analysis of mRNA of
eicosanoid enzymes in the five different human prostate cancer cell
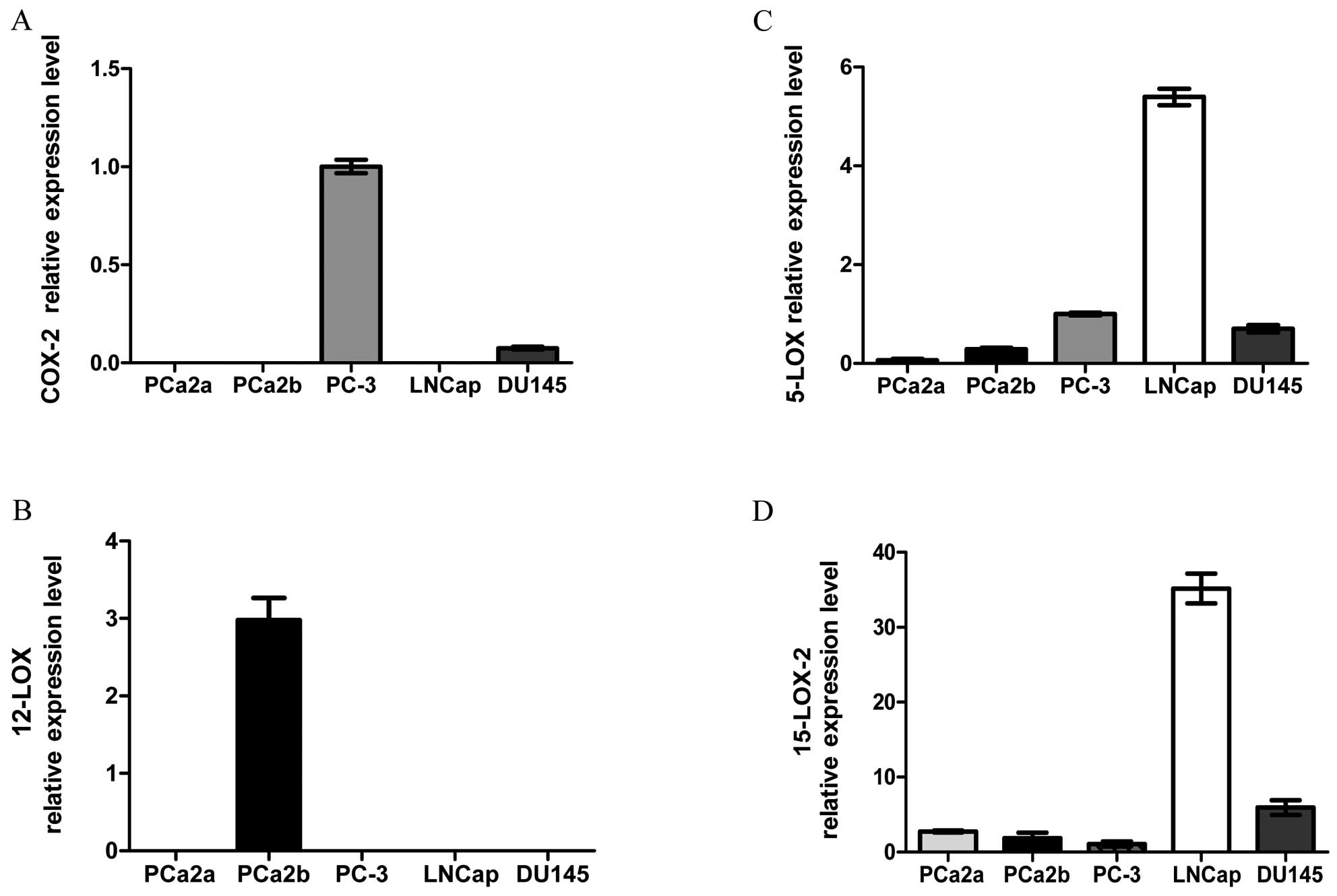
lines are shown in Fig. 1. COX-2
mRNA was detectable only in PC-3 and DU145 cells, with PC-3 cells
containing 10 times more than DU145 cells (Fig. 1A). By contrast, levels of 12-LOX
mRNA were observed only in the MDA PCa 2b metastatic prostate
cancer cells (Fig. 1B). Notably,
the level of 5-LOX mRNA in LNCaP cells was almost five times higher
than that in PC-3 cells, whereas very limited expression of 5-LOX
was detected in MDA PCa 2a and MDA PCa 2b cells (Fig. 1C). The mRNA level of 15-LOX-2 was
also highest in the LNCaP cells, relative to that in the other
cells tested (Fig. 1D).
Expression of COX-2 and the LOXs in
prostate cancer cell lines
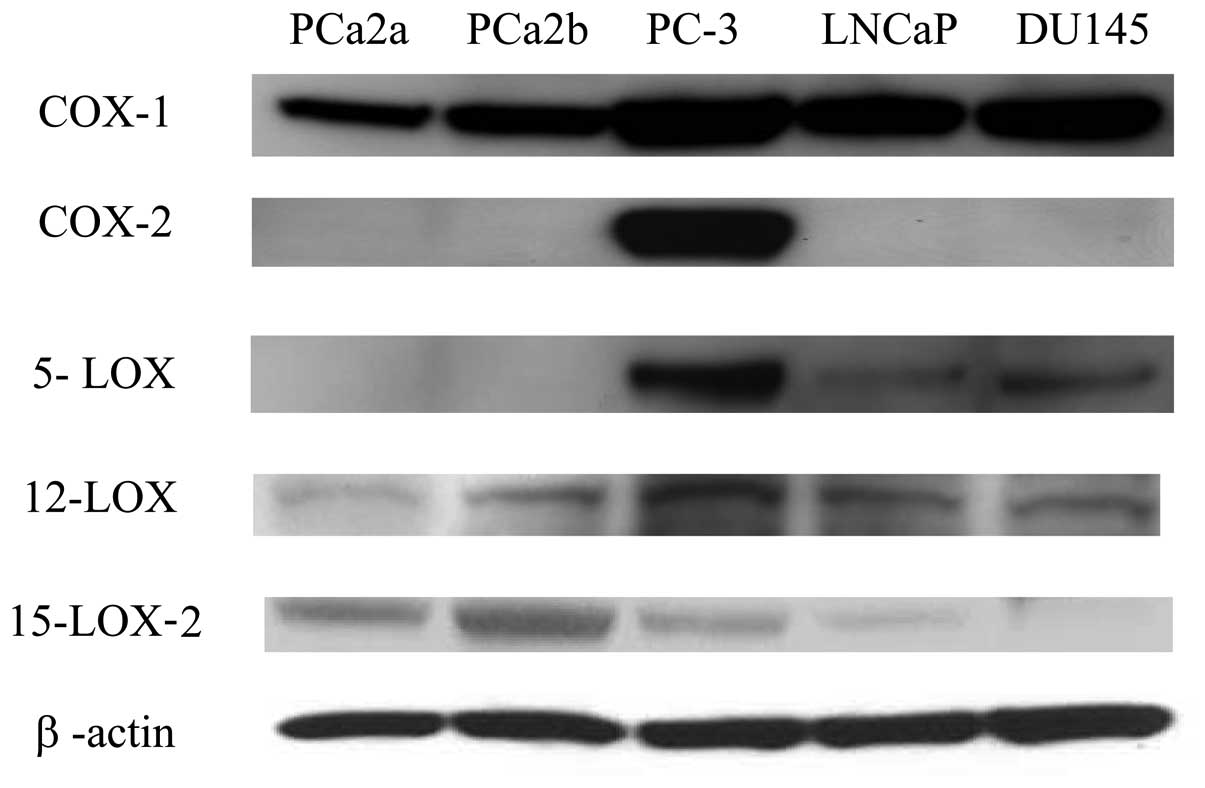
The protein expression of COX-2 and LOX enzymes was
examined in the five different prostate cancer cell lines by
Western blotting to confirm that the RNA expression of these
enzymes had resulted in protein expression. Although the protein
expression of COX-2 in PC-3 cells was similar to that of mRNA in
this cell line, the protein expression levels of 5-LOX, 12-LOX, and
15-LOX-2 were somewhat different from that of mRNA (Fig. 2). This could be reflective of
either assay differences or, more importantly, differences in mRNA
translation efficiency between the various cancer cell lines.
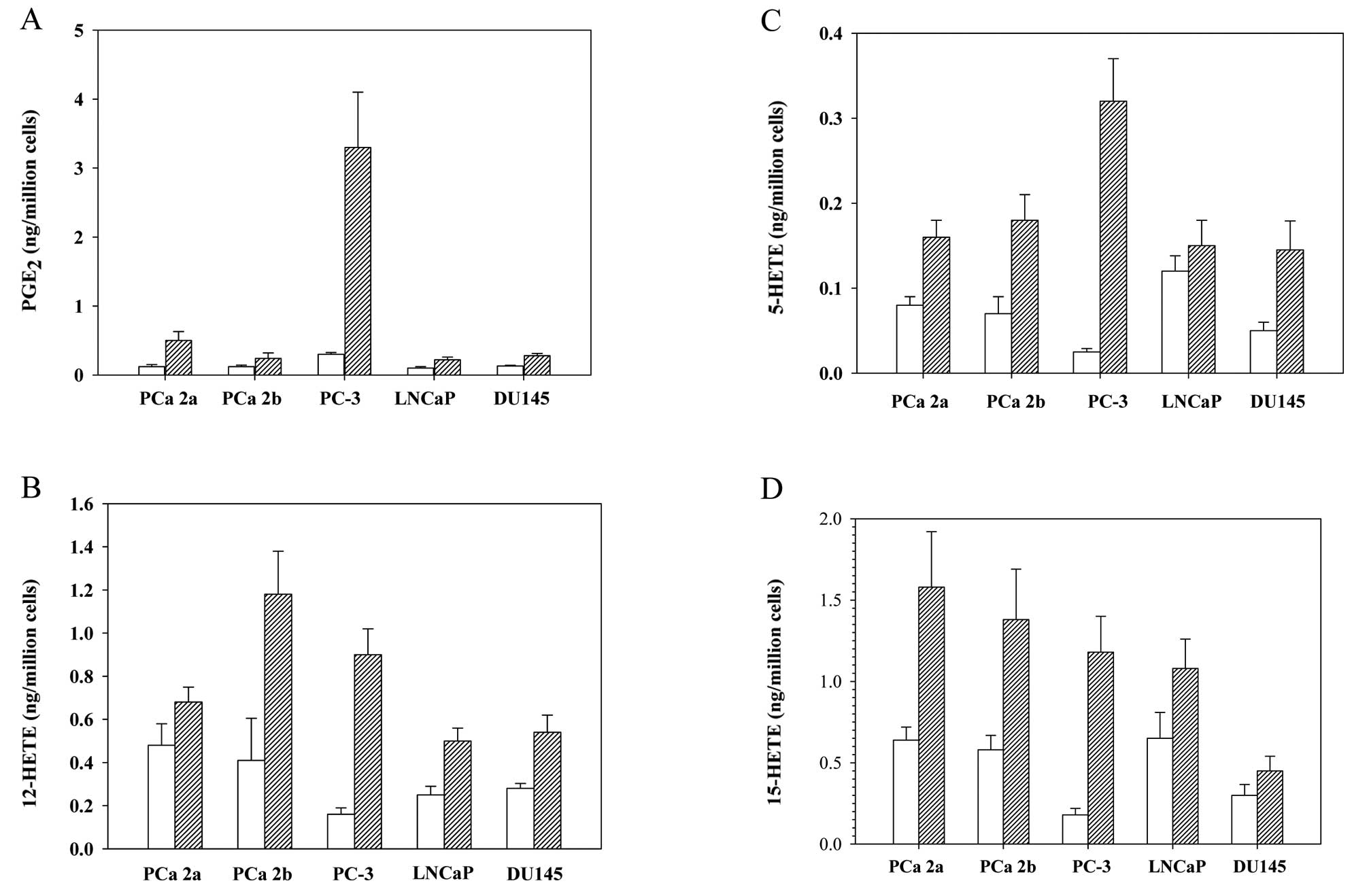
Endogenous and exogenous AA metabolism in
the human prostate cancer cell lines
The mRNA and protein levels of each eicosanoid
enzyme were differently regulated in the five prostate cancer cell
lines we tested, which suggested that the relevant metabolites of
the eicosanoids may also differ. Therefore, we measured both
endogenous and exogenous eicosanoid levels in those cell lines. As
shown in Fig. 3A, levels of both
endogenous and exogenous PGE2, a COX-2 metabolite, were
highest in the PC-3 cells, suggesting a high capacity to produce
this pro-inflammatory eicosanoid. The level of exogenous
PGE2 in PC-3 cells was almost 15 times higher than that
produced endogenously.
By comparison, the levels of endogenous 12-HETE in
the metastatic prostate cancer cells MDA PCa 2a (0.48±0.10
ng/million cells) and MDA PCa 2b (0.41±0.19 ng/million cells) were
almost twice those in the PC-3, DU145, and LNCaP cells (0.16–0.25
ng/million cells) (Fig. 3B). When
AA was used as a supplement for the cells, the formation of 12-HETE
was markedly increased in the MDA PCa 2b and PC-3 cells. The level
of exogenous 12-HETE was ranked from highest to lowest in the MDA
PCa 2b, PC-3, MDA PCa 2a, DU145, and LNCaP cells.
The level of endogenous 5-HETE was highest in the
LNCaP cells, but the exogenous level of 5-HETE was noticeably
higher in the PC-3 cells, in fact, more than 1.5 times the levels
in the other cell lines (Fig. 3C).
This suggests that the capacity to produce 5-LOX is greater in PC-3
cells than in the other cancer cell lines tested, and it correlated
with the highest expression of the 5-LOX protein in this particular
cell line (Fig. 2). The level of
endogenous 15-HETE was also highest in LNCaP cells, but after
exposure of the cell lines to AA, the MDA PCa 2a cells showed the
highest level of 15-HETE formation (Fig. 3D).
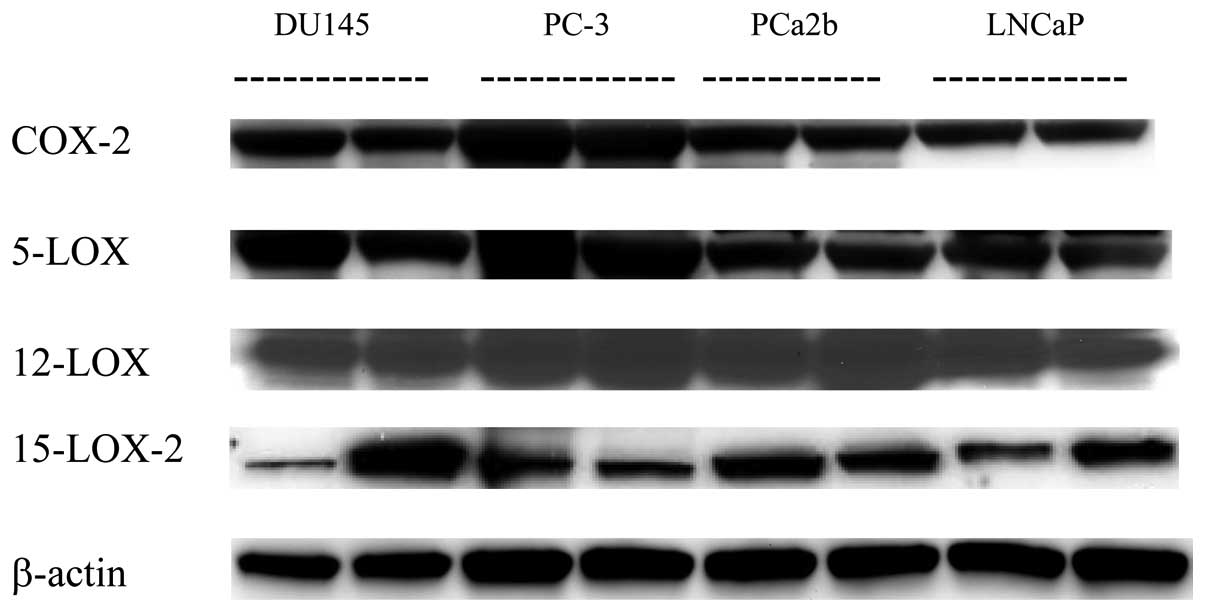
Expression of COX-2 and the LOXs in human
prostate cancer xenograft tissues
To explore the role of eicosanoids in prostate
cancer progression, we further investigated the expression of COX-2
and various LOX proteins in human prostate xenograft tissues
(Fig. 4, top panel). As the
tumorigenicity of the MDA PCa 2a cells was extremely low, we
evaluated the protein expression of eicosanoid enzymes in only the
other four human prostate cancer xenograft tissues. The protein
expression of the COX-2 and 5-LOX enzymes was highest in the PC-3
xenografts, whereas the protein levels of 12-LOX were higher in
both PC-3 and PCa 2b cells. In line with the protein expression of
15-LOX-2 in the cells in vitro, its expression was
consistently highest in the two tumor tissues derived from MDA PCa
2b xenografts (Fig. 4, top
panel).
Endogenous AA metabolism in human
prostate cancer xenograft tissues
AA metabolites were further examined in xenografts
of the DU145, LNCaP, PC-3, and MDA PCa 2b human prostate cancer
cells (Fig. 4, bottom panel). In a
pattern similar to that observed in the cultured prostate cancer
cell lines, the concentration of endogenous PGE2 was
significantly higher in PC-3 xenograft tissues (10.5±2.17 ng/mg
protein) than it was in the other three xenograft tissues (range,
0.07–1.1 ng/mg of protein) (p<0.001) (Fig. 4A, bottom panel). The levels of
12-HETE in both PC-3 and MDA PCa 2b xenograft tissues were also
much higher (13.9 and 10.9 ng/mg protein, respectively) than those
in the LNCaP xenograft tissues (3.9 ng/mg protein) (p<0.01)
(Fig. 4B, bottom panel). The
highest levels of both 5- and 15-HETE in PC3 and MDA PCa 2b tumor
tissues were only about one third or one fourth (2.07±0.46 for
5-HETE and 3.24±0.45 ng/mg protein) the levels of PGE2
and 12-HETE (Fig. 4C and D, bottom
panel). These results appear to correspond with the endogenous
levels of these AA metabolites in the same prostate cancer cells
in vitro.
Expression of COXs and LOXs in prostate
core biopsy specimens containing tumor
To test the role of AA metabolism in prostate
cancer, the expression of COX and LOX proteins in human prostate
core biopsy specimens containing tumor was compared with that in
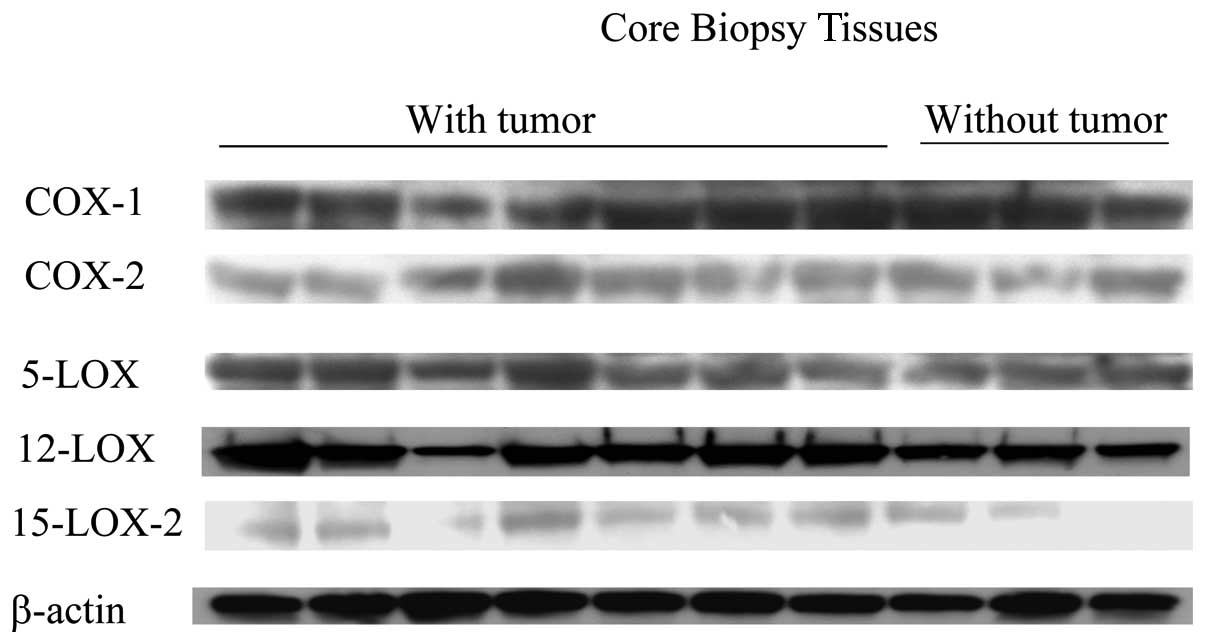
normal, nontumorous prostate tissue specimens. As shown in Fig. 5, the expression of COX-1 and COX-2
enzymes did not differ between the tumorous and normal tissues. In
almost all of the tumorous specimens, the expression of 12-LOX was
moderately greater than it was in the specimens of normal tissue.
The protein levels of 5-LOX were also greater in the cores
containing tumor than in those without, but the magnitude of the
increases was not as remarkable as that in the case of 12-LOX. We
found that the expression of 15-LOX-2 was slightly higher in most
of the tumorous cores than it was in the normal cores.
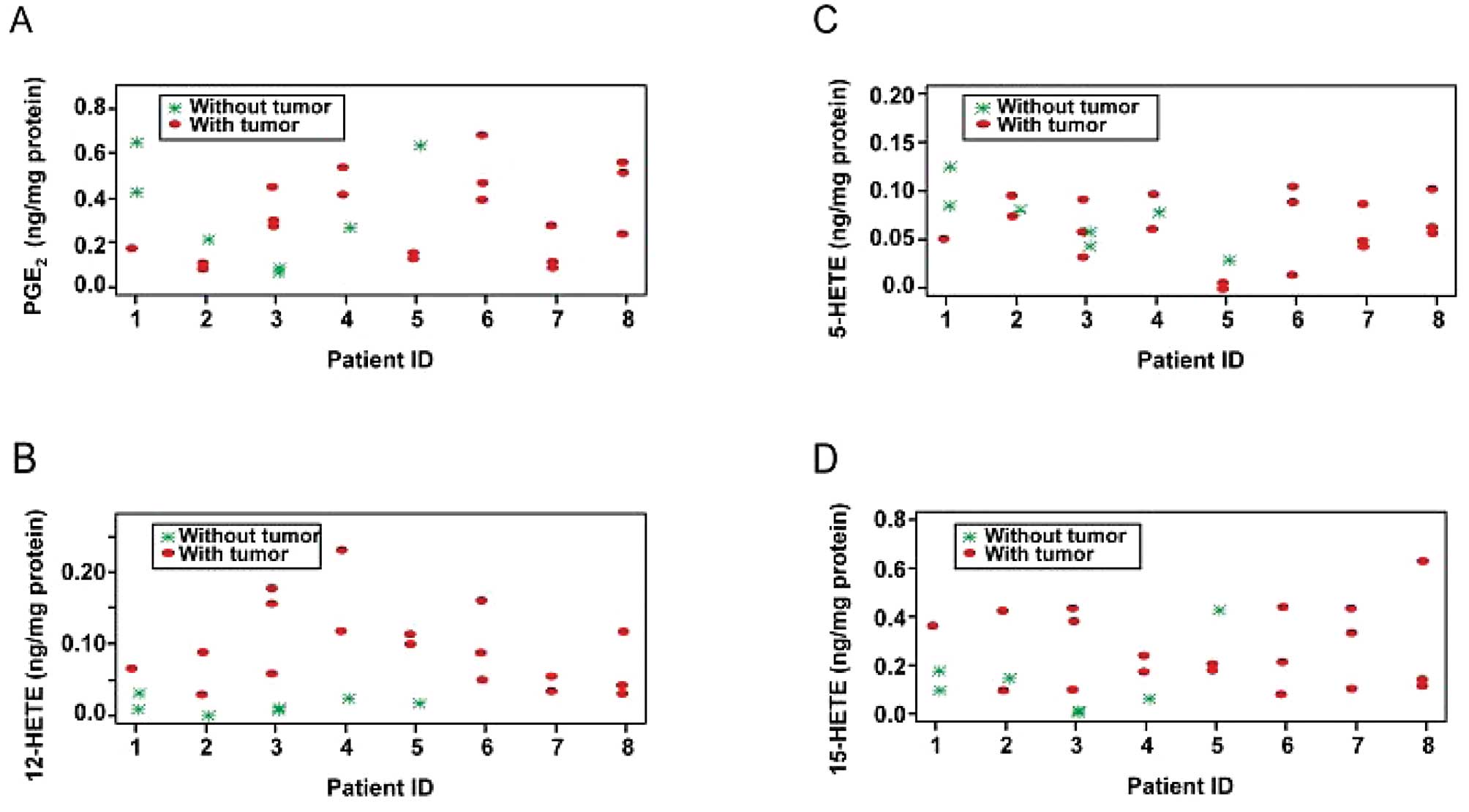
Eicosanoid profile in human prostate core
biopsy samples
AA metabolism was also examined in seven specimens
without tumor and 19 specimens containing tumor obtained from core
biopsies of prostatectomy specimens from eight patients. A
mixed-effects linear model was fitted to assess the differences
between the tumorous and normal specimens. The mean PGE2
levels in the non-tumor-containing and tumorous specimens were very
similar, as shown in Fig. 6A
(0.335 ng/mg and 0.314 ng/mg protein, respectively; p=0.703). The
mean level of endogenous 12-HETE was significantly higher in the
cores containing tumor than it was in those without (0.094 ng/mg
and 0.010 ng/mg protein, respectively; p=0.019) (Fig. 6B).
As with the PGE2 levels, the mean 5-HETE
levels were very similar in tumorous and normal tissues (0.062
ng/mg and 0.073 ng/mg protein, respectively; p= 0.483) (Fig. 6C). However, the mean level of the
15-LOX-2 metabolite 15-HETE in the tumorous cores was more than
twice that in the normal cores, although the difference was not
statistically significant (0.269 ng/mg and 0.129 ng/mg protein,
respectively; p=0.110) (Fig.
6D).
The changes in AA metabolism in the core biopsy
specimens containing malignant cells were similar to those observed
both in prostate cancer cells in vitro and in prostate
cancer xenograft tissues, suggesting that 12-LOX may play an
important role in the progression of human prostate cancer.
Discussion
AA-derived COX and LOX metabolites play a critical
role in prostate cancer progression. In this study, we found higher
levels of 12-HETE in the androgen-independent metastatic prostate
cancer cells and their corresponding xenograft tissues than in the
androgen-dependent prostate cancer cells. More striking was the
finding that among the five AA metabolites examined, the level of
endogenous 12-HETE was significantly more elevated in core biopsy
specimens containing tumor than it was in those without tumor. Our
results also suggest that the COX and LOX enzyme metabolic pathways
are modulated differently by exogenous and endogenous AA and that
this modulation may be cell type and microenvironment specific.
Previous evidence from specimens obtained during
radical prostatectomy showed that AA turnover is 10 times greater
in tumor tissue than in normal prostate tissue (7). In addition, the results of multiple
studies have supported the idea that AA has a role in the
proliferation of prostate cancer cells (8,35,36).
Investigators in one recent study reported that AA is critical in
modulating the cross talk between prostate carcinoma and bone
stromal cells that is mediated by transforming growth factor α,
interleukin 1β, and the receptor activator for nuclear factor κB
ligand (37), suggesting that AA
plays a role in the metastasis of prostate cancer to bone.
Whether COX-2 is involved in prostate cancer
progression is controversial. A higher level of expression of COX-2
or its metabolite PGE2 has consistently been observed in
premalignant lesions of high-grade prostatic intraepithelial
neoplasia (PIN) and in benign hyperplastic prostate lesions than is
expressed in normal prostate tissues (25,38–40).
However, COX-2 expression in cancerous prostate lesions or in more
invasive or metastatic disease is more controversial. Larre et
al (40) reported having found
no difference in PGE2 production between cancerous
prostate samples and adjacent normal tissues. The results of our
preclinical evaluations (25)
showed that the level of endogenous PGE2 was almost five
times lower in prostate tissue from TRAMP mice than it was in
wild-type mice. Earlier, Shappell et al (26) had examined the expression of COX-2
and 15-LOX-2 mRNAs and their related eicosanoid products in human
prostate biopsy specimens obtained during radical prostatectomy and
found that COX-2 expression was elevated only in high-grade tumors.
Our results are in agreement with the previous reported findings
with respect to the role of PGE2 in prostate cancer
progression.
Unlike the case with COX-2, the influence of the
LOXs (particularly 5-LOX and 12-LOX) and their metabolites on the
progression of prostate cancer has been extensively investigated.
We and others have examined alterations of the AA metabolites in
the TRAMP mouse model and found that 12-HETE was significantly
greater in the prostate tissues of these mice than it was in
control or wild-type mice (25,41).
The protein and mRNA of 12-LOX are overexpressed in human prostate
cancer, and its staining intensity on immunohistochemical testing
corresponded with that of advanced-stage or high-grade cancer
(42). Furthermore, other
investigators have suggested that 12-LOX is related to the
metastatic potential of prostate cancer since the expression of
12-LOX was higher in prostate cancer cell lines that metastasized
in their corresponding xenograft models (e.g., DU145) than its
expression was in PC-3 nm, a nonmetastatic subline of the PC-3 cell
line (43). Overexpression of
12-LOX also promotes the angiogenic activity and invasion and
metastatic potential of PC-3 cells. When PC-3 cells being
transiently transfected with 12-LOX were injected into
immunodeficient mice, the cells grew more rapidly and were
significantly more angiogenic than nontransfected control PC-3
cells (44). In addition, we
previously found that Zyflamend, a multiherbal anti-inflammatory
product (New Chapter) had a strong ability to downregulate both
12-LOX and 12-HETE and that this correlated with the product’s
ability to inhibit proliferation of PC-3 prostate cancer cells
(33). This new study demonstrates
for the first time the overexpression of 12-LOX and the higher
level of production of its AA metabolite 12-HETE (relative to its
production in normal prostate tissues) in highly metastatic
prostate cancer cells, at least PC-3 and MDA PCa 2b cells in
vitro, as well as in corresponding xenografts and tissues from
malignant human prostate tumors. These findings thus suggest that
12-LOX and its product 12-HETE have important roles in prostate
cancer and moreover that they may be useful as targets in the
treatment and prevention of prostate cancer.
It has been suggested (45) that the 5-LOX and 12-LOX pathways
are even more potent than the COX-2 pathways and that inhibiting
these LOX pathways would prove a more effective therapeutic
strategy than inhibiting COX-2 pathways. Our findings agree. Among
our five prostate cancer cell lines and their four xenograft
models, only PC-3 cells and the related xenograft models showed
relatively high levels of COX-2 expression and formation of its
metabolite PGE2. Furthermore, the COX-2 and
PGE2 levels in the core biopsy specimens containing
tumor were similar to those of the core biopsy specimens without
tumor. The expression of mRNA and protein of COX-2 correlated well
with the level of PGE2 in the prostate cancer cells, the
related xenograft tumors, and the human prostate biopsy specimens
with tumor. By contrast, the LOX enzyme expression and metabolite
levels in the different materials (i.e., cultured cells, xenograft
models, and core biopsy specimen cores with tumors) were highly
elevated in a number of the prostate cancer cell lines tested. The
notable variation in LOX expression and formation of its products
under different microenvironmental conditions suggested that
modulation of COX and LOX enzyme pathways differs, depending on
their microenvironments and whether the source of AA is exogenous
or endogenous. This, in turn, suggests that therapeutic
interventions targeting these pathways will be more effective than
inhibiting a specific enzyme. For example, the mRNA levels of 5-,
12-, and 15-LOXs appear to be positively correlated with the
endogenous levels of their relevant metabolites, whereas the
exogenous levels of LOX products were proportional to those of the
LOX protein (Figs. 1–3). This discord might be due to the
sensitivity of antibodies related to each LOX of interest; the
cofactors needed for production of eicosanoids, and the relative
affinities of each enzyme for their substrate, AA. Thus, these data
suggest that evaluating the expression of these eicosanoid enzymes
as well as their relevant metabolites in the human biopsy samples
is important for delineating the role of these lipid mediators in
the development and progression of prostate cancer.
In spite of our efforts and those of other
investigators, the involvement of 15-LOX in the initiation and
development of prostate cancer remains undefined. It appears that
two isoforms of 15-LOX, designated 15-LOX-1 and 15-LOX-2, have
different functions in tumor cell growth and survival. For example,
the level of 15-LOX-1 is thought to be higher in high-grade PIN
lesions than in normal prostate tissue (45), and its overexpression may reflect
its activity as a promoter of the development of PIN lesions
(46). By contrast, the expression
of 15-LOX-2 is noted to be lower in human prostate cancer and
high-grade PIN than in normal tissue (47,48).
Also, 15-LOX-2 expression was lost in all the prostate cancer cell
lines tested by Tang et al (17) and Bhatia et al (27), including PC-3, LNCaP, and DU145
cells, although it was expressed in all the normal human prostate
cells. Other investigators have suggested that the 15-LOX product
15-HETE plays a role in suppressing apoptosis. Our results from
this study demonstrate that 15-HETE levels and 15-LOX-2 expression
were actually higher in androgen-independent prostate cancer cells,
such as MDA PCa 2a and MDA PCa 2b cells, than they were in the
androgen-dependent LNCaP cells; furthermore, expression levels were
higher in prostate tumor-containing tissues than they were in the
tissues without tumor. Thus, it is apparent that the roles of
15-LOX and its metabolites in the progression of human prostate
cancer require further investigation.
The results of evaluating endogenous and exogenous
AA metabolites using human prostate cancer cell lines, their
corresponding xenograft models, and core biopsy specimens obtained
during prostatectomy suggest important roles for these metabolites,
particularly those generated through the LOX pathways. These
metabolites may serve as valuable markers of prostate cancer
prognosis and/or response to prostate cancer therapy, and their
pathways may be exploitable as potential therapeutic targets. Owing
to the limited number of biopsy specimens we tested in this study,
however, the important role of these metabolites, especially the
12- and 15-LOXs and their relevant metabolites, in the development
of prostate cancer merit further investigation in a large
population study.
Abbreviations:
|
AA
|
arachidonic acid
|
|
BHT
|
butylated hydroxytoluene
|
|
COX
|
cyclooxygenase
|
|
FBS
|
fetal bovine serum
|
|
HETE
|
hydroxyeicosatetraenoic acid
|
|
13-HODE
|
13-hydroxyoctadecadienoic acid
|
|
HPLC
|
high-pressure liquid
chromatography
|
|
LC/MS/MS
|
liquid chromatography/tandem mass
spectrometry
|
|
LT
|
leukotriene
|
|
LOX
|
lipoxygenase
|
|
PCR
|
polymerase chain reaction
|
|
PG
|
prostaglandin
|
|
PIN
|
prostatic intraepithelial
neoplasia
|
|
PPARγ
|
peroxisome proliferator-activated
receptor γ
|
Acknowledgements
We thank Karen F. Phillips, ELS, and
Beth W. Allen for their excellent editorial assistance. We also
thank Diana Chan for her technical assistance. This work was
supported in part by The University of Texas MD Anderson Cancer
Center Prostate Cancer Research Program and in part by the National
Institutes of Health through MD Anderson’s Cancer Center Support
Grant, CA016672.
References
|
1
|
Baade PD, Youlden DR and Krnjacki LJ:
International epidemiology of prostate cancer: geographical
distribution and secular trends. Mol Nutr Food Res. 53:171–184.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Coleman MP, Quaresma M, Berrino F, et al:
Cancer survival in five continents: a worldwide population-based
study (CONCORD). Lancet Oncol. 9:730–756. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Crowe FL, Key TJ, Appleby PN, et al:
Dietary fat intake and risk of prostate cancer in the European
Prospective Investigation into Cancer and Nutrition. Am J Clin
Nutr. 87:1405–1413. 2008.
|
|
4
|
Bidoli E, Talamini R, Bosetti C, et al:
Macronutrients, fatty acids, cholesterol and prostate cancer risk.
Ann Oncol. 16:152–157. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rose DP and Connolly JM: Effects of fatty
acids and eicosanoid synthesis inhibitors on the growth of two
human prostate cancer cell lines. Prostate. 18:243–254. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pandalai PK, Pilat MJ, Yamazaki K, Naik H
and Pienta KJ: The effects of omega-3 and omega-6 fatty acids on in
vitro prostate cancer growth. Anticancer Res. 16:815–820.
1996.PubMed/NCBI
|
|
7
|
Chaudry AA, Wahle KW, McClinton S and
Moffat LE: Arachidonic acid metabolism in benign and malignant
prostatic tissue in vitro: effects of fatty acids and
cyclooxygenase inhibitors. Int J Cancer. 57:176–180. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Anderson KM, Wygodny JB, Ondrey F and
Harris J: Human PC-3 prostate cell line DNA synthesis is suppressed
by eicosatetraynoic acid, an in vitro inhibitor of arachidonic acid
metabolism. Prostate. 12:3–12. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Berquin IM, Min Y, Wu R, et al: Modulation
of prostate cancer genetic risk by omega-3 and omega-6 fatty acids.
J Clin Invest. 117:1866–1875. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Higgs GA, Moncada S and Vane JR:
Eicosanoids in inflammation. Ann Clin Res. 16:287–299. 1984.
|
|
11
|
Wallace JM: Nutritional and botanical
modulation of the inflammatory cascade--eicosanoids,
cyclooxygenases, and lipoxygenases--as an adjunct in cancer
therapy. Integr Cancer Ther. 1:7–37. 2002.PubMed/NCBI
|
|
12
|
Hughes-Fulford M, Chen Y and Tjandrawinata
RR: Fatty acid regulates gene expression and growth of human
prostate cancer PC-3 cells. Carcinogenesis. 22:701–707. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Subbarayan V, Sabichi AL, Llansa N,
Lippman SM and Menter DG: Differential expression of
cyclooxygenase-2 and its regulation by tumor necrosis factor-alpha
in normal and malignant prostate cells. Cancer Res. 61:2720–2726.
2001.PubMed/NCBI
|
|
14
|
Tang K and Honn KV: 12(S)-HETE in cancer
metastasis. Adv Exp Med Biol. 447:181–191. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yin B, Yang Y, Zhao Z, et al: Arachidonate
12-lipoxygenase may serve as a potential marker and therapeutic
target for prostate cancer stem cells. Int J Oncol. 38:1041–1046.
2011.PubMed/NCBI
|
|
16
|
Ghosh J and Myers CE: Inhibition of
arachidonate 5-lipoxygenase triggers massive apoptosis in human
prostate cancer cells. Proc Natl Acad Sci USA. 95:13182–13187.
1998. View Article : Google Scholar
|
|
17
|
Tang S, Bhatia B, Maldonado CJ, et al:
Evidence that arachidonate 15-lipoxygenase 2 is a negative cell
cycle regulator in normal prostate epithelial cells. J Biol Chem.
277:16189–16201. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shureiqi I, Chen D, Lee JJ, et al:
15-LOX-1: a novel molecular target of nonsteroidal
anti-inflammatory drug-induced apoptosis in colorectal cancer
cells. J Natl Cancer Inst. 92:1136–1142. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vanaja DK, Grossmann ME, Celis E and Young
CY: Tumor prevention and antitumor immunity with heat shock protein
70 induced by 15-deoxy-delta12,14-prostaglandin J2 in transgenic
adenocarcinoma of mouse prostate cells. Cancer Res. 60:4714–4718.
2000.
|
|
20
|
Wang MT, Honn KV and Nie D:
Cyclooxygenases, prostanoids, and tumor progression. Cancer
Metastasis Rev. 26:525–534. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dassesse T, de Leval X, de Leval L,
Pirotte B, Castronovo V and Waltregny D: Activation of the
thromboxane A2 pathway in human prostate cancer correlates with
tumor Gleason score and pathologic stage. Eur Urol. 50:1021–1031.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nie D, Guo Y, Yang D, et al: Thromboxane
A2 receptors in prostate carcinoma: expression and its role in
regulating cell motility via small GTPase Rho. Cancer Res.
68:115–121. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nie D, Che M, Zacharek A, et al:
Differential expression of thromboxane synthase in prostate
carcinoma: role in tumor cell motility. Am J Pathol. 164:429–439.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang X, Colby JK, Yang P, Fischer SM,
Newman RA and Klein RD: The resistance to the tumor suppressive
effects of COX inhibitors and COX-2 gene disruption in TRAMP mice
is associated with the loss of COX expression in prostate tissue.
Carcinogenesis. 29:120–128. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shappell SB, Olson SJ, Hannah SE, et al:
Elevated expression of 12/15-lipoxygenase and cyclooxygenase-2 in a
transgenic mouse model of prostate carcinoma. Cancer Res.
63:2256–2267. 2003.PubMed/NCBI
|
|
26
|
Shappell SB, Manning S, Boeglin WE, et al:
Alterations in lipoxygenase and cyclooxygenase-2 catalytic activity
and mRNA expression in prostate carcinoma. Neoplasia. 3:287–303.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bhatia B, Maldonado CJ, Tang S, et al:
Subcellular localization and tumor-suppressive functions of
15-lipoxygenase 2 (15-LOX2) and its splice variants. J Biol Chem.
278:25091–25100. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Navone NM, Olive M, Ozen M, et al:
Establishment of two human prostate cancer cell lines derived from
a single bone metastasis. Clin Cancer Res. 3:2493–2500.
1997.PubMed/NCBI
|
|
29
|
Pfaffl MW: A new mathematical model for
relative quantification in real-time RT-PCR. Nucleic Acids Res.
29:e452001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shureiqi I, Zuo X, Broaddus R, et al: The
transcription factor GATA-6 is overexpressed in vivo and
contributes to silencing 15-LOX-1 in vitro in human colon cancer.
FASEB J. 21:743–753. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kempen EC, Yang P, Felix E, Madden T and
Newman RA: Simultaneous quantification of arachidonic acid
metabolites in cultured tumor cells using high-performance liquid
chromatography/electrospray ionization tandem mass spectrometry.
Anal Biochem. 297:183–190. 2001. View Article : Google Scholar
|
|
32
|
Yang P, Felix E, Madden T, Fischer SM and
Newman RA: Quantitative high-performance liquid
chromatography/electrospray ionization tandem mass spectrometric
analysis of 2- and 3-series prostaglandins in cultured tumor cells.
Anal Biochem. 308:168–177. 2002. View Article : Google Scholar
|
|
33
|
Yang P, Chan D, Felix E, et al:
Determination of endogenous tissue inflammation profiles by
LC/MS/MS: COX- and LOX-derived bioactive lipids. Prostaglandins
Leukot Essent Fatty Acids. 75:385–395. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang P, Collin P, Madden T, et al:
Inhibition of proliferation of PC3 cells by the branched-chain
fatty acid, 12-methyltetradecanoic acid, is associated with
inhibition of 5-lipoxygenase. Prostate. 55:281–291. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dahiya R, Yoon WH, Boyle B, Schoenberg S,
Yen TS and Narayan P: Biochemical, cytogenetic, and morphological
characteristics of human primary and metastatic prostate cancer
cell lines. Biochem Int. 27:567–577. 1992.PubMed/NCBI
|
|
36
|
Wang Y, Corr JG, Thaler HT, Tao Y, Fair WR
and Heston WD: Decreased growth of established human prostate LNCaP
tumors in nude mice fed a low-fat diet. J Natl Cancer Inst.
87:1456–1462. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Angelucci A, Garofalo S, Speca S, et al:
Arachidonic acid modulates the crosstalk between prostate carcinoma
and bone stromal cells. Endocr Relat Cancer. 15:91–100. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zha S, Gage WR, Sauvageot J, et al:
Cyclooxygenase-2 is up-regulated in proliferative inflammatory
atrophy of the prostate, but not in prostate carcinoma. Cancer Res.
61:8617–8623. 2001.PubMed/NCBI
|
|
39
|
Kirschenbaum A, Klausner AP, Lee R, et al:
Expression of cyclooxygenase-1 and cyclooxygenase-2 in the human
prostate. Urology. 56:671–676. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Larre S, Tran N, Fan C, et al: PGE2 and
LTB4 tissue levels in benign and cancerous prostates.
Prostaglandins Other Lipid Mediat. 87:14–19. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang H, Wang X, Zeng A and Yang K: Effects
of coherence on anisotropic electromagnetic Gaussian-Schell model
beams on propagation. Opt Lett. 32:2215–2217. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gao LJ, Wang XF, Yang HY, Gao XZ, Lu YC
and Cui ZJ: Effects of a lactic acid bacteria community SFC-2
treated on rice straw. Huan Jing Ke Xue. 28:1392–1396. 2007.(In
Chinese).
|
|
43
|
Timar J, Raso E, Dome B, et al:
Expression, subcellular localization and putative function of
platelet-type 12-lipoxygenase in human prostate cancer cell lines
of different metastatic potential. Int J Cancer. 87:37–43. 2000.
View Article : Google Scholar
|
|
44
|
Nie D, Hillman GG, Geddes T, et al:
Platelet-type 12-lipoxygenase in a human prostate carcinoma
stimulates angiogenesis and tumor growth. Cancer Res. 58:4047–4051.
1998.PubMed/NCBI
|
|
45
|
Kelavkar UP, Harya NS, Hutzley J, et al:
DNA methylation paradigm shift: 15-lipoxygenase-1 upregulation in
prostatic intraepithelial neoplasia and prostate cancer by atypical
promoter hypermethylation. Prostaglandins Other Lipid Mediat.
82:185–197. 2007. View Article : Google Scholar
|
|
46
|
Kelavkar UP, Parwani AV, Shappell SB and
Martin WD: Conditional expression of human 15-lipoxygenase-1 in
mouse prostate induces prostatic intraepithelial neoplasia: the
FLiMP mouse model. Neoplasia. 8:510–522. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shappell SB, Boeglin WE, Olson SJ, Kasper
S and Brash AR: 15-lipoxygenase-2 (15-LOX-2) is expressed in benign
prostatic epithelium and reduced in prostate adenocarcinoma. Am J
Pathol. 155:235–245. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gonzalez AL, Roberts RL, Massion PP, Olson
SJ, Shyr Y and Shappell SB: 15-Lipoxygenase-2 expression in benign
and neoplastic lung: an immunohistochemical study and correlation
with tumor grade and proliferation. Hum Pathol. 35:840–849. 2004.
View Article : Google Scholar : PubMed/NCBI
|