Introduction
Cell migration and invasion are two critical
cellular processes that are often deregulated during tumorigenesis.
Cancer growth is accompanied by progressive infiltration, invasion
and destruction of the surrounding tissue. The metastasis and
invasiveness of tumor cells are the most reliable features that
differentiate malignant tumors from benign ones (1). Despite our advanced understanding of
primary cancer development and progression, metastasis and the
systemic spread of the cancer to secondary sites remains the
leading cause of cancer-associated death (2). Therefore, the discovery of a new
anti-metastatic strategy is critical in order to increase the
survival and to prevent the mortality of patients with cancer.
In epithelial cells, several specialized and
distinct intercellular structures, including the gap junction,
tight junction (TJ), adherens junction and desmosome, are
responsible for the establishment of contact between neighboring
cells. Among them, TJs are the most apical component of cell/cell
complexes and play a critical role in regulating the balance
between differentiation, proliferation and cell death (3,4).
However, during metastasis, the expression pattern of TJ proteins
is frequently disrupted in epithelial tumors. Claudins, which are
major integral membrane proteins that form the backbone of TJs, can
form homodimers or heterodimers to produce paired strands between
adjacent cells and act as a barrier to the paracellular flux of
water and the solution and transmigration of other cells, thereby
determining the characteristic permeability properties of different
epithelial tissues (5–7). According to the serial analysis of
gene expression, the overexpression of claudins is frequently found
in various cancer cells. For instance, claudin-3 and -4 have been
reported to be constantly elevated in ovarian, breast, prostate and
pancreatic cancers, as well as bladder cancer (8–12).
Recent reports found that certain claudins, such as claudin-4 and
-7 are more elevated in high-grade urothelial cancer patients as
compared to low-grade patients (13,14).
Therefore, the examination of the modulation of claudin family
proteins is adjuvant to enhancing the accuracy of the pathological
diagnosis and the addition of further information for clinical
treatment (15,16).
Matrix metalloproteinases (MMPs), are a family of
zinc ion-dependent endopeptidases, that consists of more than 26
endopeptidases, which are known to act on a broad spectrum of cell
surface molecules (17–19). The three prominent roles of MMPs in
pathological conditions may be grouped into the following types:
tissue destruction, fibrosis and the weakening of the matrix. In
the tumor cell invasion process, degradation of the extracellular
matrix (ECM) is one of the pivotal steps. Therefore, the
upregulation of MMPs has been found in the majority of malignant
tumors, and has also been connected with cancer aggressiveness,
stage and prognosis (20,21). Among all the members of the MMP
family, MMP-2 (gelatinase-A) and -9 (gelatinase-B) have been known
to be involved in the invasive metastatic possess of tumor cells,
including bladder cancer cells (22–24).
Since MMPs have many physiological functions in metastasis, the
inhibition of the activity of MMPs is inherent for the activated
MMPs, thus the tissue inhibitor of metalloproteinase (TIMP)
prevents the uncontrolled action of these proteinases (21,24,25).
Bufalin is the major digoxin-like immunoreactive
component, obtained from the skin and parotid venom gland of the
toad, Bufo bufo gargarizans(26). Although bufalin has long been used
as a treatment for heart failure in Oriental medicine in Asian
countries, studies have shown that bufalin exhibits anticancer
effects. Bufalin induces cell cycle arrest, differentiation and
apoptosis in many human cancer cells such as leukemia, gastric,
colon, breast, endometrial and ovarian cancer and osteosarcoma
cells (27–33). Recently, Chueh et
al(34) demonstrated that
bufalin inhibited migration and invasion in human osteosarcoma
cells by down-regulating the levels of MMP-2, extracellular
signal-regulated kinase (ERK) and c-Jun N-terminal kinase (JNK)
signaling pathways. However, the anti-metastatic effects of bufalin
on cancer cells have not yet been thoroughly reported and knowledge
of the molecular mechanisms involved is rudimentary and remains to
be delineated. Therefore, the purpose of this study was to
elucidate the anti-metastatic potential of bufalin using the T24
human bladder carcinoma cell line and to investigate the underlying
intracellular signal transduction pathways involved in the
inhibition of metastasis. The results of this study demonstrate
that bufalin inhibits two measures of metastatic potential, cell
motility and invasiveness, through the modulation of the levels of
TJ-associated factors and the activities of MMPs.
Materials and methods
Reagents and antibodies
RPMI-1640 medium was purchased from Invitrogen Corp.
(Carlsbad, CA, USA). Fetal bovine serum (FBS) was acquired from
Gibco-BRL (Gaithersburg, MD, USA). Bufalin was purchased from
Sigma-Aldrich Chemical Corp. (St. Louis, MO, USA), dissolved in
dimethyl sulfoxide (DMSO, vehicle) and stored in aliquots at 4°C.
The primary antibodies against MMP-2, MMP-9, TIMP-1 and TIMP-2 were
purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA) and
antibodies against claudin-2, -3 and -4 were obtained from
Invitrogen. The antibody against actin was obtained from
Sigma-Aldrich. Peroxidase-labeled donkey anti-rabbit and sheep
anti-mouse immunoglobulin were acquired from Santa Cruz
Biotechnology. All other chemicals not specifically mentioned were
purchased from Sigma-Aldrich.
Cell culture and trypan blue
counting
T24 human bladder carcinoma cells were obtained from
American Type Culture Collection (Rockville, MD, USA) and cultured
in RPMI-1640 medium supplemented with 10% heat-inactivated FBS, 1%
penicillin-streptomycin (Gibco-BRL) at 37°C and 5% CO2.
For the cell viability study, T24 cells were grown to 70%
confluence, treated with bufalin and then cell viability was
assessed by a trypan blue exclusion assay.
Colony forming assay
T24 cells were seeded into 100-mm dishes at a
density of 5, 10 or 20×102 cells/dish and allowed to be
stabilized for 24 h at 37°C in a culture medium. Cells were then
treated with 100 nM of bufalin. After 15 days, colonies were fixed
with 3.7% paraformaldehyde for 20 min and stained with hematoxylin
solution for 10 min at room temperature. The dishes were washed
with PBS and then counted manually and photographed.
Cell motility and wound healing
assay
For the persistence migratory directionality assay,
T24 cells were grown to confluence on 35 mm cell culture dishes
that were coated with 20 μg/ml of rat tail collagen
(Becton-Dickinson Biosciences, Bedford, MA, USA). Confluent
monolayer cells were scratched using a 200 μl micropipette
tip. After wounding, the cultures were washed twice with FBS-free
RPMI-1640 medium to remove cell debris. The cells were supplemented
with a medium containing 0.5% FBS and treated with 50 or 100 nM of
bufalin. At the indicated time, the wound closure weight of cells
was inspected and photographed under the microscope at ×40
magnification, respectively. The assays were repeated twice and
each sample was observed in triplicate (35).
In vitro invasion assay
To observe the ability of T24 cells to penetrate the
ECM in the presence or absence of bufalin, Matrigel invasion assays
were performed. The chambers consisted of 6.5-mm inserts with
8.0-μm pore membranes (Corning Costar Corporation, Corning,
NY, USA). Each membrane was coated with 50 μl of Matrigel
(Becton-Dickinson Biosciences) diluted in an ice-cold serum-free
medium using a cooled pipette and incubated for 30 min at 37°C.
After the plates were ready to use, pre-warmed RPMI complete medium
was added to the inside of the top chamber and rehydrated for 1 h
in a 37°C incubator. Subsequently, T24 cells were seeded in
serum-free medium with or without bufalin in the upper chamber of
the transwell, and medium containing 20% FBS was added to the
bottom chambers. After overnight incubation, the membranes were
fixed with methanol for 2 min, stained with hematoxylin for 4 min
and washed with distilled water. The membranes were removed from
the insert with a small scalpel blade, mounted on a microscope
slide and coverslipped. Invasive cells were observed under a
microscope at ×200 magnification and quantified (three fields for
each membrane).
Measurement of transepithelial electrical
(TER) resistance
TER was measured using an EVOM Epithelial Tissue
Voltohmmeter (World Precision Instruments, Sarasota, FL, USA),
equipped with a pair of STX-2 chopstick electrodes. Briefly, the
T24 cells were seeded in the 8.0-μm pore size top chamber of
the Transwell (Corning Costar) with serum-free medium and an assay
medium containing 20% FBS was added to the bottom chamber. The
cells were treated with bufalin at a concentration of 50 or 100 nM
and incubated for 24 h. Two electrodes were placed at the top and
bottom chambers and then resistance was estimated with the
voltohmmeter.
Protein extraction and western blot
analysis
The cells were treated with bufalin at the indicated
time and harvested. Total cells were then gently lysed with lysis
buffer (20 mM sucrose, 1 mM EDTA, 20 μM Tris-Cl, pH 7.2, 1
mM DTT, 10 mM KCl, 1.5 mM MgCl2 and 5 μg/ml
aprotinin) for 30 min. The supernatants were collected and the
protein concentrations were quantified using a Bio-Rad protein
assay kit (Bio-Rad Laboratories, Hercules, CA, USA). For western
blot analysis, equal amounts of protein were subjected to
electrophoresis on SDS-polyacrylamide gels and then
electrotransferred onto nitrocellulose membranes (Schleicher &
Schuell, Keene, NH, USA). Blots were probed with the desired
antibodies, incubated with the diluted enzyme-linked secondary
antibodies and visualized by enhanced chemiluminescence (ECL),
according to the procedure recommended (Amersham Corp., Arlington
Heights, IL, USA).
RNA extraction and reverse
transcription-polymerase chain reaction (PCR)
Total-RNAs were isolated using a RNeasy mini kit
(Qiagen, La Jolla, CA, USA) and primed with random hexamers to
synthesize complementary DNA using AMV reverse transcpriptase
(Amersham) following the manufacturer’s instructions. PCR was
performed in a Mastercycler (Eppendorf, Hamburg, Germany) with the
primers indicated in Table I. The
conditions for PCR reactions were 1X (94°C for 3 min), 35X (94°C
for 45 sec; 58°C for 45 sec; and 72°C for 1 min) and 1X (72°C for
10 min). The amplification products obtained by PCR were
electrophoretically separated on a 1% agarose gel and visualized by
ethidium bromide (EtBr) staining.
 | Table I.Primer sequences used for RT-PCR. |
Table I.
Primer sequences used for RT-PCR.
| Gene name | | Sequence |
|---|
| GAPDH | Sense | 5′-CGG AGT CAA CGG
ATT TGG TCG TAT-3′ |
| Antisense | 5′-AGC CTT CTC CAT
GGT GGT GAA GAC-3′ |
| MMP-2 | Sense | 5′-CTT CTT CAA GGA
CCG GTT CAT-3′ |
| Antisense | 5′-GCT GGC TGA GTA
GAT CCA GTA-3′ |
| MMP-9 | Sense | 5′-TGG GCT ACG TGA
CCT ATG ACC AT-3′ |
| Antisense | 5′-GCC CAG CCC ACC
TCC ACT CCT C-3′ |
| TIMP-1 | Sense | 5′-TGG GGA CAC CAG
AAG TCA AC-3′ |
| Antisense | 5′-TTT TCA GAG CCT
TGG AGG AG-3′ |
| TIMP-2 | Sense | 5′-GTC AGT GAG AAG
GAA GTG GAC TCT-3′ |
| Antisense | 5′-ATG TTC TTC TCT
GTG ACC CAG TC-3′ |
| Snail | Sense | 5′-TAT GCT GCC TTC
CCA GGC TTG-3′ |
| Antisense | 5′-ATG TGC ATC TTG
AGG GCA CCC-3′ |
| E-cadherin | Sense | 5′-GAA CAG CAC GTA
CAC AGC CCT-3′ |
| Antisense | 5′-GCA GAA GTG TCC
CTG TTC CAG-3′ |
| Claudin-1 | Sense | 5′-TCA GCA CTG CCC
TGC CCC AGT-3′ |
| Antisense | 5′-TGG TGT TGG GTA
AGA GGT TGT-3′ |
| Claudin-2 | Sense | 5′-ACA CAC AGC ACA
GGC ATC AC-3′ |
| Antisense | 5′-TCT CCA ATC TCA
AAT TTC ATG C-3′ |
| Claudin-3 | Sense | 5′-AAG GCC AAG ATC
ACC ATC GTG-3′ |
| Antisense | 5′-AGA CGT AGT CCT
TGC GGT CGT-3′ |
| Claudin-4 | Sense | 5′-TGG ATG AAC TGC
GTG GTG CAG-3′ |
| Antisense | 5′-GAG GCG GCC CAG
CCG ACG TA-3′ |
Gelatin zymographic analysis of secreted
MMPs
The enzymatic activities of MMP-2 and -9 were
measured by gelatin zymography. Briefly, the bufalin-treated cell
culture supernatants were collected using centrifugation. The
cell-free supernatant was mixed with 10X bromophenol blue (Junsei
Chemical Co. Ltd., Tokyo, Japan) and zymography was conducted by
precast gel (10% polyacrylamide and 0.25% gelatin). Following
electrophoresis, the gel was shaken with 2.5% Triton X-100 at room
temperature for 1 h and washed with distilled water two to three
times. Next, the gel was incubated with a developing buffer
containing 1 M Tris-HCl (pH 7.5) 50 ml, 1 M CaCl2 5 ml,
10% NaN3 2 ml, 20 mM ZnCl2 50 μl,
H2O at 37°C for 24 h. The following day, the gel was
stained with 0.5% (w/v) Coomassie Brilliant Blue (Bio-Rad) for 30
min and then destained in methanol:acetic acid:water (1:1:8)
(36).
Statistical analysis
All data are presented as the means ± SD. The
significant differences among the groups were analyzed using a
one-way analysis of variance followed by an ANOVA test. A value of
p<0.05 was considered as an indication of statistical
significance.
Results
Effects of bufalin on cell proliferation
and colony formation in T24 cells
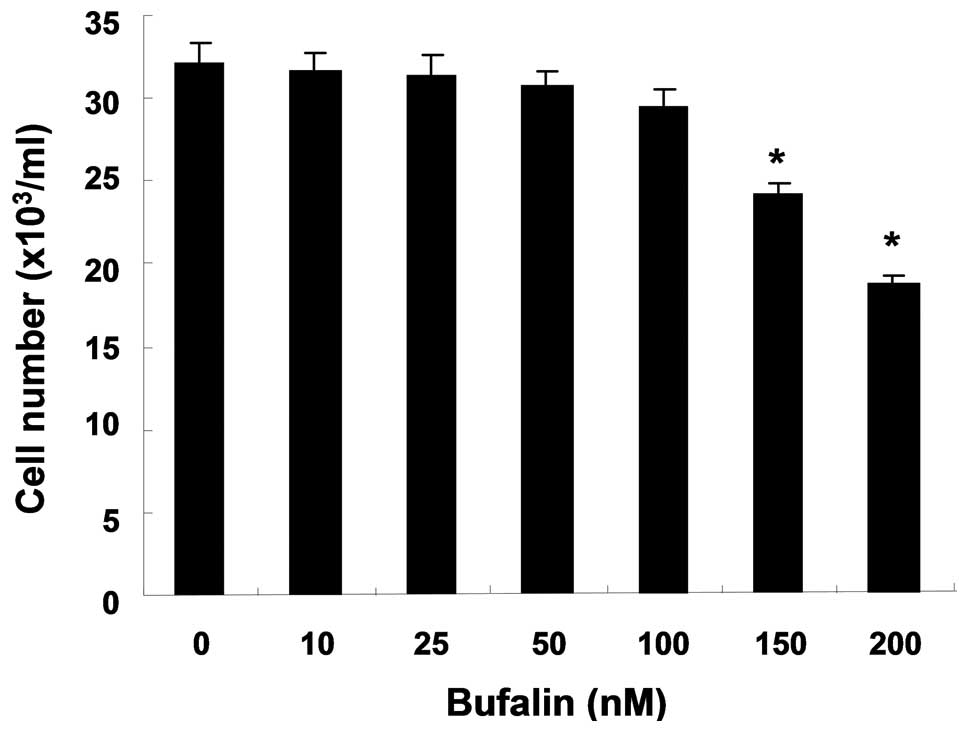
To investigate the effects of bufalin on cell
proliferation, T24 cells were treated with diverse concentrations
for 24 h and subjected to trypan blue exclusion. When compared with
the untreated control cells, bufalin showed a dose-dependent
inhibitory effect on the proliferation of T24 cells (Fig. 1). However, bufalin in the range of
10–100 nM did not have a significant cytotoxic effect on T24 cells;
therefore, a concentration of bufalin within this lower range was
used in the remaining experiments.
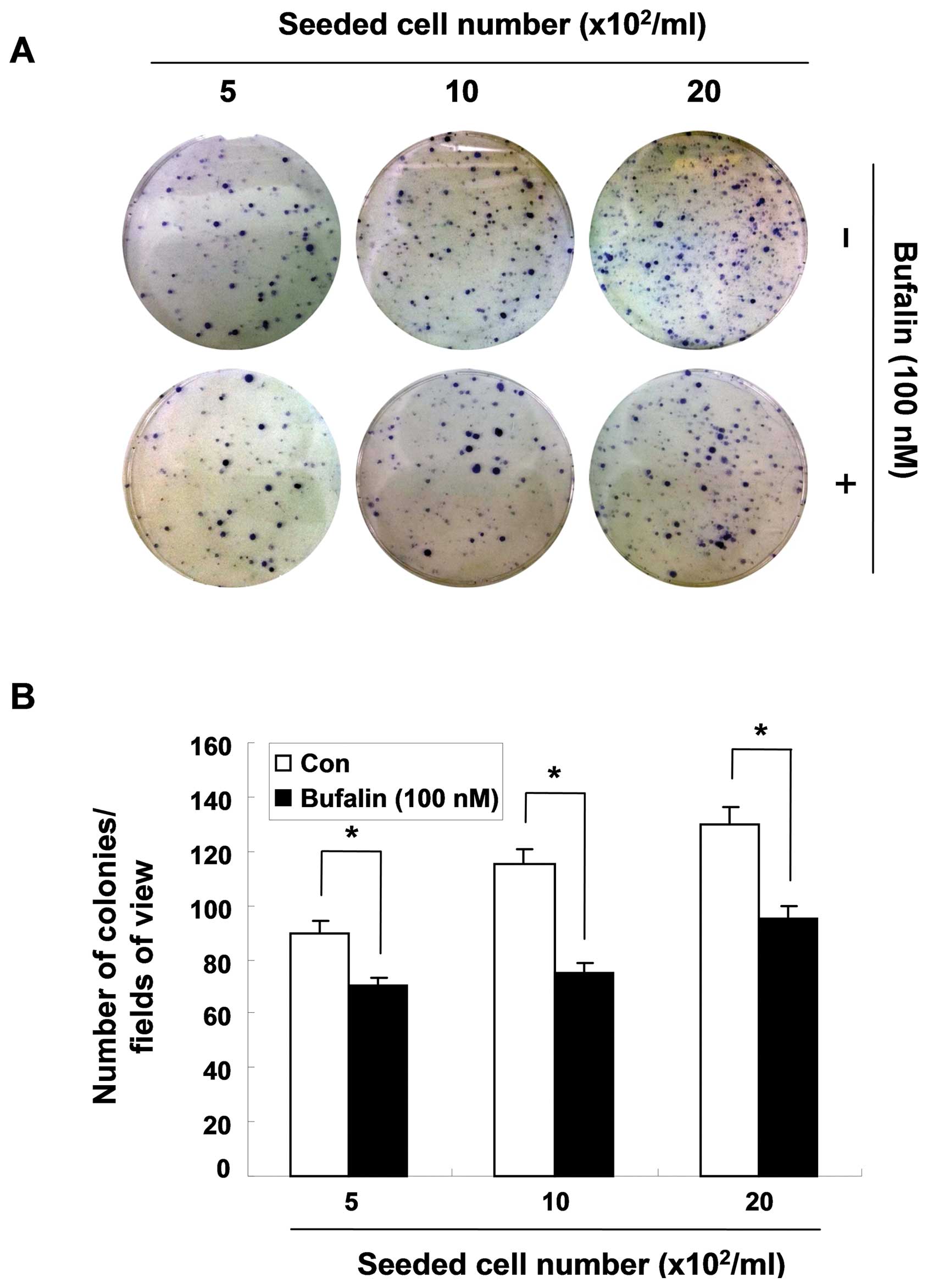
To confirm the cytostatic effect of bufalin,
clonogenic assay was also performed. Based on trypan blue exclusion
results, 100 nM concentration of bufalin were chosen for this
study. Independent of the seeded number of cells, the dishes
treated with bufalin exhibited less colony formation than the
untreated ones, suggesting that bufalin has exerts
anti-proliferative effects at a non-cytotoxic dose (Fig. 2).
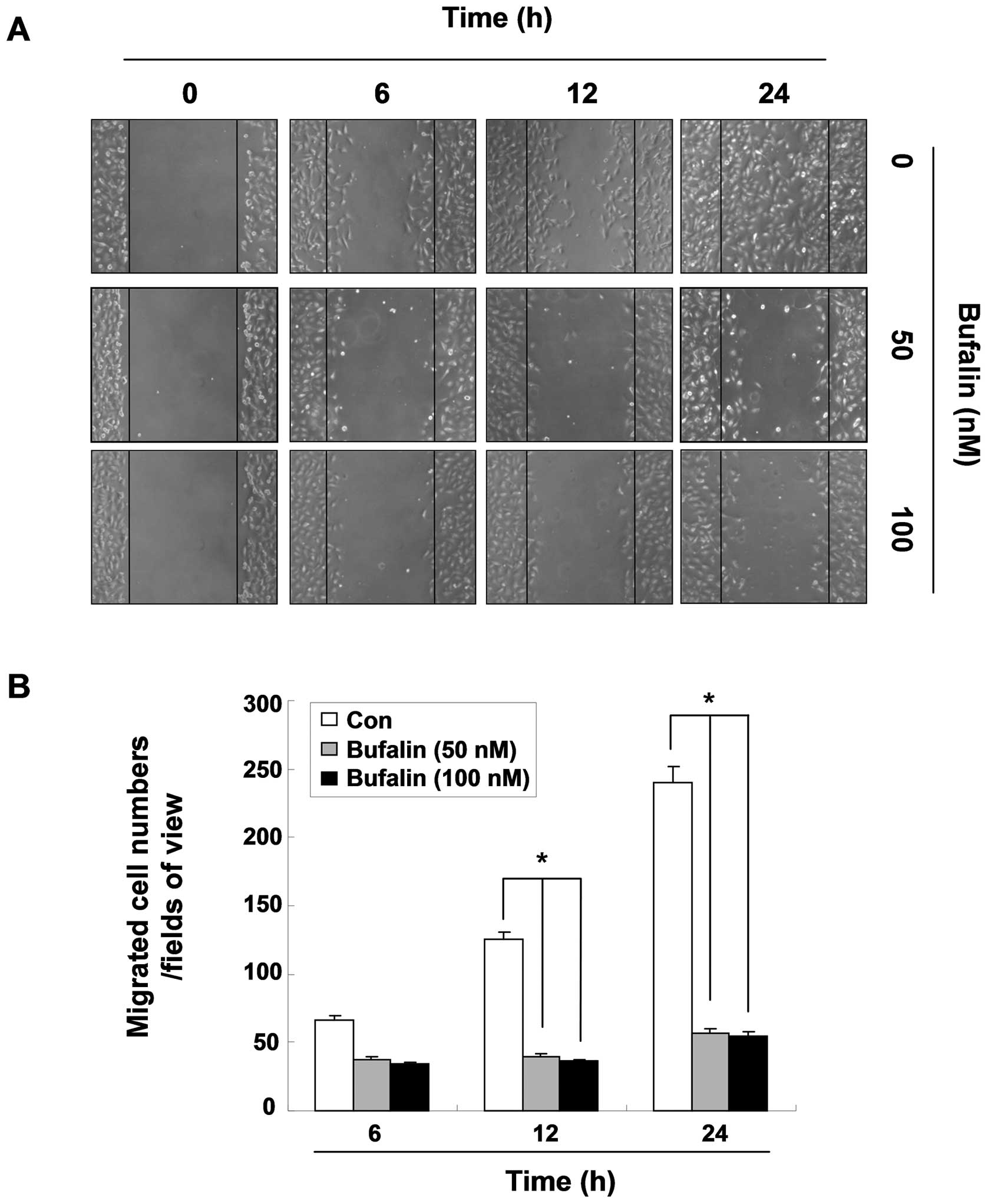
Delay of migration by bufalin in T24
cells
To measure the inhibitory effect of bufalin on the
motility of T24 cells, a wound healing assay, a widely used
qualitative method of the study of cell migration, was carried out
in the condition medium containing 0.5% FBS. After 24 h, the width
of the wound was almost completely closed in the untreated dishes;
however, a distinct gap still remained in the bufalin-treated
dishes (Fig. 3). Quantitative data
showing the migrating cells demonstrated that treatment with
bufalin impeded the migration of cells compared to the control
cells in a time-dependent manner. However, there was no significant
difference between the two doses, 50 and 100 nM of bufalin. These
results suggest that bufalin possesses the ability to reduce cell
motility and migration in T24 cells.
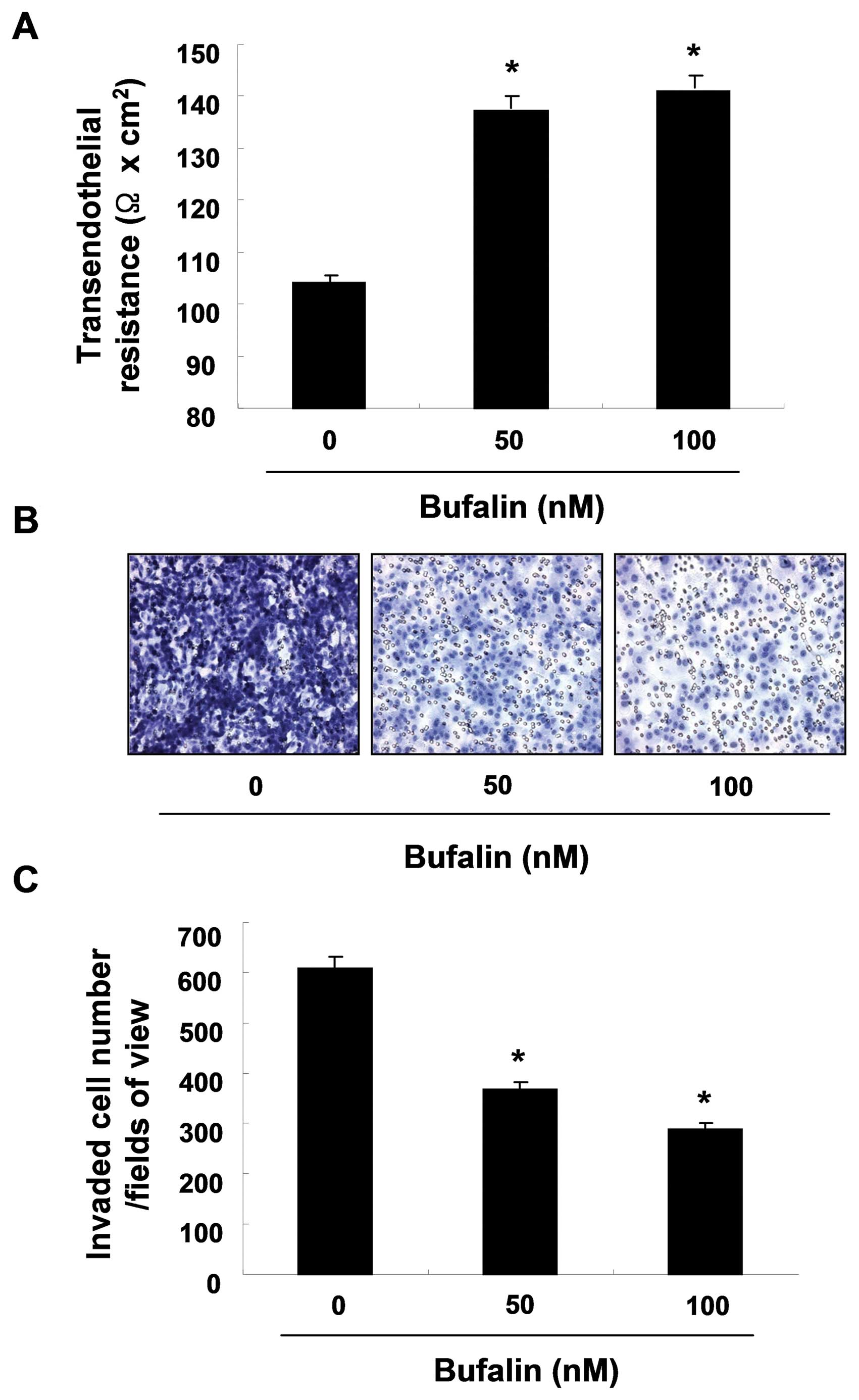
Bufalin increases TER values and
decreases cell invasion in T24 cells
To determine the interaction between the tightening
of TJs and the anti-invasive activity of T24 cells stimulated with
bufalin, the investigation of TER (a measure of TJ formation)
values was conducted. After bufalin exposure for 24 h, TER values
were considerably increased in a dose-dependent manner suggesting
that bufalin increased the tightening of TJs (approximately
1.3-fold and 1.4-fold by 50 and 100 nM of bufalin, respectively,
Fig. 4A). We subsequently
investigated whether bufalin can reduce cell invasion using a
Boyden chamber invasion assay. As shown in Fig. 4B and C, incubating cells with 50
and 100 nM of bufalin reduced cell invasion through the Matrigel
chamber to 39.5 and 53.6% of the control levels at 24 h,
respectively, suggesting that the upregulation of TER contributes
to the inhibition of cell invasion in T24 cells.
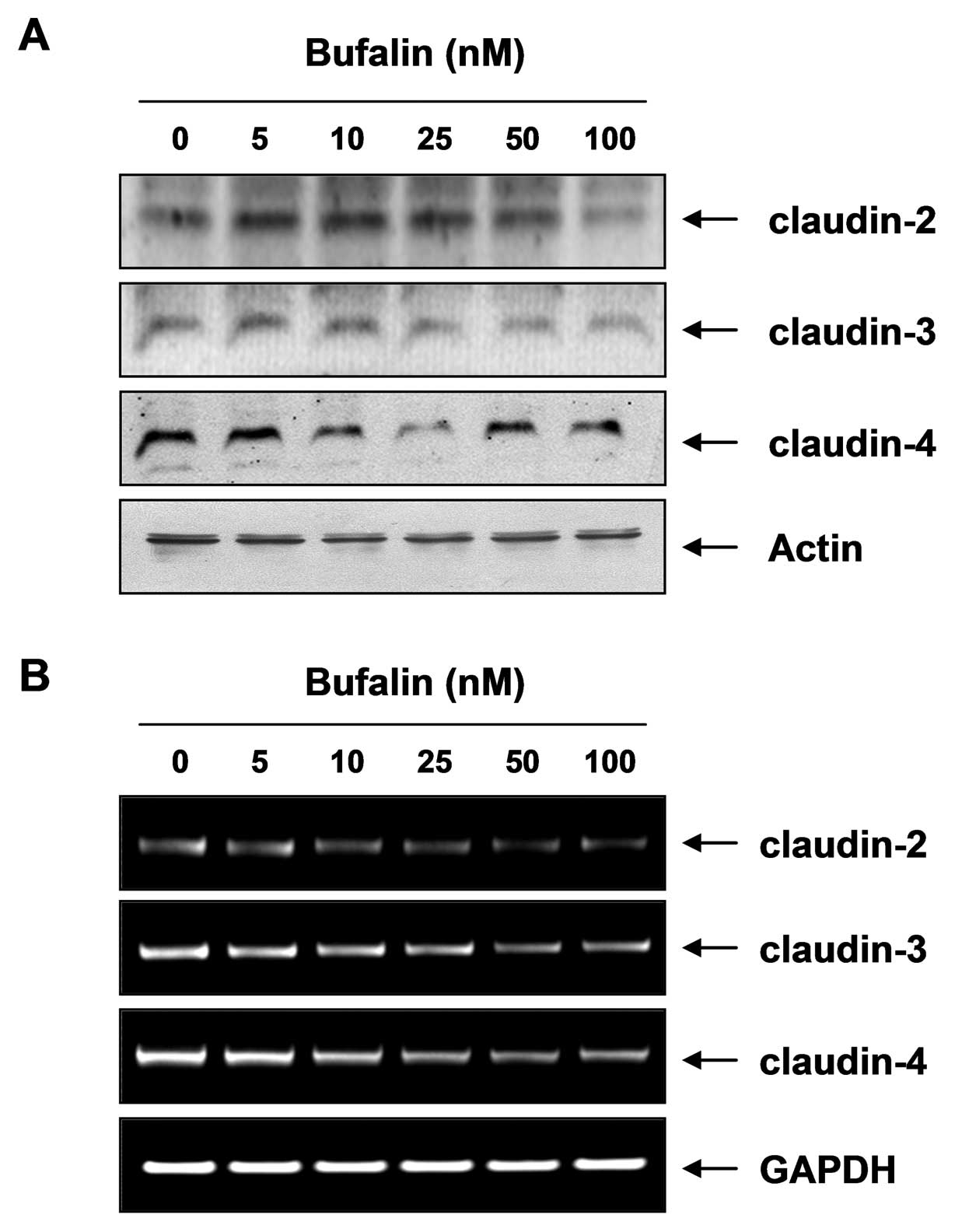
Bufalin modulates the expression of
claudin family members
To elucidate the mechanism by which bufalin enhances
TJ activity, we examined the levels of TJ components, claudins,
through western blot analysis and RT-PCR analysis. As illustrated
in Fig. 5, bufalin
dose-dependently downregulated the levels of claudin proteins
including claudin-2, -3 and -4 and considerably decreased their
mRNA levels. These results indicate that this modulation of claudin
family members by bufalin may be associated with the TJ tightening
in T24 cells.
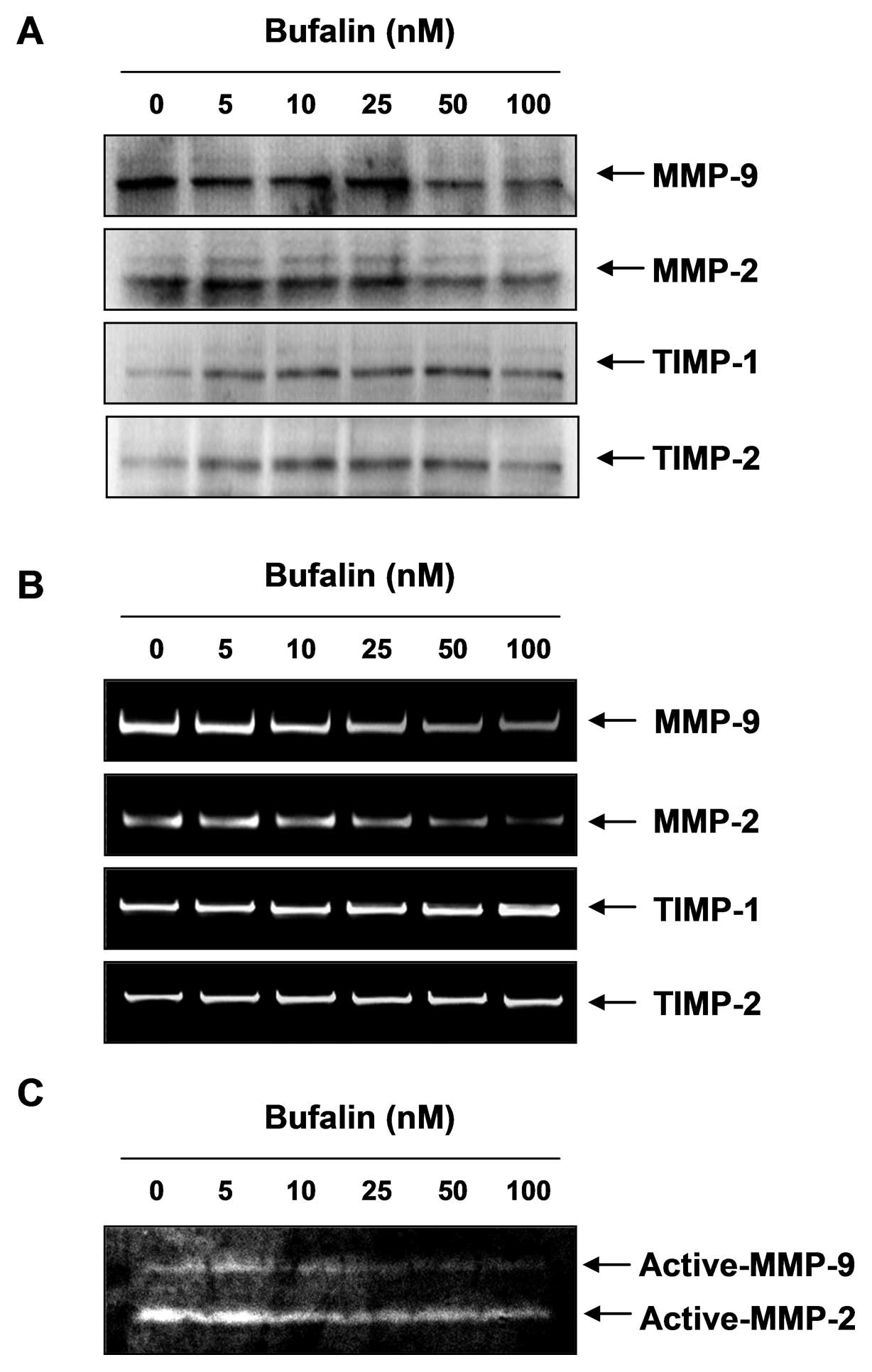
Bufalin inhibits enzyme activities and
expression of MMPs in T24 cells
As an increase in MMP enzyme activity is important
for the degradation of the ECM, which is critical to metastasis, we
investigated the effect of bufalin on MMP-2 and -9 activity using
western blot analysis, RT-PCR and gelatin zymography. As shown in
Fig. 6, bufalin treatment led to a
downregulation of the MMP protein and mRNA levels in a
dose-dependent manner, which was concomitant with decreased MMP-2
and -9 enzyme activities. However, bufalin treatment increased
TIMP-1 and -2 mRNA and protein levels in a concentration-dependent
manner. These results indicate that the anti-invasive effect of
bufalin is associated with increased TIMP levels, as well as the
inhibition of MMP expression and activity in T24 cells.
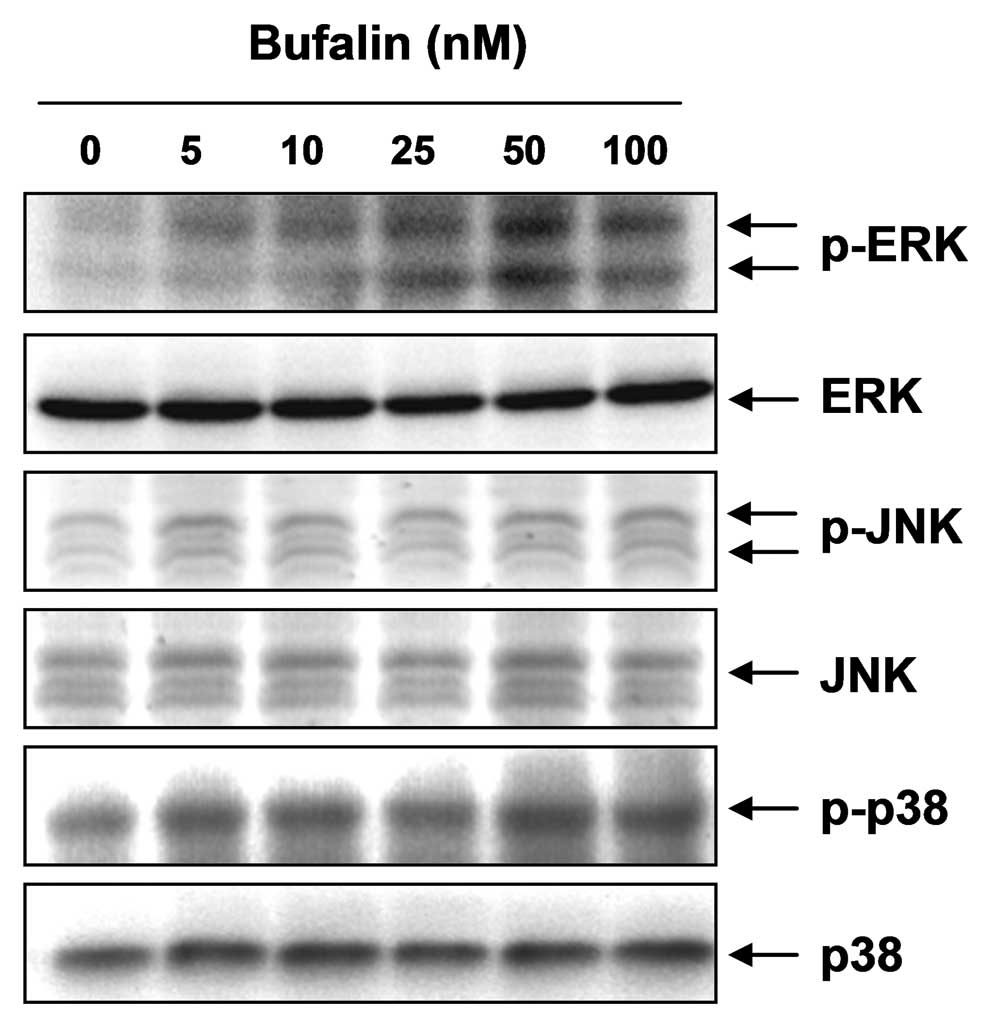
Bufalin modulates the activation of the
mitogen-activated protein kinase (MAPK) pathway in T24 cells
Several lines of evidence have implicated the MAPK
pathway in the regulation of MMP and TJ activity (37–39),
which have been implicated in a number of cellular functions
including cell survival, adhesion and metastasis. Therefore, to
investigate whether the anti-invasive effect of bufalin is mediated
through the modulation of the MAPK pathway, we examined the
activation of the three MAPKs by analyzing their phosphorylated
forms in western blots probed with specific anti-phosphokinase
antibodies. The data demonstrated that 50 nM bufalin treatment
markedly increased the phosphorylation of ERK, but not that of JNK
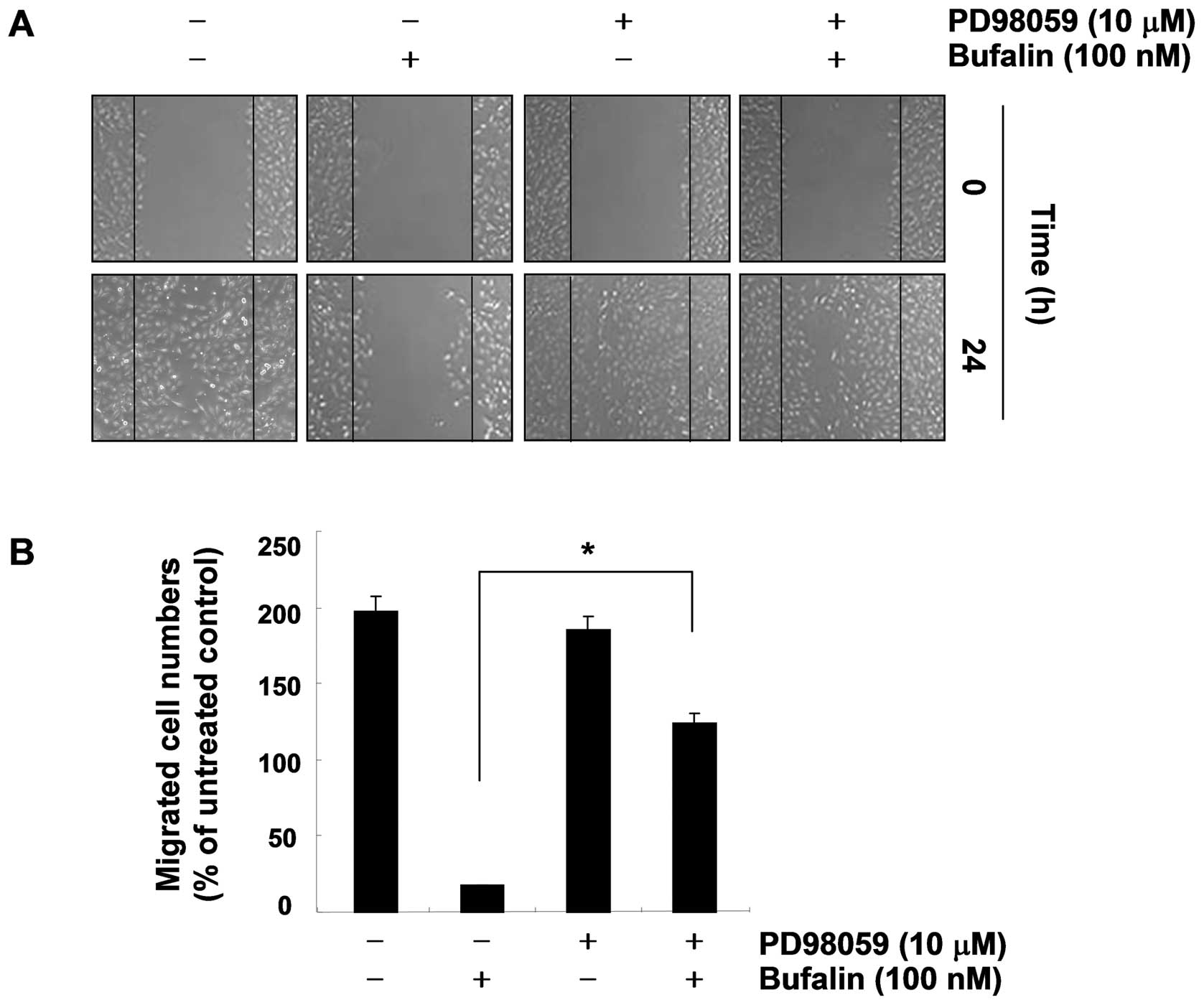
and p38 MAPK (Fig. 7). We then
evaluated the possible role of ERK signaling in bufalin-induced
anti-invasive activity. As shown in Fig. 8, pre-treatment with PD98059 (an ERK
inhibitor) markedly reversed the bufalin-mediated reduction of T24
cell migration, which indicates that bufalin-induced
anti-invasiveness is associated with the ERK pathway.
Discussion
Bufalin is a major active component of Sum su (Chan
su in Chinese), which is obtained from the skin and parotid venom
gland of the toad, Bufo bufo gargarizans(26). Sum su has a wide range of
biological applications as a cardiotonic, local anesthetic,
antimicrobial, blood pressure stimulator, and has been used in
respiratory and antitumor traditional medicines in Korea, China and
other Asian countries (26).
Moreover, a number of experimental studies on bufalin have been
performed worldwide. Bufalin has revealed distinct antitumor
activity, such as the inhibition of cell proliferation, induction
of cell differentiation and apoptosis, the disruption of the cell
cycle, inhibition of angiogenesis, the ability to overcome
anticancer drug resistance and the modulation of the immune
response (27–33,40–42).
Therefore, it is expected that bufalin may be a novel target to
reduce cancer cell metastasis. However, the anti-metastatic
activity of bufalin has not yet been extensively investigated; to
our knowledge, only one study has been published so far (34). In the present study, we found that
bufalin significantly inhibited the migratory and invasive activity
in T24 bladder carcinoma cells by enhancing TJ activity,
downregulating MMP activity and modulating TJ-related proteins
in vitro.
TJs are completely necessary to create a tight seal
of cellular sheets, which is comprised of three major integral
membrane proteins, claudin, occludin and junctional adhesion
molecules. According to previous studies, the modulation of
claudins plays an important role during oncogenic transformation
exhibiting distinct tissue- and development-specific distribution
patterns (8,43). Thus, the regulation of claudin
proteins may be the new target for cancer treatment. Our data
showed that bufalin treatment downregulated almost all the claudin
levels. In this regard, the downregulation of claudins is connected
with the tightening of TJs, the inhibition of cell migration and
the invasion in T24 cells. In addition, bufalin induced a
significant inhibition of protein and mRNA levels and the enzyme
activities of MMP-2 and -9. Simultaneously, the mRNA and protein
levels of TIMP-1 and -2 were elevated in a concentration-dependent
manner. MMP-2 and -9, two types of gelatinases, are key agonists in
tumor invasion and angiogenesis; thus, tumor metastasis may be
inhibited by blocking MMP synthesis and activation (11,44).
Therefore, the data indicated that the activity of MMPs is limited
to physiologically binding to one of four endogenous TIMPs;
therefore, the balance of secreted MMPs and TIMPs is maintained in
the connective tissue homeostasis of normal tissue. However, an
imbalance of MMPs and TIMPs has usually been exhibited to lead to
an excess of degradative activity (45). As shown in Fig. 6, the anti-invasive activity of
bufalin in T24 cells was related with the decrease in the MMP/TIMP
ratio as a key factor in the regulation of the anti-metastatic
process, which subsequently blocks the degradation of the ECM and
leads to the inhibition of cell invasion.
Although the MAPK pathway plays a critical role in
regulating cell death and survival in many physiological and
pathological settings, numerous reports have demonstrated that the
function and expression of TJs and MMPs are critically mediated by
the MAPK pathway, suggesting that these pathways also play an
important role in tumor metastasis (37–39).
Thus, we confirmed the involvement of the MAPK pathway in
bufalin-induced anti-invasive activity. In the present study, we
observed that bufalin induced the activation of ERK, but not that
of JNK and p38 MAPK, and that the inhibition of ERK suppressed the
bufalin-induced inhibition of cell migration. These results suggest
that the bufalin inhibition of cell invasion of T24 cells may
partly occur through the activation of the ERK pathway.
In view of the results thus far achieved, bufalin
suppresses the attachment to the ECM, cell migration and invasion
in T24 cells. Although further controlled trials are warranted,
these results provide evidence that bufalin inhibits the metastasis
of human bladder cancer in vitro. Overall, the present study
suggests that bufalin is a promising therapeutic agent for cancer
therapy with low toxicity and few side-effects.
Acknowledgements
This study was supported by the
National Research Foundation of Korea (NRF) grant funded by the
Korea government (2012-0000476).
References
|
1.
|
Gunawardane RN, Sgroi DC, Wrobel CN, Koh
E, Daley GQ and Brugge JS: Novel role for PDEF in epithelial cell
migration and invasion. Cancer Res. 65:11572–11580. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Timpson P, Serrels A, Canel M, Frame MC,
Brunton VG and Anderson KI: Quantitative real-time imaging of
molecular dynamics during cancer cell invasion and metastasis in
vivo. Cell Adh Migr. 3:351–354. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Choi YH, Choi WY, Hong SH, Kim SO, Kim GY,
Lee WH and Yoo YH: Anti-invasive activity of sanguinarine through
modulation of tight junctions and matrix metalloproteinase
activities in MDA-MB-231 human breast carcinoma cells. Chem Biol
Interact. 179:185–191. 2009. View Article : Google Scholar
|
|
4.
|
Boireau S, Buchert M, Samuel MS, Pannequin
J, Ryan JL, Choquet A, Chapuis H, Rebillard X, Avancès C, Ernst M,
Joubert D, Mottet N and Hollande F: DNA-methylation-dependent
alterations of claudin-4 expression in human bladder carcinoma.
Carcinogenesis. 28:246–258. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Angelow S and Yu AS: Claudins and
paracellular transport: an update. Curr Opin Nephrol Hypertens.
16:459–464. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Morin PJ: Claudin proteins in human
cancer: promising new targets for diagnosis and therapy. Cancer
Res. 65:9603–9606. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Utech M, Brüwer M and Nusrat A: Tight
junctions and cell-cell interactions. Methods Mol Biol.
341:185–195. 2006.PubMed/NCBI
|
|
8.
|
Agarwal R, D’Souza T and Morin PJ:
Claudin-3 and claudin-4 expression in ovarian epithelial cells
enhances invasion and is associated with increased matrix
metalloproteinase-2 activity. Cancer Res. 65:7378–7385. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Hewitt KJ, Agarwal R and Morin PJ: The
claudin gene family: expression in normal and neoplastic tissues.
BMC Cancer. 6:1862006. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Morin PJ: Claudin proteins in ovarian
cancer. Dis Markers. 23:453–457. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Neesse A, Griesmann H, Gress TM and Michl
P: Claudin-4 as therapeutic target in cancer. Arch Biochem Biophys.
524:64–70. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Nakanishi K, Ogata S, Hiroi S, Tominaga S,
Aida S and Kawai T: Expression of occludin and claudins 1, 3, 4 and
7 in urothelial carcinoma of the upper urinary tract. Am J Clin
Pathol. 130:43–49. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Törzsök P, Riesz P, Kenessey I, Székely E,
Somorácz A, Nyirády P, Romics I, Schaff Z, Lotz G and Kiss A:
Claudins and ki-67: potential markers to differentiate low- and
high-grade transitional cell carcinomas of the urinary bladder. J
Histochem Cytochem. 59:1022–1030. 2011.PubMed/NCBI
|
|
15.
|
Ouban A and Ahmed AA: Claudins in human
cancer: a review. Histol Histopathol. 25:83–90. 2010.
|
|
16.
|
Takahashi A, Kondoh M, Suzuki H and Yagi
K: Claudin as a target for drug development. Curr Med Chem.
18:1861–1865. 2011. View Article : Google Scholar
|
|
17.
|
Martin MD and Matrisian LM: The other side
of MMPs: protective roles in tumor progression. Cancer Metastasis
Rev. 26:717–724. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: regulators of the tumor microenvironment. Cell.
141:52–67. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Pytliak M, Vargová V and Mechírová V:
Matrix metalloproteinases and their role in oncogenesis: a review.
Onkologie. 35:49–53. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Zhong WD, Han ZD, He HC, Bi XC, Dai QS,
Zhu G, Ye YK, Liang YX, Qin WJ, Zhang Z, Zeng GH and Chen ZN:
CD147, MMP-1, MMP-2 and MMP-9 protein expression as significant
prognostic factors in human prostate cancer. Oncology. 75:230–236.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Amălinei C, Căruntu ID, Giuşcă SE and
Bălan RA: Matrix metalloproteinases involvement in pathologic
conditions. Rom J Morphol Embryol. 51:215–228. 2010.
|
|
22.
|
Zhang L, Shi J, Feng J, Klocker H, Lee C
and Zhang J: Type IV collagenase (matrix metalloproteinase-2 and
-9) in prostate cancer. Prostate Cancer Prostatic Dis. 7:327–332.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Black PC and Dinney CP: Bladder cancer
angiogenesis and metastasis-translation from murine model to
clinical trial. Cancer Metastasis Rev. 26:623–634. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Szarvas T, vom Dorp F, Ergün S and Rübben
H: Matrix metalloproteinases and their clinical relevance in
urinary bladder cancer. Nat Rev Urol. 8:241–54. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Wojtowicz-Praga SM, Dickson RB and Hawkins
MJ: Matrix metalloproteinase inhibitors. Invest New Drugs.
15:61–75. 1997. View Article : Google Scholar
|
|
26.
|
Yang LH, Zhang HZ, Zhang B, Chen F, Lai
ZH, Xu LF and Jin XQ: Studies on the chemical constituents from the
skin of Bufo bufo gargarizans Cantor. Yao Xue Xue Bao.
27:679–683. 1992.(In Chinese).
|
|
27.
|
Zhang LS, Nakaya K, Yoshida T and Kuroiwa
Y: Bufalin as a potent inducer of differentiation of human myeloid
leukemia cells. Biochem Biophys Res Commun. 178:686–693. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Jing Y, Nakaya K and Han R:
Differentiation of promyelocytic leukemia cells HL-60 induced by
daidzein in vitro and in vivo. Anticancer Res. 13:1049–1054.
1993.PubMed/NCBI
|
|
29.
|
Szabo E, Francis J and Birrer MJ:
Alterations in differentiation and apoptosis induced by bufalin in
cJun overexpressing U-937 cells. Int J Oncol. 12:403–409.
1998.PubMed/NCBI
|
|
30.
|
Takai N, Ueda T, Nishida M, Nasu K and
Narahara H: Bufalin induces growth inhibition, cell cycle arrest
and apoptosis in human endometrial and ovarian cancer cells. Int J
Mol Med. 21:637–643. 2008.PubMed/NCBI
|
|
31.
|
Jiang Y, Zhang Y, Luan J, Duan H, Zhang F,
Yagasaki K and Zhang G: Effects of bufalin on the proliferation of
human lung cancer cells and its molecular mechanisms of action.
Cytotechnology. 62:573–583. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Takai N, Kira N, Ishii T, Yoshida T,
Nishida M, Nishida Y, Nasu K and Narahara H: Bufalin, a traditional
oriental medicine, induces apoptosis in human cancer cells. Asian
Pac J Cancer Prev. 13:399–402. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Hong SH and Choi YH: Bufalin induces
apoptosis through activation of both the intrinsic and extrinsic
pathways in human bladder cancer cells. Oncol Rep. 27:114–120.
2012.PubMed/NCBI
|
|
34.
|
Chueh FS, Chen YY, Huang AC, Ho HC, Liao
CL, Yang JS, Kuo CL and Chung JG: Bufalin-inhibited migration and
invasion in human osteosarcoma U-2 OS cells is carried out by
suppression of the matrix metalloproteinase-2, ERK, and JNK
signaling pathways. Environ Toxicol. Sep 16–2011.(E-pub ahead of
print). View Article : Google Scholar
|
|
35.
|
Kim TH, Ku SK, Lee IC and Bae JS:
Anti-inflammatory functions of purpurogallin in LPS-activated human
endothelial cells. BMB Rep. 45:200–205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Wang YG, Kim SJ, Baek JH, Lee HW, Jeong SY
and Chun KH: Galectin-3 increases the motility of mouse melanoma
cells by regulating matrix metalloproteinase-1 expression. Exp Mol
Med. 44:387–393. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Mori T, Wang X, Aoki T and Lo EH:
Downregulation of matrix metalloproteinase-9 and attenuation of
edema via inhibition of ERK mitogen activated protein kinase in
traumatic brain injury. J Neurotrauma. 19:1411–1419. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Shin DY, Lu JN, Kim GY, Jung JM, Kang HS,
Lee WS and Choi YH: Anti-invasive activities of anthocyanins
through modulation of tight junctions and suppression of matrix
metalloproteinase activities in HCT-116 human colon carcinoma
cells. 25:567–572. 2011.
|
|
39.
|
Chen F, Hori T, Ohashi N, Baine AM, Eckman
CB and Nguyen JH: Occludin is regulated by epidermal growth factor
receptor activation in brain endothelial cells and brains of mice
with acute liver failure. Hepatology. 53:1294–1305. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Yin JQ, Shen JN, Su WW, Wang J, Huang G,
Jin S, Guo QC, Zou CY, Li HM and Li FB: Bufalin induces apoptosis
in human osteosarcoma U-2OS and U-2OS methotrexate300-resistant
cell lines. Acta Pharmacol Sin. 28:712–720. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Dong Y, Yin S, Li J, Jiang C, Ye M and Hu
H: Bufadienolide compounds sensitize human breast cancer cells to
TRAIL-induced apoptosis via inhibition of STAT3/Mcl-1 pathway.
Apoptosis. 16:394–403. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Qi F, Li A, Inagaki Y, Kokudo N, Tamura S,
Nakata M and Tang W: Antitumor activity of extracts and compounds
from the skin of the toad Bufo bufo gargarizans Cantor. Int
Immunopharmacol. 11:342–349. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43.
|
Myal Y, Leygue E and Blanchard AA: Claudin
1 in breast tumorigenesis: revelation of a possible novel ‘claudin
high’ subset of breast cancers. J Biomed Biotechnol.
2010:9568972010.PubMed/NCBI
|
|
44.
|
Newby AC: Matrix metalloproteinases
regulate migration, proliferation, and death of vascular smooth
muscle cells by degrading matrix and non-matrix substrates.
Cardiovasc Res. 69:614–624. 2006. View Article : Google Scholar
|
|
45.
|
Visse R and Nagase H: Matrix
metalloproteinases and tissue inhibitors of metalloproteinases:
structure, function, and biochemistry. Circ Res. 92:827–839. 2003.
View Article : Google Scholar : PubMed/NCBI
|