Introduction
Since their discovery in 1973 by Steinman, dendritic
cells (DCs) have gained the attention of immunologists because they
are extremely efficient in activating T cells (1,2). On
interaction with DCs, naïve CD4+ T cells and
CD8+ T cells can differentiate into antigen-specific
effector T cells with different functions (2). DCs can also interact with cells of
the innate immune system, including natural killer cells,
phagocytes and mast cells (1,2).
Moreover, they have an important role in controlling humoral
immunity by directly interacting with B cells (3,4). In
addition to immune activation, DCs are also involved in immune
suppression (5). These two
opposite functions are important for the normal function of immune
system. However, with respect to cancer immunotherapy,
counter-regulatory responses antagonise the ability to induce
robust and sustained immune responses that can eventually lead to
tumour regression. In the case of therapeutic vaccines, immunity
must be effectively induced when disease is already firmly
established (6–9).
For clinically effective immune responses against
tumours, there is a need to inhibit the expression of the factors
that dampen immune responses in patients. This could potentially
lead to longer activation of DCs, T and NK cells, and might be
beneficial for cancer immunotherapy. Among the immune inhibitory
molecules expressed by DCs is indoleamine 2,3-dioxygenase (IDO), an
enzyme that conveys immunosuppressive effects by degrading
tryptophan, an essential amino acid required for T-cell
proliferation and survival (10–12).
Depletion of tryptophan by IDO in the local microenvironment
induces T-cell apoptosis and impairs normal T-cell function
(13). IDO-expressing DCs can also
induce differentiation of naïve CD4+ T cells into
regulatory T cells (T-regs) that further suppress antitumor
immunity (14). Moreover, various
tryptophan metabolites such as N-formylkynurenine and quinolinate
are directly immuno-suppressive of T-cell activation (10,11).
These findings indicate that the regulation of tryptophan
metabolism by IDO in DCs is a highly adaptable modulator of
immunity. Indeed, injection of IDO-positive DCs into mice
suppressed the activation of antigen-specific T cells in the lymph
nodes draining the injection site (10). Effector T cells starved of
tryptophan were unable to proliferate and enter into G1 cell cycle
arrest. Interestingly, in vitro studies have shown that
IDO-positive DCs were susceptible to killing by naturally occurring
IDO-reactive cytotoxic T cells (15).
Based on the findings indicated above, a promising
strategy for enhancing immunity in patients with cancer would be
the blockade of IDO expression in DCs. To this end, we have
developed several effective small interfering RNA (siRNAs)
targeting IDO that exhibited a silencing potency at nanomolar
concentrations (16). Importantly,
IDO gene silencing enhanced the ability of DCs to stimulate T cells
in vitro (16). In the
present study, we sought to assess the feasibility, safety and
immunogenicity of IDO-silenced DC cancer vaccine in patients with
gynecological cancers such as ovarian cancer. Epithelial ovarian
cancer is the leading cause of death from gynecologic malignancies
(17). Despite an initial response
rate of 65 to 80% to frontline therapy, recurrence arises in the
majority of cases, leading a relative 5-year survival rate of only
47%. Hence, this situation highlights the need of new optimized
therapeutic strategies. Collectively, the data indicate that
IDO-silenced DC vaccine is safe and effective in inducing
antitumour immunity. Indeed, all four vaccinated patients developed
immune response to the vaccine. The ability to enhance the
immunostimulatory potency of DCs by silencing IDO expression would
benefit patients with cancers.
Materials and methods
Patients
The patient characteristics are summarized in
Table I. Two patients with cancer
ovarii stage III C, one patient with metastatic cancer corporis
uteri, and one patient with cancer peritonei were included in the
study. These cancer types are usually considered as ovarian cancer.
All patients had recurrent disease and had received multiple
chemotherapy treatments when they were enrolled in the IDO-silenced
DC vaccine. Prior to being included in the study, patients were
informed of the investigative nature of this study, and written
consent in accordance with institutional regulation was obtained
prior to study entry. The use of the IDO-silenced DC vaccine as
compassionate use was approved by Norwegian Medicine Agency and
conducted in accordance to the provisions of the Declaration of
Helsinki.
 | Table I.Patient characteristics. |
Table I.
Patient characteristics.
| Patient
identity | Age |
Stage/subtypea | Time at
diagnosis | Time to
progression | Chemotherapy prior
to IDO-silenced DC vaccine |
|---|
| CU-1 | 56 | Cancer Ovarii stage
III C | June 2006 | February 2008 | 4 different
chemotherapy combinations |
| CU-2 | 58 | Cancer Corporis
uteri c.met | July 2004 | March 2010 | 1 chemotherapy
combination |
| CU-3 | 45 | Cancer Ovarii stage
III C | September 2007 | September 2007 | 4 different
chemotherapy combinations |
| CU-4 | 57 | Cancer
Peritonei | January 2009 | September 2010 | 2 different
chemotherapy combinations |
Generation of DCs for vaccination
Autologous DC vaccines were produced under standard
Good Manufacturing Practice (GMP) conditions as described elsewhere
(18,19). Fast DCs were generated from
monocytes as described by Tanaka et al (20). In brief, monocyte-rich adherent
cell fractions from leukapheresis products were collected and
cultured for two days in CellGro GMP DC medium supplemented with
GM-CSF and IL-4 and then matured for one day by adding IL-1, IL-6,
TNF-α and PGE-2. The resulting fast DCs (5×107
cells/cuvette) were transfected with either hTERT or suvivin mRNA
(10 μg) in combination with IDO siRNA (15–20 μg) by
square wave electroporation as described elsewhere (18). After transfection, the cells were
cultured for one night prior to cryopreservation into separate
vaccine doses (5×106 cells/0.5 ml).
Tumour antigens and siRNAs
mRNAs encoding hTERT and survivin proteins, two
universal tumour antigens (21,22),
were produced as described previously under standard GMP conditions
(18). The synthesised mRNAs were
5’-capped in order to be translated. With respect to siRNA, the
following set of DNA oligonucleotides (MWG, Ebersberg, Germany)
were used for in vitro transcription. Underlined letters
indicate siRNA sense and antisense transcripts. Sense strand: oligo
1, 5′-CGTTTAATACGACTCACTATAGGTCCGTGAGTTTGTCCTTTCAA-3′;
oligo 2, 5′-TTGAAAGGACAAACTCACGGACTATAGTGAGTCGTATTAAACG-3′.
Antisense strand: oligo 1, 5′-CGTTTAATACGACTCACTATAGAAAGGACAAACTCACGGACTG-3′;
oligo 2, 5′-CAGTCCGTGAGTTTGTCCTTTCTATAGTGAGTCGTATTAAACG-3′. Prior
to transcription, the DNA oligonucleotides were annealed to form
double stranded DNA and siRNA sense and antisense strands were
transcribed from the double-stranded DNA template (40 μg/1
ml reaction) using Ribomax large scale RNA production system-7 as
described by the manufacturer’s instructions (Promega, Madison, WI,
USA). The transcripts were purified under standard GMP conditions
(18). The in vitro
transcribed RNA strands have the following sequence: sense strand,
5′-GGUCCGUGAGUUUGUCCUUUCAA-3′, antisense strand
5′-GAAAGGACAAACUCACGGACUG-3′.
Vaccination
IDO-silenced DC vaccines were described as indicated
above. The four patients received a combination of hTERT (right
arm) and survivin (left arm) DC vaccines supplemented with IDO
siRNA. Patients were revaccinated weekly for 4 weeks after the
first DC injection. Booster vaccinations as a follow-up treatment
were performed each 4 weeks. At each vaccination visit, a
comprehensive assessment of adverse drug reactions, blood
screening, physical examination and assessment of ECOG performance
status (PS) was performed as described previously (19). PBMCs were isolated from patient
peripheral blood using density centrifugation (Lymphoprep; Nycomed,
Oslo, Norway), washed and frozen in RPMI supplemented with 20% FCS
and 10% DMSO until use. Microbiologic and endotoxin tests of DCs,
mRNA and siRNA preparations were performed before use and were
negative at all times.
Delayed-type hypersensitivity
Delayed-type hypersensitivity (DTH) skin tests were
performed at visit one (baseline) and at all the following weekly
vaccination visits from week 2 and onwards. A positive skin
reaction was defined as erythema and induration of >5 mm
diameter 48 h after vaccine injection.
Allogeneic lymphocyte reactions
(MLRs)
Allogeneic CD4+ and CD8+ T
cells were isolated from Buffy coats by direct magnetic labelling
with the appropriate microbeads according to the manufacturer’s
instructions (Invitrogen Dynal AS, Oslo, Norway). Co-cultures were
performed as previously described (23). After 5 days in culture,
proliferation was analysed by [3H]-thymidine
incorporation. The expression of the activation markers presented
in this study was performed after DC vaccine thawing and further
culturing overnight.
Recombinant hTERT and survivin
proteins
A recombinant hTERT sequence encoding amino acids
563–735 of the hTERT protein was cloned into the NdeI site of the
Pet-28b(+) vector (Stratagene, La Jolla, CA, USA) in frame with the
N-terminal 6X His tag/thrombin sequence. The protein was produced
in E. Coli BL21 codon Plus (DE3)-RIPL (Stratagene) and
purified by NiNTA chromatography under denaturing conditions. All
fractions were tested by western blot analysis with anti-His tag
antibodies and the fractions of interest were dialyzed against PBS
buffer and sterile filtered before use. Recombinant human survivin
protein was purchased from BioVision Inc (Milpitas, CA, USA).
Monitoring of T-cell responses
Prior to vaccination and at week 9–10, Tscell
responses were assessed in vitro against recombinant hTERT
and survivin proteins. T-cell proliferation was performed as
described elsewhere and all assays were done in triplicates
(24). Briefly, thawed PBMCs were
seeded at 1.5×105 per well in 96-well plate in 250
μl ex vivo 15 medium in the presence or absence of
recombinant hTERT or survivin (10 μg/ml). After 6 days in
culture, cells were pulsed with [3H]-thymidine for 18 h
prior to harvesting. An antigen-specific response was considered
positive when the stimulatory index was more than three (24). Stimulation index was determined by
dividing the mean counts per minute (cpm) of wells incubated with
the antigen by the mean counts per minute of medium alone. To
evaluate the strength of cell proliferation, the data are presented
as cpm. Cytokine contents in culture supernatants were measured by
commercially available ELISA kits (R&D Systems).
Analysis of IDO expression by western
blotting
IDO expression in the Fast DC preparations
subsequent to siRNA transfection was analyzed using western
blotting. The expression of IDO was also analyzed in mDCs generated
using a standard method to produce monocyte-derived mDCs for
clinical use (18,19). In this protocol, monocytes obtained
from leukapheresis were cultured for five days with GM-CSF and
IL-4. Subsequently, they were cultured for two days with IL-1,
IL-6, TNF-α and PGE2, and then frozen until use.
Flow cytometry
Phenotype of DCs was analysed by direct
immunofluorescence staining of cell surface antigens using FITC or
PE conjugated antibodies against CD80, CD83, CD86, HLA-DR, CCR7,
CD40 and isotype controls. All antibodies were purchased from Dako
(Glostrup, Denmark) or eBioscience (BD Biosciences, Pharmingen, San
Diego, CA). After staining on ice for 30 min, samples were washed
twice and then analysed by flow cytometry using a FACSCanto II. The
data were analysed using FlowJo software.
Analysis of tryptophan catabolism
To analyse tryptophan catabolism, dendritic cells
were plated in Hanks’ buffered salt solution (HBSS) containing 100
μM L-tryptophan (Sigma-Aldrich, St. Louis, MO, USA).
Subsequent to overnight incubation at 37°C, supernatants were
harvested and analysed for tryptophan contents by commercially
available ELISA kit (Abnova, Taipei City, Taiwan).
Statistical analysis
All in vitro assays were performed in
triplicate, and values are plotted as mean ± standard deviation.
Values were compared using Student’s t-test. P-values <0.05 were
considered significant.
Results
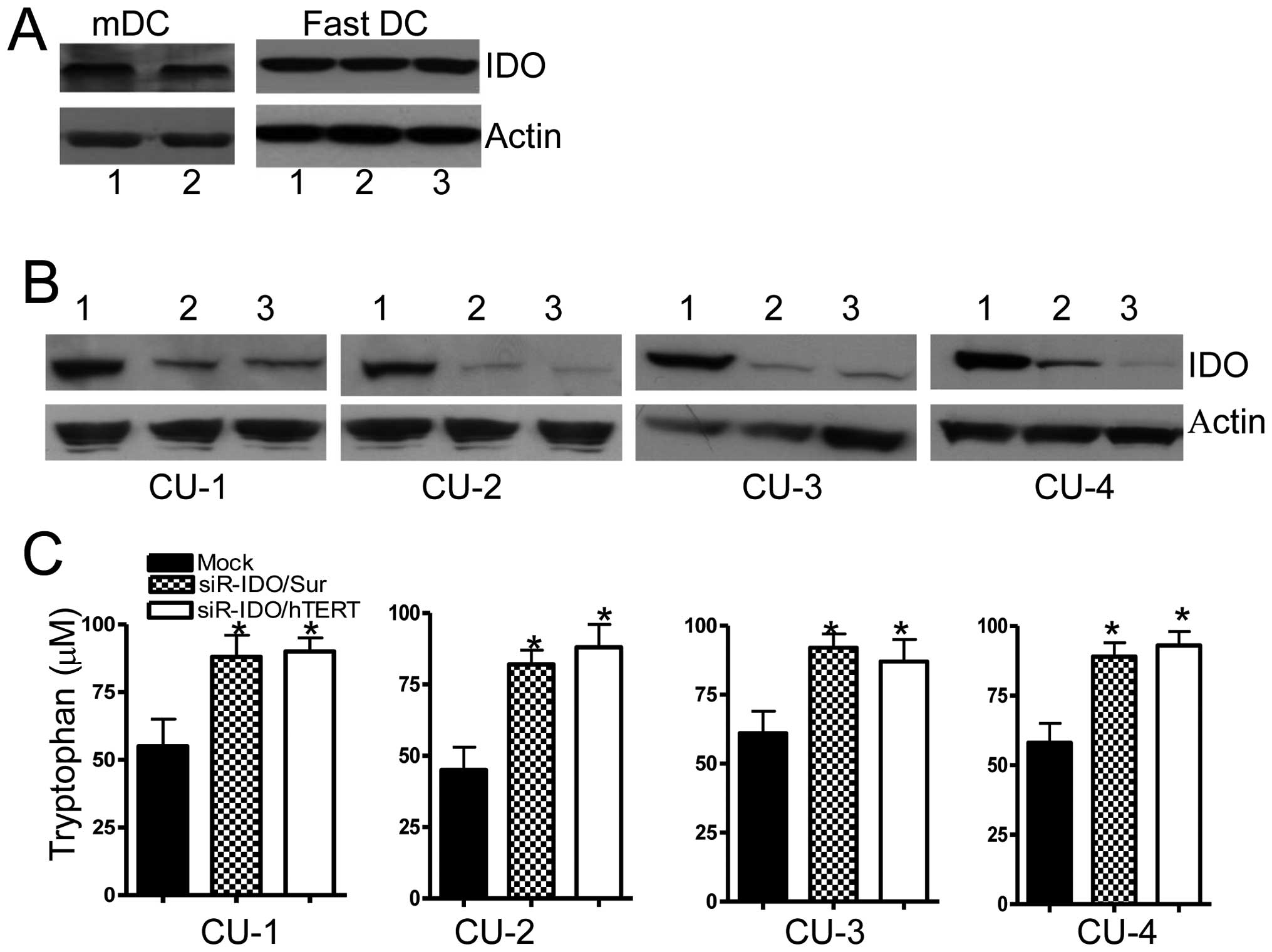
IDO expression in standard DC vaccine
preparations
Expression of IDO by DCs may create local tolerance
by directly suppressing T-cell activation and enhancing regulatory
T-cell function. Therefore, we have analysed IDO expression in
clinical grade DCs generated for therapeutic vaccination of cancer
patients. In total, 10 DC preparations were analysed by western
blotting and all expressed IDO at the protein level (Fig. 1A as an illustrative example). These
data are in accordance with recent reports showing IDO upregulation
in human DCs upon in vitro maturation (25). The induction of IDO expression
subsequent to in vitro maturation of DCs might be a major
problem for the failure of DC-based immunotherapy.
IDO gene silencing in Fast DC vaccine
preparations
Having demonstrated that IDO is expressed in DC
vaccine preparations, next we analysed the silencing potency of the
in vitro transcribed siRNAs. This siRNA was designed based
on our previous study reporting on the identification of IDO siRNAs
that function at low concentrations (16). In this study, siRNA-4 was optimized
for efficient in vitro transcription under standard GMP
conditions (18). Fast DC
preparations from patients with ovarian cancer were transfected
ex vivo with IDO siRNA along with mRNA encoding tumour
antigens hTERT or survivin. As shown in Fig. 1B, the siRNA effectively inhibited
IDO expression in all 8 fast DC vaccine preparations.
To ascertain the enzymatic activity of IDO, we have
measured tryptophan catabolism in supernatants of IDO-silenced DCs
and IDO-positive DCs using tryptophan ELISA kit. In contrast to
IDO-silenced DCs, the tryptophan concentration decreased
significantly in IDO-positive DCs (mock, P<0.05) (Fig. 1C). Therefore, IDO is a functional
enzyme in clinical DC preparations as suggested previously
(25).
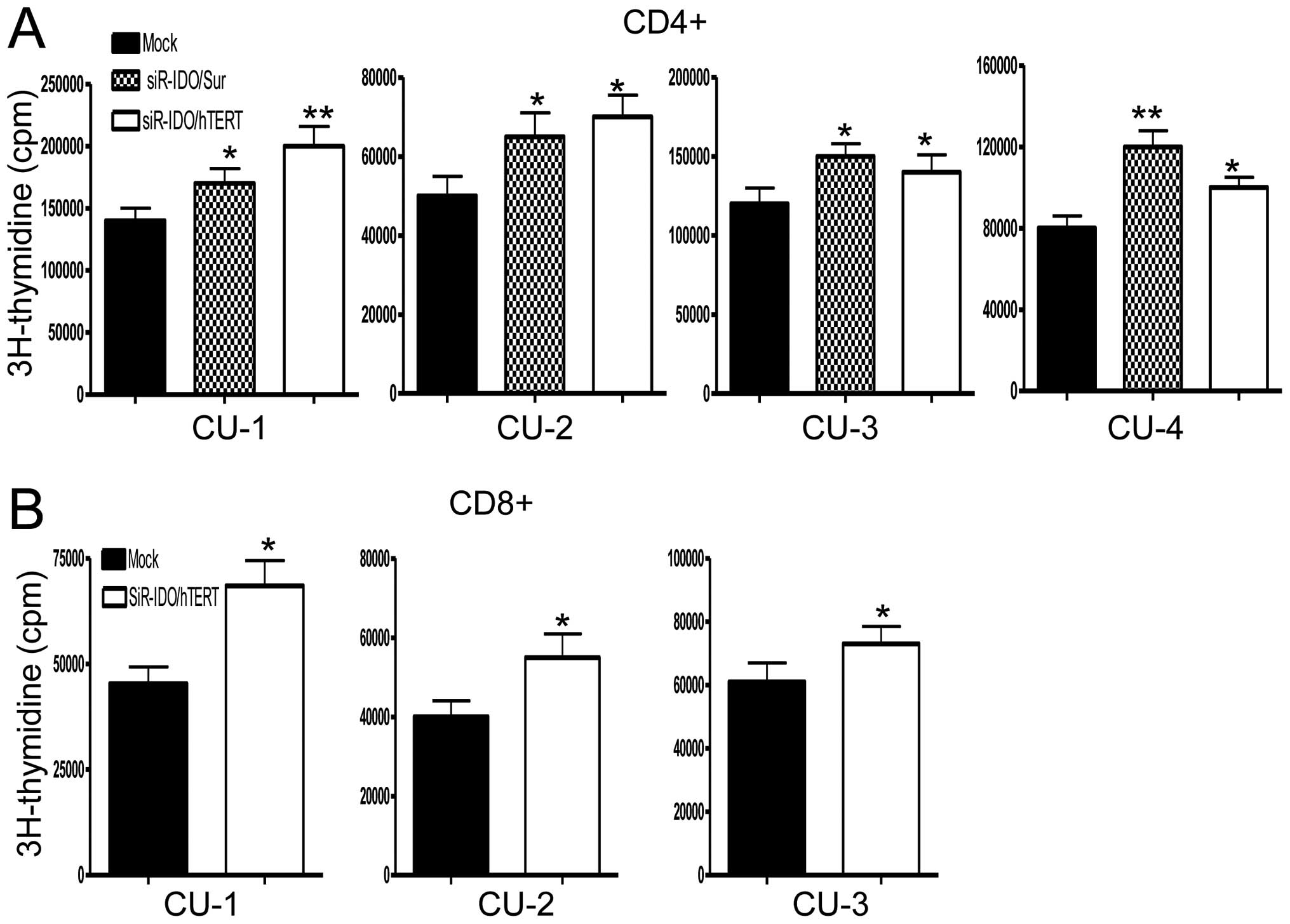
Allogeneic stimulatory potency of
IDO-silenced DC vaccines
While DC-based therapy holds great promise for
treating cancers, the clinical responses of conventional DC cancer
vaccines are very low according to many studies (7). Furthermore, it is not yet known to
what extent IDO contribute to tumour-related immune suppression in
cancer patients. To determine if IDO-silenced DC activate T cells
more efficiently than IDO-positive DCs, mixed lymphocyte reaction
assay was used to measure the allostimulatory potency of these
cells (Fig. 2A and B).
IDO-silenced DC vaccine preparations showed a higher
allostimulatory capacity of both CD4+ and
CD8+ T cells than their counterparts IDO-positive DCs
(mock DCs, P<0.05), suggesting that the presence of tryptophan
is a limiting factor in the cultures. The data are also consistent
with the consumption of tryptophan by IDO-positive DCs shown in
Fig. 1C.
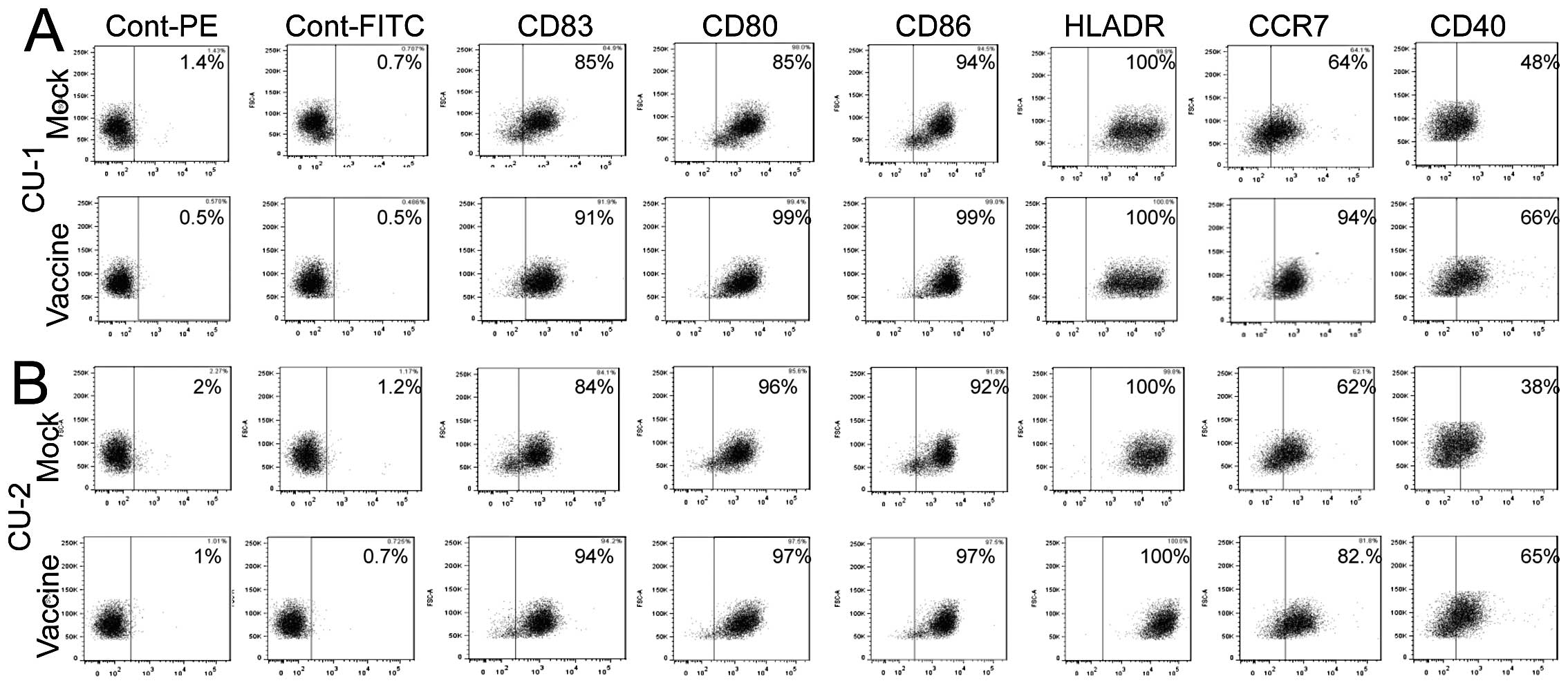
Phenotypic analysis of DC vaccine
preparations
The surface expression of various molecules related
to antigen presentation, migration and T-cell activation was
analysed by means of flow cytometry. The expression of CD80, CD83,
CD86, and HLA-DR molecules was comparable in IDO-silenced DCs
(vaccine) and IDO-positive DCs (mock) (Fig. 3A and B, as representative
examples). However, in three patients (CU-1, CU-2 and CU-3), the
expression of CD40 and CCR7 was significantly increased in
IDO-silenced DCs when compared to their mock-transfected
counterparts (P<0.05). For example, the percentages of
IDO-silenced DC expressing CCR7 from patients CU-1, CU-2 and CU-3
were 94, 82 and 67%, while that in mock DCs were 64, 62 and 47%,
respectively. CCR7 is required for DC migration to draining lymph
nodes where they activate T cells. The upregulation of CCR7 and
CD40 is most likely due to the activation of intracellular retinoid
acid inducible-gene 1 (RIG-1) by 5’-triphosphate siRNA (see
Discussion). It should be noted that the expression profiles of the
analysed markers in others fast DC preparations transfected with
mRNA encoding hTERT or survivin are similar to those of patient
mock DCs (data not shown).
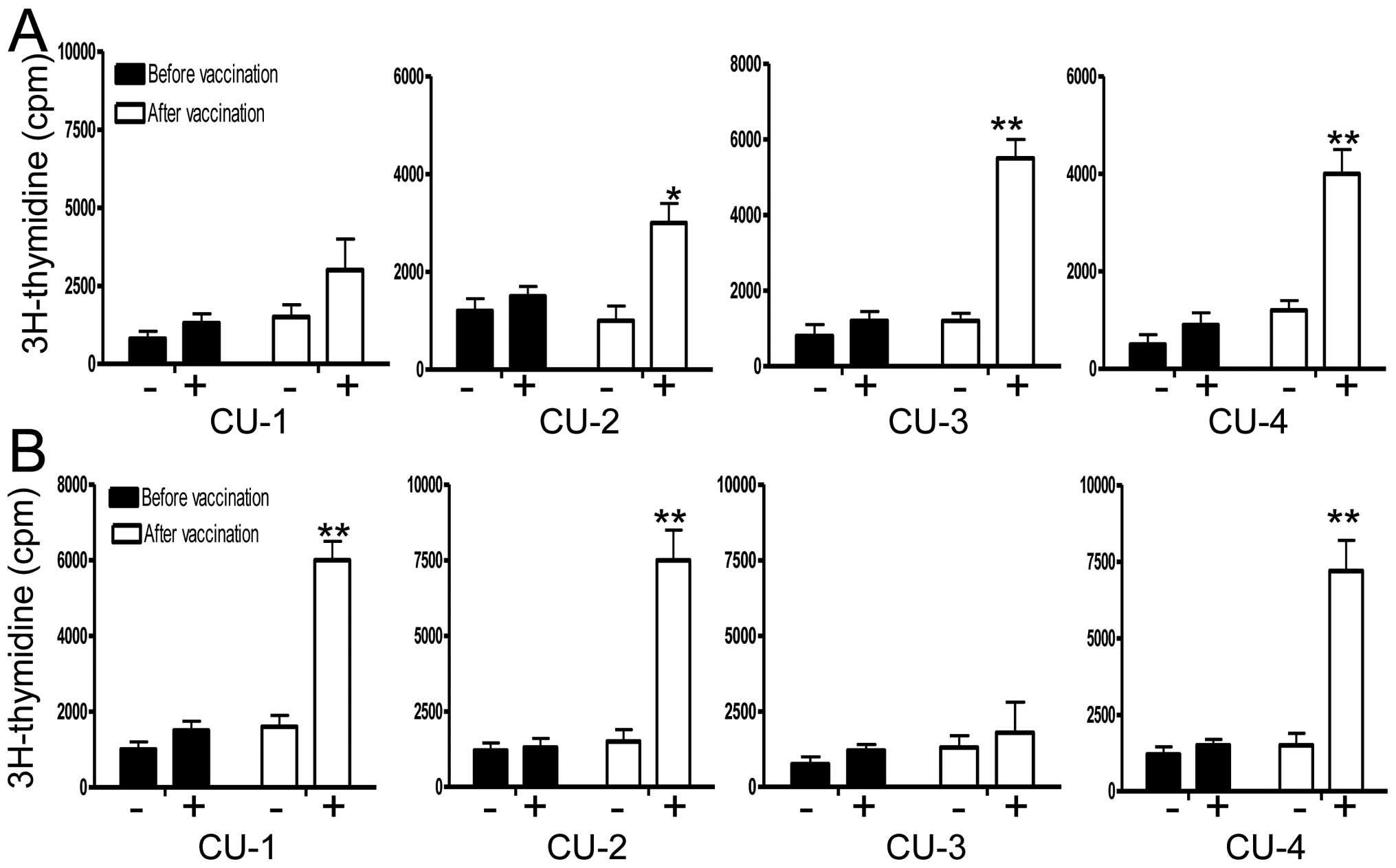
Immune and clinical responses
We next examined the safety of the DC vaccine and
whether it would induce antigen-specific immune response and
clinical improvement. The immunization was well tolerated except
for the development of flu-like symptoms, which is reminiscent of
effective induction of immunity. All four patients developed a DTH
response (Table II). The induction
of specific T-cell responses during treatment was analysed in
peripheral blood before treatment, and at week 9–10 after
vaccination. In this assay, we used recombinant hTERT and survivin
proteins, enabling the detection of most T-cell epitopes. As shown
in Fig. 4, the patients developed
T-cell response against hTERT and survivin tumour antigens.
Notably, there was a 2- to 6-fold increase in T-cell proliferation
when compared to unstimulated PBMCs.
| Table II.Immune and clinical responses to
IDO-DC vaccination. |
Table II.
Immune and clinical responses to
IDO-DC vaccination.
| Patient
identity | DTH response |
Immune-response | Chemotherapy
treatment free interval before IDO-DCs (months) | Time to progressive
disease with IDO-DCs (months) |
|---|
| CU-1 | + | + | 12 | 4.5 |
| CU-2 | + | + | 10 | 13(+9a) |
| CU-3 | + | + | 0 | 21b |
| CU-4 | + | + | 14 | 11 |
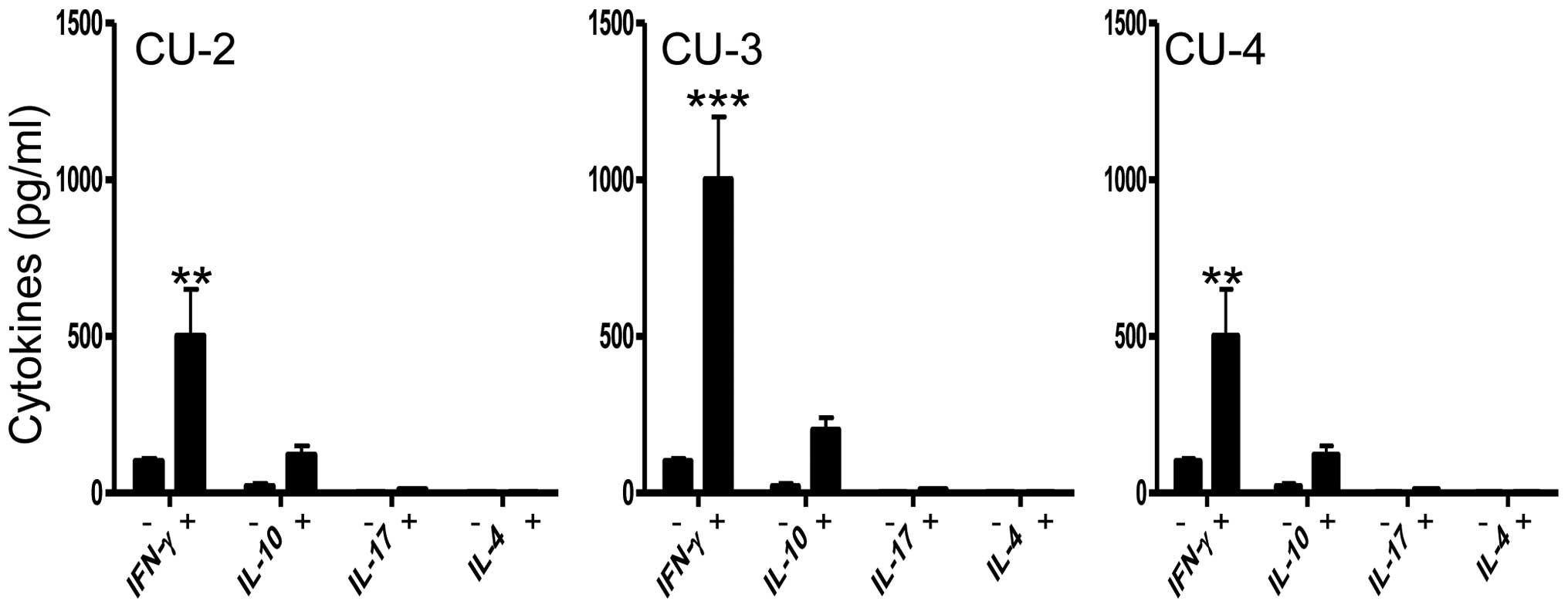
To determine the nature of the immune response, we
have assessed the levels of interferon γ (IFN-γ), interleukin
(IL)-4, IL-10 and IL-17 in T-cell culture supernatants from
patients who responded to hTERT protein. A significant increase in
IFN-γ was detected in three patients, indicating a Th-1 type
response (Fig. 5). IFN-γ helps
CD8+ T cells to differentiate into cytotoxic T
lymphocytes capable of killing MHC class I positive tumours.
With regard to the clinical response (Table II), the four vaccinated patients
maintained stable disease subsequent to vaccination. Interestingly,
patient CU-2 and CU-3 showed a significant disease free interval
compared to that obtained with only chemotherapy (13 vs 10 months,
and 21 vs 0 months, respectively). When patients progressed
following DC vaccination they were given a combination of both
chemotherapy and DC vaccine (CU-2, CU-4). In patient CU-2 this
combination resulted in a partial remission and the patient is
still under therapy. It should be noted that patient CU-3 never
achieved a complete disease free interval on chemotherapy since
2007. However, when she was given DC vaccination combined with
chemotherapy she obtained a partial remission and the decline in
tumour volume and markers continues. This observation supports the
potential benefits of combining DC vaccine and chemotherapy
(26). Fig. 6 illustrates the treatment schedule
and clinical outcome of patient CU-3.
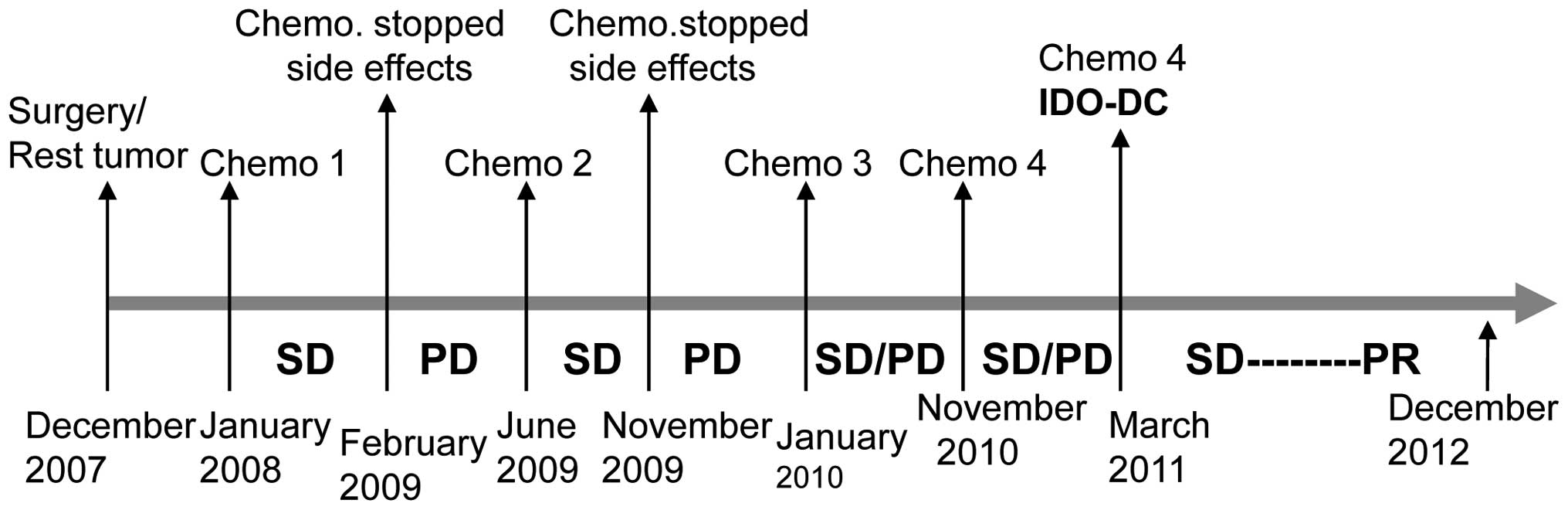 | Figure 6.Timeline for the treatment and
clinical development. The figure illustrates the treatment schedule
for the patient with ovarian cancer (CU-3). After surgery, the
patient has received four combinations of chemotherapy (chemo)
prior DC vaccine. Chemo 1, carboplatin, taxol, avastin; chemo 2,
carboplatin, caleyx; chemo 3, taxol; chemo 4, taxol, avastin; SD,
stable disease; PD, progressive disease; PR, partial remission. |
Discussion
The clinical efficacy of DC-based vaccines for
cancer immunotherapy is influenced by various interplaying,
immunomodulating factors including IL-10 and IDO. In the present
study, we evaluated the safety and efficacy of an IDO-silenced DC
vaccine in four patients with ovarian cancers extensively
pre-treated with standard chemotherapy and with recurrent disease.
We show that even in these patients, it is possible to induce
immune responses leading to clinical improvement. Indeed, all four
patients developed a DTH response and flu-like symptoms after
vaccination, and a strong hTERT-specific T-cell response was
detected in peripheral blood of three patients. Also, the patients
developed survivin-specific T-cell response. These immune responses
were related to objective clinical responses. Given these
encouraging findings, the downregulation of IDO expression in DC
cancer vaccines should be considered as a new attractive approach
for generating therapeutic immunity against tumours.
In general, the development of an immune response is
controlled by DCs (1). Once T
cells are effectively primed, pro-inflammatory cytokines such as
IFN-γ and T-reg signals such as CTLA-4, induce IDO expression in
DCs resulting in their conversion into tolerogenic DCs. Such DCs
can inhibit T-cell expansion as well as the generation of adaptive
T-reg cells from naïve CD4+ T cells (27). IFN-γ is produced primary by NK
cells, activated CD4+ and CD8+ T cells and
NKT cells. Reverse signalling via B7 molecules (CD80/86) expressed
on DCs after interaction with CD28 on T cells has also been shown
to induce IDO expression in DCs. With respect to cancer vaccines,
the induction of IDO expression can occur during in vitro
maturation of DCs as well as in vivo after T-cell activation
as indicated above. IDO gene silencing by siRNA is expected to
block IDO expression during this entire process and thereby
enhancing the potency of DC vaccine. Gene silencing in fast DCs did
not weaken after nine days in culture (data not shown). The
inclusion of siRNAs into DC cancer vaccines is a straightforward
strategy that does not require additional technology or
modification to the mRNA transfection procedure (18). Moreover, the data suggest that RNAi
using IDO siRNA is safe in humans. Indeed, none of the vaccinated
patients have developed other side-effects than flu-like symptoms
following vaccination. Flu-like symptoms are considered to be a
good sign of efficient immune responses (7). It should be noted that vaccination of
mice with suppressor of cytokine signalling-1 (SOCS-1)-silenced DCs
also enhanced antigen-specific antitumor immunity in in vivo
murine models (28). This provides
more evidence to support the knockdown of immuno-suppressive genes
in DC cancer vaccines. In this respect, a vaccine that combines the
inhibition of IDO, IL-10 and SOCS-1 is under investigation.
Out of the four vaccinated patients, only one
patient (CU-2) is HLA-A2 positive. However, pentamer staining with
a newly identified hTERT T-cell epitope (674–683 pentamer) 19 did
not detect any peripheral CTL response, indicating that the
response to recombinant hTERT was directed towards other T-cell
epitopes that are under investigation. Patient PBMC incubated with
hTERT protein produced a significant amount of IFN-γ, but not IL-4
suggesting the induction of Th-1 immune polarization that may be
contributed to the clinical responses seen in the patients.
Another feature of the IDO-silenced DC vaccine is
the upregulation of CD40 and CCR7 seen in 3 out of 4 vaccinated
patients. Previously, we have shown that in vitro
transcribed siRNA carrying uncapped 5’-triphosphate can activate
immature monocyte-derived DCs via intracellular pathogen
recognition receptor RIG-1 (16,29).
RIG-1 is a cytosolic sensor of viral RNA-bearing 5’-triphosphate,
triggering a type 1-interferon and proinflammatory cytokine
response (30,31). A recent study also showed that
in vitro transcribed siRNAs targeting Bcl-2 induced the
expression of interferons and activated DCs in a mouse model when
injected intravenously (32).
Similarly, Kubler et al (33) showed that RIG-1 activation can
trigger cytokine secretion and DC maturation. The ability of siRNA
to activate cytolytic RIG-1 in DCs coupled with the silencing of
gene expression raises the prospect of 5’-triphosphate-bearing
siRNAs being developed as therapeutics and adjuvant for cancer
vaccines. Simulating viral infection with agents such as uncapped
5’-triphosphate siRNA, rather than aseptic inflammation (using
cytokine cocktails) can induce the differentiation of monocytes
into DCs with optimal function. In this respect, Cheong et
al demonstrated that blood monocytes can be mobilized to
develop into ‘real’ DCs in vivo by microbial danger signals
such as lipopolysaccharide (34).
They acquired antigen-presenting function equal to or in some cases
superior to bone marrow derived DCs.
The discovery of RNAi has revolutioned molecular
biology by allowing selective knockdown of genes in a manner more
potent than that of any other previous strategies (35). Despite growing interest in the use
of siRNA in humans (36–39), one of the major challenges to the
clinical development of gene silencing by siRNAs has been in
vivo delivery in general, and particularly the ability to
selectively target siRNA molecules to specific cell types (40,41).
The use of siRNA in an ex vivo setting as demonstrated in
the current study should overcome the delivery and targeting
problems mentioned above. With respect to cancer immunotherapy, our
data also indicate that transfection with IDO siRNA does not have a
negative effect on DC phenotype and function. In contrast, it does
produce ‘superior’ DCs compared to their mock transfected
counterparts. The in vitro enhanced DC function seen may
contribute to the clinical improvement seen in patients following
vaccination. These encouraging results, coupled with recent
findings that siRNA-bearing 5’-triphosphate can activate DCs,
should be further explored as a platform for inducing enhanced
anti-tumor immunity in humans. The current study sets the stage for
testing IDO-silenced DC vaccine in cancer patients. Existing
DC-based vaccine clinical trials would likely benefit from
additional modifications to silence inhibitory molecules such as
IDO, IL-10 and SOCS-1.
Acknowledgements
The authors are grateful for
assistance provided by the Cell therapy members for vaccine
preparation and clinical follow-up of the patients. This study was
supported in part by the Norwegian Ministry of Health, the
Norwegian Cancer Society and Inger and John Fredriksen Foundation
for ovarian Cancer.
References
|
1.
|
Steinman RM and Banchereau J: Taking
dendritic cells into medicine. Nature. 449:419–426. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Banchereau J and Steinman RM: Dendritic
cells and the control of immunity. Nature. 392:245–252. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Jego G, Pascual V, Palucka AK and
Banchereau J: Dendritic cells control B cell growth and
differentiation. Curr Dir Autoimmun. 8:124–139. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Shaw J, Wang YH, Ito T, Arima K and Liu
YJ: Plasmacytoid dendritic cells regulate B-cell growth and
differentiation via CD70. Blood. 115:3051–3057. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Steinman RM, Hawiger D and Nussenzweig MC:
Tolerogenic dendritic cells. Annu Rev Immunol. 21:685–711. 2003.
View Article : Google Scholar
|
|
6.
|
Hoos A, Britten CM, Huber C and
O’Donnell-Tormey J: A methodological framework to enhance the
clinical success of cancer immunotherapy. Nature Biotechnol.
29:867–870. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Palucka K and Banchereau J: Cancer
immunotherapy via dendritic cells. Nature Rev. 12:265–277.
2012.PubMed/NCBI
|
|
8.
|
Sioud M and Sørensen D: Generation of an
effective anti-tumor immunity after immunization with xenogeneic
antigens. Eur J Immunol. 33:38–45. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Sioud M: Does our current understanding of
immune tolerance, autoimmunity, and immunosuppressive mechanisms
facilitate the design of efficient cancer vaccines? Scand J
Immunol. 70:516–525. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Munn DH, Sharma MD, Lee JR, Jhaver KG,
Johnson TS, Keskin DB, Marshall B, Chandler P, et al: Potential
regulatory function of human dendritic cells expressing indoleamine
2,3-dioxyggenase. Science. 297:1867–1870. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Hwu P, Du MX, Lapointe R, Do M, Taylor MW
and Young HA: Indoleamine 2,3-dioxygenase production by human
dendritic cells results in the inhibition of T cell prolifieration.
J Immunol. 164:3596–3599. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Braun D, Longman RS and Albert ML: A
two-step induction of indoleamine 2,3 dioxygenase (IDO) activity
during dendritic cell maturation. Blood. 106:2375–2381. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Munn DH, Sharma MD and Hou D: Expression
of indoleamine 2,3-dioxygenase by plasmacytoid dendritic cells in
draining-draining lymph noedes. J Clin Invest. 114:280–290. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Jonuleit H, Schmitt E, Stenbrink K and Enk
AH: Dendritic cells as a tool to induce anergic and regulatory T
cells. Trends Immunol. 22:394–400. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Sørensen RB, Berge-Hansen L, Junker N,
Hansen CA, Hadrup SR, Schumacher TN, Svane IM, Becker JC, thor
Straten P and Andersen MH: The immune system strikes back: cellular
immune responses against indoleamine 2,3-dioxygenase. PLoS One.
4:e69102010.PubMed/NCBI
|
|
16.
|
Flatekval GF and Sioud M: Modulation of
dendritic cell function and maturation with mono- and bifunctional
small interfering RNAs targeting indoleamine 2,3-dioxygenase.
Immunology. 128:e837–e848. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Guppy AE, Nathan PD and Rustin GJ:
Epithelial ovarian cancer: a review of current management. Clin
Oncol. 17:399–411. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Sæbøe-Larssen S, Fossberg E and Gaudernack
G: mRNA-based electrotransfection of human dendritic cells and
induction of cytotoxic T lymphocyte responses against the
telomerase catalytic subunit (hTERT). J Immunol Methods.
259:191–203. 2002.PubMed/NCBI
|
|
19.
|
Suso EMI, Dueland S, Rasmussen AM, Vetrhus
T, Aamdal S, Kvalheim G and Gaudernack G: hTERT mRNA dendritic cell
vaccination: complete response in a pancreatic cancer patient
associated with response against several hTERT epitopes. Cancer
Immunol Immunother. 60:809–818. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Tanaka F, Yamaguchi H, Haraguchi N,
Mashino K, Ohta M, Inoue H and Mori M: Efficient induction of
specific cytotoxic T lymphocytes to tumor rejection peptide using
functional matured 2 day-cultured dendritic cells derived from
human monocytes. Int J Oncol. 29:1263–1268. 2006.
|
|
21.
|
Liu JP, Chen W, Schwarer AP and Li H:
Telomerase in cancer immunotherapy. Biochim Biophys Acta.
1805:35–42. 2010.PubMed/NCBI
|
|
22.
|
Andersen MH, Svane IM, Becker JC and
Straten PT: The universal character of the tumor-associated antigen
survivin. Clin Cancer Res. 13:5991–5994. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Moberslein M and Sioud M: Galectin-1 and
-3 gene silencing in immature and mature dendritic cells enhances T
cell activation and interferon-γ production. J Leuko Biol.
91:461–467. 2012.PubMed/NCBI
|
|
24.
|
Sioud M, Kjeldsen-Kragh J, Quayle AJ,
Wiker HG, Sørskaar D, Natvig JB and Førre O: Immune responses to
18.6 and 30-kDa mycobacterial antigens in rheumatoid patients, and
V beta usage by specific synovial T-cell lines and fresh T cells.
Scand J Immunol. 34:803–812. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Wobser M, Voigt H and Houben R: Dendritic
cell based antitumor vaccination: impact of functional indoleamine
2,3-dioxygenase expression. Cancer Immunol Immunother.
56:1017–1024. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Hodge JW, Ardiani A, Farsaci B, Kwilas AR
and Gmeiro SR: The tiping point for combination therapy: cancer
vaccines with radiation, chemotherapy, or targeted small molecule
inhibitors. Semin Oncol. 39:323–339. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Munn DH and Mellor AL: IDO expression by
dendritic cells: tolerance and tryptophan catabolism. Nat Rev
Immunol. 4:762–774. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Shen L, Evel-Kabler K, Strube R and Chen
SY: Silencing of SOCS1 enhances antigen presentation by dendritic
cells and antigen-specific anti-tumor immunity. Nat Biotechnol.
22:1546–1553. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Furset G and Sioud M: Design of
bifunctional siRNAs: combining immunostimulation and gene-silencing
in one single siRNA molecule. Biochem Biophys Res Commun.
352:642–649. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Hornung V, Ellegast J, Kim S, Brzózka K,
Jung A, Kato H, Poeck H, et al: 5′-Triphosphate RNA is the ligand
for RIG-I. Science. 314:994–997. 2006.
|
|
31.
|
Pichlmair A, Schulz O, Tan CP, Naslund TI,
Liljestrom P, Weber F and Reis e Sousa C: RIG-I-mediated antiviral
responses to single-stranded RNA bearing 5′-phosphates. Science.
314:997–1001. 2006.PubMed/NCBI
|
|
32.
|
Poeck H, Besch R and Maihoefer C:
5′-triphosphate-siRNA: turning gene silencing and RIG-I activation
against melanoma. Nat Med. 14:1256–1263. 2008.
|
|
33.
|
Kubler K, Gehrke N and Riemann S: Targeted
activation of RNA helicase retinoic acid-inducible gene-I induces
proimmunogenic apoptosis of human ovarian cancer cells. Cancer Res.
70:5293–5304. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Cheong C, Matos I and Choi JH: Microbial
stimulation fully differentiates monocytes to DC-SIGN/CD209(+)
dendritic cells for immune T cell areas. Cell. 143:416–429.
2010.PubMed/NCBI
|
|
35.
|
Fire A, Xu S and Montgomery MK: Potent and
specific genetic interference by double-stranded RNA in
Caenorhabditis elegans. Nature. 391:806–811. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Kaiser PK, Symons RCA and Shah SM: A
randomized, double-blind, placebo-controlled study of an RNAi-based
therapy directed against respiratory syncytial virus. Proc Natl
Acad Sci USA. 107:8800–8805. 2010. View Article : Google Scholar
|
|
37.
|
Zamora MR, Budev M and Rolfe M: RNA
interference therapy in lung transplant patients infected with
respiratory syncytial virus. Am J Resp Critical Care Med.
183:531–538. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
DiGiusto DL, Krishnan A and Li L:
RNA-based gene therapy for HIV with lentiviral vector-modified
CD34(+) cells in patients undergoing transplantation for
AIDS-related lymphoma. Sci Transl Med. 2:36ra432010.PubMed/NCBI
|
|
39.
|
Leachman SA, Hickerson RP and Schwartz ME:
First-inhuman mutation-targeted siRNA Phase I trial of an inherited
skin disorder. Mol Ther. 18:442–446. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Tiemann K and Rossi JJ: RNAi-based
therapeutics-current status, challenges and prospects. EMBO Mol
Med. 1:142–151. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Behlke MA: Progress towards in vivo
use of siRNAs. Mol Ther. 13:644–670. 2006. View Article : Google Scholar : PubMed/NCBI
|