Introduction
Hepatocellular carcinoma (HCC) is currently the
sixth most common malignancy and the third leading cause of
cancer-related death worldwide, with approximately 695,900 deaths
in 2008 (1,2). Although hepatectomy, liver
transplantation, and ablation are the available treatment options
for patients with HCC, high postoperative metastasis or recurrence
rate remain the major obstacles that influence long-term survival,
while most recurrences are due to invasion-related spreading.
Therefore, there is an urgent need for a better understanding of
the molecular mechanisms that contribute to HCC metastasis and for
new potential therapeutic strategy.
Different molecular events and phenotypic changes
are involved in cancer cell disintegration and migration into
distant organs or tissues. The epithelial-to-mesenchymal transition
(EMT), which is characterized by the loss or downregulation of
epithelial markers (E-cadherin) and upregulation of mesenchymal
markers (Vimentin), is a crucial step in tumor invasion and
metastasis (3,4). Furthermore, HCC is a hypervascular
tumor and neovascularization is a common phenomenon. The
development of neovasculature plays an important role in the growth
and metastasis of HCC (5).
Mortalin, which is also known as heat shock protein
75 (HSP75), is low or undetectable in the normal tissues, while
highly expressed in several epithelial carcinomas (6,7).
Overexpression of Mortalin interacts with the wild-type tumor
suppressor protein p53 and modulates the Ras-Raf-MAPK pathway and
then increase the malignancy of tumor cells (8–10).
Therefore Mortalin appears to play a crucial role in tumorigenesis
and metastasis. Recently, it was reported that Mortalin
overexpression in HCC was associated with venous infiltration and
advanced tumor stages and thus Mortalin was implicated as a tumor
marker for predicting early recurrence (11). Nevertheless, there are no previous
studies on the role of Mortalin in HCC, EMT and angiogenesis.
In the present study, we observed that the high
expression of Mortalin is an important regulator inducing EMT, but
it does not promote angiogenesis in HCC. Furthermore, we
demonstrated that the decreased expression of Mortalin is
accompanied by an inhibition of EMT in the HCC cell line
MHCC97H.
Materials and methods
Cell culture
Five human hepatoma-derived cell lines (Hep3B,
MHCC97H, HepG2, MHCC97L and HCCLM3) with various metastatic
potentials and a normal liver cell line L02 were purchased from
Liver Cancer Institute of Zhongshan Hospital, Shanghai, China.
MHCC97-L, MHCC97-H and HCCLM3 belonged to the same genetic
background and had a stepwise increasing metastatic potential
(12,13). The cells were maintained in
Dulbecco’s modified Eagle’s medium (DMEM) (Gibco, NY, USA) with 10%
(vol/vol) fetal bovine serum (FBS) (Hyclone, Logan, UT, USA), 100
U/ml penicillin and 100 U/mll streptomycin at 37°C in a humidified
incubator supplemented with 5% CO2.
Patients and tissue samples
Tumor and paracarcinomatous tissues for
immunohistochemistry were obtained from 96 patients undergoing
curative hepatectomy for HCC at Department of Hepatic Surgery,
Anhui Provincial Hospital Affiliated to Anhui Medical University
from June, 2007 to June, 2010. Frozen HCC tumor and corresponding
paracarcinomatous tissues were obtained from 13 patients for qPCR
and western blotting. Ten normal liver samples were acquired from
the patients who had received an operation due to hepatic trauma as
the controls. No patient had received adjuvant therapies before
surgery. These patients included 76 males and 24 females with a
mean age of 56.4 years (range, 21–80 years). Tumor size was
measured as the largest dimension of the tumor by gross
examination; the pathological tumor stage was defined according to
the sixth edition of the tumor-node-metastasis (TNM) classification
of the International Union Against Cancer; tumor differentiation
was defined according to the Edmondson grading system (14). All tissue diagnoses were confirmed
by three experienced pathologist who were unaware of patients’
clinical and laboratory data. Clinicopathological parameters of the
HCC patients are described in Table
I. The study protocol conformed to the ethical guidelines of
the declaration of Helsinki. Ethics approval for the study of human
subjects was obtained from the research ethics committee of Anhui
Provincial Hospital Affiliated to Anhui Medical University and
written informed consent was obtained from all the patients.
 | Table I.Clinicopathological factors and the
expression of Mortalin in 100 patients with HCC. |
Table I.
Clinicopathological factors and the
expression of Mortalin in 100 patients with HCC.
| Characteristics | Case | Mortalin expression
| χ2 | P-value |
|---|
| Low | High |
|---|
| Age (years) | | | | | |
| ≤55 | 28 | 3 | 25 | 2.421 | 0.210 |
| >55 | 72 | 20 | 52 | | |
| Gender | | | | | |
| Male | 76 | 19 | 57 | 0.322 | 0.570 |
| Female | 24 | 4 | 20 | | |
| Tumor size
(cm) | | | | | |
| ≤5 | 38 | 5 | 33 | 3.352 | 0.067 |
| >5 | 62 | 18 | 44 | | |
| Tumor nodules | | | | | |
| Single | 68 | 18 | 50 | 0.898 | 0.343 |
| Multiple | 32 | 5 | 27 | | |
| Tumor capsula | | | | | |
| Complete | 70 | 13 | 57 | 2.584 | 0.108 |
| None | 30 | 10 | 20 | | |
| AFP (ng/ml) | | | | | |
| ≤400 | 31 | 7 | 24 | 0.004 | 0.947 |
| >400 | 69 | 16 | 53 | | |
| ICGR15
(%) | | | | | |
| ≤10 | 79 | 17 | 62 | 1.697 | 0.193 |
| >10 | 21 | 6 | 15 | | |
| HBsAg | | | | | |
| Positive | 84 | 20 | 64 | 0.014 | 0.907 |
| Negative | 16 | 3 | 13 | | |
| Liver
cirrhosis | | | | | |
| Present | 87 | 21 | 66 | 0.004 | 0.951 |
| Absent | 11 | 2 | 9 | | |
| Child-Pugh
grade | | | | | |
| A | 79 | 15 | 64 | 3.420 | 0.064 |
| B | 21 | 8 | 13 | | |
| Edmondson
grade | | | | | |
| I–II | 67 | 20 | 47 | 4.272 | 0.039 |
| III–IV | 33 | 3 | 30 | | |
| TNM stage | | | | | |
| I–II | 64 | 21 | 43 | 8.188 | 0.004 |
| III–IV | 36 | 2 | 34 | | |
| Invasion and
metastasis | | | | | |
| Absent | 61 | 19 | 42 | 4.742 | 0.029 |
| Present | 39 | 4 | 35 | | |
RNA extraction, reverse transcription and
real-time quantitative PCR (qPCR)
Total RNA was extracted from six cell lines, normal
liver tissues, snap-frozen HCC tumor and corresponding
paracarcinomatous tissues using the TRIzol reagent (Invitrogen,
Carlsbad, CA, USA). The quality of mRNA was evaluated by the
OD260/OD280 ratio and samples were used only when the ratio was
between 1.8 and 2.0. cDNA was synthesized according to the
instructions of the Revert Aid First Strand cDNA synthesis kit
(Invitrogen). The sequences of Mortalin-specific primers were
designed as follows: forward, 5′-GAGAGACAGGGGTTGATTTGAC-3′;
reverse, 5′-GCA CAGATGAGGAGAGTTCACA-3′. In addition, the sequences
of Vimentin-specific primers were designed as follows: forward,
5′-CCGACACTCCTACAAGATTTAGA-3′; reverse,
5′-CAAAGATTTATTGAAGCAGAACC-3′. For standardization of RNA quality
control, the expression of glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) in each sample was quantified by using the primer set
5′-AAGGTCATCCCTG AGCTGAAC-3′ (forward) and 5′-ACGCCTGCTTCACCACC
TTCT-3′ (reverse). Real-time quantitative PCR was performed at 95°C
for 5 sec and 60°C for 30 sec for 40 cycles using SYBR Green PCR
Master Mix (Applied Biosystems) and Applied Biosystems 7500
Real-Time PCR System. Relative gene expression level was calculated
by using the 2−ΔΔCt method. After 40 cycles of PCR, a
melting curve analysis was performed to monitor PCR product purity
and then the amplicons were analyzed on a 1.0% agarose gel
containing ethidium bromide.
Western blot analysis
Six cultured cell lines in an exponentially growing
phase were washed thoroughly twice with PBS and then proteins of
cell lines, liver tissues, snap-frozen HCC and corresponding
paracarcinomatous tissues were extracted using Total Protein
Extraction kit (KeyGen, China). After centrifugation, protein
concentrations of lysate were quantified using BCA method. Then,
equal amounts of protein samples were loaded onto 10% sodium
dodecyl sulfate-polyacrylamide electrophoresis (SDS-PAGE) gel and
electrophoretically transferred onto polyvinylidene difluoride
(PVDF) membranes (0.45 μm, Millipore, MA, USA). Membranes
were blocked with 5% non-fat milk in Tris-buffered saline
containing 0.1% Tween-20 (TBST) for 1 h and then incubated with
primary antibody of rabbit anti-Mortalin antibody (ab53098) (Abcam,
Cambridge, MA, USA), rabbit anti-Vimentin antibody (ab92547)
(Abcam) and rabbit anti-β-actin antibody (ab133626) (Abcam) at 4°C
overnight. After 1.5 h of incubation in horseradish
peroxidase-labelled secondary antibodies (ZB-2308, ZSGB-BIO), the
blots were detected using ECL western blot kits (Pierce, Rockford,
IL, USA) according to the manufacturer’s instructions. The density
of the bands was quantified with an ImageJ Software (National
Institute of Health, USA). The expression of β-actin was used as an
internal control.
Histopathological and immunohistochemical
analyses
Immunohistochemisty was used to detect the
expression of Mortalin in 10 normal liver samples and 100 HCC tumor
and corresponding paracarcinomatous samples; Vimentin and CD34 were
only examined in HCC tumor tissues. Briefly, formalin-fixed and
paraffin-embedded tissues were cut into sections with a thickness
of 2 μm. Sections were stained with hematoxylin and eosin
(HE) for histological examination. Sections were dewaxed in xylene
and rehydrated in a graded series of alcohols. Thereafter, antigen
retrieval was performed by microwaving in sodium citrate buffer (10
mM, pH 6.0) for 15 min and endogenous peroxidase activity was
blocked with 3% hydrogen peroxide for 10 min. Subsequently, rabbit
anti-Mortalin antibody (ab53098) (Abcam), mouse anti-Vimentin
antibody (ZM-0260, ZSGB-BIO), mouse anti-CD34 (ZM-0046, ZSGB-BIO)
antibody were, respectively, incubated at 4°C overnight. Following
30 min of incubation in horse-radish peroxidase-conjugated
secondary antibody (PV-6000, ZSGB-BIO), sections were stained in 3,
3-diaminobenzidine tetrahydrochloride (DAB) (ZLI-9017, ZSGB-BIO)
solution under microscopic observation and then counterstained
lightly with hematoxylin. The positive controls were used with
sections of breast cancer and the negative controls were processed
with PBS instead of primary antibody.
Mortalin and Vimentin staining were assessed by a
semi-quantitative scoring system including the intensity of
staining and the proportion of stained cells (15). At low-power (×40) microscope,
staining intensity of tissue sections were scored (0, no staining;
1, weak staining appearing as light yellow; 2, moderate staining
appearing as yellowish-brown; 3, strong staining appearing as
brown); at high-power (×400) microscope, >5 fields in one
section were scrutinized and then the percentage of positive
stained cells was calculated (0, none; 1, <10%; 2, 10%–50%; 3,
>50%). The final score of each section was obtained: [(score for
staining intensity) x (score for percentage of positive cells)].
For category analysis, immunoreactivity of tumor cells was
distinguished between high (total score ≥4) and low (total score
<4). CD34 is an antigen present in the vascular endothelial
cells. We used antibodies against CD34 to stain vascular
endothelial cells and then calculated the microvessel density (MVD)
(16). The field of maximal CD34
expression was found in tumor tissues. Within the field, the area
of maximal angiogenesis was selected and microvessels were counted
on a ×200 magnification field. The immunohistochemical results were
scored by three pathologists who were blinded to clinical data.
Plasmid extraction and RNA
interference
The small hairpin RNAs (shRNA) (Genechem, China)
against Mortalin were designed by inserting oligos with a hairpin
loop into the GV115 vector. The target sequences of Mortalin shRNA
(NM_004134, CTGGAATGGCCTTAGTCAT) were synthesized by Shanghai
GeneChem Co. Bacteria containing the plasmid were inoculated on
solid culture medium with Amp resistance in the incubator. A
bacterial colony was picked and then inoculated in liquid culture
medium on a vibration shaker at 37°C (shaker speed 150 rpm)
overnight. Harvest overnight bacterial culture by centrifuging
12,000 g for 2 min. Plasmid was extracted using the Qiagen Plasmid
Mini kit (Qiagen, China) according to the manufacturer’s
instructions, then dissolved in a suitable volume of TE buffer.
MHCC97H cells at ∼3×105/ml were seeded in a 6-well plate
and placed in antibiotic-free DMEM containing 10% FBS. When the
cells confluence reached 90%, transient transfection of MHCC97H
cells was performed. A mixture containing 10 μl/well
Lipofectamine 2000 (Invitrogen), 10 μl/well plasmid and 480
μl/well OPTI-MEM were added to plates. After 6 h, OPTI-MEM
was replaced by DMEM containing 10% FBS.
Three groups were established in this experiment:
blank group (no interference), NC (negative control) group
(transfected with NC shRNA) and shRNA group (transfected with
Mortalin shRNA). Cells were harvested 24, 48, 72 and 96 h after
transfection and subjected to MTT assay, flow cytometry, qPCR and
western blot analyses. Simultaneously, a GFP (green fluorescent
protein) plasmid was used to determine transfection efficiency and
cell viability was determined by the MTT assay (MTT Cell
Proliferation and Cytotoxicity Assay kit) and flow cytometry
(Annexin V/PI apoptosis kit). All experiments were performed in
triplicate to confirm the reproducibility.
MTT assay
[3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide]
Twenty-four hours post-transfection cells
(2.0×104/ml, 200 μl/well) were cultured in
96-well plates with DMEM among three groups. MTT (0.5%) (Beyotime,
China) with a volume of 20 μl was added to each well for 4 h
and then DMEM was aspirated from wells as far as possible without
disturbing the cells and crystals on the plastic surface.
Subsequently, each well was added with a 200-μl dimethyl
sulfoxide (DMSO) to dissolve crystals and the plate agitated on a
plate shaker for 10 min. The optical density (OD) measurements were
carried out in an enzyme-linked immunosorbent assay (ELISA) reader
at the wavelength of 490 nm. The reader was calibrated to 0
absorbance using DMEM without cells.
Flow cytometry
Twenty-four hours post-transfection, cells
(2.0×105) were collected by centrifugation among three
groups and then suspended in 500 μl 1X binding buffer.
Subsequently, 5 μl Annexin V-FITC and 10 μl propidium
iodide (PI) (Annexin V/PI apoptosis kit, Multi Sciences) were added
to each sample. Samples were gently mixed and incubated at room
temperature for 5 min in the dark. The cells were analyzed on a
Canto-II™ reader (BD Biosciences) and data were analyzed with
FlowJo software (Tree Star, Ashland, OR, USA).
Statistical analysis
Statistical analyses were performed using the
statistical package SPSS 13.0 (SPSS Inc., Chicago, IL, USA). The
difference in Mortalin mRNA and protein expression level among six
cell lines and tissue samples were examined using Student’s t-test.
Fisher’s exact or χ2 tests were performed to analyze
correlations among different protein expressions or between protein
expressions and various clinicopathological parameters. P<0.05
was considered statistically significant.
Results
The expression level of Mortalin mRNA and
protein in HCC and normal liver cell lines
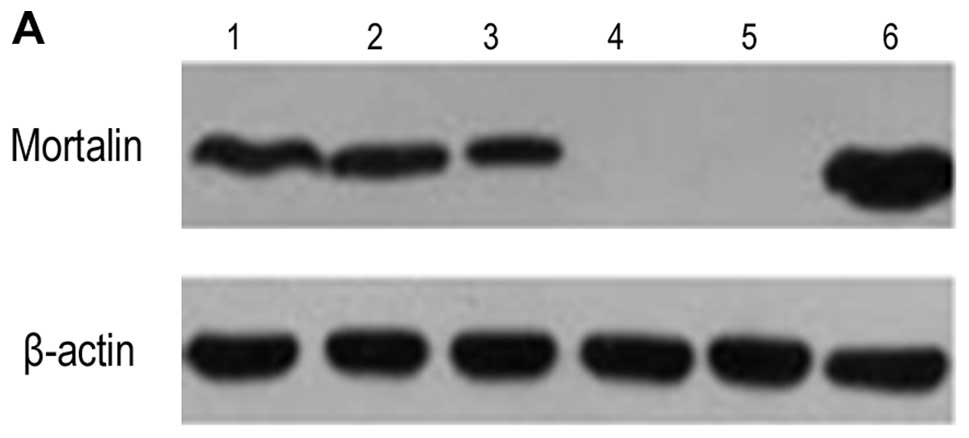
The expression of nodal mRNA was detected in all
cell lines. The expression of Mortalin protein was detected in
Hep3B, MHCC97H, HepG2 and HCCLM3, but not in MHCC97L and L02
(Fig. 1). The mean expression
level of Mortalin mRNA and protein are shown in Table II. It was found that among the
three human HCC cell lines with the same genetic background, the
expression level of Mortalin was the highest in HCCLM3, while it
was the lowest in MHCC97L (P<0.05).
| Table II.Relative quantity of Mortalin mRNA
and protein in six cell lines. |
Table II.
Relative quantity of Mortalin mRNA
and protein in six cell lines.
| Hep3B | MHCC97H | HepG2 | L02 | MHCC97L | HCCLM3 |
|---|
| mRNA | 0.45±0.08 | 1 | 0.14±0.06 | 0.04±0.01 | 0.05±0.02 | 2.38±0.27 |
| Protein | 0.72±0.11 | 1.21±0.13 | 0.35±0.07 | 0 | 0 | 2.14±0.29 |
The expression level of Mortalin mRNA and
protein in HCC tumor, paracarcinomatous and normal liver
tissues
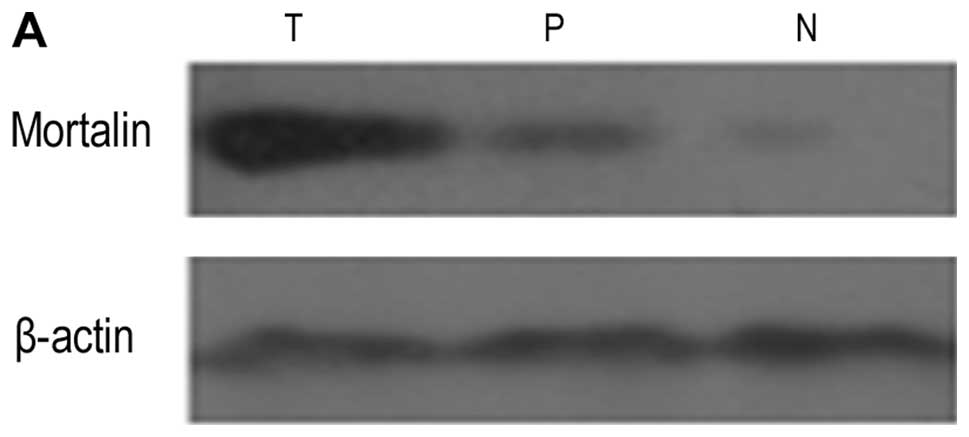
Mortalin was detected in 10 normal liver samples and
all 13 HCC tumors and corresponding paracarcinomatous samples. The
mean mRNA and protein level of Mortalin in HCC tumor,
paracarcinomatous and normal liver tissues were 1, 0.36±0.14,
0.18±0.07 and 2.58±0.21, 0.22±0.09, 0.05±0.02, respectively
(Fig. 2). The results showed that
the expression level of Mortalin in HCC tumor tissues was
significantly higher than that in paracarcinomatous and normal
tissues (P<0.05).
Immunohistochemical expression of
Mortalin in HCC tumor, paracarcinomatous and normal liver
tissues
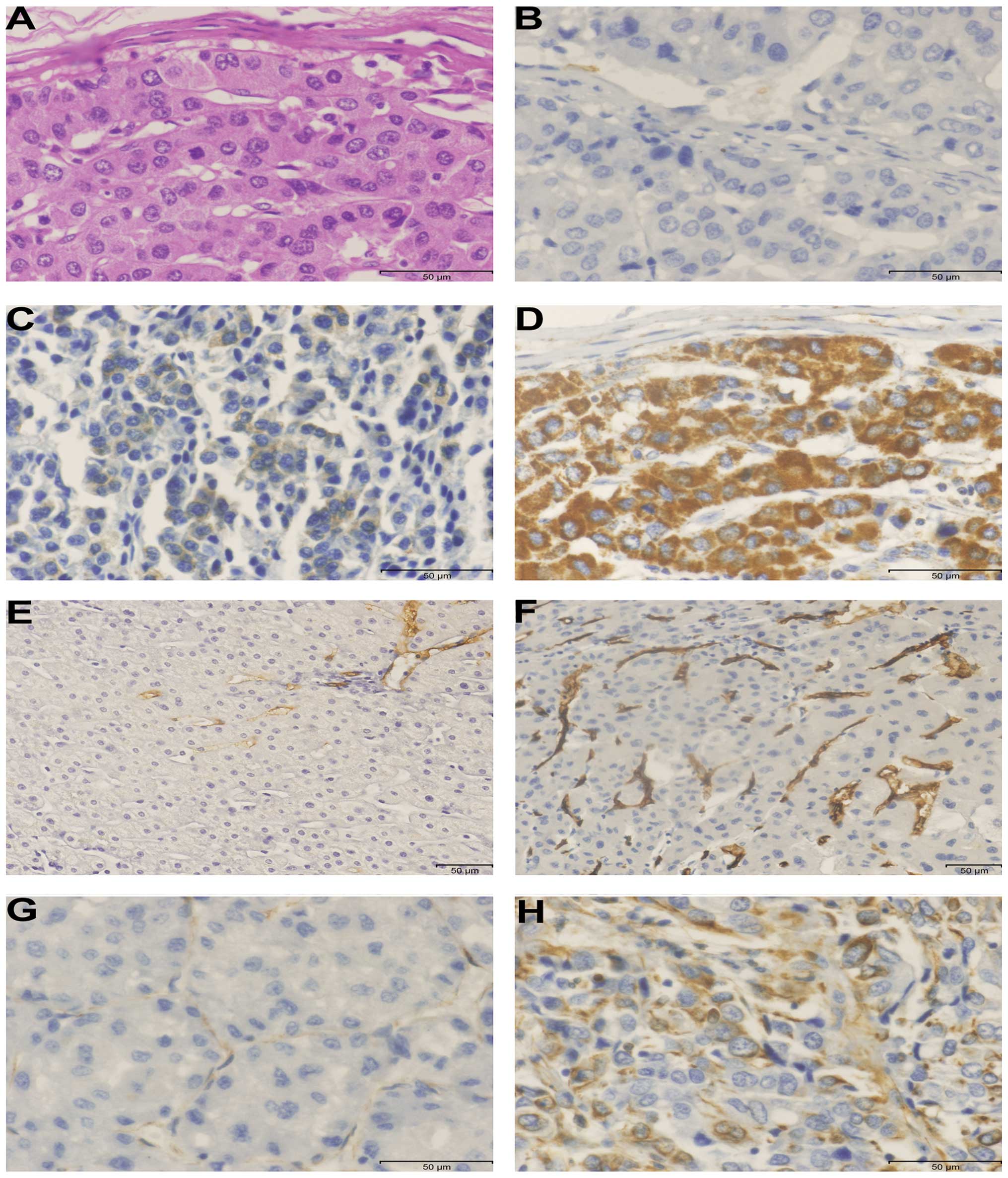
We found that positive expression of Mortalin was
mainly cytoplasmic, some tumor cells stained strongly, while others
exhibited slight or no staining at all (Fig. 3B–D). Mortalin expression was
detected in 77% (77/100) of HCC tumor tissues, which was
significantly higher than 19% (19/100) in the paracarcinomatous
tissues and 1% (1/10) in the normal liver tissues
(χ2=67.388, P<0.001; χ2=16.669,
P<0.001, respectively) (Table
III). Association between the expression of Mortalin and
clinicopathological characteristics was evaluated and was found
that the high expression of Mortalin was positively correlated with
poor Edmondson grade (χ2=4.272, P=0.039), advanced TNM
stages (χ2=8.188, P=0.004) and status of invasion and
metastasis (χ2=4.742, P=0.029), but there was no
significant correlation with other variables (Table I).
| Table III.Immunohistochemical expression of
Mortalin in HCC, paracarcinomatous tissues and normal liver
tissues. |
Table III.
Immunohistochemical expression of
Mortalin in HCC, paracarcinomatous tissues and normal liver
tissues.
| Liver tissue | Mortalin expression
| χ2 | P-value |
|---|
| Low | High |
|---|
| HCC tumor
tissue | 23 | 77 | | |
| Paracarcinomatous
tissue | 81 | 19 | 67.388 | <0.001 |
| Normal tissue | 9 | 1 | 16.669 | <0.001 |
Immunohistochemical expression of
Mortalin, Vimentin and MVD in HCC tumor tissues
Vimentin is an important hallmark of EMT; high
expression of Vimentin was observed mainly in the cytoplasmic
staining (Fig. 3G and H). Among
all patients with HCC, 52% (52/100) patients had high expression of
Vimentin. Spearman’s rank correlation test was used to analyze the
relation between Mortalin and Vimentin; a significant positive
correlation was found between the expression of Mortalin and
Vimentin (r=0.236, P=0.018) (Table
IV). Angiogenesis is an important process of growth and
metastasis in HCC; MVD was detected by using an antibody against
CD34 (Fig. 3E and F). MVD in HCC
tumor tissues ranged from 0 to 139/200 × field (median, 32/200 ×
field). We found that tumors with high expression of Mortalin had a
tendency to higher MVD than those with the low expression of
Mortalin (39.4±42.5 vs. 29.7±16.9, t=1.630, P=0.106).
| Table IV.Correlation between Mortalin and
Vimentin expression in 100 HCC patients with HCC. |
Table IV.
Correlation between Mortalin and
Vimentin expression in 100 HCC patients with HCC.
| Mortalin
expression | Vimentin expression
| r | P-value |
|---|
| Low | High |
|---|
| Low | 16 | 7 | 0.236 | 0.018 |
| High | 32 | 45 | | |
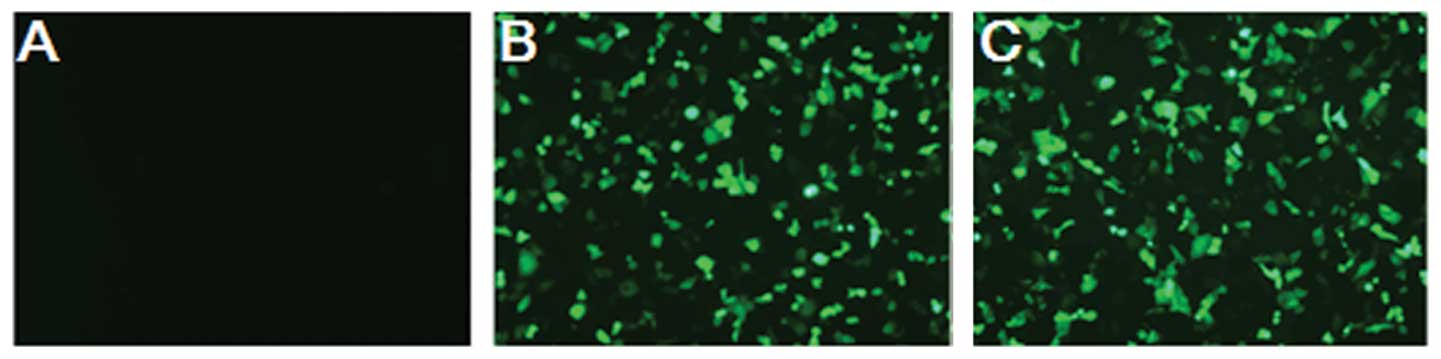
shRNA transfection in MHCC97H cells
GFP fluorescence showed that plasmid Mortalin shRNA
was successfully transfected into MHCC97H cells (Fig. 4). MTT and flow cytometry assays
were applied to determine cell viability 24 h after transfection
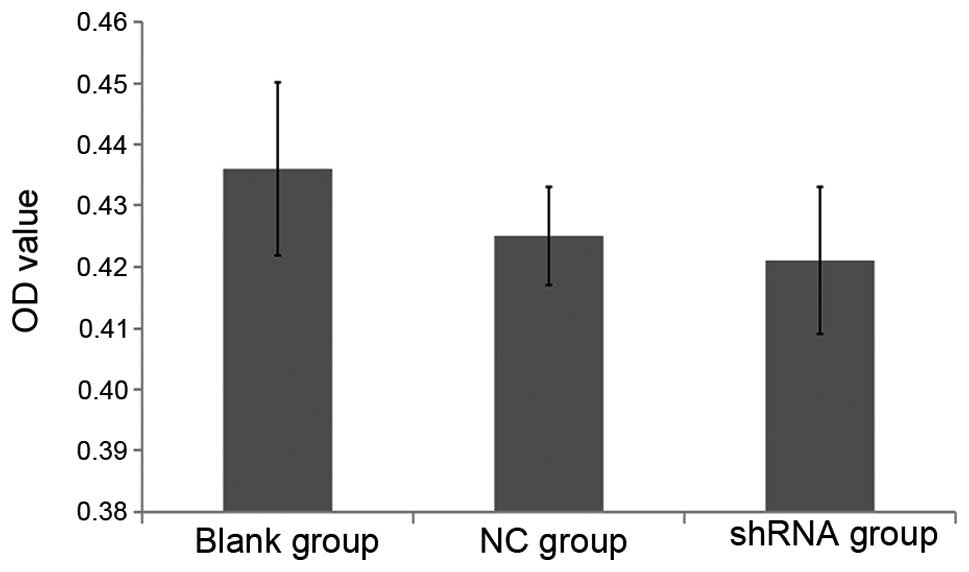
among the three groups. OD values were not significantly different
among the three groups (P>0.05) (Fig. 5). The result showed that only 2.5
and 3.5% cytotoxicity was present compared to the blank group.
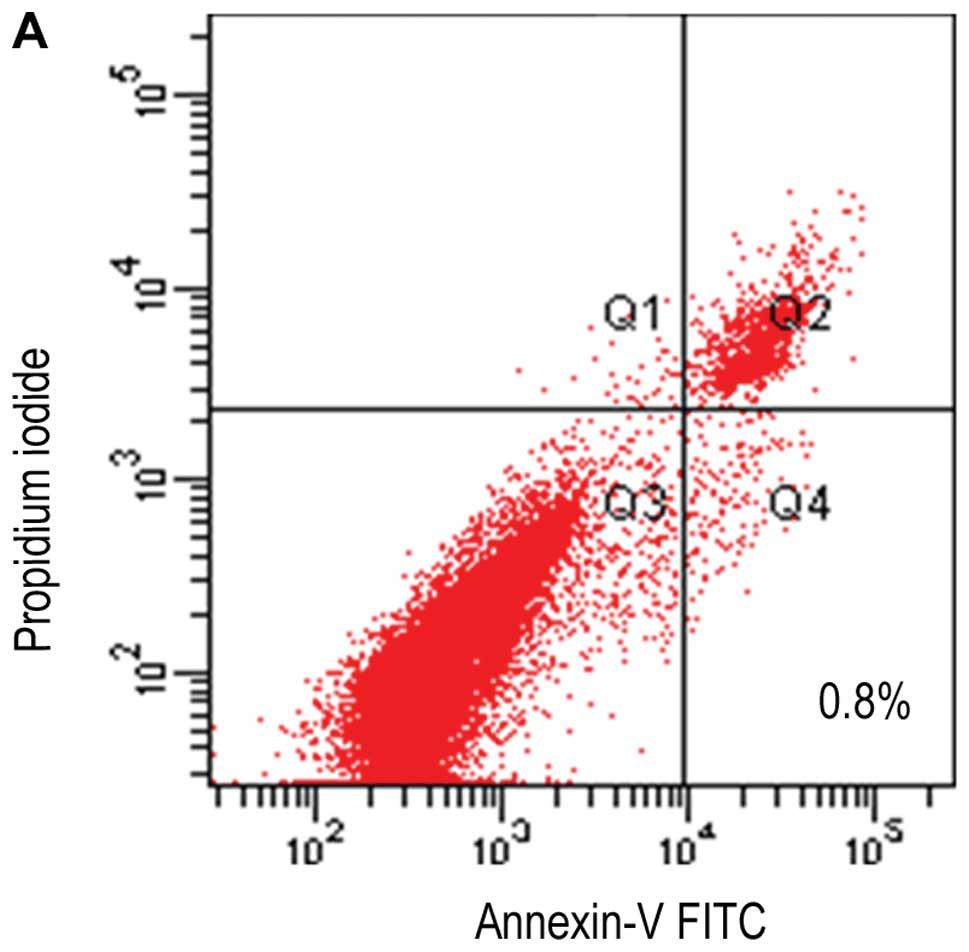
Similarly, flow cytometry analysis showed that early apoptosis
(Annexin V+/PI−) rates among the three groups
were 0.8, 4.5 and 9.2% (Fig. 6),
indicating that transfection did not cause severe cell damage. To
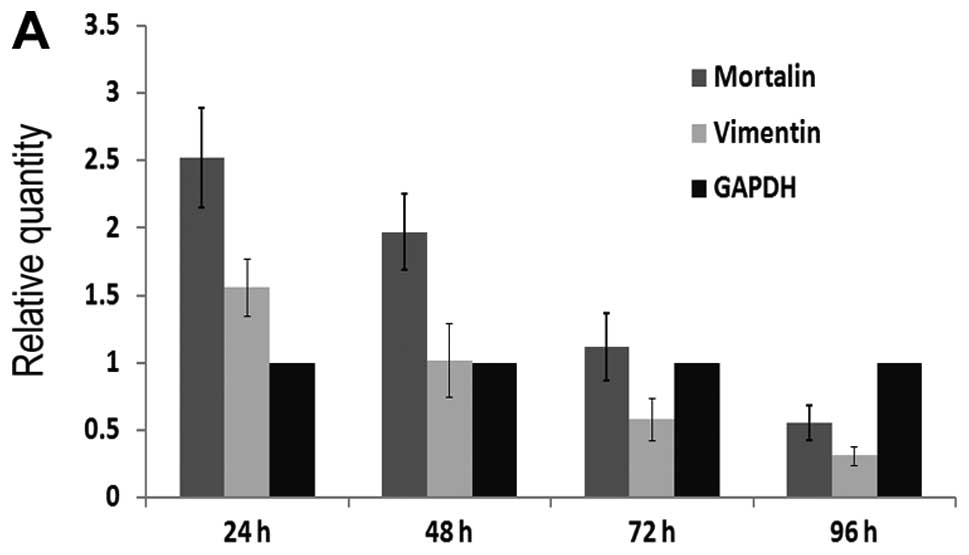
demonstrate the level of inhibition after transfection, qPCR assay
was performed. As shown in Fig.
7A, Mortalin mRNA level of cells were 2.52±0.37, 1.97±0.28,
1.12±0.25 and 0.55±0.13 in shRNA group after 24, 48, 72 and 96 h,
respectively. Simultaneously, the expression level of Mortalin mRNA
in blank group and NC group were 2.90±0.31, 2.83±0.22, 2.95±0.27,
2.90±0.23 and 2.88±0.41, 2.80±0.15, 2.93±0.30, 2.91±0.09,
respectively. The expression level of Mortalin mRNA in shRNA group
was significantly lower than that in blank group and NC group
(P<0.05, respectively). These results showed that Mortalin shRNA
was an effective sequence to suppress the target gene and the
capability of interference increased gradually upto 96 h after
transfection.
To find the relationship between Mortalin and
Vimentin, qPCR and western blot assays were applied. Vimentin mRNA
level of cells in shRNA group after 24, 48, 72 and 96 h were
1.56±0.21, 1.02±0.27, 0.58±0.16 and 0.31±0.07, respectively
(Fig. 7A). 24, 48, 72 and 96 h
after transfection, the expression level of Mortalin protein and
Vimentin protein in shRNA group were 2.09±0.37, 1.48±0.23,
0.73±0.11, 0.25±0.06 and 1.03±0.21, 0.54±0.14, 0.21±0.07,
0.11±0.02, respectively (Fig. 7B).
We found that the decreased expression of Mortalin was accompanied
by a reduction of Vimentin expression (Fig. 7C). This observation supported our
conclusion that inhibition of Mortalin expression could decrease
the expression of Vimentin and had a suppressive function in
epithelial-tomesenchymal transition.
Discussion
HCC invasion and metastasis are closely related to
adverse clinical outcome and shorter survival of cancer patients.
Although great efforts have been made to clarify molecular
mechanisms involved in HCC invasion and metastasis, the detailed
mechanisms for HCC malignancy and metastasis are still unknown. It
was reported that angiogenesis and EMT played important roles in
the occurrence and progression of HCC (16,17).
Mortalin, as a ‘stress protein’, was a highly conserved molecular
chaperone and abundant in the same tumors and played a key role in
cell cycle regulation, carcinogenesis and tumor progression
(7,18,19).
Simultaneously, Mortalin-p53 interaction resulted in inhibition of
transcriptional activation and control of centrosome duplication
functions (9,20). In addition, Lu et al
(21) found that inhibitor of
Mortalin was able to induce tumor cell apoptosis. Therefore,
Mortalin was suggested to serve as a therapeutic marker.
It was previously reported that Mortalin was
overexpressed in leukemia, brain tumors, colon carcinoma and tumor
cell lines (7,22). Yi et al (11) have demonstrated that overexpression
of Mortalin was closely associated with venous infiltration and
advanced tumor stages in HCC. These findings suggested that
overexpressed Mortalin could increase malignancy and aggressive
behavior and predicted early recurrence in HCC. Chen et al
(23) found that expression of
Mortalin was notably higher in the SMMC 7721 than in a normal liver
cell line. Lu et al (21)
showed that human HepG2 cells lacked mortalin-p53 interaction and
were resistant to apoptosis, but cell apoptosis was significantly
increased by Mortalin shRNA transfection. These findings showed the
potential for clinical application of chemotherapeutic drugs in HCC
treatment.
HCC is usually a hypervascular tumor and
upregulation of Vimentin is considered as the essential step of EMT
process. When a liver tumor was 1–2 mm in diameter, it could obtain
nutrients through diffusion without an extensive vasculature. If
tumor growth become much larger, angiogenesis must occur, also,
metastasis is required for angiogenesis (24). CD34 is considered to be a marker
for neovascularization, the expression of CD34 positive endothelial
cells played an important role in the process of angiogenesis in
HCC and metastasis. EMT was first discovered at key transition
steps during embryogenesis and was the critical event that mediated
tumor metastasis. Through activation of several specific
transcription factors, tumor cells invariably adopt a mesenchymal
phenotype to invade surrounding tissues and metastasize (4,25).
Vimentin has been recognized as a marker for EMT and its
overexpression has been strongly associated with metastatic
phenotype and poor prognosis.
To our knowledge, this was the first study to
investigate the relationship among Mortalin expression,
angiogenesis and EMT by immunohistochemical staining. We discovered
that Mortalin had a significant correlation with Vimentin, but the
high expression of Mortalin group did not have significantly higher
MVD than that in low expression of Mortalin group. This shows that
Mortalin could not promote angiogenesis, but could contribute to
the process of EMT. We also observed that the expression level of
Mortalin in HCCLM3 with the highest metastatic potential was
notably higher than in the other five cell lines with lower
metastatic potential. This suggested that Mortalin expression
correlated with the metastatic phenotype of HCC cells.
In three cell lines with the same genetic background
and various metastatic potentials, HCCLM3, with the highest
metastatic potential, exhibited the highest level of Mortalin and
MHCC97L, with the lowest metastatic potential, having the lowest
level of Mortalin. This showed that overexpression of Mortalin
could possess metastasis-inducing capabilities. We analyzed
expression of Mortalin and clinicopathological characteristics and
found Mortalin was associated with Edmondson grade, TNM stage and
tumor invasion and metastasis. By Mortalin shRNA transfection, we
also found that decreased expression of Mortalin was accompanied by
a reduction of Vimentin expression. Our conclusions are that low
expression of Mortalin was able to inhibit EMT and decrease tumor
progression and lose the metastasis-inducing capability.
Although the same studies have identified that HSP70
blocked TGF-β-induced EMT in HaCat cells (26) and HSP72 inhibited EMT by inhibiting
Smad3 activation in renal epithelial cells (27), the mechanisms by which Mortalin
(HSP75) promoted EMT and tumor metastasis are still unclear. To
solve these problems, further experiments are required.
In conclusion, our study suggests that
overexpression of Mortalin is correlated with metastatic phenotype
of HCC cells and can promote EMT, but cannot induce angiogenesis in
HCC. The decreased expression of Mortalin is accompanied by an
inhibition of EMT in the HCC cell line. Using shRNA transfection,
Mortalin knockdown may have potential for clinical application to
decrease tumor metastasis and recurrence after curative
surgery.
Acknowledgements
This study was supported by the
National Natural Science Foundation of China (No. 81272398), the
Science and Technological Fund of Anhui Province (No. 12010402112)
and the Anhui Provincial Natural Science Foundation of Institution
of Higher Education (No. KJ2011A171).
References
|
1.
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2.
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. Lancet. 379:1245–1255. 2012. View Article : Google Scholar
|
|
3.
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Christiansen JJ and Rajasekaran AK:
Reassessing epithelial to mesenchymal transition as a prerequisite
for carcinoma invasion and metastasis. Cancer Res. 66:8319–8326.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Tanaka S and Arii S: Molecular targeted
therapies in hepatocellular carcinoma. Semin Oncol. 39:486–492.
2012. View Article : Google Scholar
|
|
6.
|
Wadhwa R, Taira K and Kaul SC: An Hsp70
family chaperone, mortalin/mthsp70/PBP74/Grp75: what, when, and
where? Cell Stress Chaperones. 7:309–316. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Wadhwa R, Takano S, Kaur K, et al:
Upregulation of mortalin/ mthsp70/Grp75 contributes to human
carcinogenesis. Int J Cancer. 118:2973–2980. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Mizukoshi E, Suzuki M, Misono T, et al:
Cell-cycle dependent tyrosine phosphorylation on mortalin regulates
its interaction with fibroblast growth factor-1. Biochem Biophys
Res Commun. 280:1203–1209. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Wadhwa R, Takano S, Robert M, et al:
Inactivation of tumor suppressor p53 by mot-2, a hsp70 family
member. J Biol Chem. 273:29586–29591. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Wadhwa R, Yaguchi T, Hasan MK, Taira K and
Kaul SC: Mortalin-MPD (mevalonate pyrophosphate decarboxylase)
interactions and their role in control of cellular proliferation.
Biochem Biophys Res Commun. 302:735–742. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Yi X, Luk JM, Lee NP, et al: Association
of mortalin (HSPA9) with liver cancer metastasis and prediction for
early tumor recurrence. Mol Cell Proteomics. 7:315–325. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Li Y, Tang Y, Ye L, et al: Establishment
of a hepatocellular carcinoma cell line with unique metastatic
characteristics through in vivo selection and screening for
metastasis-related genes through cDNA microarray. J Cancer Res Clin
Oncol. 129:43–51. 2003.
|
|
13.
|
Li Y, Tang ZY, Ye SL, et al: Establishment
of cell clones with different metastatic potential from the
metastatic hepatocellular carcinoma cell line MHCC97. World J
Gastroenterol. 7:630–636. 2001.PubMed/NCBI
|
|
14.
|
Edmondson HA and Steiner PE: Primary
carcinoma of the liver: a study of 100 cases among 48,900
necropsies. Cancer. 7:462–503. 1954. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Chen MH, Yip GW, Tse GM, et al: Expression
of basal keratins and vimentin in breast cancers of young women
correlates with adverse pathologic parameters. Mod Pathol.
21:1183–1191. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Weidner N, Semple JP, Welch WR and Folkman
J: Tumor angiogenesis and metastasis - correlation in invasive
breast carcinoma. N Engl J Med. 324:1–8. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Hartl FU: Molecular chaperones in cellular
protein folding. Nature. 381:571–579. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Calderwood SK, Khaleque MA, Sawyer DB and
Ciocca DR: Heat shock proteins in cancer: chaperones of
tumorigenesis. Trends Biochem Sci. 31:164–172. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Ma Z, Izumi H, Kanai M, Kabuyama Y, Ahn NG
and Fukasawa K: Mortalin controls centrosome duplication via
modulating centrosomal localization of p53. Oncogene. 25:5377–5390.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Lu WJ, Lee NP, Kaul SC, et al:
Mortalin-p53 interaction in cancer cells is stress dependent and
constitutes a selective target for cancer therapy. Cell Death
Differ. 18:1046–1056. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Dundas SR, Lawrie LC, Rooney PH and Murray
GI: Mortalin is over-expressed by colorectal adenocarcinomas and
correlates with poor survival. J Pathol. 205:74–81. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Chen X, Xu B, Li H, et al: Expression of
mortalin detected in human liver cancer by tissue microarrays. Anat
Rec. 294:1344–1351. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Kimura H, Nakajima T, Kagawa K, et al:
Angiogenesis in hepatocellular carcinoma as evaluated by CD34
immunohistochemistry. Liver. 18:14–19. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Gupta GP and Massague J: Cancer
metastasis: building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Zhou Y, Mao H, Li S, et al: HSP72 inhibits
Smad3 activation and nuclear translocation in renal
epithelial-to-mesenchymal transition. J Am Soc Nephrol. 21:598–609.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Li Y, Kang X and Wang Q: HSP70 decreases
receptor-dependent phosphorylation of Smad2 and blocks
TGF-beta-induced epithelial-mesenchymal transition. J Genet
Genomics. 38:111–116. 2011. View Article : Google Scholar : PubMed/NCBI
|