Introduction
Endometrial cancer is the third most common
gynecologic cancer in Japan, and its morbidity and mortality have
dramatically increased in the past 30 years (1). The majority of patients with early
stage endometrial cancer are cured through surgery, while the
patients with advanced stage or recurrent lesions are treated by
chemotherapy. Paclitaxel and/or cisplatin-based chemothetrapies
have been applied to these patients, but with limited efficacy, for
which new molecular target therapies are urgently needed. Recent
studies have demonstrated the potential of molecular target therapy
against cancer stem cells (CSCs) (2).
The cells with CSC-like properties has been
demonstrated as tumor-initiating cells (TICs) in a variety of solid
tumors including breast cancer (3), brain tumors (4,5),
prostate cancer (6,7), lung cancer (8), pancreatic cancer (9), colorectal cancers (10,11)
and melanoma (12). CD133, a
5-transmembrane glycoprotein with a molecular weight of 117 kDa,
has been widely used to isolate TICs and is now considered to be a
potential marker of TICs in a variety of tumor types.
A previous study (13), together with our previous report
(14), showed that CD133 is a
potential marker of CSCs in endometrial cancer cells. Sorted
CD133+ cells had elevated levels of expression of
self-renewal genes, such as Nanog and BMI, compared to
CD133− cells (14).
CD133+ cells were able to generate both
CD133+ and CD133− cells, exhibiting
self-renewal capacity, while CD133− cells could not.
Furthermore, CD133+ cells showed increased proliferative
potential in vitro and tumorigenicity in vivo, and
showed apparent resistance to cytotoxicity from chemotherapeutic
agents. Immunohistochemical analysis of endometrial cancer
specimens revealed that overall survival was worse for tumors with
high CD133 expression than low CD133 expression (14). These studies have raised several
questions: why are CD133+ cells aggressive, leading to
the worse prognosis? What are the signaling pathways or molecules
causing the effect? Such information may support the establishment
of molecular target therapy to CSCs in endometrial cancer.
To answer these questions, we have sought to
characterize CD133+ endometrial cancer cells using
microarray analyses to identify genes involved in their CSC-like
features. Our study clearly demonstrates that an increased
chemoresistance and tumorigenic potentials of CD133+
cells are at least partly attributed to an enriched SP fraction as
well as increased MMP-1 expression.
Materials and methods
Cell culture
The human endometrial cancer cell lines, Ishikawa
and MFE280, were cultured in Dulbecco’s modified Eagle’s medium
(DMEM, Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10%
heat-inactivated fetal bovine serum (FBS), streptomycin (100
μg/ml), and penicillin (100 IU/ml) in the presence of 5%
CO2.
Flow cytometry and cell sorting
Cells were incubated in phosphate-buffered saline
(PBS) containing 0.5% bovine serum albumin (BSA) and 2 mM EDTA with
phycoerythrin (PE)-conjugated CD133/2 (clone 293C3) antibodies
(Miltenyi Biotec, Auburn, CA, USA). Mouse IgG2b-PE (Miltenyi
Biotec) was used as the isotype control antibody. To identify and
isolate SP cells, cells were stained with Hoechst 33342
(Sigma-Aldrich), either alone or in combination with 100 μM
verapamil (Sigma-Aldrich). For flow cytometry and cell sorting,
samples were analyzed using the JSAN desktop cell sorter and AppSan
software (Bay Bioscience Co. Ltd., Kobe, Japan).
Cell proliferation assay
Cell proliferation was determined using the WST-1
reagent (Roche Diagnostics, Tokyo, Japan). Briefly,
2×103 cells were seeded in 96-well plates and incubated
in normal medium conditions at 37°C. On designated days, WST-1
reagent (10 μl) was added to each well, and the cells
further incubated for 2 h at 37°C. Absorbance was measured using a
microplate reader at test and reference wavelengths of 450 and 655
nm, respectively.
Soft agar colony formation assay
Diluted single cells (5×104) were seeded
onto 60-mm dishes containing 0.33% soft agar in DMEM supplemented
with 10% heat-inactivated FBS on top of 0.5% base agar in DMEM
supplemented with 10% heat-inactivated FBS. Colonies with diameters
larger than 0.25 mm after 14 days incubation were counted.
Chemosensitivity assay
Cells (1×104) were seeded in 24-well
plates, incubated for 24 h, then treated with designated
concentrations of paclitaxel (provided by Bristol Pharmaceuticals,
Tokyo, Japan). After incubation for 48 h, the cells were counted
using a hemacytometer with trypan blue staining.
RNA analysis
Total RNA was isolated from the cells using the
RNeasy mini kit (Qiagen, Valencia, CA, USA) according to the
manufacturer’s protocol. Complementary DNA was synthesized from 2
μg of RNA using the Omniscript RT kit (Qiagen) with random
primers and amplified together with Taq polymerase (Nippon Gene,
Tokyo, Japan) for the amplification of ABCG2, MT1-MMP
and glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Primers and
PCR conditions are listed in Table
I. Real-time PCR (quantitative PCR; qPCR) was performed using a
LightCycler and a SYBR-Green system (Applied Biosystems, Foster
City, CA, USA). Microarray analyses were performed using 3D-Gene
Human Oligo chip 24k (Toray, Tokyo, Japan).
 | Table I.PCR primer and condition details. |
Table I.
PCR primer and condition details.
| Gene | Primer | Annealing
temperature | Cycle no. |
|---|
| ABCG2 | F:
GTTCTCTTCTTCCTGACGACCA
R: CCACACTCTGACCTGCTGCTA | 60 | 30 |
| MT1-MMP | F:
TCGGCCAAAGCAGCAGCTTC
R: CTTCATGGTGTCTGCATCAGC | 59 | 35 |
| GAPDH | F:
CTCAGACACCATGGGGAAGGTGA
R: ATGATCTTGAGGCTGTTGTCATA | 60 | 28 |
Western blot analysis
Whole cell extracts were prepared using 1X lysis
buffer (Cell Signaling, Danvers, MA, USA) and concentrations were
determined using the Bradford protein assay (Life Science,
Hercules, CA, USA). SDS-PAGE and western blot analysis were
performed as described previously, using 50–100 μg of
protein (15,16). MT1-MMP antibody (clone 114-6G6,
Fuji Chemical Industries Ltd, Takaoka, Japan) was used at 1:25,
ABCG2 (ab3380, Abcam) and GAPDH (Abcam) at 1:10,000.
Knockdown study of MT1-MMP
Cells were seeded and transfected with 30 nM of
negative control small interfering RNA (siRNA) or human MT1-MMP
siRNA oligonucleotides (Applied Biosystems) using Lipofectamine
2000 Transfection Reagent (Invitrogen) according to the
manufacturer’s protocol.
Invasion assay
The invasive ability of CD133+ or
CD133− cells was assayed in vitro using a Biocoat
Matrigel Invasion Chamber (Becton-Dickinson Biosciences, Bedford,
MA, USA), as described previously (17). Cells were suspended in the upper
wells of Matrigel chambers, in DMEM containing 0.1% BSA. After a
22-h incubation, cells on the upper surface of the membrane were
removed by wiping with cotton swabs, and cells that had migrated
through the membrane to the lower surface were fixed with methanol
and stained with Mayer’s Hematoxylin and eosin. The cells on the
lower surface of the membrane were counted microscopically to
obtain the invasion index. Chemotaxis assays were performed in the
same manner, except that the filters were not coated with Matrigel,
and the number of cells on the lower surface of the membrane was
considered the migration index. The invasive ability of cells was
described as the ratio of the invasion index to the migration
index.
Gelatin zymography
The supernatants of cells were subjected to gelatin
zymography with Gelatin zymography kit (Primary Cell, Sapporo,
Japan) according to the manufacturer’s directions.
Statistical analysis
Statistical analysis was carried out using the
statistical package StatView version 5.0 (Abacus Concepts,
Berkeley, CA, USA). We used the Student’s t-test for in
vitro experiments. A p-value of <0.05 was considered to
indicate statistical significance.
Results
SP cells are enriched in CD133-expressing
Ishikawa cells
In our previous study (14), we examined the frequency of
CD133+ cells in 6 endometrial cancer cell lines, among
which Ishikawa and MFE280 cells exhibited significant levels of
CD133 expression detectable by FACS analysis. Furthermore, only
Ishikawa cells had distinct side population fraction, another
hallmark of CSC. Thus, in the present study, we mainly used
Ishikawa cells for the subsequent analyses. To characterize
CD133+ cells in endometrial cancer, we first performed
cDNA microarray analyses with Ishikawa cells. A total of 440 genes
were found to be overexpressed in CD133+ cells at least
2-fold compared with CD133− cells: genes of ABC
transporters, cytokines, growth factors and invasion molecules were
included in the total (Table II).
In contrast, a total of 96 genes were downregulated in
CD133+ cells at least 2-fold compared with
CD133− cells (Table
III). Among these genes, we paid special attention to the
multi-drug resistance gene ABCG2, because of the previous
findings that CD133+ endometrial cancer cells are more
resistant to chemotherapeutic agents such as paclitaxel and
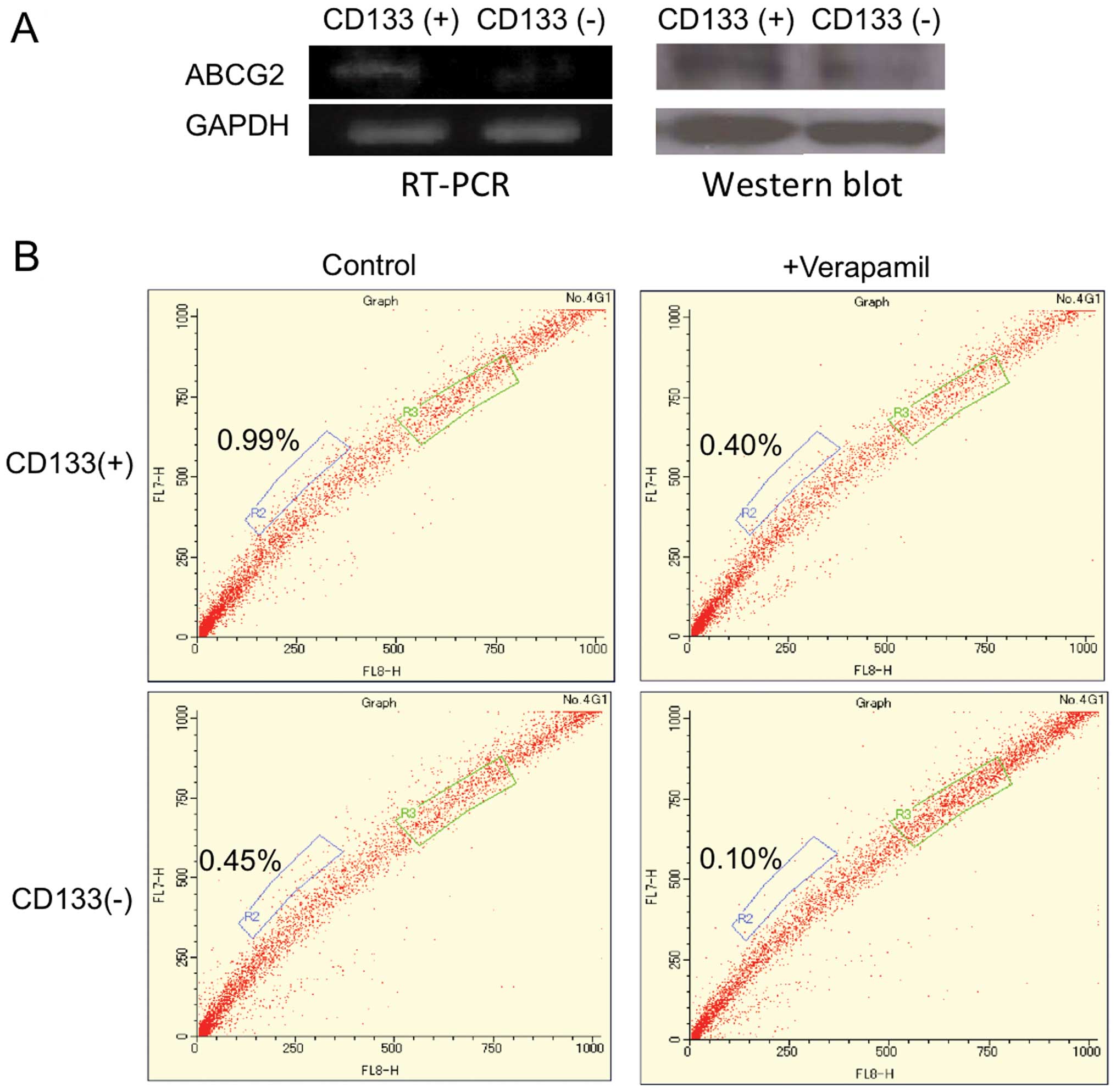
cisplatin than CD133− cells (14). Increased expression of ABCG2 in
CD133+ Ishikawa cells was confirmed by RT-PCR and
western blot analyses (Fig.
1A).
| Table II.The genes of higher expression in
CD133+ Ishikawa cells than CD133− cells. |
Table II.
The genes of higher expression in
CD133+ Ishikawa cells than CD133− cells.
| ARRDC2 | ZNF334 | GJB6 | TREX1 | MMP14(MT1-MMP) | STARD8 | Q86WM5_HUMAN | LHX1 | SLC26A3 | IGX1_HUMAN |
| ZNF137 | PLD2 | NMD3A_HUMAN | Q96DP9_HUMAN | TCL1A | CDA | HBXAP | FAM49A | DIAPH2 | DDHD1 |
| OR9G4 | CXCL13 | NKX2-4 | MARCO | GBG1_HUMAN | SENP7 | TMEM40 | ZNF584 | PCDH12 | C13orf23 |
| Q7Z2R7_HUMAN | RRAD | GSH1 | DPT | C4orf6 | Q8N134_HUMAN | GIMA5_HUMAN | CAPN3 | ABCG2 | TRPV6 |
| LAF4 | OPN1SW | C21orf63 | CBLN1 | NUDT10 | Q86VH3_HUMAN | C1QTNF3 | IER3 | GPSM1 | FSIP1 |
| GLS | ATOH1 | C20orf144 | KR108_HUMAN | Q8NH32_HUMAN | OR6Y1 | CYP3A5 | NTF3 | PCYT1B | FRMD3 |
| Q86W74_HUMAN | VSIG4 | ZC3HAV1 | DPP10 | TLL2 | C21orf121 | NOSTRIN | PITRM1 | RBM10 | P2RY1 |
| FAM53B | SLC14A1 | Q9NXW5_HUMAN | HPR | HMX2 | LRRK1 | C20orf85 | Q9P1C0_HUMAN | ADM2 | VGLL1 |
| GAL | PSG10 | Q9H357_HUMAN | SCRG1_HUMAN | Q8NGE6_HUMAN | CRSP2 | C2orf13 | EFHB | TNFRSF8 | MSH4 |
| VISL1_HUMAN | SLC27A4 | ENK13_HUMAN | Q8IY46_HUMAN | ZNF346 | SEPT4_HUMAN | Q9H8T6_HUMAN | PPY2 | BMP6 | ACOX3 |
| PPP1R14D | XDH | RALGPS2 | DEPDC1 | ALKBH | SH2D3C | SIA7B_HUMAN | ALS2CR8 | Q7Z470_HUMAN | Q8NAH5_HUMAN |
| PKN2 | ZNF195 | PAQR3_HUMAN | RAPGEF1 | GPATC4 | TMSL6 | C10orf81 | KIAA0258 | WBSCR23 | Q9NRZ3_HUMAN |
| Q9P135_HUMAN | ZNF541 | Q70YC7_HUMAN | KLF8 | SCEL | Q8N2X2_HUMAN | LMOD1 | Q9NSJ0_HUMAN | RAP1GDS1 | NF2 |
| KBTB6_HUMAN | HOXC13 | IGJ | DLX3 | PRSS23 | CE290_HUMAN | LPL | ZFX | HIP14_HUMAN | KRT23 |
| GPR19 | Q8NHA3_HUMAN | OR8D1 | ASCC3L1 | PHACTR3 | FSTL3 | SP3 | ALDH1A1 | SPARC | FHL2 |
| NT5E | EDNRA | HAMP | SLC22A16 | TMPRSS3 | DBT | C9orf60 | ASB6 | CLDN9 | HSPB3 |
| CTSG | IGHG3 | GPR87_HUMAN | CPA1 | PRKY | TUBB4Q | DSCR1L2 | ELN | TMCC3 | SLC19A2 |
| SNX9 | CEACAM5 | TEAD1 | ACCN5 | PDZK10 | CD24 | CD3G | ABCG8 | DNAJA4 | PTK2B |
| PCDHB10 | LOH11CR2A | FBXO15 | DPYS | NKPD1 | ABCG4 | TGM4 | ZIM3 | RALGPS2 | CSNK1G3 |
| FIGN | TNFRSF14 | BPESC1 | Q86XT6_HUMAN | UNC5A_HUMAN | PIP5KL1 | TIGD7 | RASGRP2 | Q8N9Q3_HUMAN | FSD1 |
| PLAG1 | CASP8 | DMRT1 | Q8NGP1_HUMAN | FXYD6 | BIRC8 | THEG | Q5VSL2_HUMAN | LEPREL1 | MMP10 |
| CRABP1 | IGFBP5 | PTHLH | Q8ND77_HUMAN | RTN1 | TFAP4 | KRTAP2-4 | FSHPRH1 | Q8N3F2_HUMAN | LRRK2 |
| TMED6 | CAV1 | Q8NAM0_HUMAN | ABCC9 | SYT3 | MLSTD1 | ZNF214 | TYB4_HUMAN | Q9P1G6_HUMAN | ITM2A |
| CABYR | KYNU | ABI3BP | ZFYVE9 | ITGB3 | APXL | RGS7 | ALOX15B | CHRAC1 | RGS16 |
| Q9Y4N5_HUMAN | TDGF1 | HAL | RGS3 | LYZL1 | LAS1L | SPG7 | LARS | ZNF93 | GRINL1A |
| Q8NEE2_HUMAN | KIF21B | GPR120 | IFR28_HUMAN | PERQ1 | GPR119 | GPR26 | SALL1 | ATXN7L1 | ADHFE1 |
| Q8N896_HUMAN | WDR9_HUMAN | SUHW4 | BEX1 | ZFP67 | OR5W2 | SLC16A5 | CLDN1 | C19orf18 | |
| XP_376267.2 | C9orf102 | HCP5 | TFF3 | OSGEPL1 | C10orf59 | FUBP3_HUMAN | IL17D | Q9NWZ4_HUMAN | |
| CD37 | RPS6KB1 | TRAM2 | PTPRE | ZNF311 | DDAH1 | Q96JG6_HUMAN | MYCNOS | INHBB | |
| Q9P143_HUMAN | ZDHHC8 | UPK1B | HS3ST3A1 | CXCL1 | ACOXL | THAP2_HUMAN | Q14560_HUMAN | IL20RA | |
| SEMA4F | COPA | OXCT2 | ABCC8 | Q96FU4_HUMAN | SPATA13 | Q9NSH8_HUMAN | Q8NEF7_HUMAN | GPRC5C | |
| ESR2 | SLMAP | ASB4 | F10 | Q8N4W5_HUMAN | SERPINB8 | C9orf90 | GK | TMEPAI | |
| OLFM1 | DIO1 | RASGEF1C | HD | DUSP16 | ATP2B4 | NID2 | CA9 | CCL15 | |
| FPRL1 | O14634_HUMAN | CM35H_HUMAN | CPEB2 | FUT10 | THSD1 | CHRNA2 | CBLN3_HUMAN | CTAG2 | |
| TBA3_HUMAN | ARHGAP25 | Q86V40_HUMAN | APEX2 | ZNF12 | ACTL7B | CLDN16 | NRGN | RHBDF1 | |
| PSMF1 | NCF2 | SYT5 | ELAC1 | ACOX2 | EBF2 | HGF | CALR3 | Q9NPS2_HUMAN | |
| CKLF | S100A12 | PLXNA4 | SIRT6_HUMAN | PPP1R1A | GK2 | ALDH1A3 | LENEP | ZNF588 | |
| TRIM58 | HPCL4_HUMAN | PBX1 | TEX15 | FSCN1 | SAMD1 | Q8N9M7_HUMAN | Q14946_HUMAN | CCL24 | |
| UTRN | TRIM6 | RAB14_HUMAN | C20orf54 | RPP30 | FGFR1 | FGFBP1 | ZNF367 | ARPM2_HUMAN | |
| SLC6A9 | Q9H8Q9_HUMAN | FZD4 | Q9HAU7_HUMAN | SERPINI2 | PIK3CG | MARK4 | Q6UXT6_HUMAN | NXN | |
| OR51E1 | ZNF623 | ALPK2 | DSG1 | PTGES | NRAP | Q9BT82_HUMAN | CTAGE6 | ZNF44 | |
| Q30181_HUMAN | C20orf96 | GABRG2 | Q8N3H6_HUMAN | CST5 | KRT17 | RERG | KCNQ3 | FBXW8_HUMAN | |
| CCDC11 | Q5VZR3_HUMAN | SPACA3 | CA6 | HIST1H1B | ADPRHL1 | IVNS1ABP | GPR126 | SCGB1A1 | |
| PCDH15 | AKR1B10 | ANKRD2 | TAGLN | AHRR | SLC22A17 | TNFRSF19L | TPM1 | NKX3-1 | |
| CEL | CMKOR1 | Q86SX0_HUMAN | WWOX | NALP2 | TLX3 | CSN1S1 | Q5TAX4_HUMAN | PROM1 | |
| OR7E5P | SOCS6 | C1S | RCN3 | C10orf63 | RTDR1 | CRLF1 | EIF2C3 | Q9NSC1_HUMAN | |
| Table III.The genes of lower expression in
CD133+ Ishikawa cells than CD133− cells. |
Table III.
The genes of lower expression in
CD133+ Ishikawa cells than CD133− cells.
| DMPK | RBP4 | PRSS1 | ZNF261 |
| Q6ZQS7_HUMAN | OR56B4 | USH1C | SEMG1 |
| GRIN2C | C20orf19 | Q86XE0_HUMAN | B3GALT3 |
| GPR54 | MAL | LHFPL2 | ARSA |
| KLF1 | ZNF322A | RHEBL1 | KIAA1683 |
| CCNA1 | ANKRD24 | TRIM48 | Q5QPC4_HUMAN |
| MLL3 | DNASE1 | MBD3L2 | O75372_HUMAN |
| Q8NI68_HUMAN | ZNF228 | TESK2 | CLCN4 |
| IGF2 | RL41_HUMAN | LPHN1 | DCHS1 |
| IGF1 | UTF1 | TNIP1 | POLL |
| BIRC4 | TLE2 | CSF3R | PAX5 |
| ITGAL | HIST1H2AB | MYO18A | ACRC |
| C20orf23 | Q8N9V4_HUMAN | GRAP | TPK1 |
| RFPL1 | Q96NA9_HUMAN | KCNC4 | MSN |
| Q6PJR0_HUMAN | ZNF431 | POMC | RFPL2 |
| RIMS2 | C6orf148 | LRRIQ1 | Q8N9H1_HUMAN |
| FEV | Q9NRE5_HUMAN | NXPH4 | DCP1B |
| C21orf81 | FGD1 | PEX6 | HAGHL |
| RHD | RHOI_HUMAN | BCL2L13 | RCOR2 |
| ATP1A2 | POLR1A | TMEFF1 | Q96MC9_HUMAN |
| ALPPL2 | TRIM43 | VPREB3 | MSMB |
| Q9HCN2_HUMAN | Q9NYD4_HUMAN | COX1_HUMAN | OR11H4 |
| RASD1_HUMAN | ZNF423 | Q8IW70_HUMAN | SOS2 |
| NTRI_HUMAN | OCRL | C14orf49 | CNTNAP4 |
Based on the microarray analyses, we were interested
in the relationship between CD133+ cells and SP cells,
since the latter specialized populations are known to highly
express ABCG2. Flow cytometry was used to examine the SP fraction
in CD133+ or CD133− Ishikawa cells. The ratio
of the SP cells in CD133+ cells was calculated as 0.69%
(±0.14%) based on the control with verapamil, whereas it was 0.49%
(±0.17%) in CD133− cells. These findings indicate that
there are more SP cells in CD133+ cells (Fig. 1B).
A recent report indicated that SP cells in
endometrial cancer are potential CSCs (18). Therefore, we confirmed such
potential of Ishikawa-SP cells by chemosensitivity or
colony-formation assay. Ishikawa-SP or non-SP cells were treated
with paclitaxel at 2 or 10 nM for 48 h and the cells counted. As
shown in Fig. 1C, SP cells are
more resistant than non-SP cells to paclitaxel at 10 nM (Fig. 1C). A colony-formation assay was
performed, in which Ishikawa-SP or non-SP cells were seeded onto
soft agar and colonies larger than 0.25 mm in diameter after
incubation for 14 days were counted. The SP cells showed
significantly greater colony-forming ability than non-SP cells
(Fig. 1D), consistent with a
previous report (18).
CD133-expressing SP cells have increased
proliferative and anchorage-independent growth
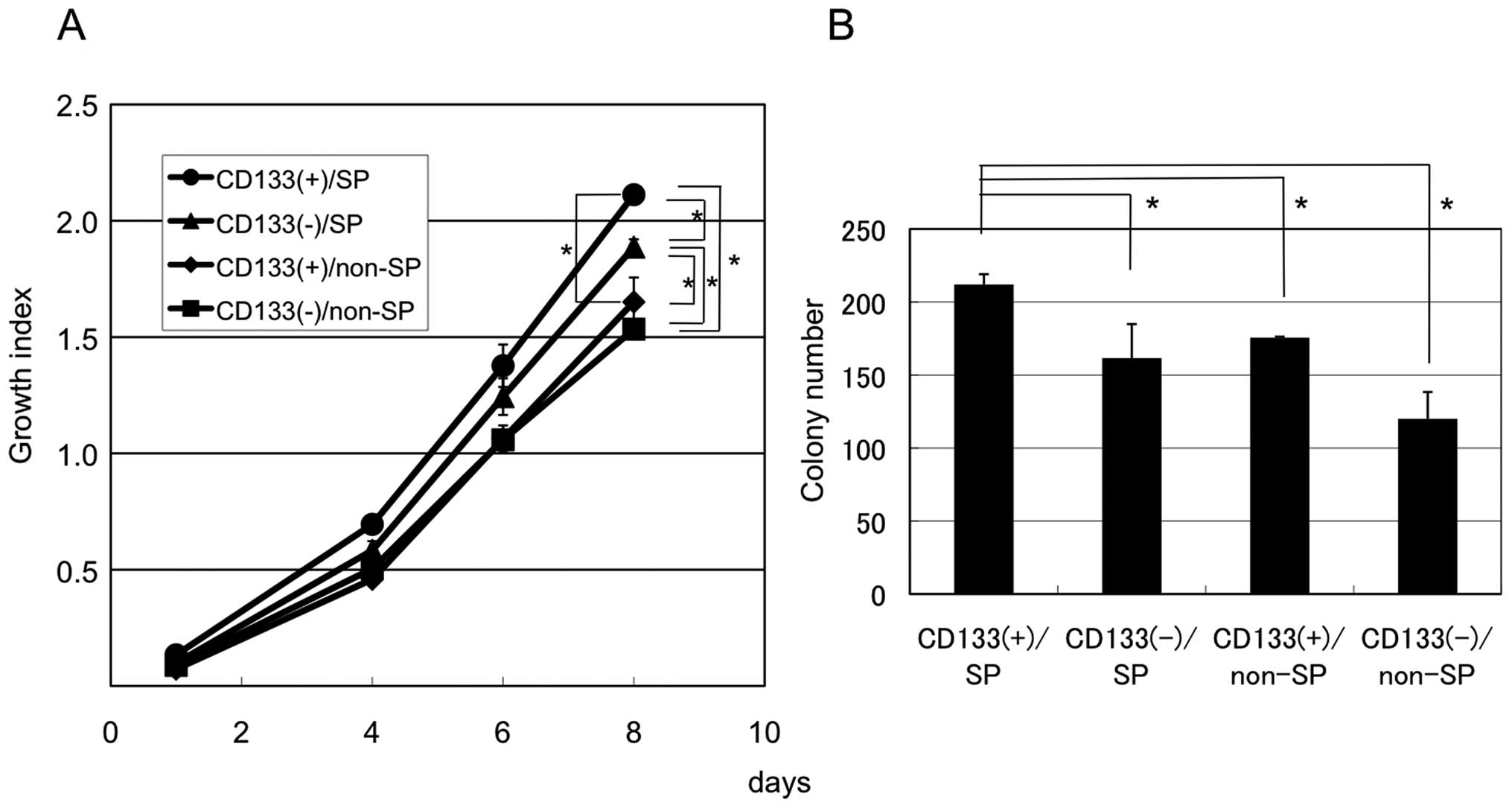
We next examined the tumorigenic potential of SP
cells with or without CD133 expression. CD133+/SP,
CD133+/non-SP, CD133−/SP and
CD133−/non-SP Ishikawa cells were sorted, purified and
cultured in normal growth medium for 8 days, and cell growth
compared by WST assay. As shown in Fig. 2A, in normal growth medium,
CD133+/SP Ishikawa cells grew significantly faster than
any other group.
To evaluate anchorage-independent growth of these
cells, we assessed their colony-forming ability in soft agar.
CD133+/SP cells formed more colonies than
CD133+/non-SP, CD133−/SP or
CD133−/non-SP cells (Fig.
2B). These results suggest that CD133+/SP Ishikawa
cells have the highest potential as CSCs.
CD133-expressing cells have increased
invasive ability via elevated levels of MT-MMT expression
We previously reported CD133 to be a prognostic
factor in endometrial cancer (14). The precise mechanisms, however,
remained unclear, but we speculated that increased invasive ability
of CD133+ cells might be the key. Among the genes highly
expressed in CD133+ cells, we took particular notice of
matrix metalloproteinase genes (MMPs) involved in cellular
invasion. We investigated the expression of a total of 26 MMPs in
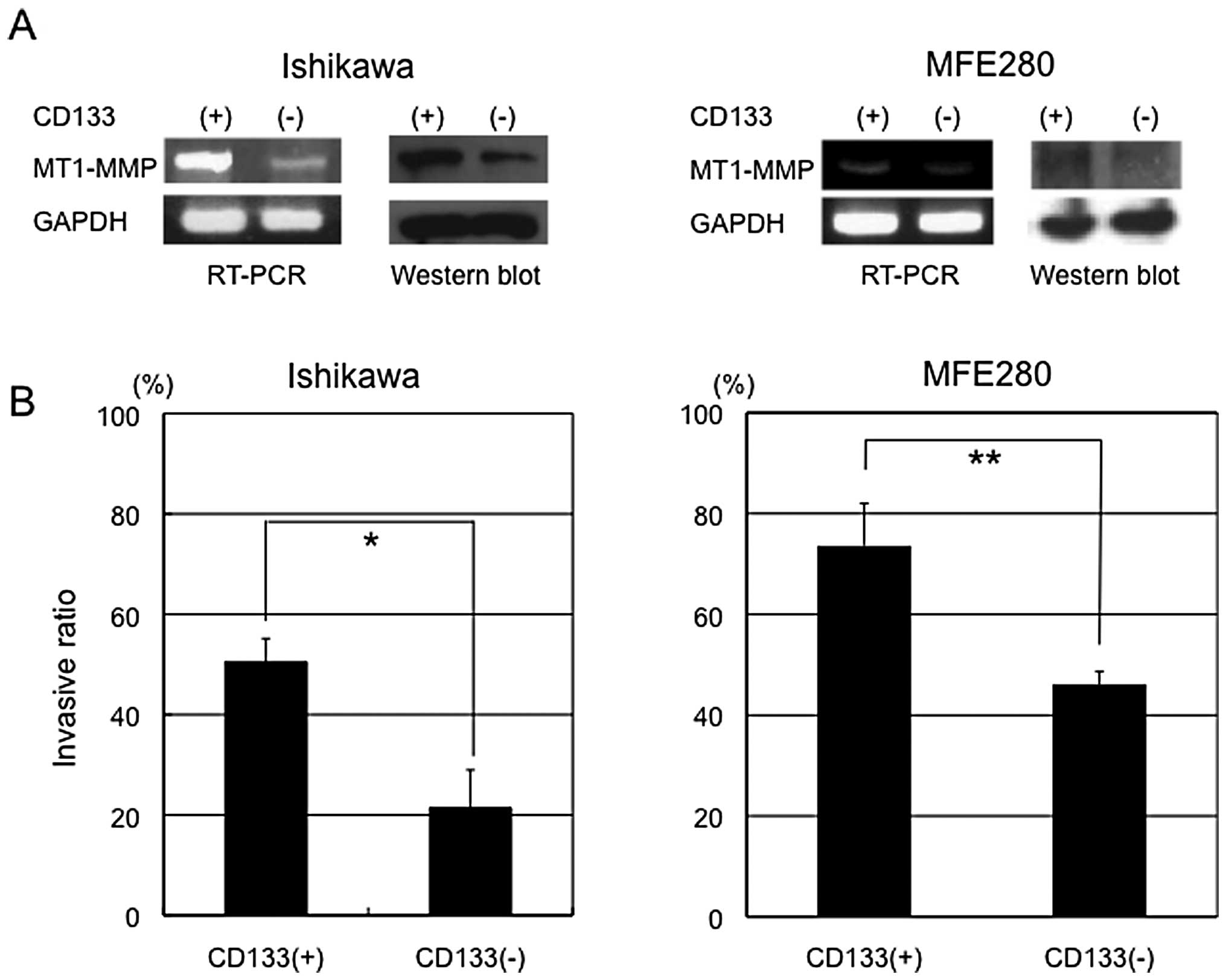
both CD133+ and CD133− Ishikawa cells and
found that only MMP14 (MT1-MMP) showed higher expression
(>2-fold) in CD133+ cells than CD133−
cells. We confirmed by RT-PCR and western blot analysis that
MT1-MMP was preferentially expressed in CD133+ cells
(Fig. 3A).
MT1-MMP plays a critical role in tumor invasion and
metastasis. We evaluated the invasion ability of CD133+
or CD133− endometrial cancer cells using an in
vitro invasion assay. Significant differences in invasive
ability were observed between CD133+ and
CD133− Ishikawa cells (50.5 vs 21.4%) (Fig. 3B), which were confirmed in another
cell type (MFE280 cells): CD133+ MFE280 cells showed
higher invasive ability than CD133− cells (73.6 vs
46.1%). These results suggest that CD133+ endometrial
cancer cells have increased invasive activity compared with
CD133− cells.
To confirm whether elevated MT-MMP1 expression is
essential for increased invasive ability of CD133+
endometrial cancer cells, we performed siRNA knockdown experiments
of MT1-MMP. RT-PCR and western blot analysis showed
successful knockdown of MT1-MMP by siRNA (Fig. 3C). Gelatin zymography also showed
significant inhibition of enzymatic activity of MT1-MMP by siRNA.
In vitro invasion assay revealed that the knockdown of
MT1-MMP led to decreased invasive ability in both Ishikawa
and MFE280 cells, from 147.6 to 52.0% and from 38.0 to 19.1%,
respectively (Fig. 3C), confirming
that MT1-MMP influences their invasive capacity. We also examined
the invasive ability of sorted CD133+ and
CD133− endometrial cancer cells, with or without
knockdown of MT1-MMP. In CD133+ Ishikawa and
MFE280 cells, invasive ability was significantly decreased by
successful knockdown of MT1-MMT from 159.7 to 63.6%, and 10.6 to
6.3%, respectively: knockdown efficacies in mRNA expression were 60
and 45%, respectively, compared with mock transfected cells
(Fig. 3D). Knockdown of
CD133− Ishikawa and MFE280 cells was not sufficient, and
invasive ability was not affected in these cells (47.9 vs 51.6%,
3.7 vs 2.3%, respectively). This was because constitutive levels of
mRNA MT1-MMP expression were much lower in CD133−
Ishikawa and MFE280 cells than in CD133+ Ishikawa and
MFE280 cells. These findings indicate that increased expression of
MT1-MMP in CD133+ endometrial cancer cells contributes
to their invasive ability.
Discussion
Several lines of evidence have identified CSC
populations using various CSC markers in many malignant tumors
(19). The characteristic of CSCs
are high potential for tumorigenicity, tumor invasion and
metastasis (20,21), and chemoresistance (22). Previously, we demonstrated that
CD133 is not only a CSC marker but also an independent prognostic
factor in endometrial cancer. In this study, we focused on the
invasive ability of CD133+ cells, in order to dissect
the mechanisms of the aggressive behavior of endometrial CSCs.
The SP phenotype is mediated by expression of ABCG2
protein, a superfamily of ATP-binding cassette (ABC) transporters,
which is associated with multi-drug resistance (23,24).
SP cells are known to be resistant to chemotherapeutic agents and
have been identified as CSCs in malignant solid tumors including
hepatocellular carcinoma (25),
lung cancer (26), ovarian cancer
(27), breast cancer (28) and pancreatic cancer (29). Our study demonstrated that Ishikawa
cells contained SP cells (0.69%). Ishikawa-SP cells are more
resistant to paclitaxel than non-SP cells. Furthermore, Ishikawa-SP
cells exhibited increased colony-forming ability in soft agar,
compared with non-SP cells, which shows that they have potential as
CSCs. This is consistent with a recent study (18). We speculate that the enriched SP
fraction in CD133+ cells contributes to their increased
chemoresistance.
Our data indicate that in Ishikawa cells, both
CD133+ and SP cells were capable of exhibiting the CSC
phenotype. What does this mean? Are there multiple types of
distinct CSCs or multiple markers of CSCs in this tumor type? Do
CD133+ cells significantly overlap with SP cells? The
ratio of the SP cells was 0.69% in CD133+ cells,
compared with 0.47% in CD133− cells: therefore,
overlapping population was not large. Nevertheless,
CD133+ and SP cells showed CSC-like characteristics
in vitro. Although we have not done in vivo analysis,
Kato et al recently observed a CSC-like tumorigenic
phenotype of SP cells (18). Thus,
both CD133 and SP may be independently considered as CSC markers
according to the current experimental criteria. We further
investigated the characteristics of SP or non-SP cells in
CD133+ and CD133− cells. SP/CD133+
Ishikawa cells had the greatest advantage of proliferation and
tumorigenicity in vitro compared with SP/CD133−,
non-SP/CD133+ and non-SP/CD133− cells. The
frequency of CD133+ cells in Ishikawa cells was
approximately 10.0%, while the ratio of SP fraction in
CD133+ Ishikawa cells was 0.69%. Based on these results,
about 0.069% SP/CD133+ cells were contained in Ishikawa
cells, which exhibit the highest CSC activity. Taken together, we
speculated that multiple types of CSCs with distinct markers may be
present, at least satisfying the minimum experimental conditions
for the definition of CSCs, but the small subset with concurrent
expression of markers appears to have the highest CSC activity.
Accumulating evidence has revealed that CSCs have
great invasive ability (30–32).
We confirmed that CD133+ endometrial cancer cells
exhibited higher expression of MT1-MMP, through which they
appeared to show increased invasive ability. Annabi et al
reported that MT1-MMP and MMP9 contributed to the invasive
phenotype in CD133+ brain cancer stem cells (30), and Kohga et al showed that
MMP2, which is activated by MT1-MMP, is required for invasive
ability in CD133+ hepatocellular carcinoma cells
(31), which is basically
consistent with our results.
Invasion of cancer cells, including lymph node and
distant metastasis, is believed to be associated with
epithelialmesenchymal transition. Kabashima et al
demonstrated that TGF-β-induced epithelial-mesenchymal transition
(EMT)-and invasion-associated gene alterations such as reduction of
E-cadherin and induction of Snail and MMP2 in a side population of
pancreatic cancer (32).
Circulating tumor cells in patients with advanced prostate and
breast cancer expressed epithelial protein such as adhesion
molecule, mesenchymal proteins including N-cadherin and vimentin,
and the CSC marker CD133 (33).
Our experimental model, in which CD133+ endometrial
cancer cells exhibited increased invasive capacity via elevated
MT1-MMP, might be suitable to study the role of EMT in metastasis.
We are currently investigating whether CD133+
endometrial cancer cells are likely to show EMT phenotypes during
the process of invasion.
The present microarray analyses revealed a total of
440 genes upregulated and 96 genes downregulated in
CD133+ cells, compared to CD133− cells. There
might be some genes other than MT1-MMP involved in aggressive
behavior of CD133+ cells. For example, Q9HCN2, a gene
encoding p53AIP, known to be a p53-regulated apoptosis-inducing
protein (34), was downregulated
in CD133+ cells. Thus, impaired apoptosis pathway might
be associated with aggressive phenotypes of CD133+
cells. Further extensive analysis with microarray data will
hopefully identify genes critical for determining phenotypes of
CD133+ cancer stem cells.
In summary, we found the characteristic features of
CD133+ endometrial cancer cells, enriched SP cells and
elevated MT1-MMP expression, through which they achieve increased
chemoresistance as well as invasive capacity. A subpopulation of SP
cells with CD133 expression showed the greatest CSC-like activity.
Further characterization of CD133+ cells is required to
identify the more condensed population of CSCs and to provide a
novel molecular target for this tumor type.
Acknowledgements
This study was supported by a
Grant-in-Aid for Scientific Research from the Japan Society for the
Promotion of Science (JSPS).
References
|
1.
|
Matsuda T, Marugame T, Kamo K, et al:
Cancer incidence and incidence rates in Japan in 2006: Based on
data from 15 population-based cancer registries in the monitoring
of cancer incidence in Japan (MCIJ) project. Jpn J Clin Oncol.
42:139–147. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Kyo S: Endometrial cancer stem cells: Are
they a possible therapeutic target? Curr Obstet Gynecol Rep.
2:1–10. 2013. View Article : Google Scholar
|
|
3.
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Singh SK, Hawkins C, Clarke ID, et al:
Identification of human brain tumour-initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Bao S, Wu Q, McLendon RE, Hao Y, Shi Q and
Hjelmeland AB: Glioma stem cells promote radioresistance by
preferential activation of the DNA damage response. Nature.
444:756–760. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Gu G, Yuan J, Wills M and Kasper S:
Prostate cancer cells with stem cell characteristics reconstitute
the original human tumor in vivo. Cancer Res. 67:4807–4815. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Miki J, Furusato B, Li H, et al:
Identification of putative stem cell markers, CD133 and CXCR4, in
hTERT-immortalized primary nonmalignant and malignant tumor-derived
human prostate epithelial cell lines and in prostate cancer
specimens. Cancer Res. 67:3153–3161. 2007. View Article : Google Scholar
|
|
8.
|
Eramo A, Lotti F, Sette G, et al:
Identification and expansion of the tumorigenic lung cancer stem
cell population. Cell Death Differ. 15:504–514. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Hermann PC, Huber SL, Herrler T, et al:
Distinct populations of cancer stem cells determine tumor growth
and metastatic activity in human pancreatic cancer. Cell Stem Cell.
1:313–323. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
O’Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110.
2007.PubMed/NCBI
|
|
11.
|
Ricci-Vitiani L, Lombardi DG, Pilozzi E,
Biffoni M, Todaro M and Peschle C: Identification and expansion of
human colon cancer - initiating cells. Nature. 445:111–115. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Monzani E, Facchetti F, Galmozzi E, et al:
Melanoma contains CD133 and ABCG2 positive cells with enhanced
tumourigenic potential. Eur J Cancer. 43:935–946. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Rutella S, Bonanno G, Procoli A, et al:
Cells with characteristics of cancer stem/progenitor cells express
the CD133 antigen in human endometrial tumors. Clin Cancer Res.
15:4299–4311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Nakamura M, Kyo S, Zhang B, et al:
Prognostic impact of CD133 expression as a tumor-initiating cell
marker in endometrial cancer. Hum Pathol. 41:1516–1529. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Nakamura M, Bodily JM, Beglin M, Kyo S,
Inoue M and Laimins LA: Hypoxia-specific stabilization of
HIF-1alpha by human papillomaviruses. Virology. 387:442–448. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Mizumoto Y, Kyo S, Mori N, et al:
Activation of ERK1/2 occurs independently of KRAS or BRAF status in
endometrial cancer and is associated with favorable prognosis.
Cancer Sci. 98:652–658. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Mizumoto Y, Kyo S, Ohno S, et al: Creation
of tumorigenic human endometrial epithelial cells with intact
chromosomes by introducing defined genetic elements. Oncogene.
25:5673–5682. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Kato K, Takao T, Kuboyama A, et al:
Endometrial cancer side-population cells show prominent migration
and have a potential to differentiate into the mesenchymal cell
lineage. Am J Pathol. 176:381–392. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Visvader JE and Lindeman GJ: Cancer stem
cells in solid tumours: accumulating evidence and unresolved
questions. Nat Rev Cancer. 8:755–768. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Maeda S, Shinchi H, Kurahara H, et al:
CD133 expression is correlated with lymph node metastasis and
vascular endothelial growth factor-C expression in pancreatic
cancer. Br J Cancer. 98:1389–1397. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Kojima M, Ishii G, Atsumi N, Fujii S,
Saito N and Ochiai A: Immunohistochemical detection of CD133
expression in colorectal cancer: a clinicopathological study.
Cancer Sci. 99:1578–1583. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Ma S, Lee TK, Zheng BJ, Chan KW and Guan
XY: CD133+ HCC cancer stem cells confer chemoresistance
by preferential expression of the Akt/PKB survival pathway.
Oncogene. 27:1749–1758. 2008.
|
|
23.
|
Kim M, Turnquist H, Jackson J, et al: The
multidrug resistance transporter ABCG2 (breast cancer resistance
protein 1) effluxes Hoechst 33342 and is overexpressed in
hematopoietic stem cells. Clin Cancer Res. 8:22–28. 2002.PubMed/NCBI
|
|
24.
|
Doyle LA and Ross DD: Multidrug resistance
mediated by the breast cancer resistance protein BCRP (ABCG2).
Oncogene. 22:7340–7358. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Chiba T, Kita K, Zheng YW, et al: Side
population purified from hepatocellular carcinoma cells harbors
cancer stem cell-like properties. Hepatology. 44:240–251. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Ho MM, Ng AV, Lam S and Hung JY: Side
population in human lung cancer cell lines and tumors is enriched
with stem-like cancer cells. Cancer Res. 67:4827–4833. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Szotek PP, Pieretti-Vanmarcke R, Masiakos
PT, et al: Ovarian cancer side population defines cells with stem
cell-like characteristics and Mullerian Inhibiting Substance
responsiveness. Proc Natl Acad Sci USA. 103:11154–11159. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Christgen M, Ballmaier M, Bruchhardt H,
von Wasielewski R, Kreipe H and Lehmann U: Identification of a
distinct side population of cancer cells in the Cal-51 human breast
carcinoma cell line. Mol Cell Biochem. 306:201–212. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Wang YH, Li F, Luo B, et al: A side
population of cells from a human pancreatic carcinoma cell line
harbors cancer stem cell characteristics. Neoplasma. 56:371–378.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Annabi B, Rojas-Sutterlin S, Laflamme C,
et al: Tumor environment dictates medulloblastoma cancer stem cell
expression and invasive phenotype. Mol Cancer Res. 6:907–916. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Kohga K, Tatsumi T, Takehara T, et al:
Expression of CD133 confers malignant potential by regulating
metalloproteinases in human hepatocellular carcinoma. J Hepatol.
52:872–879. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Kabashima A, Higuchi H, Takaishi H, et al:
Side population of pancreatic cancer cells predominates in
TGF-beta-mediated epithelial to mesenchymal transition and
invasion. Int J Cancer. 124:2771–2279. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Armstrong AJ, Marengo MS, Oltean S, et al:
Circulating tumor cells from patients with advanced prostate and
breast cancer display both epithelial and mesenchymal markers. Mol
Cancer Res. 9:997–1007. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Oda K, Arakawa H, Tanaka T, et al:
p53AIP1, a potential mediator of p53-dependent apoptosis, and its
regulation by Ser-46-phosphorylated p53. Cell. 102:849–862. 2000.
View Article : Google Scholar : PubMed/NCBI
|