Introduction
Epithelial-mesenchymal transition (EMT) is a
critical process in cell differentiation, morphogenesis, growth and
change of function (1). EMT is a
cellular mechanism recognized as a central feature of normal
physiological development such as organogenesis during embryonic
development and wound healing (2).
Several developmental milestones, including gastrulation, neural
crest formation, and heart morphogenesis, rely on the plastic
transition between epithelial cells and mesenchymal cells (3). However, dysregulated EMT appears to
occur in cancer progression and metastasis, as well as the
pathogenesis of chronic degenerative fibrotic disorders in several
organs (4). Epithelial and
mesenchymal cells differ in function and characteristics.
Epithelial cells are closely adjoined by specialized membrane
structures such as desmosomes, as well as tight, adherent, and gap
junctions, and have apical-basolateral polarization (5). Conversely, mesenchymal cells do not
form an organized cell layer, nor do they have the same
apical-basolateral organization, polarization of the cell surface
molecules (6). In culture,
epithelial cells grow in clusters, whereas mesenchymal cells have a
spindle shaped and fibroblast-like morphology (7). During the EMT process, epithelial
cells are removed of cellular polarity and epithelial cell-cell as
well as cell-matrix adhesion contacts are remodeled (8). Epithelial cells also acquire
fibroblast-like properties and show reduced intercellular adhesion
and increased motility (9). EMT is
characterized by increased expression of mesenchymal markers
(vimentin, thrombospondin, N-cadherin, vitronectin), increased
expression of extracellular matrix compounds (fibronectin),
decreased expression of epithelial markers (E-cadheirn, collagen
IV, occludin, desmoplakin, and mucin 1), altered location of
transcription factors (β-catenin, Snail, Slug, Twist, sox 10, and
NF-κB) and activation of kinase (Erk1/2 and PI3K/Akt) (10).
A critical molecular feature of EMT is
downregulation of E-cadherin, a cell adhesion molecule that acts as
a calcium-dependent transmembrane glycoprotein in the plasma
membrane of most normal epithelial cells in embryonic and adult
tissues (11). Two main types of
consensus binding sites have been shown to downregulate E-cadherin
expression, Ets sites and palindromic E-box (E-pal) (3). E-boxes are characterized by the
consensus sequence, 5′-CACCTG-3′, and consist of three E-boxes in
the human E-cadherin promoter: two upstream from the transcription
start site and one in exon 1 (12). Transcriptional repression of
E-cadherin has several important consequences that are of direct
relevance to the EMT process (13,14).
Several transcriptional factors that repress transcription of
E-cadherin have been identified during development and
translocation, including Snail, Slug, Twist, and ZEB1/2
downregulation, which induces EMT (15). In particular, the zinc finger
transcriptional repressors Snail and Slug have been implicated in
repression of E-cadherin transcription in vitro (16), as well as in mesoderm formation
together with Twist (17).
Ovarian cancer is the leading cause of death from
gynecological tumors, and epithelial ovarian cancers originating
from the ovarian surface epithelium (OSE) constitute ~90% of human
ovarian neoplasms (18).
Unfortunately, this disease is often not diagnosed until in
advanced stages and not before metastatic setting of the tumor due
to lack of early stage detection (19). OSE covering a nonovulating ovary is
a stationary mesothelium that exhibits epithelial and mesenchymal
characteristics with the capacity to give rise to inclusion cysts
through loss of mesenchymal characteristics (20). Mesodermally derived normal OSE
shows epithelial and mesenchymal features characterized by the
expression of both keratin and vimentin, and OSE cells can be
converted to a mesenchymal, fibroblast-like phenotype, at least
in vitro (21).
Interestingly, only low levels of E-cadherin are prominent in OSE
cells, while E-cadherin is present in OSE cells covering deep
clefts, inclusion cysts and ovarian tumors (22). Repression of E-cadherin has been
described in a high percentage of borderline ovarian tumors and
carcinomas when compared with benign tumors. Moreover, ascites
cells with low E-cadherin expression were found to be more invasive
than solid tumor cells (23). This
initial shift towards a more differentiated phenotype early in
tumor progression appears to be followed by reacquisition of
mesenchymal features in more advanced ovarian tumors, involving a
secondary loss of E-cadherin (24). Therefore, ovarian carcinomas show
unique features when compared to other epithelial derived
cancers.
We focused on EMT marker expression among ovarian
types. We report that E-cadherin was highly expressed in benign
tumors when compared to normal ovaries or malignant tumors. As a
result, expression of EMT-associated marker is dependent on ovarian
status, and is related to transcription factor via regulation of
the expression of E-cadherin. These results support the hypothesis
that detection of EMT-related marker alteration in OSE cells may be
important in the prognosis and diagnosis of ovarian neoplasms and
thus provide protection from the initiation of neoplastic
transformation.
Materials and methods
Clinical human ovarian cancer sample
Clinical specimens of human normal ovaries and
ovarian cancers were provided by Dr T.H. Kim at the Department of
Obstetrics and Gynecology (Soonchunhyang University, Bucheon,
Kyeonggido, Korea) between 2011 and 2013. Our ovarian materials
consist of 44 normal ovarian samples, 15 benign and 3 borderline
ovarian tumors and 12 malignant tumor samples as shown in Table I. Ovarian tissue samples from a
total of 74 patients who had undergone primary surgery for newly
diagnosed advanced stage of ovarian abnormalities were included in
this study. Histological type and grade were evaluated according to
the World Health Organization (WHO) by an experienced pathologist
at Soonchunhyang University Hospital. The specimens were divided
into three pieces and then stored at −80°C until extraction of RNA
and protein samples or fixed in 10% normal formalin for
histopathological analysis. Most ovarian tissues were obtained by
cystectomy, salpingooophorectomy, and salpingectomy. Additionally,
only patients who had not undergone chemotherapy or radiotherapy
prior to surgery were included in the study.
 | Table IClassification of normal ovaries and
ovarian cancers. |
Table I
Classification of normal ovaries and
ovarian cancers.
| Number |
|---|
| Type |
| Normal | 44 |
| Benign | 15 |
| Borderline | 3 |
| Malignant | 12 |
| Total | 74 |
| Ages |
| ≤45 | 35 |
| >46 | 39 |
| Weight of mass
(ave.) |
| Normal | - |
| Benign | 179.73±68.55 |
| Borderline | 826.50±327.39 |
| Malignant | 272.63±246.23 |
Cell culture
Three ovarian cancer cell lines of human origin were
used, SKOV-3, OVCAR-3, and BG-1 cells. The ovarian adenocarcinoma
cell lines, SKOV-3 and OVCAR-3, were purchased from the Korean Cell
Line Bank (KCLB, Seoul, Korea). An adenocarcinoma cell line, BG-1,
was provided by Dr K.S. Korach (National Institute of Environmental
Health Sciences, Research Triangle Park, NC, USA). SKOV-3 and BG-1
cells were routinely cultured in Dulbecco’s modified Eagle’s medium
(DMEM; Hyclone Laboratories, Inc., Logan, UT, USA) supplemented
with 10% (v/v) heat inactivated fetal bovine serum (FBS; Hyclone
Laboratory Inc.), antibiotics (100 U/ml penicillin and 100 μg/ml
streptomycin; Cellgro Mediatech Inc., Manassas, VA, USA), and 10 mM
HEPES (Gibco, Carlsbad, CA, USA) in a humidified incubator at 37°C
under 5% CO2. OVCAR-3 cells were grown in RPMI-1640
medium containing 15% FBS and maintained in a humidified atmosphere
of 5% CO2 at 37°C. Cells were trypsinized with
trypsin/EDTA and medium was changed twice a week.
Total RNA extraction and polymerase chain
reaction (PCR)
Total RNA of three ovarian cancer cells and clinical
samples was extracted using RNA extraction solution and TRIzol
reagent (Invitrogen Life Technologies, San Diego, CA, USA)
according to the general protocols. The complementary DNA (cDNA)
was obtained from 1 μg of total RNA using murine leukemia virus
reverse transcriptase (MMLV-RT; iNtRON Biotechnology, Sungnam,
Kyeonggido, Korea), 10 pM dNTP (Bioneer, Daejeon, Korea), random
nonamer primer (Takara Bio., Shiga, Japan), 5X RT buffer (iNtRON
Biotechnology) and RNase inhibitor (iNtRON Biotechnology) for 1 h
at 37°C. Prepared cDNA was amplified using 2.5 U of Taq polymerase
(iNtRON Biotechnology), 10X PCR buffer (iNtRON Biotechnology), 10
pM of dNTP (Bioneer), and sense/antisense primers for E-cadherin,
vimentin, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH)
genes. Each primer is listed in Table
II. The PCR reaction consisted of 30 cycles of denaturation at
95°C for 30 sec, annealing at 58°C for 30 sec and extension at 72°C
for 30 sec. Amplified products were separated on 1.5% agarose gel
containing ethidium bromide (EtBr; Sigma-Aldrich Co., St. Louis,
MO, USA) and confirmed using a Gel Doc 2000 (Bio-Rad Laboratories
Inc., Hercules, CA, USA).
| Table IISequences of reverse-transcription or
real-time PCR primer pairs. |
Table II
Sequences of reverse-transcription or
real-time PCR primer pairs.
| mRNA | Sequence
(5′-3′) |
|---|
| E-cadherin | F
TCCCATCAGCTGCCCAGAAA
R ATTGTCCTTGTGTCCTCAGT |
| Vimentin | F
ACGCCATCAACACCGAGTTCA
R AACTGTTACGCAGAGCCAGTG |
| Snail | F
AAGCTTCCATGGCGCGCTCTTTCCTCGTCAGGAAGCCC
R GGATCCTCAGCGGGGACATCCTGAGCAGCCGGACTCTTG |
| Slug | F
CCTTCCTGGTCAAGAAGCAT
R CACAGTGATGGGGCTGTATG |
| Twist | F
GGATCCATGATGCAGGACGTGTCCAGCTCGCCA
R CTCGAGCTAGTGGGACGCGGACATGGACCAGGC |
| GAPDH | F
ATGTTCGTCATGGGTGTGAACCA
R TGGCAGGTTTTTCTAGACGGCAG |
Real-time PCR
The real-time PCR mixture was composed of 2X SYBR
green premix (Takara Bio.), ROX (Takara Bio.) as a reference dye,
and reverse and forward primers (Bioneer) specific for the
EMT-related transcriptional factor gene. The reaction consisted of
denaturation at 95°C for 15 sec, followed by annealing at 58°C for
20 sec and extension at 72°C for 15 sec (40 cycles). The real-time
PCR reaction was carried out in triplicate for each sample. GAPDH
was used for normalization and the mRNA levels of Snail, Slug, and
Twist genes were compared. The mRNA levels of these genes were
determined using the 2−ΔΔCt method and the specific
primer pairs listed in Table
II.
Protein extraction and
quantification
To analyze the levels of EMT related protein, whole
lysates were extracted from ovarian cancer cell lines and clinical
ovary samples. First, 1X RIPA solution (50 mM Tris-HCl, pH 8.0, 150
mM NaCl, 1% NP-40, 0.5% deoxycholic acid, and 0.1% sodium dodecyl
sulfate) was applied to the three ovarian cancer cell lines
cultured in dishes to obtain the protein. Next, part of the excised
normal ovaries or ovarian cancers from patients was used to prepare
the whole tissue lysate. Briefly, clinical ovarian samples were
homogenized in PRO-PREP® lysis buffer (iNtRON
Biotechnology). After overnight incubation at 4°C, samples were
centrifuged at 14,000 rpm for 30 min at 4°C. The concentration of
total protein of ovarian cancer cells and clinical samples was then
determined using bicinchoninic acid (BCA; Sigma-Aldrich Co.) and
copper (II) sulfate (Sigma-Aldrich Co.) mixture. Next, 150 μl of
the solution was mixed with total protein and incubated at 37°C.
After 30 min, the absorbance was measured at 640 nm. Subsequently,
40 μg of proteins were mixed with protein loading buffer (Bio-Rad
Laboratories Inc.) containing β-mercaptoethanol and heated at 100°C
for 5 min.
Western blot analysis
Whole cell lysates were resolved by 12%
SDS-polyacrylamide gel electrophoresis (SDS-PAGE), after which
fractionated proteins were transferred to polyvinylidene difluoride
(PVDF) transfer membrane (Bio-Rad Laboratories Inc.). The membrane
was then blocked with 5% skim milk (Bio-Rad Laboratories Inc.) for
2 h to inhibit non-specific interaction with primary antibody, then
washed four times in 1X TBS buffer (adjusted to pH 7.6 using HCl)
containing 0.1% (v/v) Tween-20 (Bio-Rad Laboratories Inc.).
Membrane was incubated with primary antibody, mouse monoclonal
anti-E-cadherin (1:1,000 dilution, Abcam plc., Cambridge, UK),
anti-vimentin (1:1,000 dilution, Abcam plc.), or anti-GAPDH
(1:1,000 dilution, Santa Cruz Biotechnology, CA, USA) in 1X TBS
with 3% (w/v) BSA (Sigma-Aldrich Co.) and 0.1% (v/v) Tween-20
overnight at 4°C. The next day, samples were incubated with
specific antibody followed by the appropriate horseradish
peroxidase (HRP) conjugated secondary antibody, goat anti-mouse IgG
(1:3,000 dilution, Bio-Rad Laboratories Inc.), for 2 h. Positive
immunoreactive proteins were detected using an ECL kit (West-Q
Chemiluminescent Substrate Plus kit, GenDEPOT, Barker, TX,
USA).
Hematoxylin and eosin (H&E)
staining
Clinical human normal ovaries and ovarian cancer
tissue were fixed in 10% normal formalin (Sigma-Aldrich Co.),
embedded in paraffin blocks and cut with a sliding microtome (3-μm
sections). After deparaffination and rehydration with xylene and
ethanol, the slides were stained using hematoxylin and eosin
(Sigma-Aldrich Co.). To prevent sample contamination, stained
slides were hydrated and mounted using a hydrophobic mounting
solution. The morphology of tissues derived from ovaries was
observed by light microscopy using a BX51 microscope (Olympus,
Japan).
Immunohistochemistry (IHC)
For histological analysis, paraffin-embedded
sections were deparaffinized and rehydrated by xylene, ethanol, and
tap water. Antigen retrieval was accomplished by boiling the slides
in a microwave for 10 min in a chamber containing 0.01 M citrate
buffer (pH 6.0). To quench the endogenous peroxidase, tissue slides
were placed in 0.3% methanol/hydrogen peroxidase (Sigma-Aldrich
Co.) for 30 min. To block the non-specific binding of primary
antibodies, slides were exposed to 10% normal goat serum (Vector
Laboratories, Burlingame, CA, USA) for 1 h. Subsequently, the
slides were incubated with a mixture of anti-E-cadherin (1:100,
Abcam plc.) or anti-vimentin (1:100, Abcam plc) as the primary
antibody in 5% bovine serum albumin (Sigma-Aldrich Co.) in a
humidified chamber at 4°C overnight. The next day, the slides were
washed three times in 1X PBS-T (pH 7.4) and then incubated with
appropriate biotinylated secondary antibodies (1:500 dilution,
Vector Laboratories) for 30 min at room temperature. Slides were
subsequently rinsed with 1X PBS-T for 10 min, after which
Vectastain Universal Elite ABC kit reagent (Vector Laboratories)
was applied for 30 min. Immunoreactive complexes were detected
using DAB substrate (Sigma-Aldrich Co.) and counter stained with
hematoxylin. Finally, slides were mounted with a cover slip using
mounting solution. All slides were visualized under a BX51 light
microscope for digital photography.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism 5 (GraphPad Software, San Diego, CA, USA). Statistical
differences in RNA expression levels were identified by one-way
ANOVA followed by Tukey’s test. Results were expressed as the mean
± SD. Provability values <0.05 were considered statistically
significantly.
Results
Expression of EMT related markers in
ovarian cancer cells
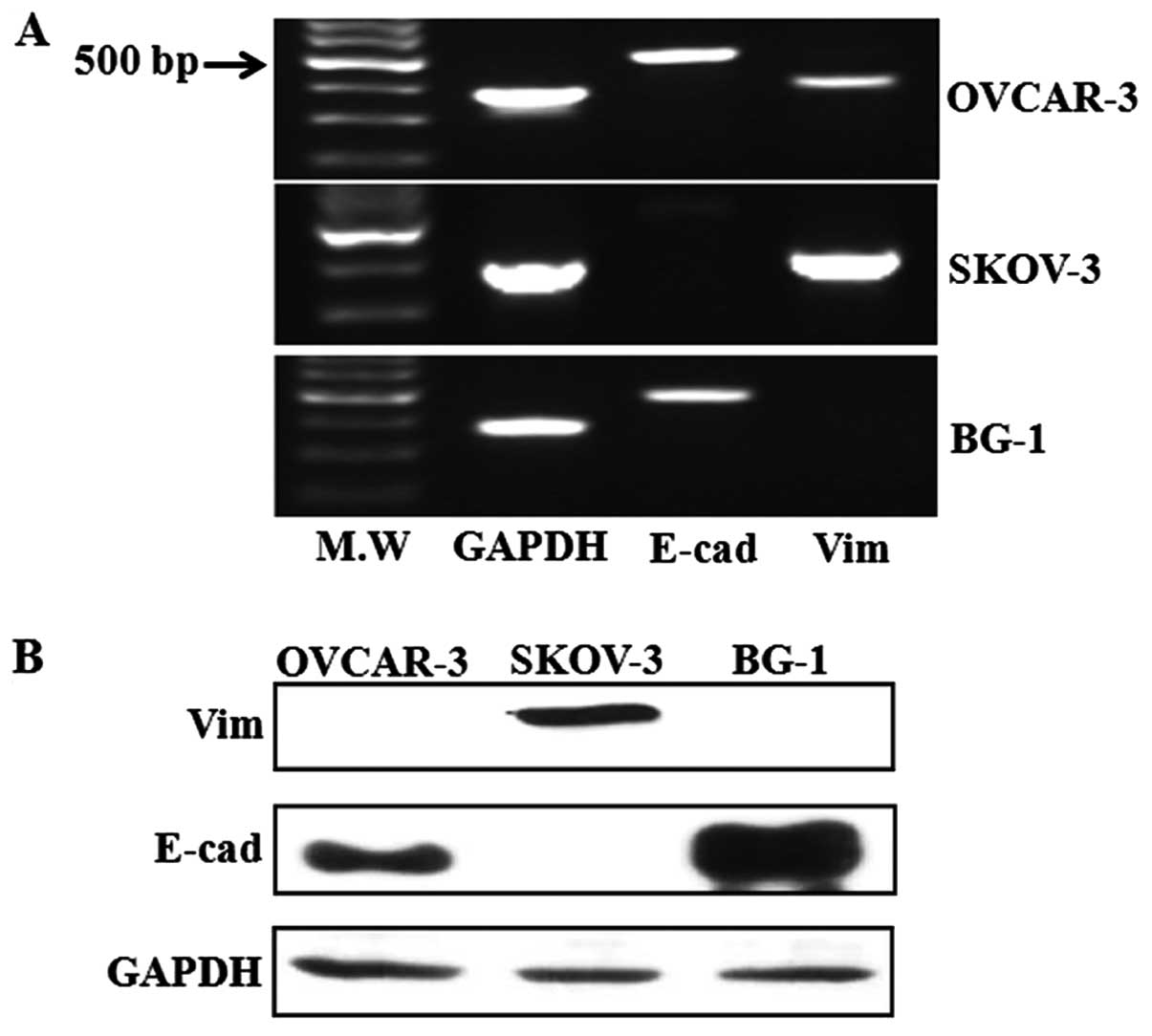
To observe the fate of EMT related markers in the
three ovarian cancer cell lines, we conducted reverse-transcription
PCR and western blot analysis to detect E-cadherin and vimentin
without any treatment. Reverse-transcription PCR showed that
E-cadherin expression was significantly higher in OVCAR-3 and BG-1
ovarian cancer cells, while its expression was absent in SKOV-3
cells (Fig. 1A). Additionally, the
expression levels of vimentin RNA as a typical mesenchymal cell
marker were observed in SKOV-3 and OVCAR-3 ovarian cancer cells. In
addition to RNA analysis, E-cadherin protein was expressed in
OVCAR-3 and BG-1 cells, but vimentin was only expressed in SKOV-3
cells (Fig. 1B). Of the three
ovarian cancer cell lines assayed, two cells showed a lack of
vimentin and the presence of E-cadherin, which is a typical pattern
for EMT related markers. These two cells lines (OVCAR-3 and BG-1)
have a cuboidal, epithelial morphology, while SKOV-3 cells have a
spindle-shaped morphology (data not shown).
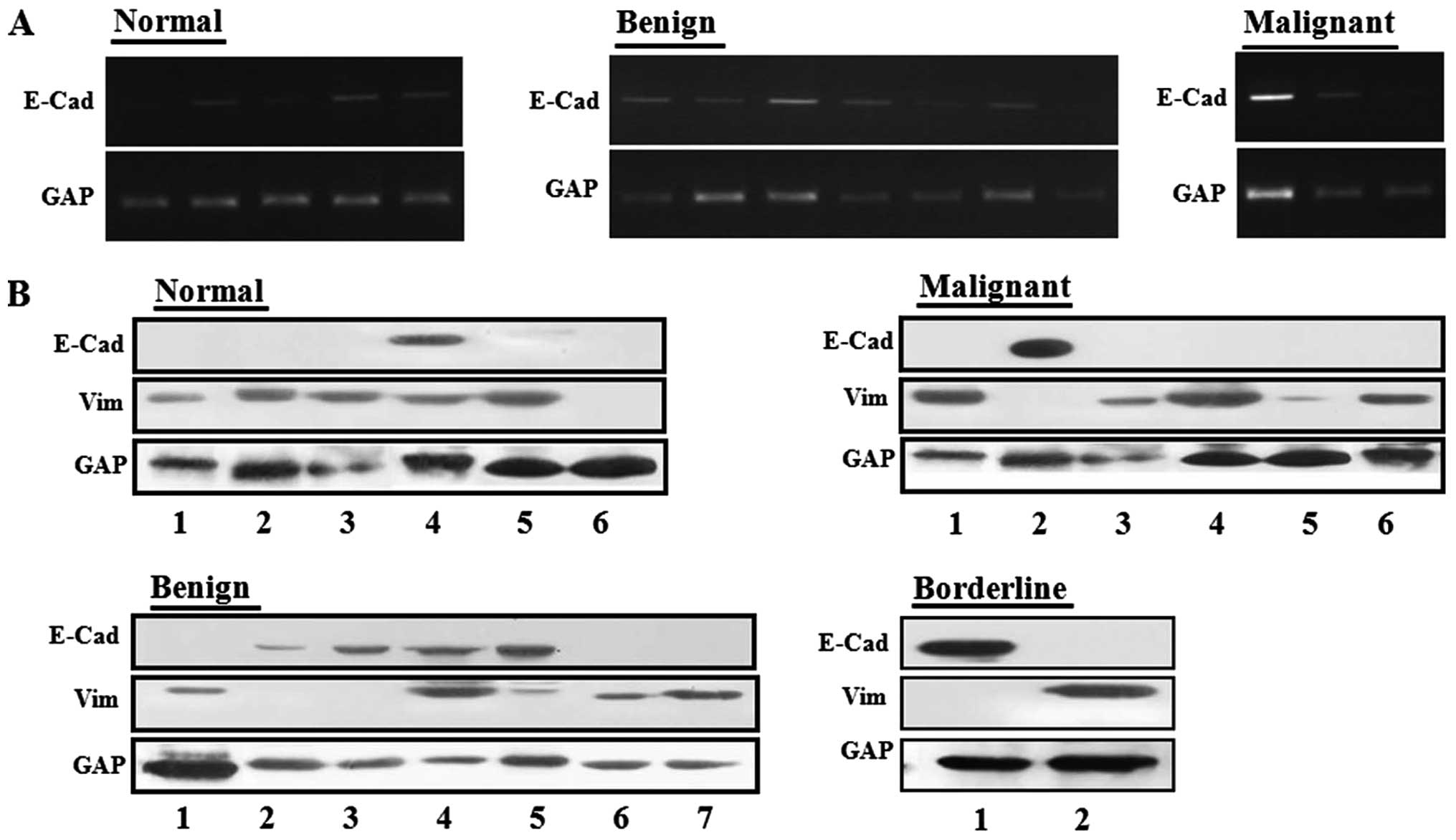
E-cadherin/vimentin expression in
clinical normal ovary and ovarian cancers
To measure the expression levels of E-cadherin RNA
in clinical ovarian cancers and normal ovary samples, we randomly
selected four groups: 1, six normal ovaries; 2, seven benign
ovarian cancers; 3, three malignant ovarian cancers; and 4, two
borderline ovarian cancers. Upon analysis of E-cadherin RNA
expression, E-cadherin was expressed in 50% of normal ovaries and
66.6% of malignant ovarian cancers, whereas it was expressed in
85.7% of benign ovarian cancers (Fig.
2A).
We also conducted western blot analysis to measure
protein levels of vimentin and E-cadherin in human clinical samples
obtained from patients. E-cadherin was observed in 16.7, 57.1, 50
and 16.7% of normal ovaries, benign, borderline, and malignant
ovarian tumors, respectively (Fig.
2B). Conversely, alteration of vimentin protein was observed in
83.3, 57.1, 50 and 66.7% of normal ovaries, benign, borderline, and
malignant tumors respectively. Therefore, the E-cadherin gene was
upregulated in benign ovarian tumors compared to normal ovary
tissues and decreased in malignant ovarian tumors.
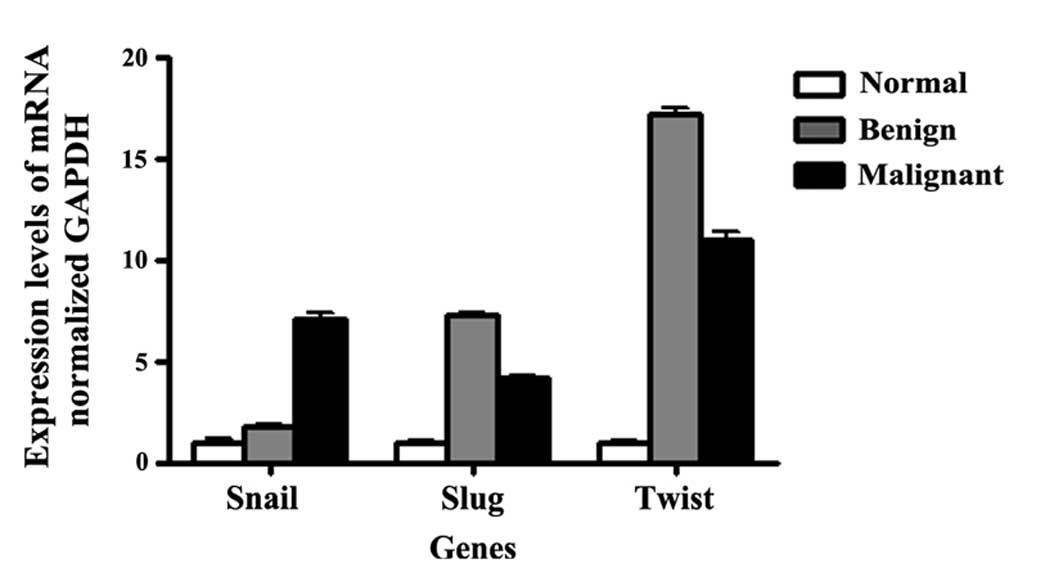
Quantification of EMT-related factors in
the ovarian tissue samples
To identify differences in the expression of
EMT-associated transcription factor between normal ovaries and
ovarian cancers, we conducted additional analyses focusing on genes
previously shown to be directly or indirectly involved in the EMT
process. EMT-associated factors, including Snail, Slug and Twist,
were overexpressed in the benign and malignant ovarian tumors
compared to normal ovaries (Fig.
3). Technical validation by real-time PCR using SYBR green
confirmed the relative overexpression of Snail by 1.8- and
7.1-fold, Slug by 7.3- and 4.1-fold, and Twist by 17.2- and
11.0-fold in benign and malignant ovary samples, respectively.
These transcriptional factors were significantly upregulated in
benign and malignant ovarian tumors relative to normal ovaries.
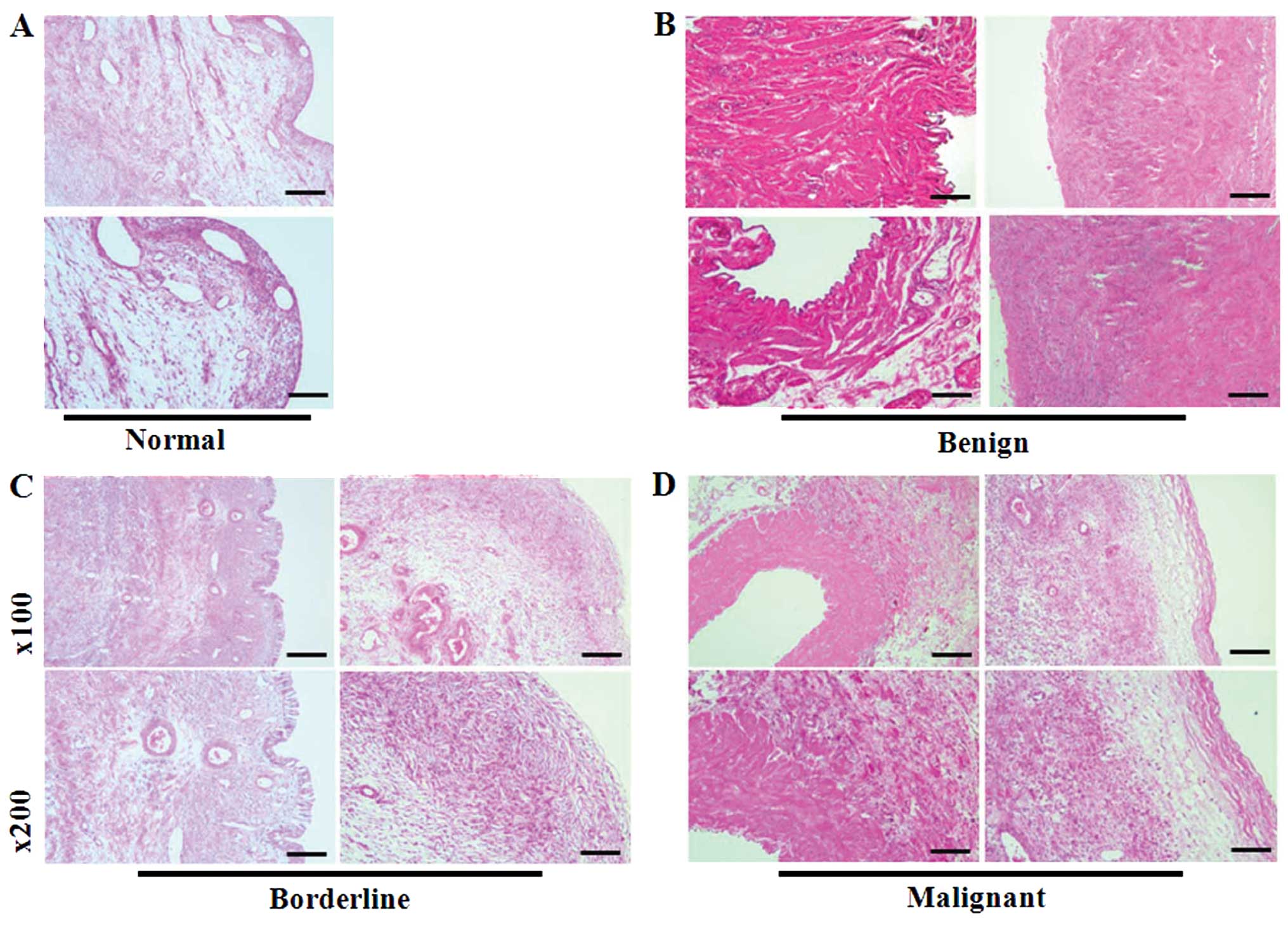
Histopathological analysis of human
normal ovaries and ovarian tumors
To investigate the histological features of normal
ovaries or ovarian cancers, tissue slides were stained with
H&E. As shown in Fig. 4A, the
specimen is a normal ovary specimen with a cystic follicle. A high
magnification view of the cyst lining demonstrates the luteinized
granulosa and theca cell layers on the top and bottom,
respectively. Benign tumor contained mature cystic teratoma
composed of multilocular cysts (Fig.
4B). Borderline tumors (mucinous endocervical type) consisted
of gray white cystic masses with fragments of solid mass containing
mucinous material and a capsule that had previously ruptured.
Additionally, these tumors had a smooth inner lining wall that was
thick and fibrotic. Low magnification revealed a papillary
architecture indistinguishable from that seen in serous borderline
tumors (Fig. 4C). Finally, serous
adenocarcinoma was used to represent malignant tumors. The cut
surface of these ovaries was yellow-gray and friable with extensive
necrosis (Fig. 4D).
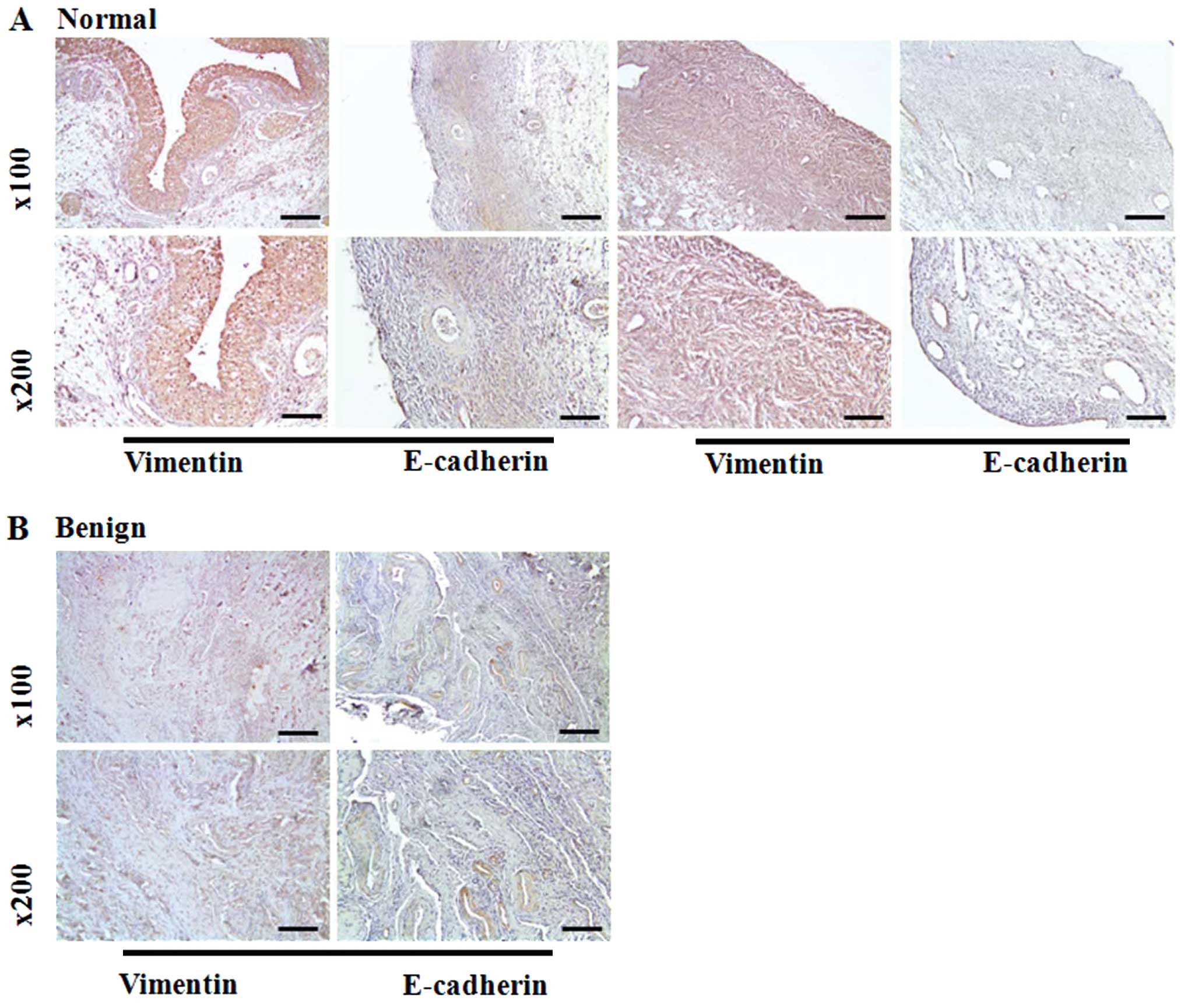
E-cadherin and vimentin expression in
clinical tissue samples
To observe the expression of EMT marker in clinical
samples, we conducted IHC analysis in normal ovaries and ovarian
cancer tissues. Initially, vimentin was significantly upregulated
in normal ovaries and malignant ovarian cancers (Fig. 5A and D). However, benign ovarian
tumors showed low expression of the vimentin protein in the tissues
(Fig. 5B and C). Epithelial cell
marker E-cadherin immunoreactivity was increased in benign and
malignant ovarian tumors relative to normal ovary tissues.
Conversely, the protein was detected in 50% of borderline ovarian
tumor samples.
Discussion
EMT is a phenomenon typically associated with
epithelial cells in which the normal physiological state is
characterized by abundant cortical E-cadherin expression and lack
of vimentin. In contrast, normal ovarian surface epithelium is
mesodermally-derived and expresses high levels of vimentin without
cortical E-cadherin (25). The
cellular features of EMT are a loss of epithelium-like polygonal
morphology, apicobasolateral cell polarity and adhesive contacts,
development of a fibroblast-like shape, reorganization of
cytoskeletal filaments, increased cell motility, and induction of
proteases for extracellular matrix (ECM) degradation, which is
prerequisite for migration and invasion (26).
In this study, we confirmed the expression of
EMT-related markers in three ovarian cancer cell lines, BG-1,
OVCAR-3, and SKOV-3. The morphology of SKOV-3 cells revealed a
fibroblastic shape that appeared to be mesenchymal. Additionally,
vimentin protein was expressed in SKOV-3 ovarian cancer cells, but
E-cadherin protein was not. The other two ovarian cancer cells
showed features of epithelial cell type such as expression of
E-cadherin and lack of vimentin protein. Based on these results,
the expression of typical EMT-related markers, E-cadherin and
vimentin, were altered based on ovarian cancer types, or
morphology. Upon further investigation, alteration of EMT markers
was observed in human clinical ovarian cancers or normal ovaries
provided by patients. In the case of E-cadherin protein, benign
ovarian cancers were upregulated by 3.4-fold when compared to
normal ovaries or malignant ovarian cancers. Conversely, expression
of vimentin protein was higher in normal ovaries or malignant
ovarian cancer (1.5-fold). Expression of E-cadherin and vimentin
protein was demonstrated by IHC staining in the human clinical
samples. Generally, vimentin protein was strongly expressed in
normal ovaries or malignant ovarian cancers, but E-cadherin protein
was barely expressed in the samples. Based on these results, normal
ovaries or malignant ovarian cancers displayed features such as
mesenchymal cells. While most epithelia expressed abundant
E-cadherin, it was absent from mesenchymally derived normal ovarian
surface epithelia, and aberrant epithelial differentiation is an
early event in epithelial ovarian carcinogenesis (27). In contrast to most carcinomas that
lose E-cadherin expression with progression, its protein is
abundant in primary differentiated ovarian carcinoma and available
data indicate a subsequent decrease of E-cadherin staining in
peritoneal metastases (28).
Post-translational modification of E-cadherin function was
suggested by data showing soluble E-cadherin in ascite or cystic
fluids from ovarian cancer patients and ratios of cystic
fluid/peripheral blood levels of soluble E-cadherin were
significantly higher in cystadenocarcinoma and borderline ovarian
tumors than benign tumors (29).
Moreover, low E-cadherin expression was observed in more invasive
tumors than benign tumors and associated with high tumor grade,
presence of peritoneal seeding and low overall survival (30). Such a phenomenon seems paradoxical,
but is re-expressed in metastatic lesions (31). The expression of E-cadherin is
altered drastically during peritoneal dissemination in ovarian
cancer, and the specific repressors may play an important role in
E-cadherin expression depending on the microenvironment of
metastatic sites.
Ovarian carcinoma cells often undergo an EMT process
before they detach or undergo metastasis. In ovarian carcinoma, the
E-cadherin expression of cancer cells floating in ascites and at
metastatic sites is lower than in primary ovarian tumors (32). Loss of E-cadherin gene expression
is primarily due to upregulation of the transcriptional repressors,
Snail, Slug, Twist or Sip1/ZEB1/2, which repress E-cadherin
transcription (33). During
malignant transformation, ovarian cancer cells lose
E-cadherin-mediated cell-cell interaction, upregulate N- or
P-cadherins and allow mesenchymal signaling through clustering of
collagen binding integrins (e.g., α2β1- and α3β1-integrin)
(34). In this study, several
transcription factors were analyzed to demonstrate the effect of
altered expression of EMT-related marker by real-time PCR. This
study showed the expression of transcriptional repressors for
E-cadherin in normal ovaries and ovarian cancers. Among the
repressors Snail, Slug, and Twist, malignant ovarian tumor
expressed higher levels of Snail, but not Slug or Twist. The
expression of Snail showed a stepwise increase from benign to
malignant tumors. Snail disrupts the functions of adherens
junctions as well as other cell-cell junctions, including tight
junctions, and blocks E-cadherin transcription by binding to CANNTG
(35). In another study,
overexpression of Snail was shown to be correlated with a higher
state or to be a marker of poor prognosis in malignancies because
it confers resistance to apoptosis induced by DNA damage (36,37).
In ovarian cancer cell lines, upregulation of Snail and Slug has
been correlated with resistance to radiation and paclitaxel and
shown to participate directly in p53-mediated pro-survival
signaling (38). Twist has been
found to be expressed at high levels in a number of human cancers,
including those of the breast, prostate, esophagus, and uterus, and
is associated with EMT and intravasation (39). Although increased Slug and Twist
gene expression was observed in this study, there was almost no
alteration of EMT-related markers or transcriptional factors
according to ovarian cancer types.
Taken together, in contrast to other epithelial
cancers, E-cadherin protein was highly expressed in ovarian benign
tumors, but barely expressed in normal ovaries. Additionally, the
results indicate that Snail expression might be more important than
that of other transcriptional repressors in ovarian tumor
prognosis. Moreover, upregulation of Snail expression followed by
the downregulation of E-cadherin enhances the invasiveness of
ovarian tumors. Further studies may be needed to clarify the role
of Snail in the aggressive behavior of ovarian cancer and the role
of metastasis.
Acknowledgements
This study was supported by a grant from the
Next-Generation BioGreen 21 Program (no. PJ009599), Rural
Development Administration, Republic of Korea.
References
|
1
|
Lee JM, Dedhar S, Kalluri R and Thompson
EW: The epithelial-mesenchymal transition: new insights in
signaling, development, and disease. J Cell Biol. 172:973–981.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hay ED: The mesenchymal cell, its role in
the embryo, and the remarkable signaling mechanisms that create it.
Dev Dyn. 233:706–720. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Larue L and Bellacosa A:
Epithelial-mesenchymal transition in development and cancer: role
of phosphatidylinositol 3′ kinase/AKT pathways. Oncogene.
24:7443–7454. 2005. View Article : Google Scholar
|
|
4
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vares G, Cui X, Wang B, Nakajima T and
Nenoi M: Generation of breast cancer stem cells by steroid hormones
in irradiated human mammary cell lines. PLoS One. 8:e771242008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Thiery JP: Epithelial-mesenchymal
transitions in development and pathologies. Curr Opin Cell Biol.
15:740–746. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aktas B, Tewes M, Fehm T, Hauch S, Kimmig
R and Kasimir-Bauer S: Stem cell and epithelial-mesenchymal
transition markers are frequently overexpressed in circulating
tumor cells of metastatic breast cancer patients. Breast Cancer
Res. 11:R462009. View
Article : Google Scholar
|
|
9
|
Kurrey NK, Jalgaonkar SP, Joglekar AV, et
al: Snail and slug mediate radioresistance and chemoresistance by
antagonizing p53-mediated apoptosis and acquiring a stem-like
phenotype in ovarian cancer cells. Stem Cells. 27:2059–2068. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Turley EA, Veiseh M, Radisky DC and
Bissell MJ: Mechanisms of disease: epithelial-mesenchymal
transition - does cellular plasticity fuel neoplastic progression?
Nat Clin Pract Oncol. 5:280–290. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sarrio D, Rodriguez-Pinilla SM, Hardisson
D, Cano A, Moreno-Bueno G and Palacios J: Epithelial-mesenchymal
transition in breast cancer relates to the basal-like phenotype.
Cancer Res. 68:989–997. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hemavathy K, Ashraf SI and Ip YT:
Snail/slug family of repressors: slowly going into the fast lane of
development and cancer. Gene. 257:1–12. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ohkubo T and Ozawa M: The transcription
factor Snail downregulates the tight junction components
independently of E-cadherin downregulation. J Cell Sci.
117:1675–1685. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Savagner P, Kusewitt DF, Carver EA, et al:
Developmental transcription factor slug is required for effective
re-epithelialization by adult keratinocytes. J Cell Physiol.
202:858–866. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nakayama K, Nakayama N, Katagiri H and
Miyazaki K: Mechanisms of ovarian cancer metastasis: biochemical
pathways. Int J Mol Sci. 13:11705–11717. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Batlle E, Sancho E, Franci C, et al: The
transcription factor snail is a repressor of E-cadherin gene
expression in epithelial tumour cells. Nat Cell Biol. 2:84–89.
2000. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Grille SJ, Bellacosa A, Upson J, et al:
The protein kinase Akt induces epithelial mesenchymal transition
and promotes enhanced motility and invasiveness of squamous cell
carcinoma lines. Cancer Res. 63:2172–2178. 2003.PubMed/NCBI
|
|
18
|
Auersperg N, Wong AS, Choi KC, Kang SK and
Leung PC: Ovarian surface epithelium: biology, endocrinology, and
pathology. Endocr Rev. 22:255–288. 2001.PubMed/NCBI
|
|
19
|
Hipp S, Berg D, Ergin B, et al:
Interaction of Snail and p38 mitogen-activated protein kinase
results in shorter overall survival of ovarian cancer patients.
Virchows Arch. 457:705–713. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Okamoto S, Okamoto A, Nikaido T, et al:
Mesenchymal to epithelial transition in the human ovarian surface
epithelium focusing on inclusion cysts. Oncol Rep. 21:1209–1214.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wong AS and Auersperg N: Normal ovarian
surface epithelium. Cancer Treat Res. 107:161–183. 2002.PubMed/NCBI
|
|
22
|
Strauss R, Li ZY, Liu Y, et al: Analysis
of epithelial and mesenchymal markers in ovarian cancer reveals
phenotypic heterogeneity and plasticity. PLoS One. 6:e161862011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cho EY, Choi Y, Chae SW, Sohn JH and Ahn
GH: Immunohistochemical study of the expression of adhesion
molecules in ovarian serous neoplasms. Pathol Int. 56:62–70. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ahmed N, Maines-Bandiera S, Quinn MA,
Unger WG, Dedhar S and Auersperg N: Molecular pathways regulating
EGF-induced epithelio-mesenchymal transition in human ovarian
surface epithelium. Am J Physiol Cell Physiol. 290:C1532–C1542.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ahmed N, Thompson EW and Quinn MA:
Epithelial-mesenchymal interconversions in normal ovarian surface
epithelium and ovarian carcinomas: an exception to the norm. J Cell
Physiol. 213:581–588. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Demir AY, Groothuis PG, Nap AW, et al:
Menstrual effluent induces epithelial-mesenchymal transitions in
mesothelial cells. Hum Reprod. 19:21–29. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Symowicz J, Adley BP, Gleason KJ, et al:
Engagement of collagen-binding integrins promotes matrix
metalloproteinase-9-dependent E-cadherin ectodomain shedding in
ovarian carcinoma cells. Cancer Res. 67:2030–2039. 2007. View Article : Google Scholar
|
|
28
|
Imai T, Horiuchi A, Shiozawa T, et al:
Elevated expression of E-cadherin and alpha-, beta-, and
gamma-catenins in metastatic lesions compared with primary
epithelial ovarian carcinomas. Hum Pathol. 35:1469–1476. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sundfeldt K, Ivarsson K, Rask K, Haeger M,
Hedin L and Brannstrom M: Higher levels of soluble E-cadherin in
cyst fluid from malignant ovarian tumours than in benign cysts.
Anticancer Res. 21:65–70. 2001.PubMed/NCBI
|
|
30
|
Vergara D, Merlot B, Lucot JP, et al:
Epithelial-mesenchymal transition in ovarian cancer. Cancer Lett.
291:59–66. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu J, Ikeguchi M, Nakamura S and Kaibara
N: Re-expression of the cadherin-catenin complex in lymph nodes
with metastasis in advanced gastric cancer: the relationship with
patient survival. J Exp Clin Cancer Res. 21:65–71. 2002.PubMed/NCBI
|
|
32
|
Lengyel E: Ovarian cancer development and
metastasis. Am J Pathol. 177:1053–1064. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Elloul S, Elstrand MB, Nesland JM, et al:
Snail, Slug, and Smad-interacting protein 1 as novel parameters of
disease aggressiveness in metastatic ovarian and breast carcinoma.
Cancer. 103:1631–1643. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Patel IS, Madan P, Getsios S, Bertrand MA
and MacCalman CD: Cadherin switching in ovarian cancer progression.
Int J Cancer. 106:172–177. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cano A, Perez-Moreno MA, Rodrigo I, et al:
The transcription factor snail controls epithelial-mesenchymal
transitions by repressing E-cadherin expression. Nat Cell Biol.
2:76–83. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yin T, Wang C, Liu T, Zhao G, Zha Y and
Yang M: Expression of snail in pancreatic cancer promotes
metastasis and chemoresistance. J Surg Res. 141:196–203. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Miow QH, Tan TZ, Ye J, et al:
Epithelial-mesenchymal status renders differential responses to
cisplatin in ovarian cancer. Oncogene. May 26–2014.(Epub ahead of
print). View Article : Google Scholar
|
|
38
|
Haslehurst AM, Koti M, Dharsee M, et al:
EMT transcription factors snail and slug directly contribute to
cisplatin resistance in ovarian cancer. BMC Cancer. 12:912012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lo HW, Hsu SC, Xia W, et al: Epidermal
growth factor receptor cooperates with signal transducer and
activator of transcription 3 to induce epithelial-mesenchymal
transition in cancer cells via up-regulation of TWIST gene
expression. Cancer Res. 67:9066–9076. 2007. View Article : Google Scholar
|