Introduction
Cancer is a major public health problem in the
United States as well as many other countries (1). It is the second leading cause of
death after heart disease, one quarter of Americans dying from
cancers. Billions of dollars are spent every year for treatment and
prevention of cancer (2). Among
cancers, malignant pleural mesothelioma (MPM), which occurs in the
peritoneum and pleura, is rare but progressive and rapidly fatal
(3). The onset has been related to
exposure to occupational and environmental asbestos (4). The disease often develops in male and
the incidence increases with age (3). Diagnosis or management for MPM has
not been precisely defined. Diagnosis is determined from the
immunocytochemical analysis and histological examination of biopsy
or diagnostic laparoscopic specimens (3). Median survival time is only 5–12
months and mean symptoms-to-survival time is 345 days due to lack
of effective treatment (5,6). Other treatments are possible, but
local excision and radical resection are preferred (7,8).
Chemotherapy has an important role in the palliative treatment.
Recently, application of natural products has gained attention as
cancer chemopreventive therapy (9). Licorice root is a common source of
licorice used in traditional and herbal medicines. The
pharmacological activity of licorice has been used for the
treatment of human diseases such as cancer, gastric ulcers,
bacterial infections and immunodeficiency (10). LCA is a flavonoid extracted from
licorice and exerts anti-tumor properties, anti-parasites and
anti-microbial activity (11–13).
LCA inhibits TNF-α-induced nuclear factor-κB activation, leading to
downregulation of inflammatory cytokines with concomitant reduction
of carrageenan-induced inflammatory responses (14,15).
However, it remains unclear whether LCA induces apoptosis by
suppressing the expression of specificity protein 1 (Sp1) in human
MPM cells.
Sp1 is one of the first eukaryotic transcription
factors to be identified and cloned and it is ubiquitously
expressed in mammalian cells (16,17).
It is also a sequence-specific DNA-binding protein and plays an
important role in various physiological processes, including
angiogenesis, cell cycle regulation, hormonal activation and
apoptosis (18). In many studies,
the expression levels of Sp1 are higher in cancer cells such as
human pancreatic cancer, breast cancer, colorectal cancer, gastric
carcinoma, hepatocellular carcinoma and thyroid carcinoma than in
normal cells (19). Further, there
is also evidence that Sp1 protein functions as an important factor
in the development of tumors, growth and metastasis (17). However, involvement of Sp1 protein
in the pathogenesis of MPM is unknown. In order to verify its
therapeutic potential, we investigated whether LCA could induce
apoptosis by suppressing Sp1 protein level in MSTO-211H and H28
cells.
Materials and methods
Plant material
The roots of Glycyrrhiza inflata were
purchased from Chonnam Herb Association. A voucher specimen
(MNUYG-003) was deposited in the College of Pharmacy, Mokpo
National University, Muan, Korea.
Extraction and isolation
The air-dried, powdered G. inflata roots (600
g) were extracted twice with MeOH (4 l) by sonication for 3 h.
After filtration, the MeOH extract was evaporated and suspended in
distilled water and then defatted with n-hexane (1 l). The
aqueous layer was partitioned with methylene chloride (3×1 l). The
evaporation residue (5 g) was subjected to flash silica gel
chromatography, using n-hexane:EtOAc:MeOH solvent system
(2:1:0.1-1:1:0.1-100% MeOH), to afford 10 fractions. Fractions 2, 3
and 4 were subjected to further flash silica gel chromatography,
with a chloroform:MeOH (100:1) eluent system, to afford LCA (50
mg). LCA was further purified by column chromatography using RP18
to an analytically acceptable purity.
Cell culture
The human MPM cells MSTO-211H and H28 were purchased
from the American Tissue Culture Collection (Manassas, VA, USA) The
MSTO-211H and H28 cells were grown in RPMI-1640 medium,
supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 100
U/ml each of penicillin and streptomycin (Thermo Scientific, Logan,
UT, USA) at 37°C in a humidified atmosphere of 5% CO2
and 95% air.
MTS assay
Effects of LCA on the cell viability were determined
using the MTS assay according to the manufacturer’s instructions.
The cells were plated on a 96-well plate at a density of
8×104 cells/well for MSTO-211H and 3×104
cells/well for H28 and then treated with various concentrations of
LCA for 24 and 48 h. After addition of MTS solution to culture
media, the absorbance was measured at 490 nm using an Enspire
Multimode Plate reader (Perkin-Elmer, Akron, OH, USA). The data
were expressed as the percentage of cell viability compared with
the control.
DAPI staining
The number of cells undergoing apoptosis, which was
induced by LCA treatment, was quantified using DAPI staining. The
cells featuring nuclear condensation and fragmentation were
determined using nucleic acid stained with
4′-6-diamidino-2-phenylindole (DAPI) (Sigma-Aldrich). Forty-eight
hours after treatment with different doses of LCA (10, 20 and 40
μM), the MSTO-211H and H28 cells were harvested and fixed in 100%
MeOH at RT for 20 min. The cells were seeded on slides, stained
with DAPI (2 μg/ml) and then analyzed under a FluoView confocal
laser microscope (Fluoview FV10i, Olympus Corp., Tokyo, Japan).
Cell cycle analysis
Following treatment of the cells with LCA for 48 h,
the detached cells and adherent cells were collected separately.
The cells were washed with cold PBS and then centrifuged before
being fixed in 70% EtOH for 3 h at −20°C. Before flow cytometry
analysis, the cells were centrifuged and incubated for 30 min at
37°C in PBS to allow for the release of low-molecular weight DNA.
Following centrifugation, the cell pellets were suspended and
treated with Muse™ cell cycle reagent using a Muse™ Cell Analyzer
(Merck Millipore, Billerica, MA, USA).
Reverse-transcription polymerase chain
reaction (RT-PCR)
Total RNAs were isolated using TRIzol®
Reagent (Life Technologies, Carlsbad, CA, USA) and 2 μg of RNA was
used to synthesize cDNA using the HelixCript™ 1st-strand cDNA
synthesis kit (NanoHelix). cDNA was obtained by PCR amplification
using β-actin-specific and Sp1-specific primers using the following
PCR conditions: 35 cycles of 1 min at 95°C, 1 min at 56°C and 1 min
at 72°C. The β-actin primers used were: forward,
5′-GTG-GGG-CGC-CCC-AGG-CAC-CA-3′; and reverse,
5′-CTC-CTT-AAT-GTC-ACG-CAC-GAT-TTC-3′. The Sp1 primers used were:
forward, ATG CCT AAT ATT CAG TAT CAA GTA; and reverse, CCC TGA GGT
GAC AGG CTG TGA. PCR products were analyzed by 2% agarose gel
electrophoresis.
Mitochondrial membrane potential
assessment
To investigate the mitochondrial membrane
permeability, MSTO-211H and H28 cells were cultured with LCA at
given doses for 48 h. After washing with PBS, cells were
dissociated by trypsin. For detection of the depolarized
mitochondria of cells undergoing apoptosis, MitoPotential working
solution to the MPM cells was added for 20 min incubation in the
dark. Muse MitoPotential 7-AAD reagent was added to each well and
then kept in the dark for 5 min. The experiment was analyzed by
Muse™ Cell Analyzer.
Multi-caspase analysis
MSTO-211H and H28 were seeded on a 10-cm dish and
treated with various concentrations of LCA (10, 20 and 40 μM) for
48 h. Caspase activity was measured using the Multi-caspase cell
kit (Merck Millipore) and analyzed by Muse™ Cell Analyzer.
Annexin V staining
The cells (MSTO-211H and H28) were seeded on a 10-cm
dish and treated with various concentrations of LCA (10, 20 and 40
μM) for 48 h. Both adherent and floating cells were harvested,
washed once with PBS. For detection of apoptosis, cells were
incubated with Muse™ Annexin V & Dead Cell reagent (Merck
Millipore) for 20 min at RT in the dark. Apoptotic and necrotic
cells were analyzed by Muse™ Cell Analyzer.
Western blot analysis
The MSTO-211H and H28 cells were treated with LCA
for 48 h and lysed in RIPA buffer (Intron Biotechnology, Seoul,
Korea) containing a protease inhibitor. Protein concentrations were
determined by BCA assay (Pierce Chemical, Rockford, IL, USA).
Proteins were separated by SDS-PAGE and transferred to
polyvinylidene fluoride (PVDF) membrane (GE Healthcare, Little
Chalfont, UK). The membrane was blocked by 3 or 5% fat-free skim
milk, then probed with primary and secondary antibodies. Bands of
interest were visualized using ECL Western blotting detection
regents (GE Healthcare).
Statistical analysis
The experimental data are expressed as the mean ±
standard deviation and statistical significance between different
groups was determined using Student’s t-test. Triplicate
experiments were performed on the data obtained for the
experimental groups. P-value of <0.05 was considered
statistically significant.
Results
LCA inhibits cell growth of MPM
In order to investigate the anti-proliferative
activity of licochalcone, MPM cells (MSTO-211H and H28) were
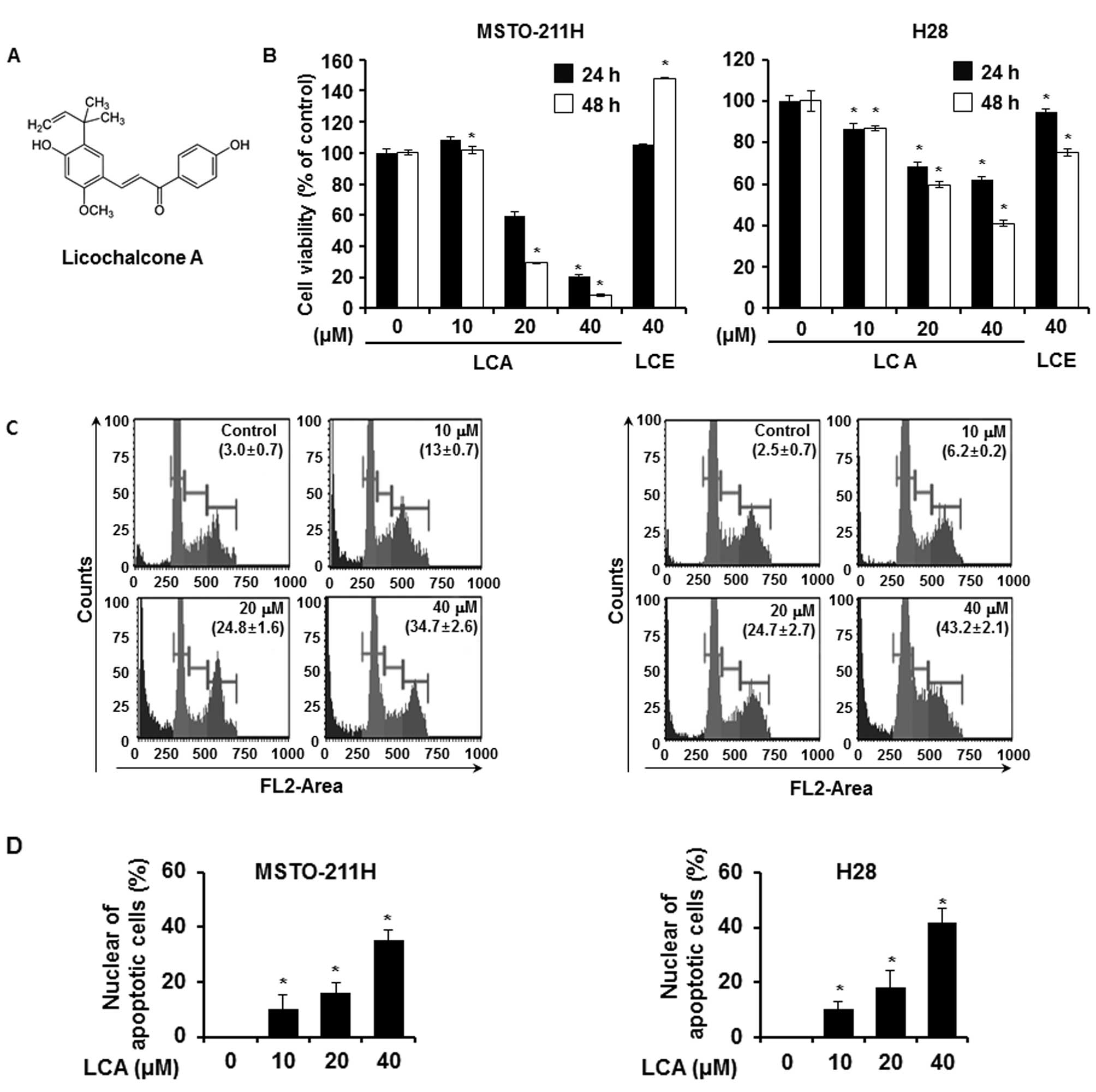
treated with LCA or LCE. The structure of LCA is depicted in
Fig. 1A. LCA and LCE were isolated
from the roots of Glycyrrhiza inflata (20). Structure of LCA is similar to that
of LCE, but the positions of the methyl groups of allyl group in
LCA were different from those of LCE (20,21).
To determine inhibitory effects of LCA and LCE on cell viability,
MTS assay was done after 24 and 48 h. As a results, it was found
that the IC50 value of LCA was 26 and 30.4 μM in
MSTO-211H and H28, respectively. LCA inhibited cell survival of
MSTO-211H cell in a dose-dependent manner, 101.6, 29.0 and 8.4%
over the concentrations used (10, 20 and 40 μM). Similarly, H28
cells represented cell viability of 86.7, 59.7 and 41.0% at the
concentrations used (10, 20 and 40 μM), respectively. However, LCE
had a weak cytotoxic activity in H28 but not MSTO-211H for 48 h
(Fig. 1B).
LCA treatment regulates the expression of
cell cycle arrest and induces apoptosis in MPM cells
In order to examine correlation of altered cell
cycle to growth inhibitory effects of LCA, we carried out PI
staining of LCA-treated cells and then flow cytometric analyses. We
analyzed cell cycle profiles of cells treated with LCA (10, 20 and
40 μM) for 48 h by Muse™ Cell Analyzer. As a result, we observed
that sub-G1 phase was increased in a dose-dependent
manner by LCA, indicating that sub-G1 phase was
increased from 3.0±0.7 to 34.7±2.6% for MSTO-211H (Fig. 1C, left) and was accumulated from
2.5±0.7 to 43.2±2.1% for H28 (Fig.
1C, right). LCA induced considerably fragmentation and
condensation of DNA of MPM cells in a dose-dependent manner
(Fig. 1D). Annexin V staining
indicated that phosphatidylserine was exposed on the outer leaflet
of the membrane during cell death. Early apoptosis of cells was
progressed to late apoptosis by LCA treatment. In MSTO-211H and
H28, late apoptotic cell populations were increased from 2.8±1.5 to
47.1±0.9% (Fig. 5C) and from
2.7±0.1 to 17.6±4.9%, respectively (Fig. 5D).
LCA modulates Sp1 protein level in MPM
cells
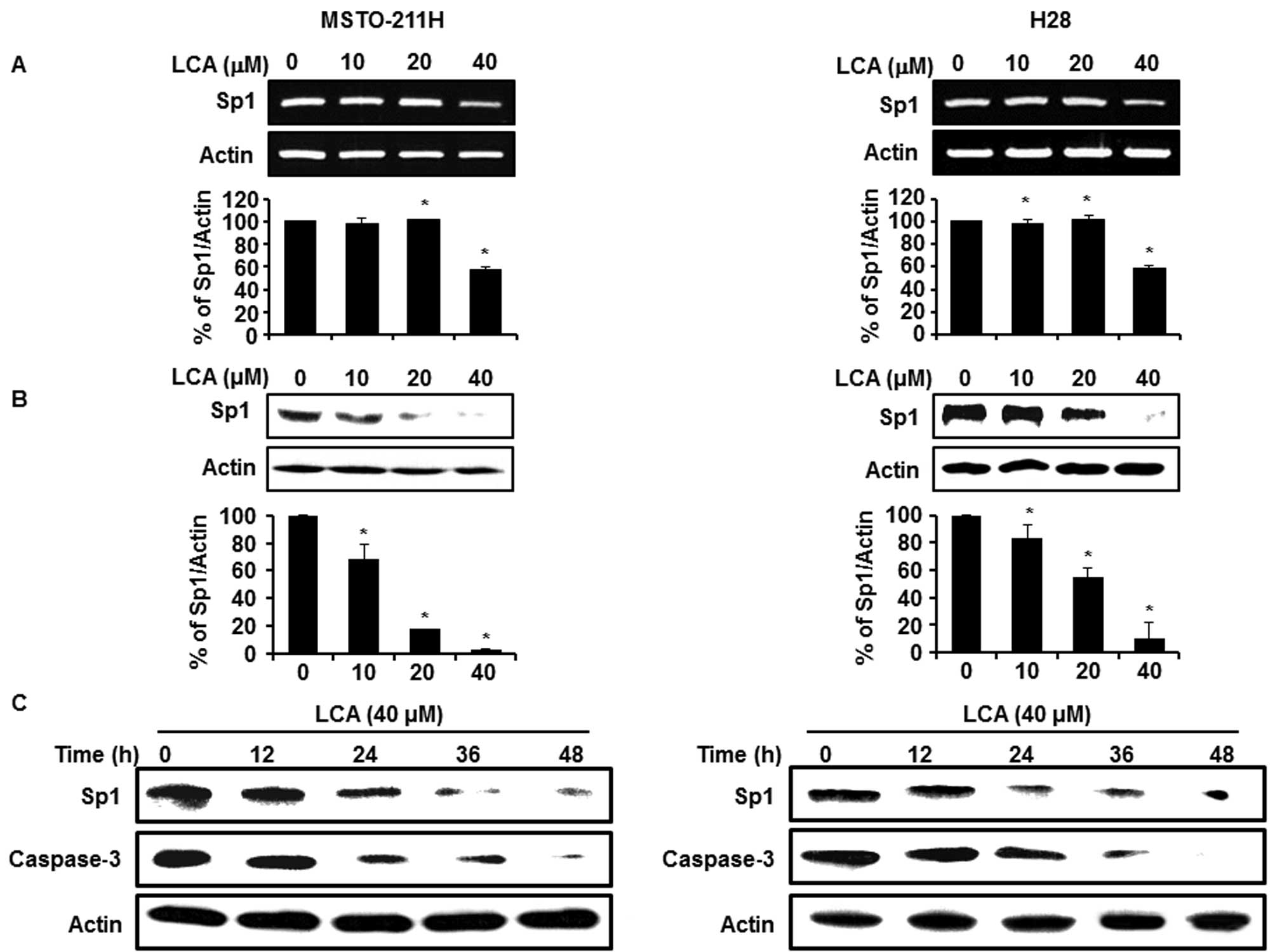
To demonstrate the link between Sp1 and cell death,
expression levels of Sp1 were primarily examined by western
blotting after treatment of MPM cells with LCA (10, 20 and 40 μM).
Fig. 2A and B show that Sp1 mRNA
and protein expression levels were reduced by LCA in a
dose-dependent manner. Also, we observed that the caspase-3
associated with cell death was cleaved in a time-dependent manner
(12, 24, 36 and 48 h; Fig. 2C).
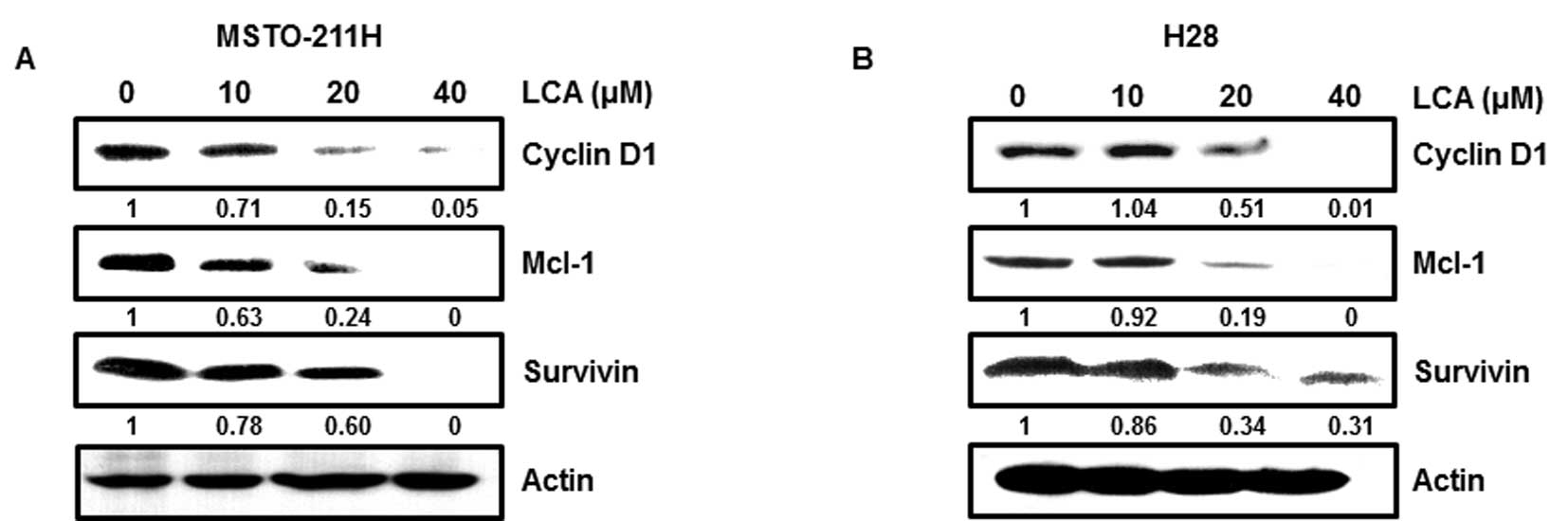
Sp1 is a transcription factor that modulates cell cycle regulation,
anti-proliferative and apoptotic cell death by regulating the
promoter of the target gene. Sp1 regulates the expression of Cyclin
D1, Mcl-1 and Survivin contributing to cell proliferation (23). In line with previous results, Sp1
target proteins are significantly reduced by LCA treatment
(Fig. 3).
LCA affects the permeability of the
mitochondrial membrane
Mitochondrial membrane potential is closely
associated with Bcl-2 family-mediated apoptosis. To investigate
whether LCA could affect the mitochondrial membrane potential, the
degree of depolarization was measured by staining of MPM cells with
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine
iodide reagent. In MSTO-211H (Fig.
4A), total depolarized cell population was 38.7±1.3, 51.9±1.2
and 72.1±1.7% and in H28 it was 14.1±4.7, 62.3±1.9 and 94.4±1.8% in
a dose-dependent manner, respectively. We measured the levels of
proteins associated with extrinsic apoptosis pathway (24). CHOP, which is increased by ER
stress, activates TNF-related apoptosis-inducing ligand (TRAIL)
death receptors DR4 and DR5 (25).
The expression levels of CHOP, DR4 and DR5 were substantially
increased in MSTO-211H (Fig. 4C)
and H28 cells (Fig. 4D).
LCA regulates apoptotic and
anti-apoptotic molecules in MPM cells
To observe the molecular mechanisms of cell
apoptosis in MSTO-211H (Fig. 5A)
and H28 (Fig. 5B) cells, we
examined the expression of target proteins. First, after LCA
treatment (10, 20 and 40 μM) the expression of pro- and
anti-apoptotic proteins was investigated. As a result, BID,
Bcl-xL Caspase-3 and C-PARP expression were specifically
decreased and cleaved in MPM cell lines. On the other hand, Bax
expression of pro-apoptotic protein was increased. Multi-caspase
activity was increased in MPM cells treated with various
concentrations of LCA (10, 20 and 40 μM) for 48 h (Fig. 5E and F). Total multi-caspase
activity in MSTO-211H (Fig. 5E)
was 14.2±1.6, 28.1±3.4 and 81.3±0.7% that in H28 (Fig. 5F) was 16.7±0.5, 32.1±2.9 and
37.9±3.0% in a dose-dependent manner, respectively.
Discussion
MPM is a cancer that develops in the mesothelium
that covers many of the internal organs of the body (4). The most common anatomical location of
mesothelioma is the pleura and it is most commonly caused by
exposure to asbestos (26).
Currently, surgery and radiotherapy have been recommended, but the
prognosis is poor. Therefore, chemotherapy is gaining attention
(27).
LCA is a novel estrogenic flavonoid and a natural
phenol product. It can be derived from roots of Glycyrrhiza
inflata and shows beneficial activities such as anti-malarial,
anticancer, anti-bacterial and anti-viral effects (12,28).
There is a growing body of evidence that it exhibits an anticancer
effect, but it is not clear whether LCA would be effective against
MPM. In this study, we investigated whether LCA could induce
apoptosis via Sp1 in MPM cells (MSTO-211H and H28). First, MTS
assay was performed to assess how the LCA could affect cell
proliferation. Cell growth was inhibited in a
concentration-dependent manner by LCA. In addition, we treated LCE
(40 μM) bearing the similar allyl group on different positions of
methyl groups compared with LCA. In contrast to LCA, LCE was not
effective in MPM cells (21,29).
The different allyl group of LCA might be critical for its
anticancer activity against MPM. PI staining was performed to
demonstrate that LCA had an anti-proliferative effect and induced
apoptotic cell death. We checked the changes in percentage of
sub-G1 fraction. Percentage of cells present in
sub-G1 peak was increased. Annexin V staining and
caspase activity were measured to further verify apoptotic cell
death (30). Caspase activity as
well as the numbers of early and late apoptotic cells was
increased. Accordingly, it is evident that the cytostatic effect of
LCA in MPM cells is due to induction of cell death.
To prove that apoptosis is mediated by regulation of
Sp1 transcription factor, we examined the apoptosis-related factors
using western blotting. Sp1 is a zinc finger transcription factor
that binds to GC-rich motifs of many promoters (23) and it is involved in many cellular
processes, including cell differentiation, cell growth, apoptosis,
immune responses, response to DNA damage and chromatin remodeling
(18). LCA downregulated
expression levels of mRNA and protein level of the Sp1 in a
concentration and time-dependent manner. Consistent with Sp1
levels, Sp1 regulatory proteins (Cyclin D1, Mcl-1 and Survivin)
were also diminished, hypothesizing that the LCA induced cell death
via downregulation of Sp1.
Pathologically, apoptosis is referred to as the
normal process of active cell death of organisms to remove damaged
or unnecessary cells. Mitochondria not only produced energy in
cells but also regulates cell fate factors (31,32).
In this study, we revealed how LCA could be involved in the
morphological control of mitochondria in MPM cells and further
delineated the mechanisms associated with the induction of cell
death.
Apoptosis involves two different pathways. One is
the extrinsic pathway of caspase activation that is mediated by the
cell surface death receptor and the other is the mitochondria
initiated pathway (33,34). Extrinsic pathway begins with the
binding of ligands to a subset of the members of the TNF receptor
superfamily (35–37). TRAIL is able to interact with the
membrane receptor DR4/TRAIL-R1 and DR5/TRAIL-R2 bearing a different
functional death domain (38,39).
An increase in expression of CHOP elevates the levels of membrane
receptors DR4 and DR5, which operate cleavage of Bid into tBid.
Cytochrome C induces the activation of caspase-8 and -9, leading to
downstream activation of caspase-3 to promote the final cell death
mechanism (35). We found that the
expression of CHOP, DR4 and DR5 was upregulated while the levels of
both caspase-3 and Bid decreased in the LCA-treated MPM cells.
These results suggest that LCA may be able to induce apoptosis in
MPM cells via the receptor-mediated pathway. The second pathway of
apoptosis involves the participation of mitochondria, regulated by
the anti-apoptotic and pro-apoptotic members of the Bcl-2 family
(40). Bcl-2 family is composed of
pro-apoptotic proteins such as Bax, Bak and Bid and anti-apoptotic
proteins such as Bcl-2, Bcl-xL and Mcl-1. Bax or Bak are
bound to the mitochondria membrane, releasing cytochrome C into
cytosol (41,42). The released cytochrome C in turn
activates caspases, which transmit cell death signals by dividing
PARP (43–46). Decreased expression of the
cytosolic Bax and concomitant increase of the mitochondrial Bax
indicated that LCA induced the translocation of Bax from cytosol to
the mitochondrial membrane. Western blot results demonstrated that
LCA upregulated the expression level of Bax and C-PARP and
downregulated the level of Bcl-2 in MPM cells. In combination with
the effect of LCA on mitochondrial membrane potential in MPM cells,
these results indicate that LCA induces apoptosis in MPM cells via
the mitochondria-mediated intrinsic pathway.
Taken together, this study reveals the relevance of
Sp1, MPM, LCA and mitochondria to apoptosis. We demonstrated that
LCA induced apoptosis by suppressing the expression of Sp1 in MPM
cells. LCA-mediated Sp1 down-regulation induced morphological
changes of mitochondria, leading to apoptotic cell death.
Therefore, our studies strongly suggest a potential novel strategy
to use LCA as an effective anticancer therapeutic agent for MPM
patients with Sp1 overexpression.
Acknowledgements
This study was supported by Gloden Seed Project
(2130051), Ministry of Agriculture, Food and Rural Affairs.
Abbreviations:
|
MPM
|
malignant pleural mesothelioma
|
|
LCA
|
licochalcone A
|
|
Sp1
|
specificity protein 1
|
|
RPMI-1640
|
Roswell Park Memorial Institute
|
|
FBS
|
fetal bovine serum
|
|
PBS
|
phosphate-buffered saline
|
|
Mcl-1
|
myeloid cell leukemia-1
|
|
PARP
|
poly(ADP-ribose)
polymerase;MTS,(3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxy-phenyl)-2-(4-sulfophenyl)-2H-tetrazolium)
|
|
DAPI
|
4′-6-diamidino-2-phenylindole
|
|
PI
|
propidium iodide
|
|
LCE
|
licochalcone E
|
|
TNF
|
tumor necrosis factor
|
|
TRAIL
|
TNF-related apoptosis-inducing
ligand
|
References
|
1
|
Siegel R, Ma J, Zou Z, et al: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tiwari RC, Ghosh K, Jemal A, et al: A new
method of predicting US and state-level cancer mortality counts for
the current calendar year. CA Cancer J Clin. 54:30–40. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ahmed I, Ahmed Tipu S and Ishtiaq S:
Malignant mesothelioma. Pak J Med Sci. 29:1433–1438. 2013.
|
|
4
|
Linton A, Cheng YY, Griggs K, et al: An
RNAi-based screen reveals PLK1, CDK1 and NDC80 as potential
therapeutic targets in malignant pleural mesothelioma. Br J Cancer.
110:510–519. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cunha P, Luz Z, Seves I, et al: Malignant
peritoneal mesothelioma - diagnostic and therapeutic difficulties.
Acta Med Port. 15:383–386. 2002.(In Portuguese).
|
|
6
|
de Pangher Manzini V: Malignant peritoneal
mesothelioma. Tumori. 91:1–5. 2005.PubMed/NCBI
|
|
7
|
Kusamura S, Deraco M, Baratti D, et al:
Cytoreductive surgery followed by intra peritoneal hyperthermic
perfusion in the treatment of peritoneal surface malignancies:
morbidity and mortality with closed abdomen technique. J Exp Clin
Cancer Res. 22:207–212. 2003.
|
|
8
|
Rodriguez D, Cheung MC, Housri N, et al:
Malignant abdominal mesothelioma: defining the role of surgery. J
Surg Oncol. 99:51–57. 2009. View Article : Google Scholar
|
|
9
|
Yuan X, Li D, Zhao H, et al: Licochalcone
A-induced human bladder cancer T24 cells apoptosis triggered by
mitochondria dysfunction and endoplasmic reticulum stress. Biomed
Res Int. 2013:4742722013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yao K, Chen H, Lee MH, et al: Licochalcone
A, a natural inhibitor of c-Jun N-terminal kinase 1. Cancer Prev
Res. 7:139–149. 2014. View Article : Google Scholar
|
|
11
|
Kim JK, Shin EK, Park JH, et al: Antitumor
and antimetastatic effects of licochalcone A in mouse models. J Mol
Med. 88:829–838. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xiao XY, Hao M, Yang XY, et al:
Licochalcone A inhibits growth of gastric cancer cells by arresting
cell cycle progression and inducing apoptosis. Cancer Lett.
302:69–75. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yo YT, Shieh GS, Hsu KF, et al: Licorice
and licochalcone-A induce autophagy in LNCaP prostate cancer cells
by suppression of Bcl-2 expression and the mTOR pathway. J Agric
Food Chem. 57:8266–8273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cui Y, Ao M, Li W, et al:
Anti-inflammatory activity of licochalcone A isolated from
Glycyrrhiza inflata. Z Naturforsch C. 63:361–365. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Funakoshi-Tago M, Tanabe S, Tago K, et al:
Licochalcone A potently inhibits tumor necrosis factor
alpha-induced nuclear factor-kappaB activation through the direct
inhibition of IkappaB kinase complex activation. Mol Pharmacol.
76:745–753. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li L and Davie JR: The role of Sp1 and Sp3
in normal and cancer cell biology. Ann Anat. 192:275–283. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Safe S and Abdelrahim M: Sp transcription
factor family and its role in cancer. Eur J Cancer. 41:2438–2448.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Courey AJ and Tjian R: Analysis of Sp1 in
vivo reveals multiple transcriptional domains, including a novel
glutamine-rich activation motif. Cell. 55:887–898. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chuang JY, Wu CH, Lai MD, et al:
Overexpression of Sp1 leads to p53-dependent apoptosis in cancer
cells. Int J Cancer. 125:2066–2076. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yoon G, Jung YD and Cheon SH: Cytotoxic
allyl retrochalcone from the roots of Glycyrrhiza inflata. Chem
Pharm Bull. 53:694–695. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fu Y, Chen J, Li YJ, et al: Antioxidant
and anti-inflammatory activities of six flavonoids separated from
licorice. Food Chem. 141:1063–1071. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Goldberg JL, Zanella CL, Janssen YM, et
al: Novel cell imaging techniques show induction of apoptosis and
proliferation in mesothelial cells by asbestos. Am J Respir Cell
Mol Biol. 17:265–271. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bouwman P and Philipsen S: Regulation of
the activity of Sp1-related transcription factors. Mol Cell
Endocrinol. 195:27–38. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen Q, Liu XF and Zheng PS: Grape seed
proanthocyanidins (GSPs) inhibit the growth of cervical cancer by
inducing apoptosis mediated by the mitochondrial pathway. PLoS One.
9:e1070452014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Condamine T, Kumar V, Ramachandran IR, et
al: ER stress regulates myeloid-derived suppressor cell fate
through TRAIL-R-mediated apoptosis. J Clin Invest. 124:2626–2639.
2014. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gultekin KE, Yurdakonar MK, Yaman E, et
al: Effects of cisplatin and panobinostat on human mesothelial
(Met-5A) and malignant pleural mesothelioma (MSTO-211H) cells.
Genet Mol Res. 12:5405–5413. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee YJ, Park IS, Lee YJ, et al:
Resveratrol contributes to chemo-sensitivity of malignant
mesothelioma cells with activation of p53. Food Chem Toxicol.
63:153–160. 2013. View Article : Google Scholar
|
|
28
|
Tsai JP, Hsiao PC, Yang SF, et al:
Licochalcone A suppresses migration and invasion of human
hepatocellular carcinoma cells through downregulation of MKK4/JNK
via NF-kappaB mediated urokinase plasminogen activator expression.
PLoS One. 9:e865372014. View Article : Google Scholar
|
|
29
|
Park HG, Bak EJ, Woo GH, et al:
Licochalcone E has an antidiabetic effect. J Nutr Biochem.
23:759–767. 2012. View Article : Google Scholar
|
|
30
|
Chang SF, Chang CA, Lee DY, et al: Tumor
cell cycle arrest induced by shear stress: Roles of integrins and
Smad. Proc Natl Acad Sci USA. 105:3927–3932. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li H, Zhu H, Xu CJ, et al: Cleavage of BID
by caspase 8 mediates the mitochondrial damage in the Fas pathway
of apoptosis. Cell. 94:491–501. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kaufmann T, Strasser A and Jost PJ: Fas
death receptor signalling: roles of Bid and XIAP. Cell Death
Differ. 19:42–50. 2012. View Article : Google Scholar :
|
|
33
|
Teng CL, Han SM, Wu WC, et al: Mechanistic
aspects of lauryl gallate-induced differentiation and apoptosis in
human acute myeloid leukemia cells. Food Chem Toxicol. 71:197–206.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Law CK, Kwok HH, Poon PY, et al:
Ginsenoside compound K induces apoptosis in nasopharyngeal
carcinoma cells via activation of apoptosis-inducing factor. Chin
Med. 9:112014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fossati S, Ghiso J and Rostagno A: TRAIL
death receptors DR4 and DR5 mediate cerebral microvascular
endothelial cell apoptosis induced by oligomeric Alzheimer’s Abeta.
Cell Death Dis. 3:e3212012. View Article : Google Scholar
|
|
36
|
Haase G, Pettmann B, Raoul C, et al:
Signaling by death receptors in the nervous system. Curr Opin
Neurobiol. 18:284–291. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wilson NS, Dixit V and Ashkenazi A: Death
receptor signal transducers: nodes of coordination in immune
signaling networks. Nat Immunol. 10:348–355. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rizzardi C, Leocata P, Ventura L, et al:
Apoptosis-related factors (TRAIL, DR4, DR5, DcR1, DcR2, apoptotic
cells) and proliferative activity in ameloblastomas. Anticancer
Res. 29:1137–1142. 2009.PubMed/NCBI
|
|
39
|
Almasan A and Ashkenazi A: Apo2L/TRAIL:
apoptosis signaling, biology and potential for cancer therapy.
Cytokine Growth Factor Rev. 14:337–348. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Martin SJ and Green DR: Protease
activation during apoptosis: death by a thousand cuts? Cell.
82:349–352. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jin Z and El-Deiry WS: Overview of cell
death signaling pathways. Cancer Biol Ther. 4:139–163. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kim EJ, Lee YJ, Shin HK, et al: Induction
of apoptosis by the aqueous extract of Rubus coreanum in HT-29
human colon cancer cells. Nutrition. 21:1141–1148. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Oliver FJ, de la Rubia G, Rolli V, et al:
Importance of poly(ADP-ribose) polymerase and its cleavage in
apoptosis. Lesson from an uncleavable mutant. J Biol Chem.
273:33533–33539. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wong WW and Puthalakath H: Bcl-2 family
proteins: the sentinels of the mitochondrial apoptosis pathway.
IUBMB Life. 60:390–397. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
45
|
El-Hamoly T, Hegedus C, Lakatos P, et al:
Activation of poly(ADP-ribose) polymerase-1 delays wound healing by
regulating keratinocyte migration and production of inflammatory
mediators. Mol Med. 20:363–371. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Toth-Zsamboki E, Horvath E, Vargova K, et
al: Activation of poly(ADP-ribose) polymerase by myocardial
ischemia and coronary reperfusion in human circulating leukocytes.
Mol Med. 12:221–228. 2006. View Article : Google Scholar
|