Introduction
Metastatic chondrosarcoma of mesenchymal origin is
the second most common bone malignancy and does not respond either
to chemotherapy or radiation; therefore, the search for new
therapies is relevant and urgent (1–4).
We have described recently that tumor growth
inhibiting cytostatic proline-rich polypeptide 1, PRP-1 (galarmin),
statistically significantly upregulated tumor suppressor miRNAs,
downregulated onco-miRNAs in human chondrosarcoma JJ012 cell line,
compared to chondrocyte culture (5). Among significantly downregulated
oncomiRNAs in chondrosarcoma JJ012 cell line after the treatment
with this peptide was miR302c, downregulated 6.46-fold. miR302-367
cluster has been described to be differentially expressed in mouse
embryonic stem cells (mESCs) and human embryonic stem cells (hES
cells), as well as in certain tumors, but not in adult mesenchymal
stem cells (hMSCs) and normal tissues (6). The gene coding for the cluster
miR302-367 is a Pol II gene with a capped and polyadenylated
transcription product. The miR302-367 coding gene contains three
exons and two introns with alternative splicing, which may or may
not include exon 2; the miRNA cluster is located within the first
intron. Eight miRNA loci (miR-302b, miR-302b*, miR-302c,
miR-302c*, hsc-3, miR-302a*, miR-302d, and
miR-367) are located within an ~700-bp region on chromosome 4. Cell
biology data indicated that the miR302-367 promoter activity
depends on the ontogeny and hierarchical cellular stage. The
promoter activity is functional during embryonic development
(hESCs, mESCs, and hECCs), but it is turned off later in
development (hMSCs and multiple transformed cell lines). Even
though the promoter transcriptional activity is restricted to an
embryonic stage, the promoter transcriptional activity was reported
to be off in not only adult stem cells (hMSCs) but also embryonic
cell lines with no stem cell potential (7,8).
Emerging evidence suggests that miRNAs also may play an essential
role in stem cell self-renewal and differentiation (9). The expression of the miR302/367
cluster rapidly and efficiently reprograms mouse and human somatic
cells to an iPSC state without a requirement for exogenous
transcription factors (10,11).
To maintain self-renewal and pluripotency stem cells have to
prevent differentiation and development. It is thought that
overexpressed miRNAs from the miR302/367 cluster in stem cells
primarily repress development. This study pursued the
identification of functional marker in cancer stem cells, mechanism
of its epigenetic regulation, correlated to selective
antiproliferative activity of PRP-1.
Materials and methods
Human chondrosarcoma JJ012 cell line
tissue culture
The chondrosarcoma cells JJ012 were cultured in
monolayer. The chondrosarcoma cell line JJ012 was provided by Dr
Joel A. Block (Department of Rheumatology, Rush-Presbyterian St.
Luke’s Medical Center, Chicago, IL, USA), then cultured and
propagated in our laboratory. Media consisted of Dulbecco’s
modified Eagle’s medium (DMEM/MEM), supplemented with F12, 10%
fetal bovine serum (ATCC), 25 μg/ml ascorbic acid, 100 ng/ml
insulin, 100 nM hydrocortisone, and 1% penicillin/streptomycin
(Sigma-Aldrich). The cells were trypsinized and used either for the
rapid cell proliferation assays or for the mRNA extraction and
miRNA arrays. The control samples were not treated with the
peptide, whereas 10 μg/ml PRP-1 was added to the experimental
series. PRP-1 was synthesized in our laboratory.
Rapid cell proliferation assay
Cell Proliferation kit, EMD Biosciences (QIA127),
was used for this assay. Cells were seeded at 5×104
cells/100 μl culture in the multi-well plate and incubated
overnight at 37°C in 5% CO2 incubator. The actual assay
was performed the next day. The rapid cell proliferation assay is
based on the activity of mitochondrial enzymes active in viable
cells. PRP-1 was added to corresponding wells just after seeding,
before overnight incubation. The colorimetric 96-well assay
measures the colorful product formazan formed by WST-1 tetrazolium
salt cleavage by the mitochondrial dehydrogenases. The formazan
formation was then quantified by measuring the change in absorbance
at 450 nm in a microplate reader. The activity of mitochondrial
dehydrogenases is proportional to the cell number. No washing,
harvesting, or solubilization steps are required. In this series of
experiments, PRP-1 was added to corresponding wells immediately
after seeding and cell attachment followed by overnight incubation
according to the manufacturer’s instructions.
Glioblastoma cellular ATP assay
A-172, T98G, and U87MG glioblastoma cell lines
(purchased from ATCC) were cultured in Dulbecco’s modified Eagle’s
medium supplemented with 10% fetal bovine serum, 100 U/ml
penicillin and 100 μg/ml streptomycin (Life Technologies). Cells
were grown for 24 h at a density of 500 cells per well in 384-well
plates. IBET-151, PRP peptide, or vehicle controls (DMSO for
IBET-151, saline for PRP) were added directly into the wells. Each
condition was tested in triplicate. After 72 h, a CellTiter-Glo
Luminescent Cell Viability assay (Promega) was performed according
to the manufacturer’s recommendations. Briefly, the CellTiter-Glo
reagents were added to the wells and the plate was briefly
centrifuged. The plate was incubated 15 min at room temperature,
and luminescence was read using an EnVision Multilabel plate reader
(Perkin-Elmer). Dose-response curves were analyzed in GraphPad
prism and fitted using a non-linear regression analysis.
Gel electrophoresis and western
blotting
Upon confluency, the cells were trypsinized and
seeded in 6-well cluster dishes at a concentration of
1×106 cells/ml. The experimental samples were treated
with PRP-1 in corresponding concentrations, whereas control samples
were not treated with the peptide. The cells were incubated for 24
h in a 5% CO2 incubator at 37°C. The following day, the
cells were washed with ice-cold phosphate-buffered saline. A
protease inhibitor was added to the cell lysis buffer (C2978;
Sigma-Aldrich, St. Louis, MO, USA) in a 1:100 ratio. The cells were
collected with a scraper and centrifuged at 15, 000 × g at 4°C. The
supernatant was collected and the protein concentration was
measured. The pellets were frozen at −80°C until loading on the gel
(20 μg/lane). Polyacrylamide gel electrophoresis and western
blotting reagents were supplied by Lonza, Inc. (Allendale, NJ,
USA), and all the related procedures followed the company’s
protocol. The catalog numbers for the reagents and the suppliers
are listed below.
Pager Gold Precast Gels (59502; 10% Tris-Glycine;
Lonza, Inc.); ECL reagent (RPN2109; GE Healthcare, Little Chalfont,
UK); Western Blocker solution (W0138; Sigma-Aldrich); ProSieve Quad
Color Protein marker (4.6-300 kDa, 00193837; Lonza, Inc.); 20×
reducing agent for ProSieve ProTrack Dual Color Loading buffer
(00193861; Lonza, Inc.); ProTrack Loading buffer (00193861; Lonza,
Inc.); ProSieve ProTrack Dual Color Loading buffer EX running
buffer (00200307; Lonza, Inc.); ProSieve EX Western Blot Transfer
buffer (00200309; Lonza, Inc.); Immobilon®-P
Polyvinylidene difluoride membranes (P4188; Sigma-Aldrich).
Culture of marrow-isolated adult
multilineage-inducible (MIAMI) cells
MIAMI cells were grown as previously described (REF:
PMID: 15173316). Briefly, whole BM cells were plated at
1×105/cm2 in T75 flasks (Costar) in the
presence of D-MEM low glucose, 3% FBS, 100 U/ml penicillin (Gibco),
1 mg/ml streptomycin (Gibco). The cells were incubated at 37°C in a
100% humidified atmosphere of 3% O2, 5% CO2,
and 92% N2. Half of the medium was changed after a week;
thereafter, half the medium was replaced twice a week. MIAMI cells
were cultured to 40–50% confluence. For expansion, MIAMI cells were
replated at a density of 100 cells/cm2 in
fibronectin-coated vessels in 95% D-MEM-low glucose, 3%
lot-selected FBS, and 100 U penicillin/1,000 U streptomycin
(expansion medium) at 3% O2, with 50% of the medium
changed twice a week.
MIAMI cell growth assay with PRP-1
MIAMI cells were plated in triplicate at 2,000
cells/cm2 in 6-well plates in expansion medium. The
following day cells were supplemented with 1, 2, and 10 μg/ml
PRP-1. At the end of each assay-day, 7, 10, and 14, 21 cells were
rinsed with PBS and detached with trypsin-EDTA, and then counted
with a Neubauer hemacytometer chamber.
Antibodies
Primary: rabbit polyclonal anti-CDK/2 (M2), cat no.
sc-1632 Santa Cruz Biotechnologies; mouse monoclonal (9E10)
anti-c-Myc, cat no. SC-40 Santa Cruz Biotechnologies; rabbit
polyclonal anti-p-c-Myc (Thr58/Ser 62), cat no. Sc-8000R, Santa
Cruz Biotechnologies; mouse monoclonal anti-SCML2 (SCMAD14a), cat
no. ab51506 Abcam; rabbit polyclonal antip-Src (Tyr416), cat no.
2101S, Cell Signaling; rabbit polyclonal anti-Src antibody, cat no.
2108S, Cell Signaling, rabbit monoclonal anti-p27 Kip1 (D69C12)
XP® cat no. 3686; Santa Cruz Biotechnology; mouse
monoclonal p21 (F-5), cat no. sc-6246, Santa Cruz Biotechnologies;
mouse monoclonal anti Nanog, clone 7F7-1, cat no. MABD24, EMD
Millipore; rabbit polyclonal anti-Bmi-1 antibody, cat no. ab38295,
Abcam. Mouse monoclonal anti-tubulin, cat no. T5168, Sigma.
Secondary: anti-mouse IgG (A9044; Sigma-Aldrich); and goat
anti-rabbit IgG peroxidase conjugate (A0545; Sigma-Aldrich).
Statistical analysis
All experiments were performed in triplicate, and
P<0.05 was considered statistically significant. Data analysis
was perform using one-way analysis of variance (ANOVA) unpaired
t-test (GraphPad Prism; GraphPad Software, San Diego, CA, USA).
Results
Comparison of antiproliferative activity
of PRP-1 in human JJ012 chondrosarcoma cell line, glioblastoma cell
lines and marrow-isolated adult multilineage inducible (MIAMI)
cells
This study pursued the identification of functional
marker in cancer stem cells, correlated to peptides
antiproliferative activity by comparing different cell lines, where
miR302-367 cluster either induces or inhibits stemness properties,
as well as marrow-isolated adult multilineage inducible (MIAMI)
cells that express embryonic stem cell markers. We have previously
demonstrated antiproliferative effect of PRP-1 reaching 80–90%
inhibition in human chondrosarcoma cells (2,4).
PRP peptide does not inhibit the growth
or viability of glioblastoma cells
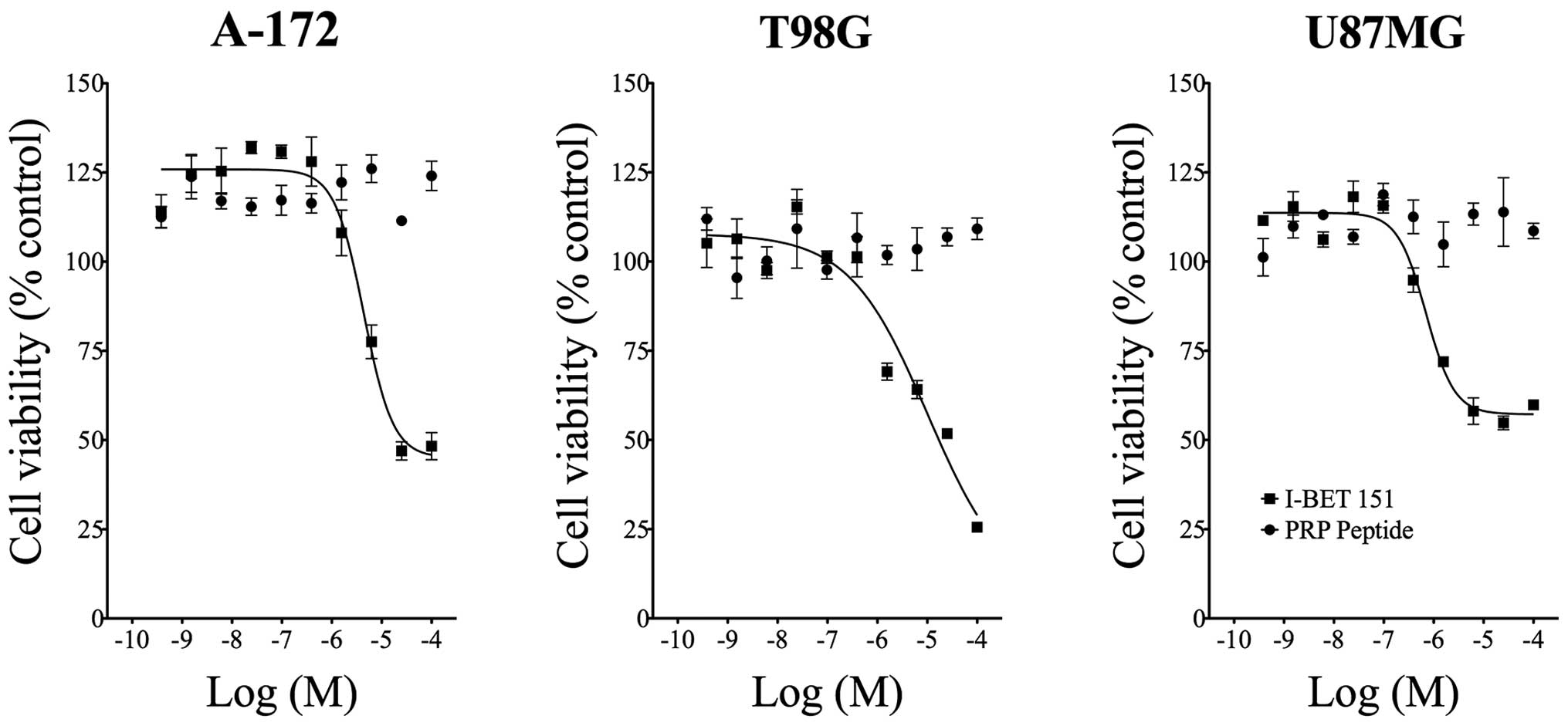
The BET bromodomain inhibitor IBET-151 reduced
glioblastoma cellular ATP levels with potency similar to that we
previously reported (IC50 = 4.8 μM in A-172 cells, 9.3
μM in T98G cells, and 0.764 μM in U87MG cells) (12,13).
In contrast, the PRP peptide showed no effect in any cell line
tested, indicating this compound does not reduce glioblastoma
cellular proliferation or viability (Fig. 1).
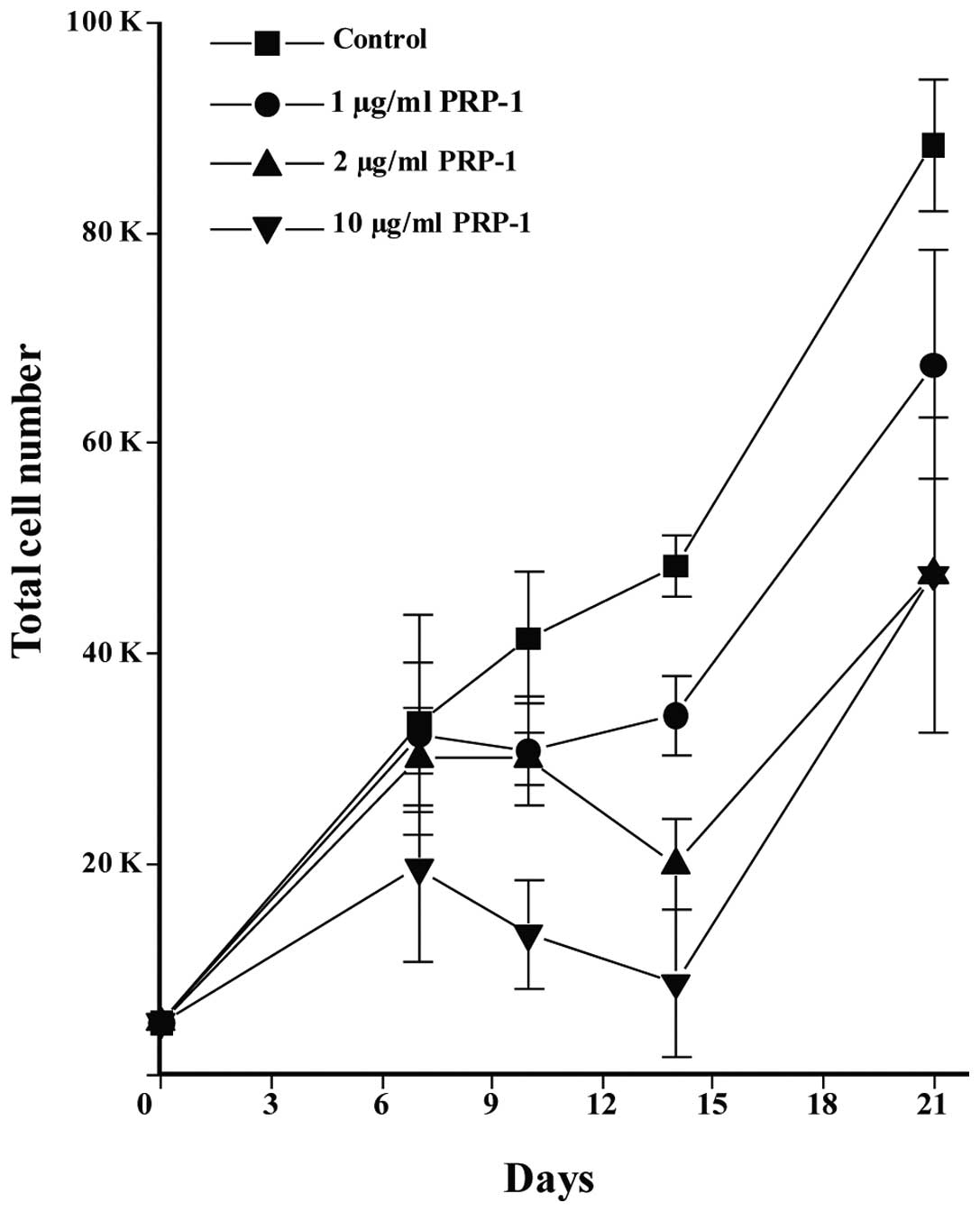
PRP-1 inhibits MIAMI cells
MIAMI cells resemble primitive stem cells in their
capacity to differentiate, at least in vitro into
mature-like cells from all three germ layers. The expression of
embryonic stem cell markers indicate the developmentally immature
status of MIAMI cells (14,15).
Therefore, it comes as no surprise that the peptide inhibited the
growth of these cells. The dose-response inhibitory effect of
PRP-1, reaching maximum at 10 μg/ml of the peptide in comparison to
untreated control cells is depicted in Fig. 2.
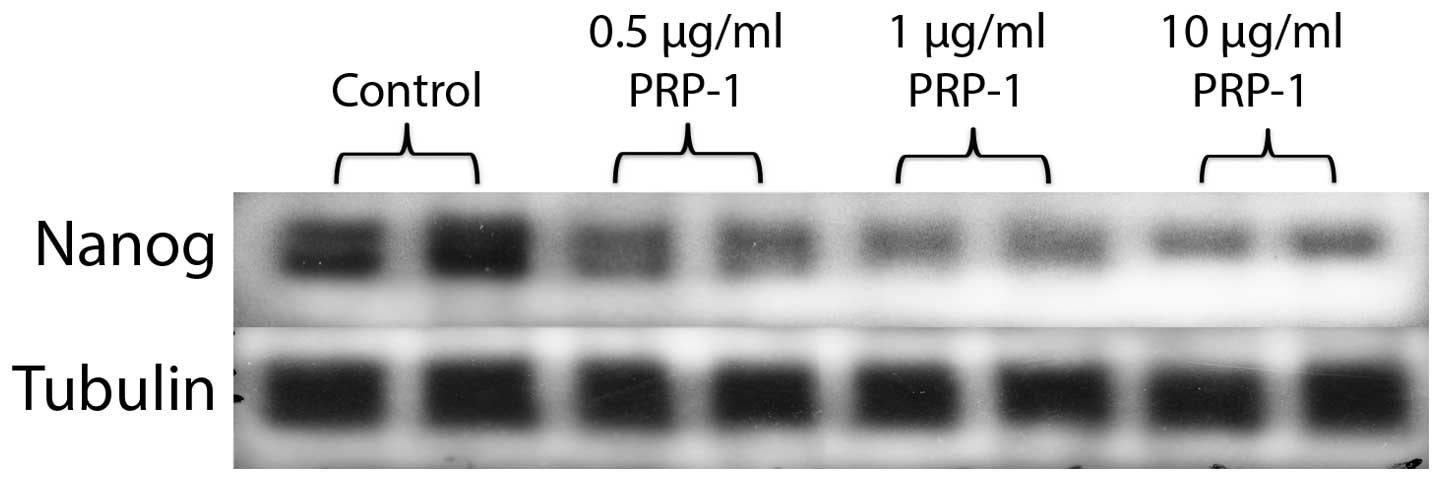
PRP-1 attenuated the expression of the
miR302-367 targets the embryonic stem cell marker Nanog and
polycomb protein Bmi-1, while increasing SCML2 expression
levels
The embryonic stem cell marker Nanog is one of the
targets for miR302-367 cluster and it is expressed in many cancers.
Nanogs expression was substantially decreased in human JJ012
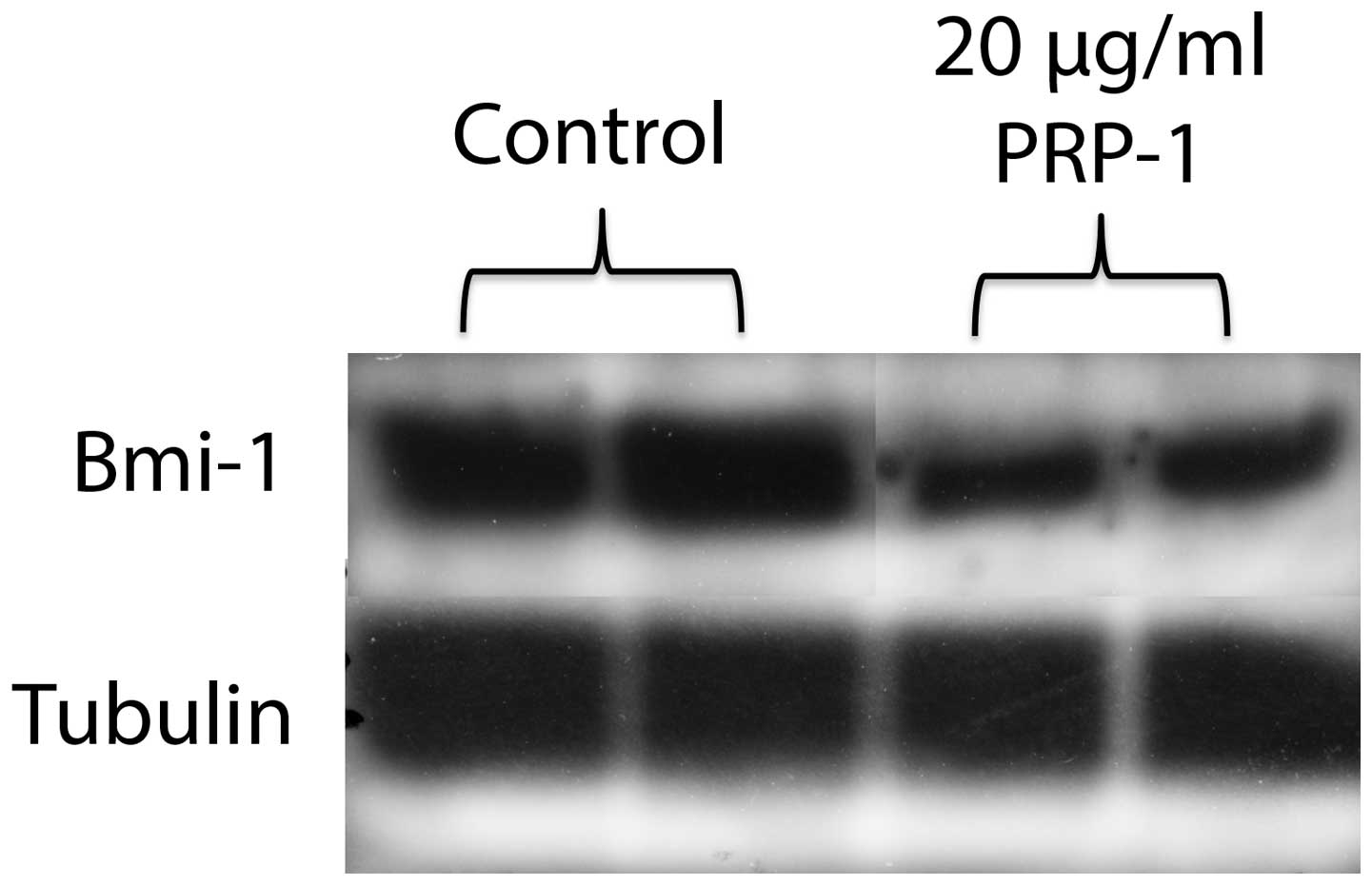
chondrosarcoma cell line after the treatment with PRP-1 (Fig. 3). The polycomb protein Bmi-1 is
also a target for the miR302-367 cluster. Treatment with PRP-1 (20
μg/ml) resulted in strong attenuation of Bmi-1 expression level in
comparison to untreated control. Tubulin is demonstrated here as
housekeeping protein (Fig. 4). On
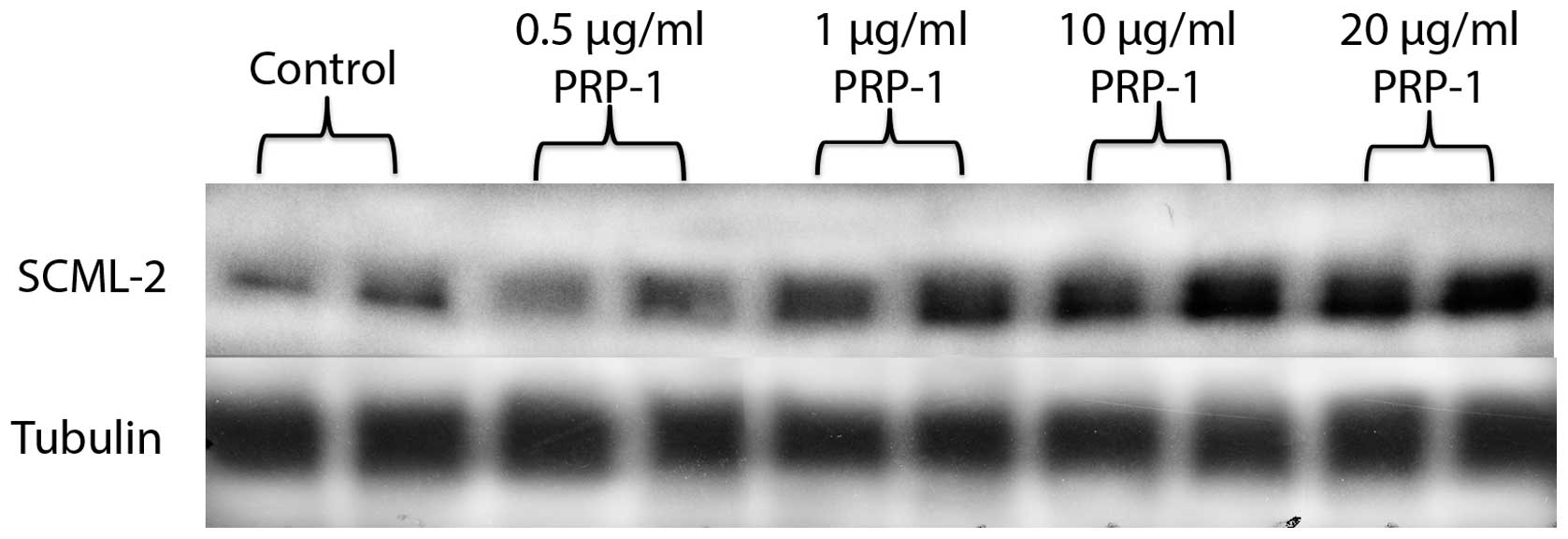
the contrary, SCML2 expression was increased by PRP-1 in a
dose-response manner. SCML2 is not a direct target for miR302-367
cluster, but it is known to repress transcription and is considered
as tumor suppressor (Fig. 5).
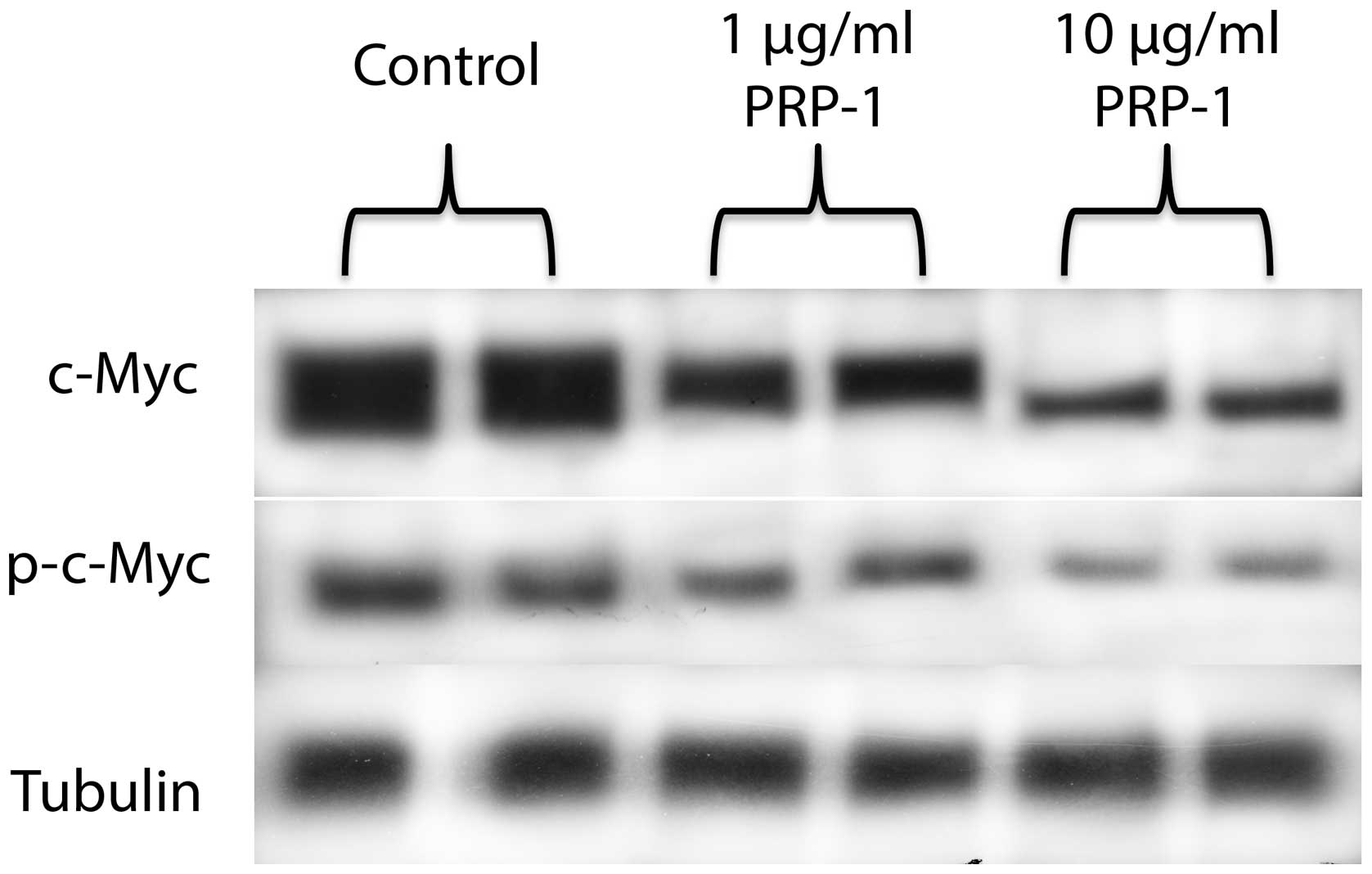
PRP-1 decreased c-Myc, p-c-Myc and Src,
but not p-Src levels
Western blot analysis revealed that PRP-1 reduced
c-Myc (oncogene target for miR302c) and phosphorylated p-c-Myc
expression (Fig. 6).
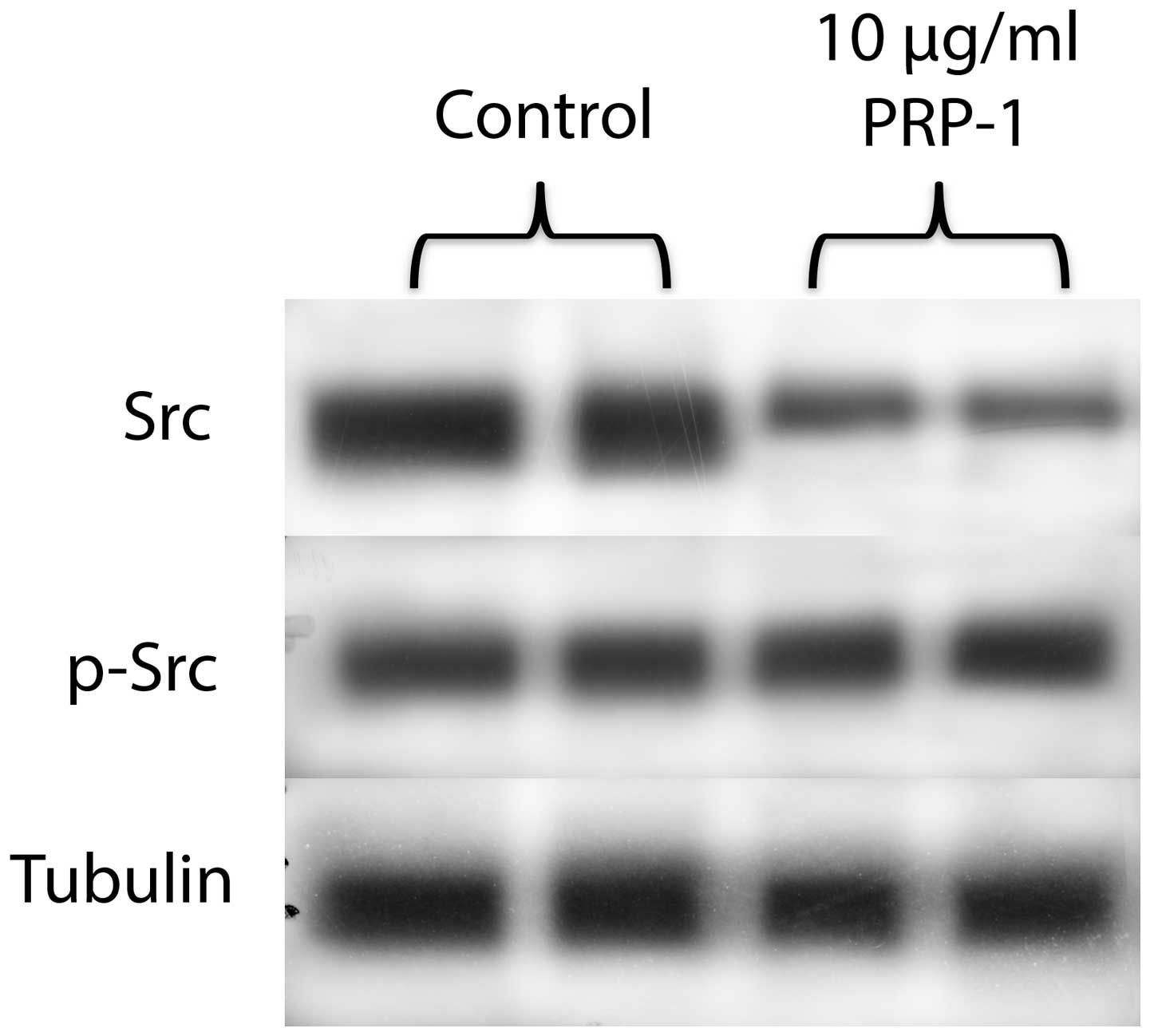
The peptide was tested for its effect on the other
oncogene, Src (albeit, its not the target for miR302c) and its
phosphorylated form. PRP-1 decreased Src protein levels, but not
p-Src expression (Fig. 7).
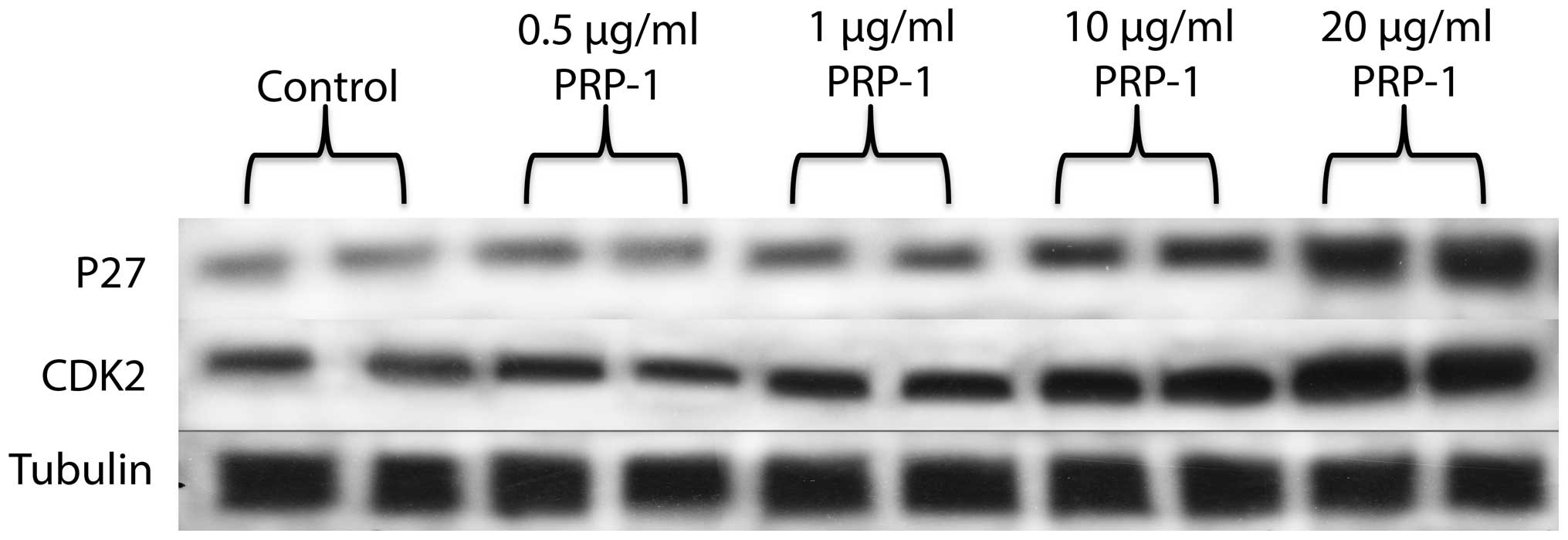
PRP-1 effect on cell cycle regulatory
proteins p27, p21 and CDK2
It was important to check the expression of cell
cycle regulatory proteins with or without the peptide treatment.
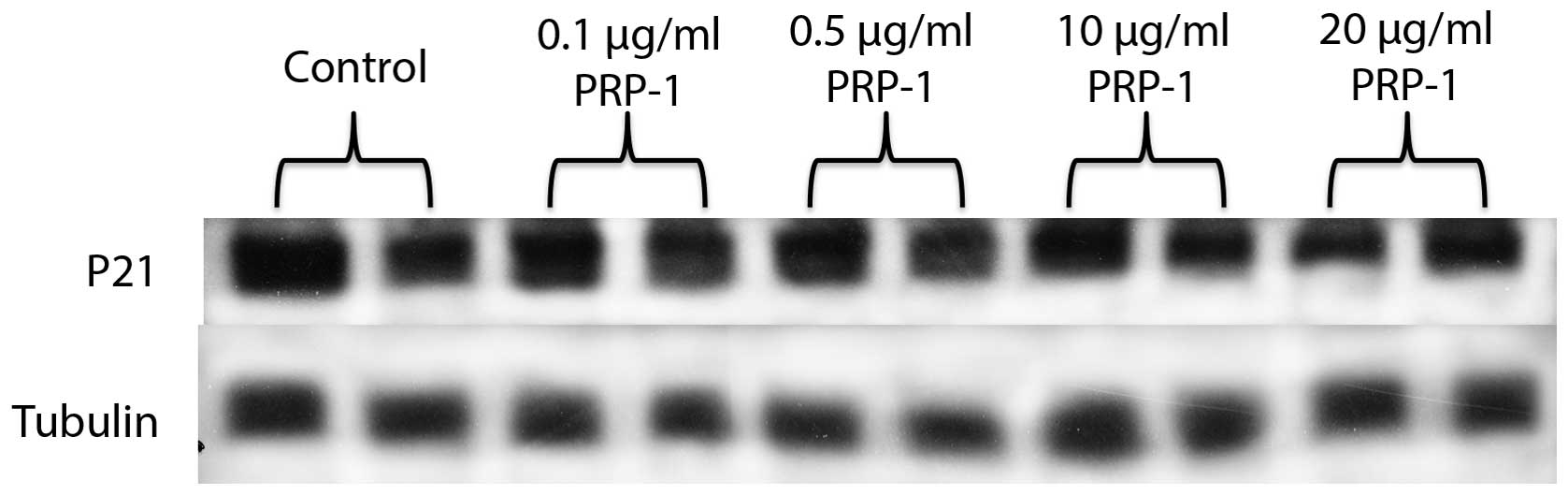
PPP-1 increased the expression levels of p27, and CDK2 (Fig. 8). P21 expression was reduced after
the treatment with PRP-1 in a dose-response manner (Fig. 9).
Discussion
In the present study, we have demonstrated that
miR302c, part of miR302-367 cluster, downstream factor of the
embryonic stem cell regulation network determined the
antiproliferative activity of cytostatic PRP-1. The cluster is
known as a potential stemness regulator in human embryonic stem
cells and its expressed in certain tumors but not in adult hMSC and
normal cells. Metastatic chondrosarcoma of mesenchymal origin is
the second most common bone malignancy and does not respond either
to chemotherapy or radiation; therefore, the search for new
therapies is relevant and urgent. We described recently that tumor
growth inhibiting proline-rich polypeptide 1 (PRP-1, galarmin)
significantly upregulated tumor suppressor miRNAs, downregulated
onco-miRNAs in human chondrosarcoma JJ012 cell line, compared to
chondrocyte culture. Among the miRNAs was miRNA302c*
6.46-fold downregulated after PRP-1 (10 μM) treatment (5). This study pursued the identification
of the functional marker in cancer stem cells, correlated to
peptide antiproliferative activity and understanding the epigenetic
regulation underlying its effect.
We have reported that inhibition of human
chondrosarcoma cells, including primary and JJ012 cells with PRP-1
reached >80% (16). In
contrast, three cell lines of glioblastoma were not affected by
PRP-1 treatment (Fig. 1).
Glioblastoma multiforme (GBM) is the most common form of primary
brain tumor in adults, often characterized by poor survival. The
absence of antiproliferative effect of PRP-1 was expected, taking
into consideration that the miR-302-367 cluster is strongly induced
during serum-mediated stemness suppression in glioblastoma. Stable
miR-302-367 cluster expression is sufficient to suppress the
stemness, self-renewal, and cell infiltration within a host brain
tissue, through inhibition of the CXCR4 pathway. Furthermore,
inhibition of CXCR4 leads to the disruption of the sonic hedgehog
(SHH)-GLI-NANOG network, which is involved in self-renewal and
expression of the embryonic stem cell-like signature. miR302-367
cluster is able to efficiently trigger a cascade of inhibitory
events leading to the disruption of glioma initiating (GiCs)
stem-like cells and tumorigenic properties (17). PRP-1 inhibited unique subpopulation
of human stromal cells from bone marrow, termed marrow-isolated
adult multilineage inducible (MIAMI) cells in a dose- and
time-response manner (Fig. 2).
These cells are known as the developmentally immature cells with
embryonic stem cell markers expression (14,15).
They resemble primitive stem cells in their capacity to
differentiate at least in vitro into mature-like cells from
all three germ layers. The observed correlation between the
antiproliferative activity of PRP-1 and its effect on
downregulation of miR302c, as a stemness marker, explains also the
opposite effects of the peptides on the upregulation of
proliferation of adult mesenchymal stem cells (MSC), and 2-fold
inhibition of the proliferation of human bone giant-cell tumor
stromal cells (18).
Somatic cells reprogram to an embryonic stem cell
(ESC) comparable induced pluripotent stem (iPS) cell state upon
forced expression of exogenously delivered transcription factors,
however, expression of exogenous miR-302 cluster (without miR-367)
is efficient in attaining a fully reprogrammed iPS state.
Methyl-DNA binding domain protein 2, (MBD2), is an epigenetic
suppressor, blocking full reprogramming of somatic to iPS cells
through direct binding to Nanog promoter elements preventing
transcriptional activation. When miR-302 cluster (without miR367)
was overexpressed, significant increase in conversion of partial to
fully reprogrammed iPS cells by suppressing MBD2 expression,
thereby increasing Nanog expression was observed (19). PRP-1 inhibited Nanog expression as
miR302c target expression both in MIAMI cells (preliminary not
shown results) and in JJ012 chondrosarcoma human cells (Fig. 3). Nanog was not the only target for
this miRNA cluster to be decreased by PRP-1. Fig. 4 depicted decreased levels of Bmi-1
protein expression after the treatment with the peptide. Bmi-1 is a
component of the Polycomb group (PcG) multiprotein PRC1 complex, a
complex required to maintain the transcriptionally repressive state
of many genes, including Hox genes, throughout the development.
Bmi-1 can act as an oncogene that is particularly potent for the
initiation of cancer progression. Bmi-1 is a part of the pathways
that are deregulated during tumor development, and thus believed to
contribute to neoplastic proliferation and cancer stem cell renewal
(20–22). Tumor recurrence following treatment
remains a major clinical challenge. Evidence from xenograft models
and human trials indicates selective enrichment of
cancer-initiating cells (CICs) in tumors that survive therapy.
Together with recent reports showing that CIC gene signatures
influence patient survival, these studies predict that targeting
self-renewal, the key ‘stemness’ property unique to CICs, may
represent a new paradigm in cancer therapy. Downregulation of Bmi-1
inhibits the ability of colorectal CICs to self-renew, resulting in
the abrogation of their tumorigenic potential. Bmi-1 was
demonstrated to play a central part in self-renewal of colorectal
cancer initiating cells, as cancer cells were reliant on Bmi-1 to
sustain growth and clonal maintenance (23). PRP-1 increased the levels of the
polycomb protein SCML2 (Fig. 5).
Even though it is not a direct target for miR302-367 cluster, SCML2
interacts with ncRNAs through an RNA-binding region (RBR),
contributing to the recruitment of PRC1 to target genes and also
directly collaborates in gene repression, particularly repressing
transcription. The repressive properties of SCML2 are independent
of Bmi-1. SCML2 is thought to be a tumor suppressor (24). Upregulation of tumor suppressor
function by PRP-1 is a characteristic feature of this peptide
(5,25). Among other affected by PRP-1
miR302c targets was c-Myc. c-Myc is known to be sufficient to
activate the fraction increase of CSCs and to activate the ES
program (26). Previously, we have
reported Myc oncogene inactivating effect of PRP-1 in
chondrosarcoma in luciferase assay (2,27),
this time we were able to demonstrate with western blot experiments
that PRP-1 caused downregulation of p-c-Myc and c-Myc (Fig. 6), which indicated the involvement
of peptides in the posttranslational modification and stabilization
of c-Myc. The effect of PRP-1 was also tested for p-Src and Src
(though Src is not a direct target for miR302-367 cluster)
(Fig. 7). An elevated level of
c-Src tyrosine kinase activity is suggested to be linked to cancer
progression by promoting other signals. The evidence underlying
this hypothesis is largely based on the observation that both Src
protein levels and, to a greater degree, Src protein kinase
activity, are frequently elevated in human neoplastic tissues when
compared to adjacent normal tissues (28). Certain cancer cells have similar
properties to stem cells (29).
Both can avoid cell division stop signals and keep dividing to form
new cells. In some cases, a subset of cancer cells within some
tumors are cancer stem cells. In stem cells, miRNAs are required to
bypass the normal G1/S checkpoint for appropriate stem cell
renewal. We demonstrated in our previous study (4) that PRP regulates growth rates of
chondrosarcoma cell line, causing cells to accumulate in phase S.
In our experiments, PRP-1 increased p27 levels in a dose-response
manner (Fig. 8). p27 mediates
response to growth inhibitory cytokines, has important
antiproliferative role and induces differentiation (30). It is very possible that by inducing
p27 levels this peptide causes S phase delay, in the manner of
Gatifloxacin action in pancreatic cancer cell lines (31). Gatifloxacin did not induce
apoptosis but caused an arrest of cells in S and G2-phase in
pancreatic carcinoma cells, inducing p21 and p27. In out
experiments, however, p21 levels (Fig.
9) decreased upon the peptide treatment. This finding is in
accord with the literature data and can be applied to breast cancer
as well, where in rat mammary carcinogenesis increased expression
of p21(Cip1), associated with decreased expression of p27(Kip1) was
observed (32). Interestingly, the
cdk2 levels upon the peptide treatment also increased (Fig. 8). It is possible that PRP-1 causes
cytosolic mislocalization of p27 and CDK2, which secures its
antiproliferative properties similarly to the
anti-migratory/invasive effects resembling those of Nodal in
trophoblast cells (33). p27 can
still perform its antiproliferative function despite being
transported out to the cytosol because of the concomitant nuclear
export of CDK2. It is generally accepted that CDK2 promotes G1/S
transition by phosphorylating and thereby inactivating Rb,
resulting in the activation of E2F transcription factors, as these
events take place in the nucleus, cytoplasmic mislocalization of
CDK2 would render it inactive in promoting cell cycle progression.
Moreover, the epigenetic mechanisms, which are involved lead to
antiproliferative activity of PRP-1. miR302c expression was
reported to be induced by JMJD2 demethylase (34,35)
binding in its promoter region and reduces H3K9me2 methylation.
JMJD1C knockdown reduces miR-302 expression (36). JMJD1A and JMJD2C are critical
regulators of ES cells, their depletion leads to embryonic stem
cell differentiation, which is accompanied by a reduction in the
expression of embryonic stem cell-specific genes and an induction
of lineage marker genes. Our experimental results proved that PRP-1
strongly inhibited H3K9 activity, comprised of a pool of JMJD1 and
JMJD2 in human chondrosarcoma (25). Jmjd2c regulates the expression of
downstream effector Nanog through demethylation of H3K9Me3 at the
promoter regions of Tcl1, Tcfcp2l1, and Zfp57 and positively
regulates the expression of these pluripotency-associated genes.
Jmjd2c is required to reverse the H3K9Me3 marks at the Nanog
promoter region and consequently prevents transcriptional
repressors HP1 and KAP1 from binding (33). The presented experimental data
demonstrate that PRP-1 substantially downregulated miR302c targets,
stemness markers Nanog, c-Myc, and polycomb protein Bmi-1. We
conclude that inhibition of H3K9 activity by PRP-1 leads to
downregulation of miR302c and its targets, defining the PRP-1
antiproliferative role. The significance of the presented data
underlies the correlation between antiproliferative activity of
PRP-1 in human metastatic chondrosarcoma cells and other tumors
with the expression of stemness inducing miR302c* that
can be a predictive marker for this peptide antitumorigenic
activity, targeting cancer stem cells. Our future efforts will be
focusing on isolating PRP-1 receptors in cancer stem cells, which
is of great therapeutic importance, considering that PRP-1 can be
quantified in blood (37).
References
|
1
|
Ozaki T, Hillmann A, Lindner N, Blasius S
and Winkelmann W: Metastasis of chondrosarcoma. J Cancer Res Clin
Oncol. 122:625–628. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Galoian K, Temple TH and Galoyan A:
Cytostatic effect of the hypothalamic cytokine PRP-1 is mediated by
mTOR and cMyc inhibition in high grade chondrosarcoma. Neurochem
Res. 36:812–818. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Galoian KA, Temple HT and Galoyan AA:
mTORC1 inhibition and ECM-cell adhesion-independent drug resistance
via PI3K-AKT and PI3K-RAS-MAPK feedback loops. Tumour Biol.
33:885–890. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Galoian KA, Temple TH and Galoyan A:
Cytostatic effect of novel mTOR inhibitor, PRP-1 (galarmin) in MDA
231 (ER-) breast carcinoma cell line. PRP-1 inhibits mesenchymal
tumors. Tumour Biol. 32:745–751. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Galoian KA, Guettouche T, Issac B, Qureshi
A and Temple HT: Regulation of onco and tumor suppressor MiRNAs by
mTORC1 inhibitor PRP-1 in human chondrosarcoma. Tumour Biol.
35:2335–2341. 2014. View Article : Google Scholar
|
|
6
|
Suh MR, Lee Y, Kim JY, Kim SK, Moon SH,
Lee JY, Cha KY, Chung HM, Yoon HS, Moon SY, et al: Human embryonic
stem cells express a unique set of microRNAs. Dev Biol.
270:488–498. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Barroso-del Jesus A, Lucena-Aguilar G and
Menendez P: The miR-302-367 cluster as a potential stemness
regulator in ESCs. Cell Cycle. 8:394–398. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Barroso-delJesus A, Romero-López C,
Lucena-Aguilar G, Melen GJ, Sanchez L, Ligero G, Berzal-Herranz A
and Menendez P: Embryonic stem cell-specific miR302-367 cluster:
Human gene structure and functional characterization of its core
promoter. Mol Cell Biol. 28:6609–6619. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang B, Pan X and Anderson TA: MicroRNA:
A new player in stem cells. J Cell Physiol. 209:266–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Anokye-Danso F, Trivedi CM, Juhr D, Gupta
M, Cui Z, Tian Y, Zhang Y, Yang W, Gruber PJ, Epstein JA, et al:
Highly efficient miRNA-mediated reprogramming of mouse and human
somatic cells to pluripotency. Cell Stem Cell. 8:376–388. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim J, Chu J, Shen X, Wang J and Orkin SH:
An extended transcriptional network for pluripotency of embryonic
stem cells. Cell. 132:1049–1061. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dawson MA, Prinjha RK, Dittmann A,
Giotopoulos G, Bantscheff M, Chan WI, Robson SC, Chung CW, Hopf C,
Savitski MM, et al: Inhibition of BET recruitment to chromatin as
an effective treatment for MLL-fusion leukaemia. Nature.
478:529–533. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pastori C, Daniel M, Penas C, et al: BET
bromodomain proteins are required for glioblastoma cell
proliferation. Epigenetics. 9:611–620. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
D’Ippolito G, Howard GA, Roos BA and
Schiller PC: Sustained stromal stem cell self-renewal and
osteoblastic differentiation during aging. Rejuvenation Res.
9:10–19. 2006. View Article : Google Scholar
|
|
15
|
D’Ippolito G, Diabira S, Howard GA, Menei
P, Roos BA and Schiller PC: Marrow-isolated adult multilineage
inducible (MIAMI) cells, a unique population of postnatal young and
old human cells with extensive expansion and differentiation
potential. J Cell Sci. 117:2971–2981. 2004. View Article : Google Scholar
|
|
16
|
Galoian K, Scully S, McNamara G, Flynn P
and Galoyan A: Antitumorigenic effect of brain proline-rich
polypeptide-1 in human chondrosarcoma. Neurochem Res. 34:2117–2121.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fareh M, Turchi L, Virolle V, Debruyne D,
Almairac F, de-la-Forest Divonne S, Paquis P, Preynat-Seauve O,
Krause KH, Chneiweiss H, et al: The miR 302-367 cluster drastically
affects self-renewal and infiltration properties of
glioma-initiating cells through CXCR4 repression and consequent
disruption of the SHH-GLI-NANOG network. Cell Death Differ.
19:232–244. 2012. View Article : Google Scholar :
|
|
18
|
Chailakhyan RK, Gerasimov YV, Chailakhyan
MR and Galoyan AA: Proline-rich hypothalamic polypeptide has
opposite effects on the proliferation of human normal bone marrow
stromal cells and human giant-cell tumour stromal cells. Neurochem
Res. 35:934–939. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee MR, Prasain N, Chae HD, Kim YJ, Mantel
C, Yoder MC and Broxmeyer HE: Epigenetic regulation of NANOG by
miR-302 cluster-MBD2 completes induced pluripotent stem cell
reprogramming. Stem Cells. 31:666–681. 2013. View Article : Google Scholar
|
|
20
|
Molofsky AV, Pardal R and Morrison SJ:
Diverse mechanisms regulate stem cell self-renewal. Curr Opin Cell
Biol. 16:700–707. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu S, Dontu G, Mantle ID, Patel S, Ahn
NS, Jackson KW, Suri P and Wicha MS: Hedgehog signaling and Bmi-1
regulate self-renewal of normal and malignant human mammary stem
cells. Cancer Res. 66:6063–6071. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang L, Li J and Song L: Bmi-1, stem
cells and cancer. Acta Biochim Biophys Sin (Shanghai). 41:527–534.
2009. View Article : Google Scholar
|
|
23
|
Kreso A, van Galen P, Pedley NM,
Lima-Fernandes E, Frelin C, Davis T, Cao L, Baiazitov R, Du W,
Sydorenko N, et al: Selfrenewal as a therapeutic target in human
colorectal cancer. Nat Med. 20:29–36. 2014. View Article : Google Scholar
|
|
24
|
Bonasio R, Lecona E, Narendra V, Voigt P,
Parisi F, Kluger Y and Reinberg D: Interactions with RNA direct the
Polycomb group protein SCML2 to chromatin where it represses target
genes. eLife. 3:e026372014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Galoian K, Qureshi A, Wideroff G and
Temple HT: Restoration of desmosomal junction protein expression
and inhibition of H3K9-specific histone demethylase activity by
cytostatic proline-rich polypeptide-1 leads to suppression of
tumorigenic potential in human chondrosarcoma cells. Mol Clin
Oncol. 3:171–178. 2015.
|
|
26
|
Wong DJ, Liu H, Ridky TW, Cassarino D,
Segal E and Chang HY: Module map of stem cell genes guides creation
of epithelial cancer stem cells. Cell Stem Cell. 2:333–344. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Galoian K, Scully S and Galoyan A:
Myc-oncogene inactivating effect by proline-rich polypeptide
(PRP-1) in chondrosarcoma JJ012 cells. Neurochem Res. 34:379–385.
2009. View Article : Google Scholar
|
|
28
|
Irby RB and Yeatman TJ: Role of Src
expression and activation in human cancer. Oncogene. 19:5636–5642.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Clarke MF and Fuller M: Stem cells and
cancer: Two faces of eve. Cell. 124:1111–1115. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Durand B, Gao FB and Raff M: Accumulation
of the cyclin-dependent kinase inhibitor p27/Kip1 and the timing of
oligodendrocyte differentiation. EMBO J. 16:306–317. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yadav V, Sultana S, Yadav J and Saini N:
Gatifloxacin induces S- and G2-phase cell cycle arrest in
pancreatic cancer cells via p21/p27/p53. PLoS One. 7:e477962012.
View Article : Google Scholar
|
|
32
|
Jang TJ, Kang MS, Kim H, Kim DH, Lee JI
and Kim JR: Increased expression of cyclin D1, cyclin E and
p21(Cip1) associated with decreased expression of p27(Kip1) in
chemically induced rat mammary carcinogenesis. Jpn J Cancer Res.
91:1222–1232. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nadeem L, Brkic J, Chen YF, Bui T, Munir S
and Peng C: Cytoplasmic mislocalization of p27 and CDK2 mediates
the anti-migratory and antiproliferative effects of Nodal in human
trophoblast cells. J Cell Sci. 126:445–453. 2013. View Article : Google Scholar
|
|
34
|
Cloos PA, Christensen J, Agger K and Helin
K: Erasing the methyl mark: Histone demethylases at the center of
cellular differentiation and disease. Genes Dev. 22:1115–1140.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Loh YH, Zhang W, Chen X, George J and Ng
HH: Jmjd1a and Jmjd2c histone H3 Lys 9 demethylases regulate
self-renewal in embryonic stem cells. Genes Dev. 21:2545–2557.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang J, Park JW, Drissi H, Wang X and Xu
RH: Epigenetic regulation of miR-302 by JMJD1C inhibits neural
differentiation of human embryonic stem cells. J Biol Chem.
289:2384–2395. 2014. View Article : Google Scholar :
|
|
37
|
Abrahamyan SS, Davtyan TK, Khachatryan AR,
Tumasyan NV, Sahakyan IK, Harutyunyan HA, Chailyan SG and Galoyan
AA: Quantification of the hypothalamic proline-rich polypeptide 1in
rat blood serum. Neurochem J. 8:38–43. 2014. View Article : Google Scholar
|