Introduction
Lung cancer is the most frequently diagnosed cancer
and remains the leading cause of cancer death among males, and has
surpassed breast cancer as the leading cause of cancer death among
females in most developed countries (1). Currently, the typical non-small cell
lung cancer (NSCLC) patients account for 85% of the total lung
cancer patients (2).
miRNAs are a family of evolutionarily conserved
small non-coding RNAs that constitute a prevalent gene regulation,
they often block mRNA translation or negatively regulate its
stability through binding site within the 3′-UTR of target mRNA
(3). Recent studies suggested that
miRNA appears to be a key determinant in diagnosis and prognosis of
many diseases. miRNAs associated with tumorigenesis display either
tumor-promoting properties or tumor-suppressing characteristics.
For example, miR-212 in lung cancer (4) and the miR-17-92 cluster in malignant
lymphoma (5) act as oncogenes. In
contrast, let-7 in lung cancer (6)
and miR-551a/miR-483 in colorectal cancer (7) act as tumor suppressors.
Retinoic acid (RA), the major bioactive metabolite
of retinol or vitamin A, plays critical roles in cell growth,
differentiation and cell apoptosis (8). It has been successfully employed for
decades in the treatment of acute promyelocytic leukemia (9). In recent years, it was reported that
RA could regulate the expression level of miRNAs in cancer, such as
in breast cancer cells (10,11).
miR-512-5p is located at chromosome 19q13.42 and is a member of
miR-512 cluster, which includes two copies of miR-512 (miR-512-1
and miR-512-2) and 46 duplicates of miR-519 (12). From previous observations and
studies, miR-512-5p is significantly upregulated in cervical
intraepithelial neoplasia specimens compared to normal cervical
epithelium samples (13).
Activation of miR-512-5p by epigenetic treatment induces apoptosis
of gastric cancer cells by suppressing Mcl-1 (14).
p21WAF/CIP1 is located at chromosome
6p21.2, and it belongs to the Cip/Kip family of CKIs (15). Early studies supported the view
that p21 is a master effector of the p53-dependent G1 arrest
(16). p21 regulates its various
biological activities primarily by binding to and inhibiting the
cyclin/CDK activity, leading to growth arrest in the cell cycle
(17). However, recent studies
also showed that p21 has context-dependent function in apoptosis
(18).
Based on the present investigation we report that
miR-512-5p is downregulated in lung cancer tissues versus paired
normal lung tissues. Overexpression of miR-512-5p induced cell
apoptosis, and inhibited glycolysis in NSCLC cells. Further
evidence showed that p21 was the target of miR-512-5p, whose
downregulation may be responsible for the function of
miR-512-5p.
Materials and methods
Cell culture
A549, H1299 (American Type Culture Collection,
Manassas, VA, USA) and bulk-selected H1299 were cultured in
RPMI-1640 medium (Gibco, Grand Island, NY, USA), supplemented with
10% FBS (Biowest, Barcelona, Catalonia, Spain), 1% penicillin and
streptomycin and 1% sodium pyruvate (Gibco) at 37°C in a humidified
atmosphere of 5% CO2.
Human tissue samples
Twelve NSCLC tumor samples and their corresponding
normal tissues were collected from Shanghai Pulmonary Hospital,
snap-frozen in liquid nitrogen and stored at −70°C until analysis,
between August 2012 and January 2014. None of the patients received
pre-surgical chemo/radiation therapy. All NSCLC patients gave
written informed consent for the use of clinical specimens for
medical research. The study was reviewed and approved by the
Committee for Ethical Review of Research Involving Human Subjects
at Shanghai Pulmonary Hospital.
Expressing constructs
The pEGP-miR-512 construct was cloned as described
(19). In brief, the genomic
fragment of hsa-miR-512 precursor was amplified. The PCR primers
were: 5′-TCGAGGATCCACCTTTCACTATTGAGCA ACA-3′ and
5′-TCGAGCTAGCTGAAGTCCAGTTTAT GGCGCA-3′. The PCR product was
digested with EcoRI and NheI restriction enzymes and
cloned into the pEGP-miR vector (Cell Biolabs, San Diego, CA, USA).
The construct was designated as pEGP-miR-512.
Transient transfection and bulk-selected
H1299 cells
All transfection were carried out using
Lipofectamine™ 2000 (Invitrogen, Carlsbad, CA, USA) according to
the manufacturer's procedure. For H1299 cells stably expressing
miR-512, H1299 cells transfected with pEGP-miR-512 were selected
with 5 μg/ml puromycin (Sigma, St. Louis, MO, USA) for two weeks.
The bulk-selected H1299 cells stably expressing miR-512 were
maintained in RPMI-1640 (Gibco) with 2.5 μg/ml puromycin (Gibco).
Bulk cultures were used to avoid clone specific effects.
RNA isolation and quantitative
RT-PCR
Total RNA was extracted with Trizol (Invitrogen),
and miRNA was extracted with miRcute miRNA Isolation kit (Tiangen,
Shanghai, China). The RNA was converted to cDNA using Primer Script
RT Reagent kit (Takara, Tokyo, Japan), which was incubated for 15
min at 37°C followed by 5 min at 85°C. The specific reverse
transcription (RT) primer of miR-512-5p was
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGC ACTGGATACGACGAAAGTG-3′.
Real-time quantitative PCR was run on a Light-Cycler Roche 480
(Roche Applied Science, Shanghai, China) with reactions consisted
of a hot start (10 min at 95°C), followed with 40 cycles of 15 sec
at 95°C and 60 sec at 60°C. The expression level of miR-512-5p and
p21 was normalized to U6 and GAPDH, respectively. Each sample was
run in triplicate to ensure accuracy. The PCR primers for
miR-512-5p were 5′-CGGCGGCACTCAGCCTTGAGGG-3′ and
5′-GTGCAGGGTCCGAGGT-3′. The PCR primers for p21 were
5′-CTACTGAGGAGCCAGCGTCTA-3′ and 5′-CTGCCCATCATCATGACCT-3′. The PCR
primers for GAPDH were 5′-CTTAGATTTGGTCGTATTGG-3′ and
5′-GAAGATGGTGATGGGATT-3′.
The miRNA mimics, miRNA inhibitor and
siRNA
miR-512-5p mimic and miR-512-5p inhibitor were
synthesized by Biotend (Shanghai, China), and p21 siRNA were
synthesized by GenePharma (Shanghai, China). The sequence of
miR-512-5p inhibitor was 5′-GAAAGUGCCCUCAAGGCUGAGUG-3′. Two pairs
of p21 siRNA were synthesized. The sequences of p21 siRNA-1
(sip21-1) were 5′-AAUGGCGGGCUGCAU CCAGGA-3′ and
5′-UCCUGGAUGCAGCCCGCCAUU-3′, and p21 siRNA-2 (sip21-2) were
5′-CUUCGACUUUGUCA CCGAG-3′ and 5′-CUCGGUGACAAAGUCGAAG-3′.
Cell apoptosis assay
Apoptosis rate was assessed by the FITC-Annexin V/PI
Apoptosis Detection kit (BD Pharmingen, San Diego, CA, USA)
according to the manufacturer's instruction. At 48 h after
transfection, cells were collected and assayed with Annexin V
binding buffer containing FITC-Annexin V and PI for 15 min at 37°C
in the dark. Cells were analyzed by a flow cytometer (BD
FACSCalibur, BD Bioscience, San Jose, CA, US) and CellQuest
software.
Glucose consumption and lactate
production
Cells were seeded on a 12-well plate well, and the
culture medium was changed to RPMI-1640 medium without FBS 4 h
after transfection. The concentrations of glucose and L-lactate
were measured after another 36 h of incubation with a glucose test
kit (Sigma) and L-lactate assay kit (Sigma), respectively.
Cell proliferation assay
Cell Counting Kit-8 (Dojindo Laboratories, Kumamoto,
Japan) was used to perform cell proliferation assays according to
the protocol of the manufacturers on days 1, 2, 3 and 4
post-plating. Briefly, 5000 cells were plated in 96-well culture
plate per well and were incubated in 10 μl CCK-8 per well for 2 h,
then examined. Absorbance was measured at 450 and 630 nm (used as
reference) with a Microplate Reader ELx808 (Bio-Tek Instruments,
Winooski, VT, USA). Each experiment was performed in
sextuplicate.
Cell migration
For cell migration assay, 5×104 cells in
0.1 ml media containing 1% FBS were seeded into the upper chamber,
with 0.6 ml of medium (10% FBS) under the upper chamber. The cells
were then cultured for 72 h, and the non-invading cells were wiped
off with a cotton swab. Cells under the chamber were fixed with
methanol, stained with DAPI (Sigma), and washed with
phosphate-buffered saline (PBS). All of the cells on the bottom of
the chamber were counted under the microscope.
Dual luciferase assay
H1299 cells were co-transfected with 50 pmol
miR-512-5p, 1 μg firefly luciferase reporter containing the wild-
or mutant-type putative target site in 3′ UTR of p21 gene and 0.15
μg Renilla luciferase control vector pRL-TK (Promega,
Madison, WI, USA). Assays were assessed 48 h later using a
SpectraMax M5 plate reader (Molecular Devices, Sunnyvale, CA, USA).
Co-transfections of luciferase report construct and pRL-TK with
miR-512-5p or NC were performed with the same procedure.
Western blotting
Immunoblot was performed as follows. Whole cells
were lysed in RIPA buffer (CW, Shanghai, China) supplemented with
protease inhibitors (100 μg/ml PMSF, Sigma-Aldrich; Complete
Protease Inhibitor Cocktail, Roche) for 15 min. Protein extracted
from lysates were submitted to SDS-PAGE (10%) and transferred to
polyvinylidenedifluoride plus membrane (Merck Millipore, Darmstadt,
Germany). Membranes were incubated in 5% non-fat milk prepared with
Tris-buffered saline containing 0.1% Tween-20 (TBST) at room
temperature (RT) for 1.5 h, and the membranes were immunoblotted
with rabbit anti-p21 polyclonal antibody (1:1000, Proteintech,
Chicago, IL, USA) or with mouse anti-tubulin monoclonal antibody
(1:5000, Sigma) overnight at 4°C, washed two times using TBST at
RT, and then incubated in secondary antibodies against rabbit or
mouse IgG (1:5000, KPL, Gaithersburg, MD, USA) for 1.5 h at RT. The
membranes were washed three times and antigen-antibody complexes
were visualized by enhanced chemiluminescence (GE Healthcare,
Little Chalfont, Buckinghamshire, UK) following the manufacturer's
instructions.
Statistical analysis
Data are presented as mean ± SD of at least three
independent experiments for each cellular experimental group.
Samples were analyzed by two-tailed unpaired Student's t test, and
a P<0.05 was considered statistically significant. We used one,
two, and three asterisks in the figures to represent P<0.05,
P<0.01, and P<0.001, respectively. All histograms were drawn
using GraphPad Prism 5.
Results
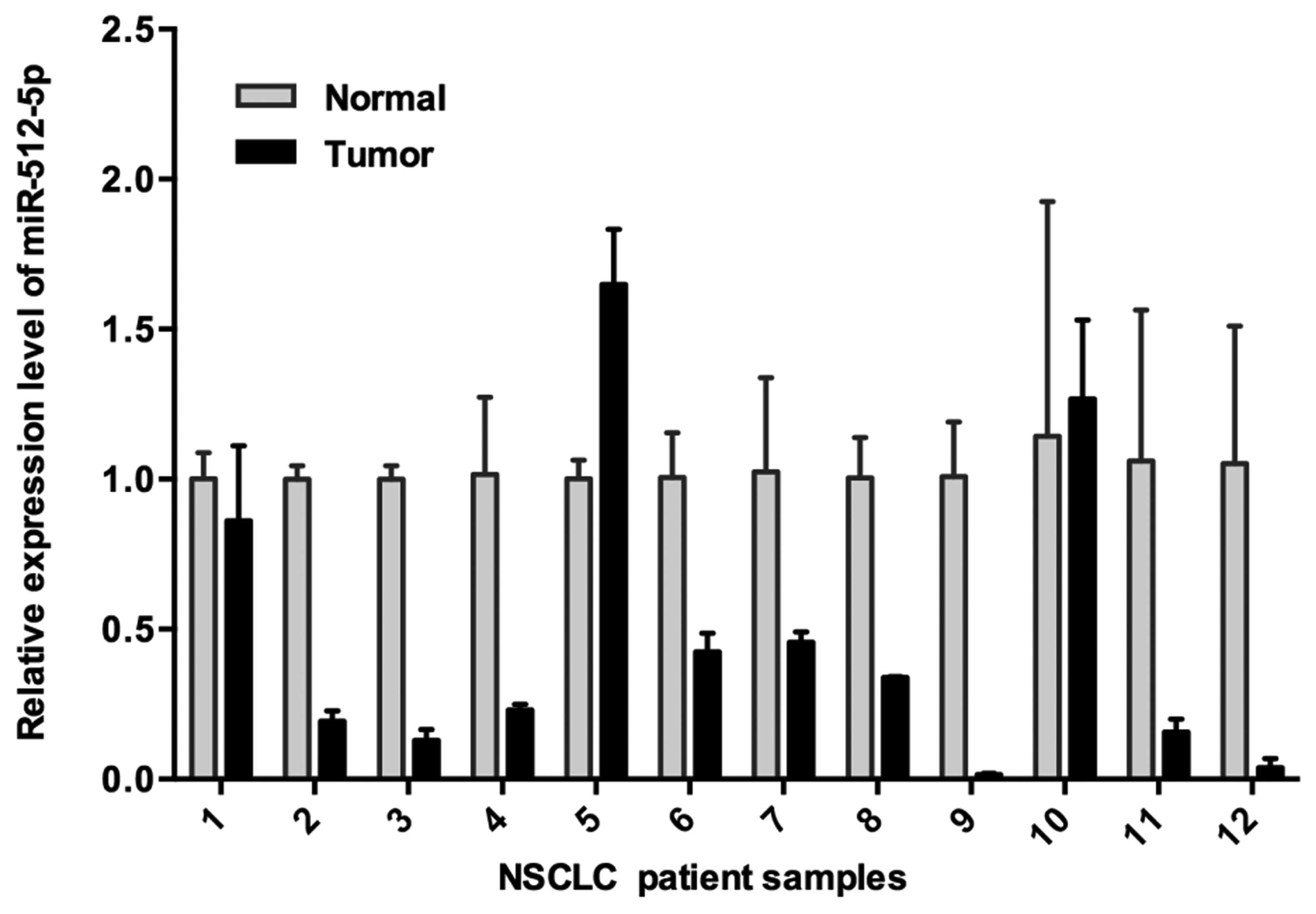
miR-512-5p is suppressed in human NSCLC
tissues
miR-512-5p is reported to be significantly
upregulated in cervical intraepithelial neoplasia specimens
compared to normal cervical epithelium samples (13). To assess the expression levels of
miR-512-5p in NSCLC, we collected 12 pairs of flash-frozen tumor
vs. normal lung tissues from 12 NSCLC patients. Of the 12 NSCLC
patients, 9 patients were subtyped as adenocarcinoma, 3 were
subtyped as squamous carcinoma (data not shown). Expression of
miR-512-5p was examined by RT-qPCR. Relative miR-512-5p expression
levels in NSCLC tumors were presented as-fold change (2ΔCt
tumor−ΔCt normal) of tumor versus paired normal tissues,
expression level of normal tissues was assigned as 1.0. Compared to
paired normal tissues, miR-512-5p was repressed in tumor samples in
83% (10/12) of NSCLC cases; while two paired samples showed
increased expression of miR-512-5p in tumor tissue (Fig. 1). This result suggested miR-512-5p
might act as a tumor suppressor in NSCLC.
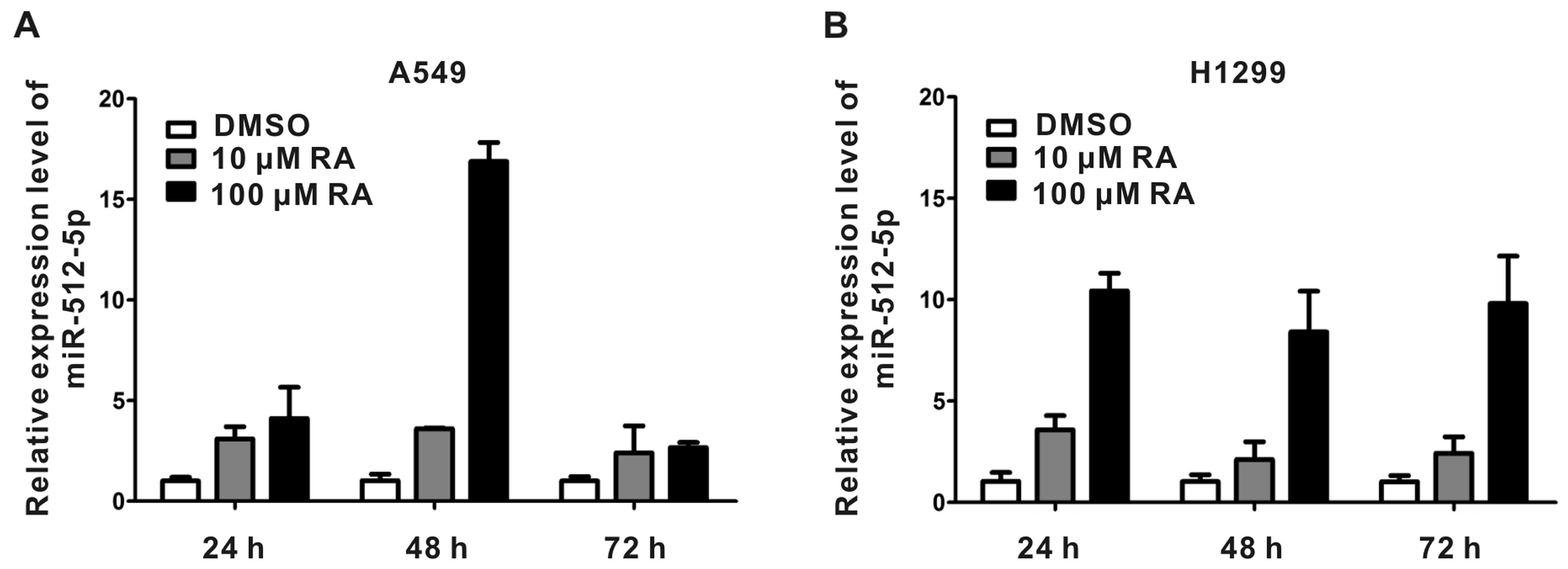
RA induces the expression of
miR-512-5p
To explore whether miR-512-5p played a role in the
progress of NSCLC cells, we set out to analyze the expression level
of miR-512-5p in A549 and H1299 with and without RA treatment at 10
μM or 100 μM at different time points. Cells treated with dimethyl
sulfoxide (DMSO) were used as control. As shown in Fig. 2A, the expression of miR-512-5p
increased about 3-fold compared with that of control when exposed
to 10 μM RA for 24, 48 and 72 h in A549 cells. As the RA
concentration was expanded to 100 μM, the expression of miR-512-5p
increased by 5-, 17- and 3-fold at 24, 48 and 72 h, respectively.
In H1299 cells, the expression of miR-512-5p increased less than
4-fold compared with that of control when treated with 10 μM RA.
However, when exposed to 100 μM RA, the expression level of
miR-512-5p increased to about 10-fold at these three time points
(Fig. 2B).
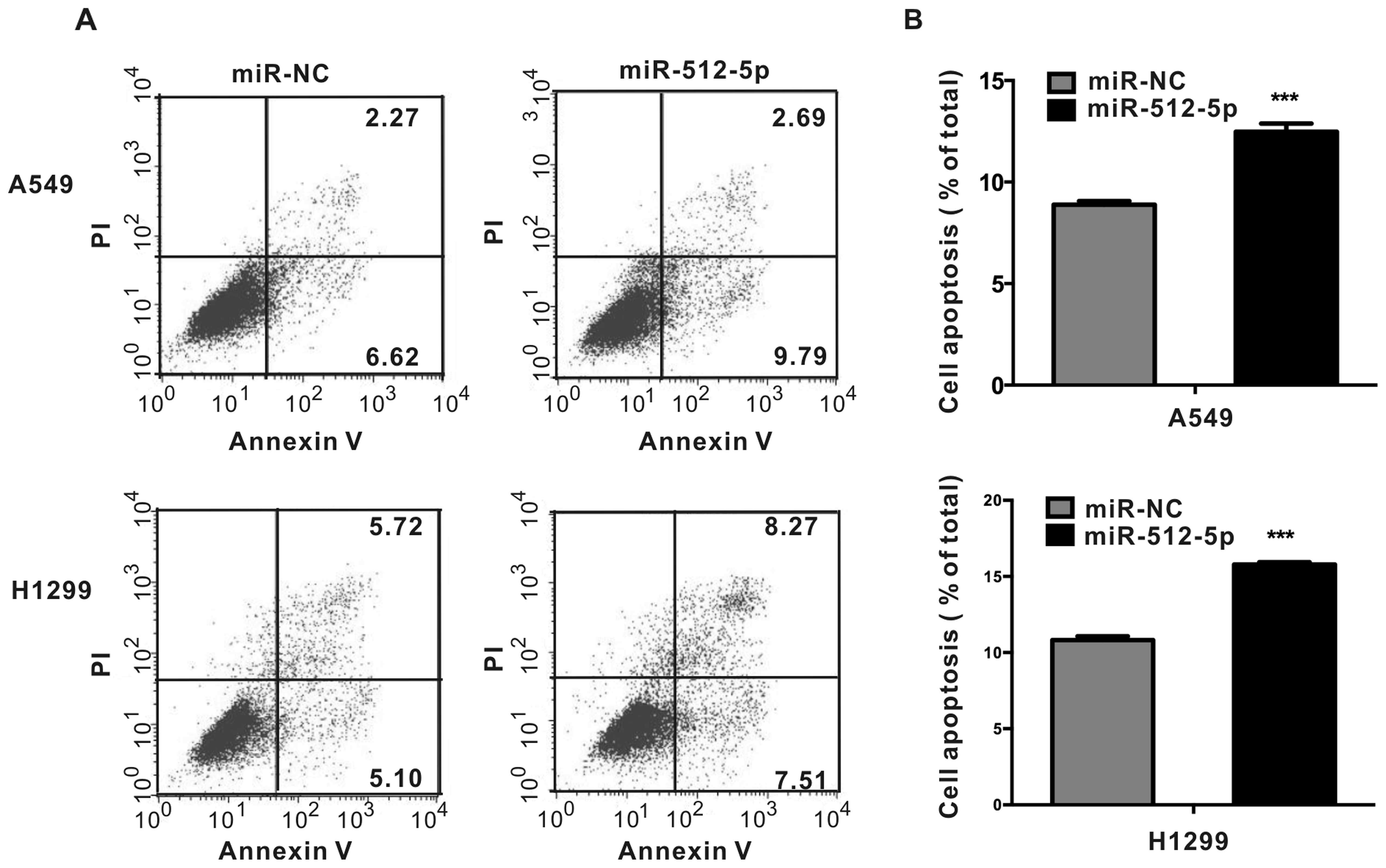
Overexpression of miR-512-5p promotes
cell apoptosis
A549 and H1299 cells were cultured for 48 h after
transfection with 50 nM miR-NC or miR-512-5p mimics, then stained
with FITC-Annexin V and PI, and analyzed by flow cytometry. We
found that cells overexpressing miR-512-5p had a larger population
of apoptotic cells than control cells. In A549 cells,
overexpression of miR-512-5p caused 40% increase of apoptotic cells
compared with that of control cells. In H1299 cells, the increase
of apoptotic cells was about 46% (Fig.
3).
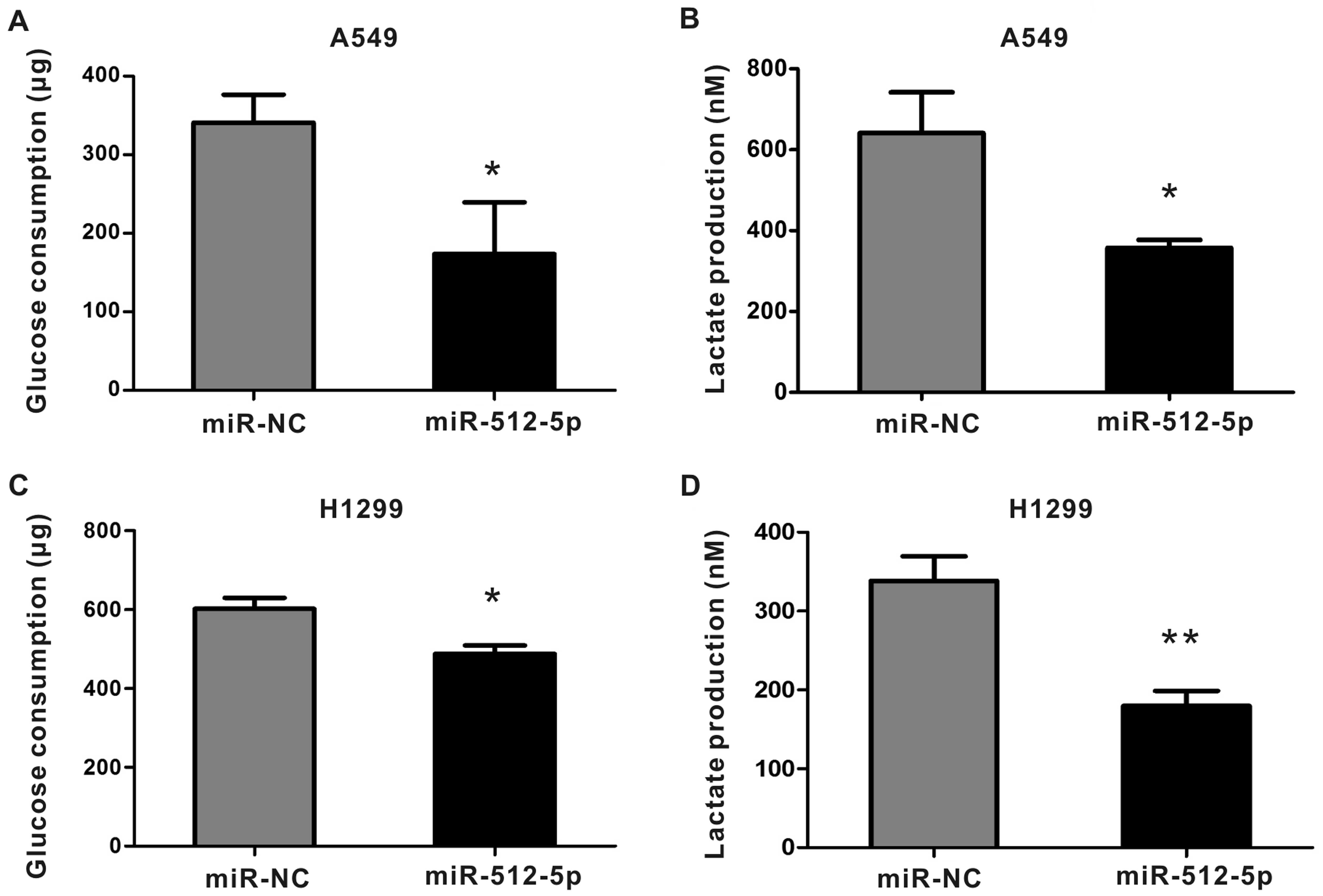
miR-512-5pinhibits glycolysis in NSCLC
cells
The Warburg effect of energy metabolism is critical
to the survival and proliferation of cancer cells (20). We asked whether miR-512-5p affected
cancer cell energy metabolism. To this end, we transfected A549 and
H1299 with miR-512-5p and 4 h later the culture medium was changed
to RPMI-1640 without FBS. After another 36 h of culture, the
glucose consumption and lactate production were detected. As shown
in Fig. 4, the rates of glucose
consumption and lactate production were strongly decreased by
miR-512-5p overexpression in A549 and H1299 cells.
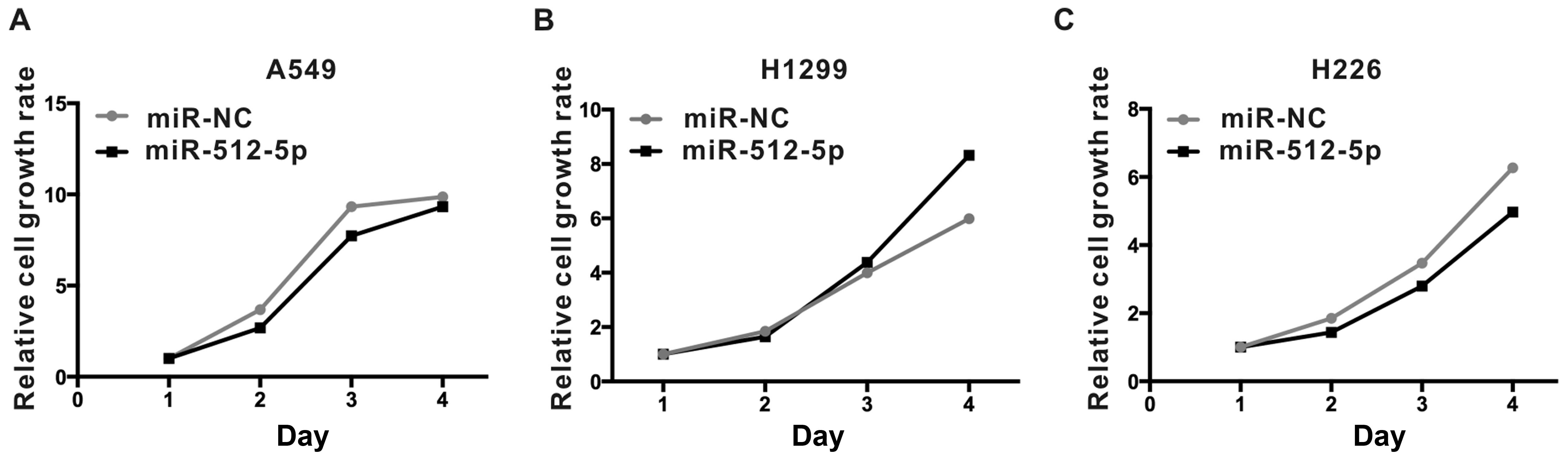
miR-512-5p has no effect on cell
proliferation
We assessed cell growth rate at day 1, 2, 3 and 4 by
CCK-8 assay to study the role of miR-512-5p on cell proliferation
in A549, H1299 and H226 cells. The cells were transfected with
either miR-512-5p or miR-NC. Compared with miR-NC, miR-512-5p
mimics exhibited almost no effect on cell proliferation on A549,
H1299 and H226 cells (Fig. 5).
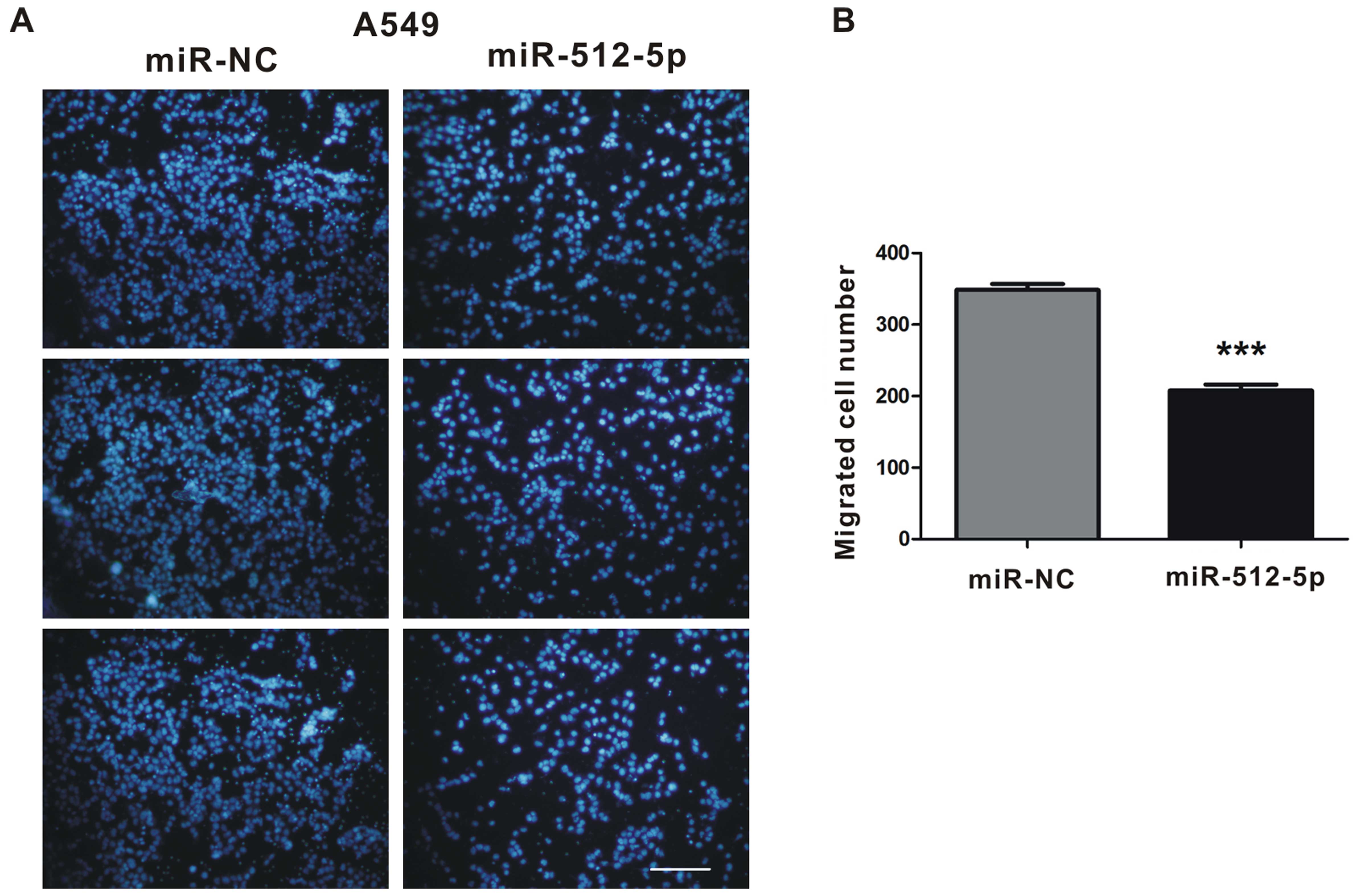
miR-512-5p attenuates the capacity of
cell migration in A549 cells
Metastasis is a process that involves multiple
steps, including cancer cell adhesion, invasion and migration
(21). To explore the role of
miR-512-5p in these processes, we transfected A549 cells with
miR-NC or miR-512-5p and seeded cells into the transwell chambers.
After incubation for 72 h, medium in chamber was removed and all of
the cells on the bottom of the chamber were counted under the
microscope. We found overexpression of miR-512-5p attenuated A549
cells migration by 39% compared to that of control (Fig. 6).
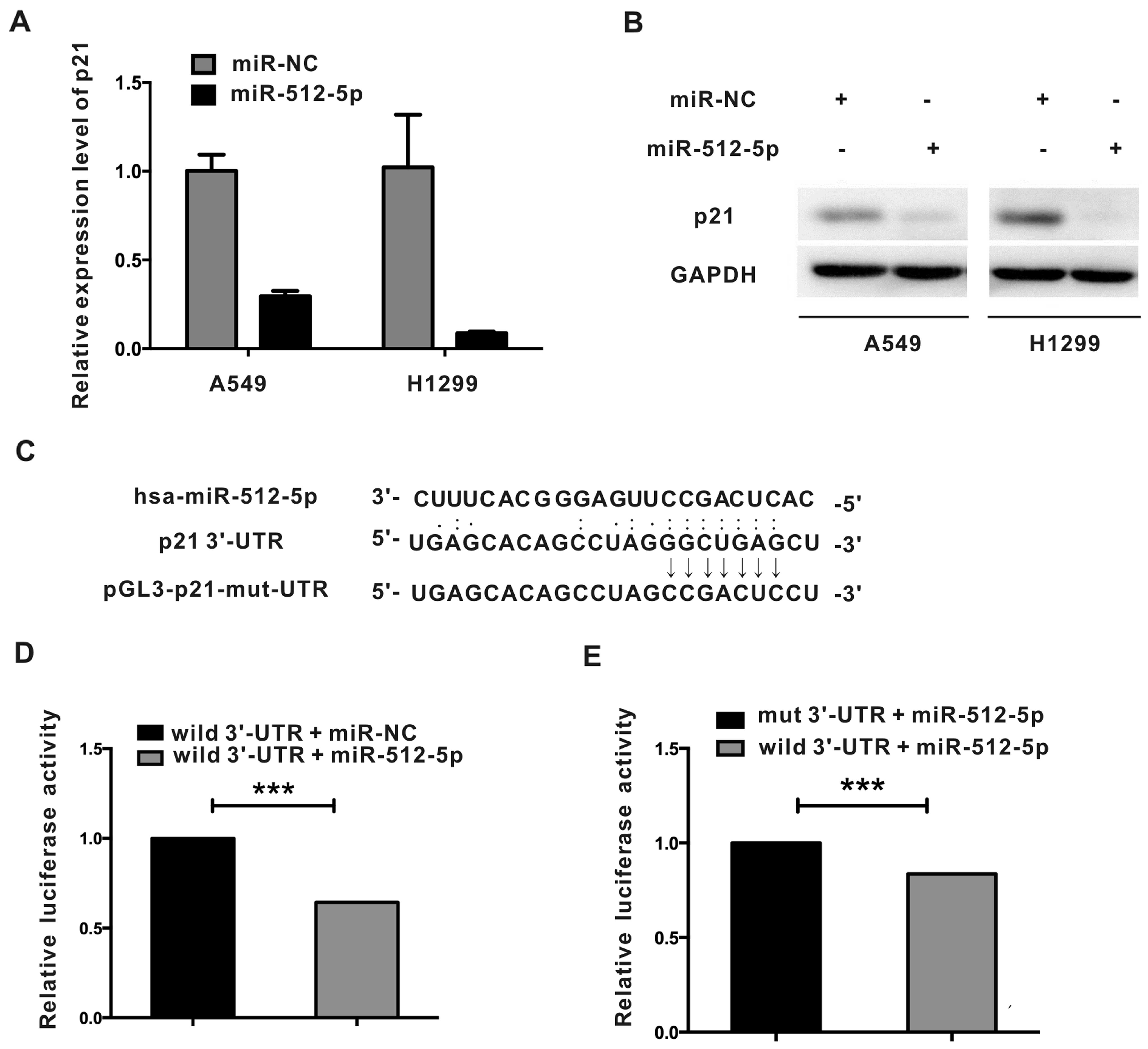
Overexpression of miR-512-5p decreases
p21 mRNA and protein
In the screening of genes that could be affected by
miR-512-5p overexpression, we found that p21 was down-regulated by
miR-512-5p. A549 and H1299 cells were transiently transfected with
miR-NC or miR-512-5p mimics for 48 h, and the expression of p21
mRNA and protein were detected by real-time PCR and western
blotting. As shown in Fig. 7A,
overexpression of miR-512-5p decreased p21 mRNA by 71% and 91% in
A549 and H1299 cells, respectively. Compared with cells transfected
with miR-NC, p21 protein was decreased significantly in cells
transfected with miR-512-5p (Fig.
7B). The experiments showed miR-512-5p inhibited the expression
of p21 in NSCLC cells.
p21 is a direct target gene of
miR-512-5p
To explore the mechanism of miR-512-5p in
downregulating p21, we first performed a computational screen for
genes with complementary sites to miR-512-5p in its 3′-UTR using
open-access software including TargetScan (www.targetscan.org), PicTar (http://pictar.bio.nyu.edu), Sanger microRNA target
(http://microrna.sanger) to check if p21 was a
target of miR-512-5p. However, no binding site in p21 was
identified. We then carried out a blast to examine whether there
was complementary site between p21 3′-UTR and miR-512-5p. We found
a potential binding site of miR-512-5p within p21 3′-UTR (Fig. 7C). To test whether p21 is a direct
target of miR-512-5p, mutation in the potential binding site was
generated in the p21 3′-UTR sequence of in the complementary site
(Fig. 7C). p21 3′-UTR fragment
containing wild-type or mutant type binding site was cloned into
pGL3-prometor vector. Luciferase reporter construct containing
wild-type 3′-UTR was cotransfected with pRL-TK, as well as miR-NC
or miR-512-5p mimics. Furthermore, luciferase reporter construct
containing wild-type 3′-UTR or mutant type 3′-UTR was cotransfected
with miR-512-5p mimics and pRL-TK. As shown in Fig. 7D, the luciferase activity of
wild-type 3′-UTR cotransfected with miR-512-5p decreased by 36%
compared with the wild-type 3′-UTR cotransfected with miR-NC.
Whereas, the luciferase activity of miR-512-5p cotransfected with
wild-type p21 3′-UTR was decreased by 17% compared with that
cotransfected with mutant-type p21 3′-UTR (Fig. 7E). Collectively, these results
indicated that miR-512-5p could bind to p21 3′-UTR to repress p21
expression.
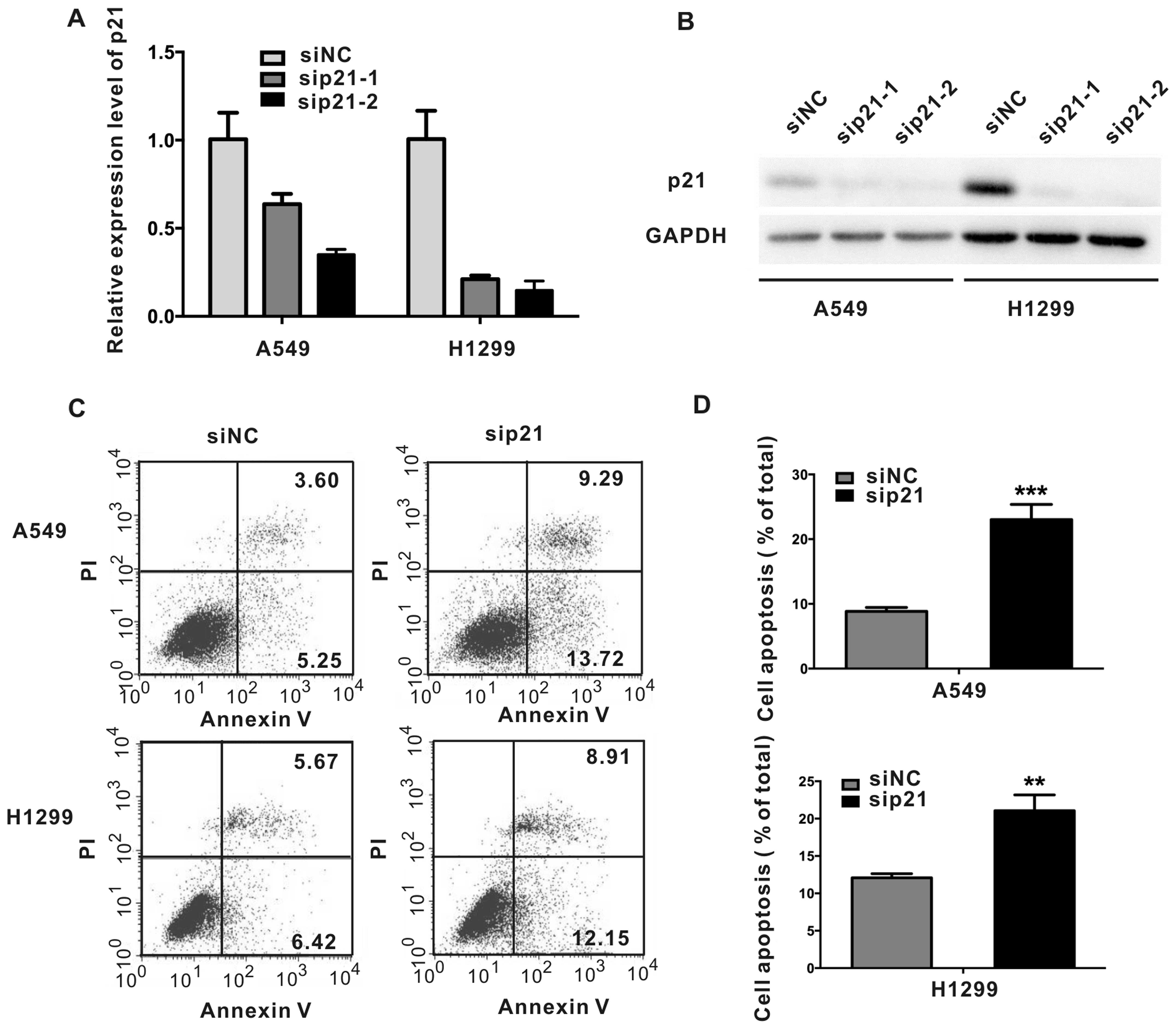
Knockdown of p21 promotes apoptosis in
NSCLC cells
We identified that p21 is a target gene of
miR-512-5p. To evaluate how p21 is involved in the functioning of
miR-512-5p, we used p21 small interfering RNA (siRNA) to knock down
the expression of p21 and investigated NSCLC cell apoptosis. siRNAs
against p21 (sip21) were synthesized according to previous studies
(22,23), and the silencing efficiency was
examined. A siRNA lacking homology to the genome was used as
negative control. Both sip21-1 and sip21-2 had good silencing
efficiency in A549 and H1299 cells, and sip21-2 showed better
silencing efficiency (Fig. 8A and
B). Thus, we used sip21-2 (sip21 for short) for further
analyses. Cell apoptosis assay was then performed. As shown in
Fig. 8C and D, knockdown of p21
significantly promoted cell apoptosis in A549 and H1299 cells. The
results were in congruent with miR-512-5p overexpression.
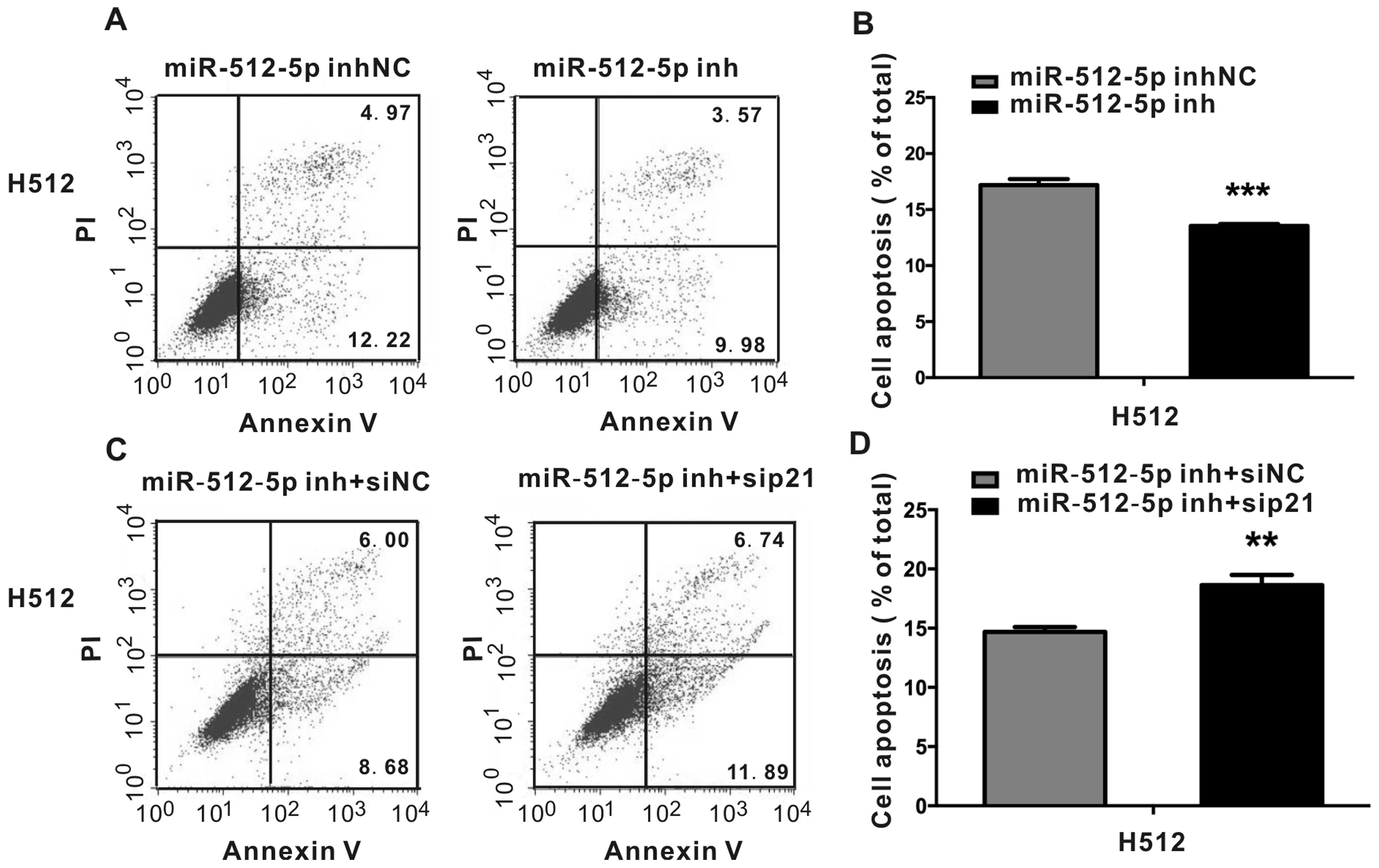
Knockdown of miR-512-5p in H1299 cells
stably expressing miR-512 attenuates apoptosis and the effect could
be reversed by p21 siRNA
To confirm that miR-512-5p may affect cell apoptosis
through targeting p21, we also detected the impact of p21 siRNA on
cell apoptosis in the presence of miR-512-5p inhibitor in H1299
cells stably expressing miR-512-5p. To clone miR-512-5p into
pEGP-miR vector, we amplified a 300 bp genomic fragment containing
miR-512-5p according to the instruction. This fragment contains the
precursor of miR-512-3p, which can also transcribe miR-512-3p. We
selected H1299 cells stably expressing miR-512 (designated as H512)
or EGP control. To avoid clone-specific effects, we used
bulk-selected cells for functional analysis.
We then detected the impact of p21 siRNA on cell
apoptosis in the presence of miR-512-5p inhibitor in H512. The
efficiency of miR-512-5p inhibitor in knocking down miR-512-5p
expression was examined by RT-PCR. The result showed miR-512-5p
inhibitor inhibited 82% of miR-512-5p expression compared with
inhibitor NC (data not shown). miR-512-5p inhibitor was transfected
into H512 to knockdown the expression of miR-512-5p, and apoptosis
assays were performed. As shown in Fig. 9, miR-512-5p inhibitor attenuated
apoptosis in H512 cells (Fig. 9A and
B). In the presence of p21 siRNA, the effect of miR-512-5p
inhibitor on apoptosis could be rescued (Fig. 9C and D).
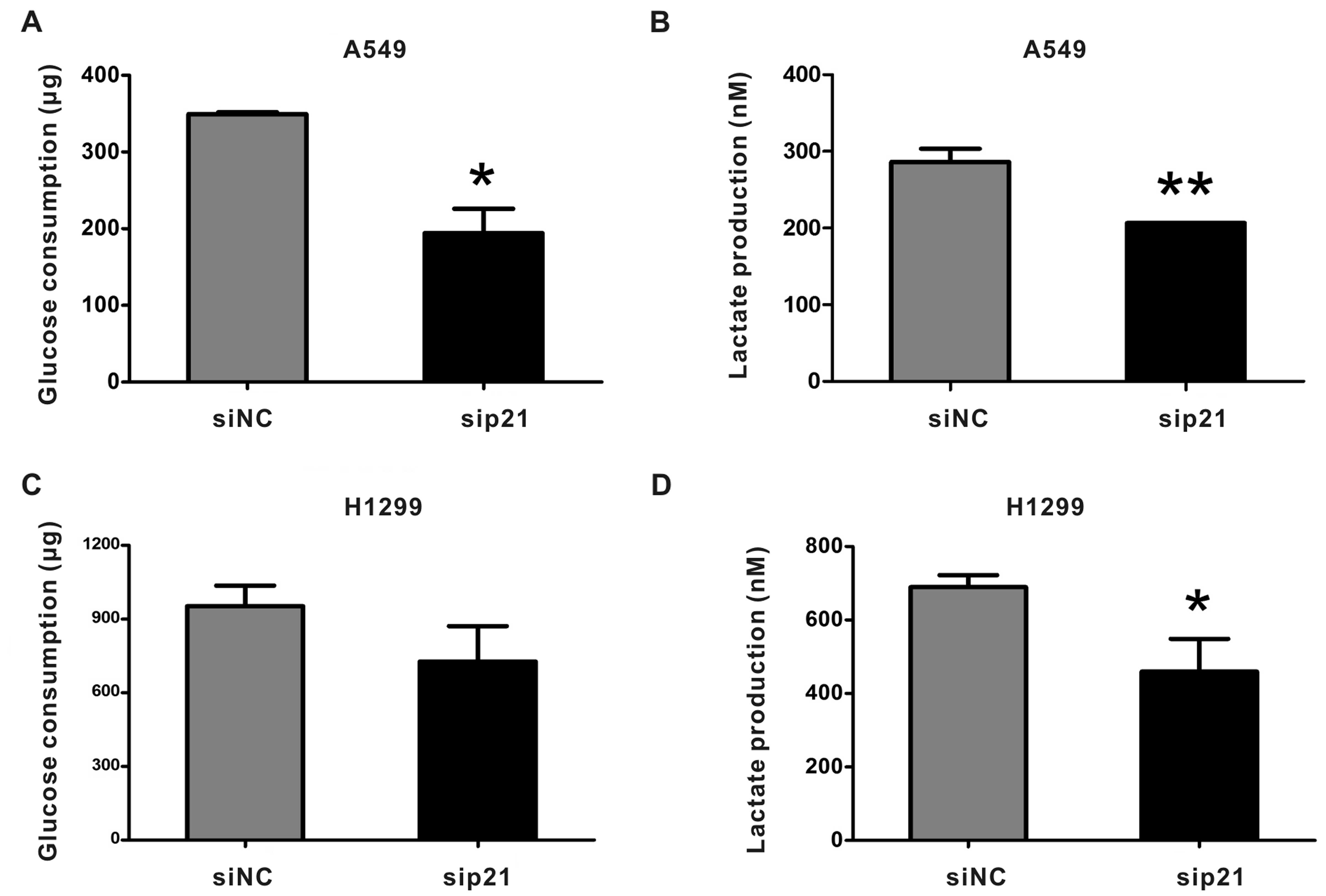
Knockdown of p21 decreases glycolysis in
NSCLC cells
To further verify that miR-512-5p affected NSCLC
cell glycolysis through targeting p21, we transfected A549 and
H1299 with sip21 for 4 h and then the culture medium was changed to
RPMI-1640 without FBS. After incubation for 36 h, we detected
glucose consumption and lactate production. As expected, the rates
of glucose consumption and lactate production were strongly
decreased by sip21 in A549 cells (Fig. 10A and B). In H1299 cells, glucose
consumption showed a trend of decreasing (Fig. 10C), while the lactate production
had a significant decrease (Fig.
10D). The results were consistent with miR-512-5p
overexpression.
Discussion
miR-512-5p is the 5′ terminal of miR-512 precursor.
It was reported that epigenetic treatment could rescue the
expression of miR-512-5p, and induce gastric cancer cell apoptosis
(14). Reactivation of both
miR-512-5p and miR-373 was able to accelerate cisplatin-induced
apoptosis inhibiting cell migration (24).
In this study, we first detected the expression of
miR-512-5p in 12 paired normal lung tissues and NSCLC tumor
tissues. We found that the expression of miR-512-5p was
downregulated in 83% (10 out of 12) of the NSCLC patient samples,
which suggested miR-512-5p acts as a tumor suppressor. Retinoic
acid (RA) played an important role in cell differentiation, cell
proliferation and cell apoptosis (25). Previous studies showed
dysregulation of RA pathway was related to tumorigenesis, including
lung cancer (26). Thus, we
treated NSCLC cells with RA to test whether RA affect the
expression of miR-512-5p. Real-time PCR showed that when A549 and
H1299 cells were exposed to 10 μM RA or 100 μM RA, the expression
of miR-512-5p increased.
To investigate the influence of miR-512-5p on NSCLC
cells, we overexpressed miR-512-5p by transient transfection. We
found that overexpression of miR-512-5p promoted cell apoptosis,
inhibited cell glycolysis and attenuated A549 cells migration, but
had no effect on cell proliferation in A549, H1299 and H226 cells.
Adi Harel and colleagues (24)
reported miR-512-5p inhibited A549 cells proliferation, which was
not consistent with our study. A plausible but yet-to-be validated
possibility is that the different cell culture condition may
attribute to the cell proliferation in A549 cells in the presence
of miR-512-5p.
To overcome bioenergetic and biosynthetic demands of
abnormal proliferation, cancer cells must reprogram their
metabolism and become more dependent on aerobic glycolysis
(27). Recent studies reported
that low intracellular concentration of glucose may induce ATP
depletion, stimulate mitochondrial death pathway cascade, and may
induce oxidative stress and trigger of bax-associated events
(28–30). Taken together with the function of
miR-512-5p in cell apoptosis and cell glycolysis, the inhibition of
glucose uptake by miR-512-5p might play a role in the development
of NSCLC cells.
To illuminate the effect of miR-512-5p in NSCLC
cells, we identified that p21 is a new target of miR-512-5p.
Although open-access software did not reveal p21 as a miR-512-5p
target, blast screen did find a miR-512-5p complementary site in
the p21 3′-UTR. The complementary site mainly includes the seed
region of miR-512-5p, as well as a few bases in the 3′ region. The
luciferase reporter assay confirmed that the disruption of
complementary site in miR-512-5p seed region showed higher
luciferase activity compared with the wild-type binding site.
Computational prediction of miRNA target genes is difficult in
animal genomes due to the imperfect pairing of the miRNA with the
corresponding target site and the software often generate many
false positive interactions. Our result showed that Blast-based
method might help predicting the miRNA target.
As a member of the cyclin-dependent kinase
inhibitors, p21 plays a significant role in cell cycle by acting as
cyclin kinase inhibitor (CKI). p21 also directly blocks DNA
replication and mediates DNA repair processes by binding to
proliferating cell nuclear antigen (PCNA) (31). In addition, p21 associates directly
with E2F1 (32), STAT3 (33) and MYC (34) thereby suppressing transcription
activities.
Although best known for its important role in
growth-inhibition, p21 also protects cells against apoptosis in
growth factor deprivation, p53 overexpression and during the
differentiation of monocytes (35). Under these conditions,
cytoplasmically localized p21 may bind to and attenuate the
activity of proteins directly associated with apoptosis, including
apoptosis signal-regulating kinase 1 (ASK1), stress-activated
protein kinases (SAPKs), procaspase 3, caspase 8 and caspase 10
(35,36). However, p21 might improve apoptosis
under certain stimuli, such as activation of tumor necrosis factor
family of death receptors or effects on DNA repair (37). Therefore, the function of p21
depends on the specific circumstances, suggests that it can be a
tumor suppressor or an oncogene.
We found knockdown of p21 promoted cell apoptosis,
and decreased cell glycolysis. The results were consistent with
miR-512-5p overexpression. However, p21siRNA exhibited a more
obvious effect on cell apoptosis in A549 and H1299 cells. It is
well known that a single miRNA can bind to and regulate many
different mRNA targets. miR-512-5p may target other genes to
regulate cell apoptosis, which might neutralize the effect of
targeting p21. We also tested the effect of miR-512-5p in H1299
cells stably expressing miR-512. miR-512-5p inhibitor significantly
depressed cell apoptosis, and in the presence of miR-512-5p
inhibitor, p21siRNA was able to rescue cell apoptosis. The results
revealed miR-512-5p induces NSCLC cell apoptosis by directly
regulating p21.
In conclusion, our study demonstrates miR-512-5p
induced NSCLC cells apoptosis and inhibited glycolysis by
regulating p21 in A549 and H1299 cells. We also found that
miR-512-5p is decreased in 83% (10/12) of NSCLC patient lung cancer
samples compared with normal sample from the same patient, which
suggests that miR-512-5p may act as a tumor suppressor in
NSCLC.
Acknowledgements
This study was supported by Shanghai Municipal
Natural Science foundation 14ZR1403700.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li J, Bi L, Sun Y, Lu Z, Lin Y, Bai O and
Shao H: Text mining and network analysis of molecular interaction
in non-small cell lung cancer by using natural language processing.
Mol Biol Rep. 41:8071–8079. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Joshi P, Jeon YJ, Laganà A, Middleton J,
Secchiero P, Garofalo M and Croce CM: MicroRNA-148a reduces
tumorigenesis and increases TRAIL-induced apoptosis in NSCLC. Proc
Natl Acad Sci USA. 112:8650–8655. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li Y, Zhang D, Chen C, Ruan Z, Li Y and
Huang Y: MicroRNA-212 displays tumor-promoting properties in
non-small cell lung cancer cells and targets the hedgehog pathway
receptor PTCH1. Mol Biol Cell. 23:1423–1434. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mendell JT: miRiad roles for the miR-17-92
cluster in development and disease. Cell. 133:217–222. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Johnson SM, Grosshans H, Shingara J, Byrom
M, Jarvis R, Cheng A, Labourier E, Reinert KL, Brown D and Slack
FJ: RAS is regulated by the let-7 microRNA family. Cell.
120:635–647. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Loo JM, Scherl A, Nguyen A, Man FY,
Weinberg E, Zeng Z, Saltz L, Paty PB and Tavazoie SF: Extracellular
metabolic energetics can promote cancer progression. Cell.
160:393–406. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Okazawa H, Shimizu J, Kamei M, Imafuku I,
Hamada H and Kanazawa I: Bcl-2 inhibits retinoic acid-induced
apoptosis during the neural differentiation of embryonal stem
cells. J Cell Biol. 132:955–968. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Burnett A, Wetzler M and Löwenberg B:
Therapeutic advances in acute myeloid leukemia. J Clin Oncol.
29:487–494. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fisher JN, Terao M, Fratelli M, Kurosaki
M, Paroni G, Zanetti A, Gianni M, Bolis M, Lupi M, Tsykin A, et al:
MicroRNA networks regulated by all-trans retinoic acid and
Lapatinib control the growth, survival and motility of breast
cancer cells. Oncotarget. 6:13176–13200. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Khan S, Wall D, Curran C, Newell J, Kerin
MJ and Dwyer RM: MicroRNA-10a is reduced in breast cancer and
regulated in part through retinoic acid. BMC Cancer. 15:3452015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lichner Z, Páll E, Kerekes A, Pállinger E,
Maraghechi P, Bosze Z and Gócza E: The miR-290–295 cluster promotes
pluripotency maintenance by regulating cell cycle phase
distribution in mouse embryonic stem cells. Differentiation.
81:11–24. 2011. View Article : Google Scholar
|
|
13
|
Cheung TH, Man KN, Yu MY, Yim SF, Siu NS,
Lo KW, Doran G, Wong RR, Wang VW, Smith DI, et al: Dysregulated
microRNAs in the pathogenesis and progression of cervical neoplasm.
Cell Cycle. 11:2876–2884. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Saito Y, Suzuki H, Tsugawa H, Nakagawa I,
Matsuzaki J, Kanai Y and Hibi T: Chromatin remodeling at Alu
repeats by epigenetic treatment activates silenced microRNA-512-5p
with down-regulation of Mcl-1 in human gastric cancer cells.
Oncogene. 28:2738–2744. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sherr CJ and Roberts JM: CDK inhibitors:
Positive and negative regulators of G1-phase progression. Genes
Dev. 13:1501–1512. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Deng C, Zhang P, Harper JW, Elledge SJ and
Leder P: Mice lacking p21CIP1/WAF1 undergo normal development, but
are defective in G1 checkpoint control. Cell. 82:675–684. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Harper JW, Adami GR, Wei N, Keyomarsi K
and Elledge SJ: The p21 Cdk-interacting protein Cip1 is a potent
inhibitor of G1 cyclin-dependent kinases. Cell. 75:805–816. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Abbas T and Dutta A: p21 in cancer:
Intricate networks and multiple activities. Nat Rev Cancer.
9:400–414. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu X, Gao G, Chu K, Yang X, Ren S, Li Y,
Wu H, Huang Y and Zhou C: Inhibition of RAC1-GEF DOCK3 by
miR-512-3p contributes to suppression of metastasis in non-small
cell lung cancer. Int J Biochem Cell Biol. 61:103–114. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the Warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen J and Thompson LU: Lignans and
tamoxifen, alone or in combination, reduce human breast cancer cell
adhesion, invasion and migration in vitro. Breast Cancer Res Treat.
80:163–170. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang J, Attar E, Cohen K, Crumpacker C
and Scadden D: Silencing p21(Waf1/Cip1/Sdi1) expression increases
gene transduction efficiency in primitive human hematopoietic
cells. Gene Ther. 12:1444–1452. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rao A, Coan A, Welsh JE, Barclay WW,
Koumenis C and Cramer SD: Vitamin D receptor and p21/WAF1 are
targets of genistein and 1,25-dihydroxyvitamin D3 in human prostate
cancer cells. Cancer Res. 64:2143–2147. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Adi Harel S, Bossel Ben-Moshe N, Aylon Y,
Bublik DR, Moskovits N, Toperoff G, Azaiza D, Biagoni F, Fuchs G,
Wilder S, et al: Reactivation of epigenetically silenced miR-512
and miR-373 sensitizes lung cancer cells to cisplatin and restricts
tumor growth. Cell Death Differ. 22:1328–1340. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
di Masi A, Leboffe L, De Marinis E, Pagano
F, Cicconi L, Rochette-Egly C, Lo-Coco F, Ascenzi P and Nervi C:
Retinoic acid receptors: From molecular mechanisms to cancer
therapy. Mol Aspects Med. 41:1–115. 2015. View Article : Google Scholar
|
|
26
|
Chen F, Cao Y, Qian J, Shao F,
Niederreither K and Cardoso WV: A retinoic acid-dependent network
in the foregut controls formation of the mouse lung primordium. J
Clin Invest. 120:2040–2048. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jones RG and Thompson CB: Tumor
suppressors and cell metabolism: A recipe for cancer growth. Genes
Dev. 23:537–548. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Muñoz-Pinedo C, El Mjiyad N and Ricci JE:
Cancer metabolism: Current perspectives and future directions. Cell
Death Dis. 3:e2482012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
El Mjiyad N, Caro-Maldonado A,
Ramírez-Peinado S and Muñoz-Pinedo C: Sugar-free approaches to
cancer cell killing. Oncogene. 30:253–264. 2011. View Article : Google Scholar
|
|
30
|
Raina K, Agarwal C, Wadhwa R, Serkova NJ
and Agarwal R: Energy deprivation by silibinin in colorectal cancer
cells: A double-edged sword targeting both apoptotic and autophagic
machineries. Autophagy. 9:697–713. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li R, Waga S, Hannon GJ, Beach D and
Stillman B: Differential effects by the p21 CDK inhibitor on
PCNA-dependent DNA replication and repair. Nature. 371:534–537.
1994. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Delavaine L and La Thangue NB: Control of
E2F activity by p21Waf1/Cip1. Oncogene. 18:5381–5392. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Coqueret O and Gascan H: Functional
interaction of STAT3 transcription factor with the cell cycle
inhibitor p21WAF1/CIP1/SDI1. J Biol Chem. 275:18794–18800. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kitaura H, Shinshi M, Uchikoshi Y, Ono T,
Iguchi-Ariga SM and Ariga H: Reciprocal regulation via
protein-protein interaction between c-Myc and p21(cip1/waf1/sdi1)
in DNA replication and transcription. J Biol Chem. 275:10477–10483.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Roninson IB: Oncogenic functions of tumour
suppressor p21(Waf1/Cip1/Sdi1): Association with cell senescence
and tumour-promoting activities of stromal fibroblasts. Cancer
Lett. 179:1–14. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dotto GP: p21(WAF1/Cip1): More than a
break to the cell cycle? Biochim Biophys Acta. 1471:M43–M56.
2000.PubMed/NCBI
|
|
37
|
Gartel AL: The conflicting roles of the
cdk inhibitor p21(CIP1/WAF1) in apoptosis. Leuk Res. 29:1237–1238.
2005. View Article : Google Scholar : PubMed/NCBI
|