Introduction
Oral squamous cell carcinoma (OSCC) is the most
common form of oral cancers (1).
Despite advances in the diagnosis and the treatment of OSCC, the
prognosis remains poor, and metastasis to the cervical lymph nodes
and the remote organs is an important issue in determining the
prognosis of OSCC patients (2,3).
Recent studies have shown that epithelial-mesenchymal transition
(EMT) plays an important role in tumor invasion and metastasis
(4–8). In addition, many studies have shown
that expression of the EMT factors are significantly associated
with tumor invasion, tumor metastasis, and survival in patients
with SCC, including patients with OSCC (9–19).
These findings strongly indicate that EMT suppression may improve
the prognosis of patients with OSCC.
Hypoxic microenvironment is a characteristic feature
of solid tumors (20). Hypoxia
inducible factor-1α (HIF-1α), an oxygen-dependent α subunit of HIF
which activates the transcription of metastatic genes, is strongly
induced under hypoxic conditions (21). Increased expression of HIF-1α
induces tumor invasion and metastasis, and is associated with the
activation of EMT factors (22–26).
We previously demonstrated that transcutaneous CO2
suppresses the growth of primary human OSCC and metastasis to the
regional lymph nodes by both improving hypoxia and increasing
mitochondrial apoptosis in treated tissue (27).
According to the above background, we hypothesized
that improving hypoxia by transcutaneous CO2 could
suppress EMT. In the present study, we investigated whether oxygen
conditions affect the expression levels of HIF-1α and EMT factors
in OSCC in vitro and whether transcutaneous CO2
affects these factors in OSCC in vivo.
Materials and methods
Cell culture
An oral cancer cell line, HSC-3 was used in this
study (Health Science Research Resources Bank, Osaka, Japan). HSC-3
cells were established from a metastatic deposit of poorly
differentiated OSCC of the tongue in a mid-internal jugular lymph
node from a 64-year-old man (28).
HSC-3 cells in Eagle's minimum essential medium (Sigma-Aldrich, St.
Louis, MO, USA) supplemented with 10% fetal bovine serum
(Sigma-Aldrich) and 1000 U/ml penicillin/streptomycin solution
(Sigma-Aldrich) were routinely cultured in an incubator in 5%
CO2 at 37°C. Trypsin (0.25%) and
ethylenediaminetet-raacetic acid (0.02%) (Sigma-Aldrich) solution
were used to isolate cells for subculture, as previously described
(27,29).
Cell experiments
To investigate the effect of changing oxygen
conditions on OSCC cells in vitro, a total of
1.5×105 HSC-3 cells per well was seeded into a 6-well
plate and cultured for 24 h. Then, HSC-3 cells were incubated for
96 h in two different conditions: normoxic group (20%
O2, 5% CO2, 75% N2) or hypoxic
group (1% O2, 5% CO2, 94% N2).
After the incubation, the mRNA expression of Snail, Slug,
E-cadherin, N-cadherin and Vimentin in the cells were evaluated
using quantitative real-time PCR, and the expression of HIF-1α in
the cells was evaluated using immunoblot analysis.
Animal models
Male athymic BALB/c nude mice, aged 7-weeks, were
obtained from CLEA Japan (Tokyo, Japan). The animals were
maintained under pathogen-free conditions, in accordance with
institutional guidelines. All animal experiments were performed in
accordance with the Guidelines for Animal Experimentation at Kobe
University Animal Experimentation Regulations (permission number:
P120602) and were approved by the Institutional Animal Care and Use
Committee.
HSC-3 cells were implanted into the back of 24 mice
at doses of 2.0×106 cells in 500 μl phosphate buffered
saline (PBS), as previously described (27,29).
Mice were randomly divided into two treatment groups: a
CO2-treated group (n=12) and a control group (n=12).
Transcutaneous CO2
treatment
Transcutaneous CO2 treatment was
performed as previously described (27,29).
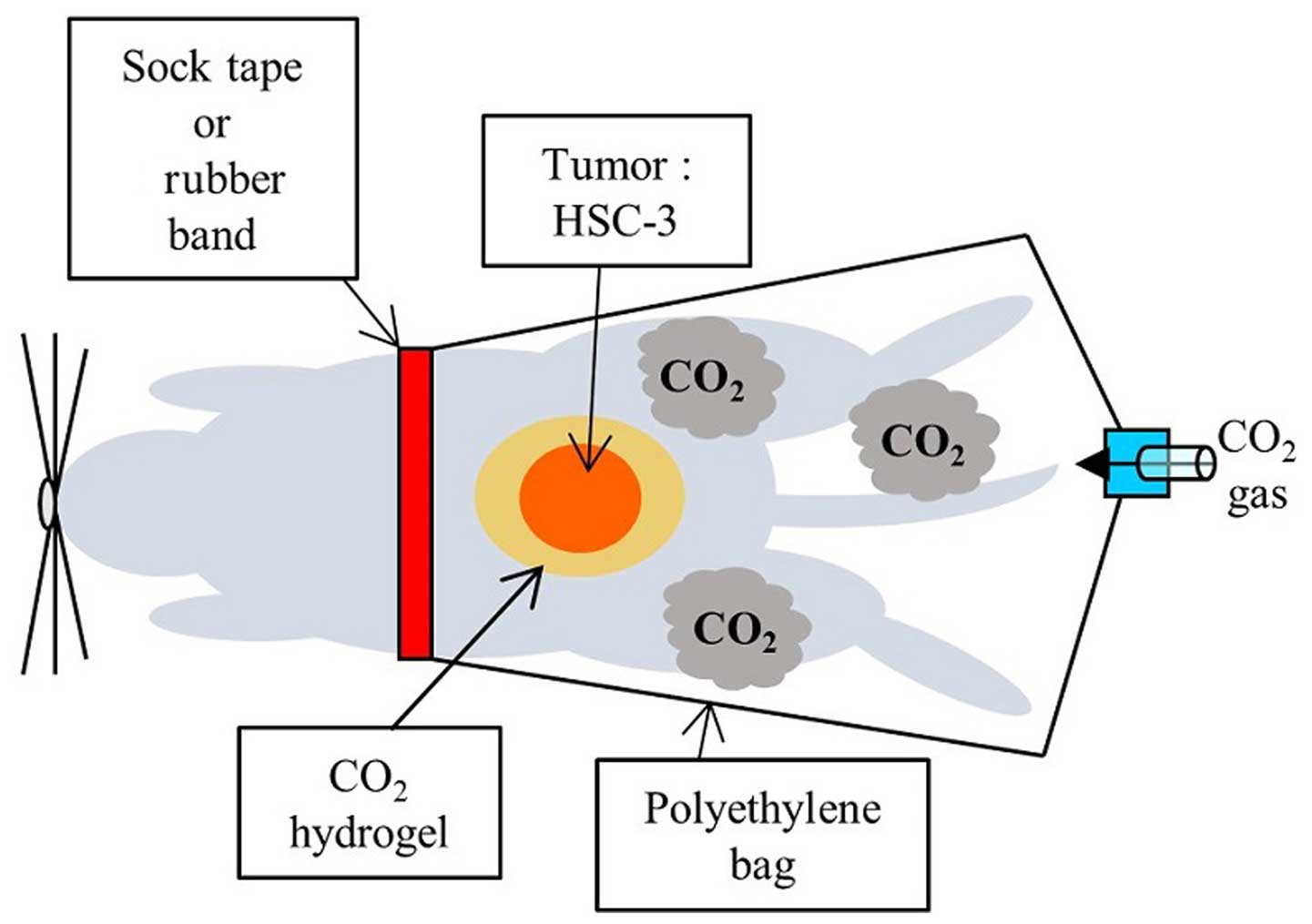
Briefly, the area of skin around the implanted tumor was covered
with CO2 hydrogel. This area was then sealed with a
polyethylene bag, and 100% CO2 gas was administered into
the bag (Fig. 1). Each treatment
was performed for 20 min. Control animals were treated similarly,
replacing the CO2 with room air.
Treatment commenced one week after HSC-3 cell
implantation and was performed twice a week for three weeks. After
24 h of final treatment, treated tumors were removed, and total RNA
and cell lysate were extracted from half of the tumor, immediately.
The other half of the tumor was formalin-fixed and
paraffin-embedded for staining. Serial 10-μm thick transverse
sections were prepared from each block.
Quantitative real-time PCR
The mRNA expression levels of β-actin, Snail,
Slug, E-cadherin, N-cadherin and Vimentin were analyzed
using quantitative real-time PCR. Total RNA was extracted from the
cells and treated tumors by selective binding to a silica-gel-based
membrane using an RNeasy Mini Kit, following the manufacturer's
instructions (Qiagen, Valencia, CA, USA). The cDNA was synthesized
(300 ng total RNA) using the High Capacity cDNA Transcription kit
(Applied Biosystems, Foster City, CA, USA).
Quantification of mRNA transcription was performed
using a StepOne Real-Time PCR System (Applied Biosystems).
Real-Time PCR was performed in a 20 μl reaction mixture using SYBR
Green Master Mix reagent (Applied Biosystems). PCR conditions were
as follows: 1 cycle at 95°C for 10 min followed by 40 cycles at
95°C for 15 sec and 60°C for 1 min. The expression of each target
gene was normalized to β-actin and relative expression to the
control group was calculated (the ΔΔCT method; Applied Biosystems).
All primers were obtained from Invitrogen (Carlsbad, CA, USA).
Primer sequences were as follows: β-actin, forward (5′-GAT
GAG ATT GGC ATG GCT TT-3′) and reverse (5′-CAC CTT CAC CGT TCC AGGT
TT-3′); Snail, forward (5′-TGC AGG ACT CTA ATC CAA AGT TTA
CC-3′) and reverse (5′-GAG GGA TGG CTG CCA GC-3′); Slug,
forward (5′-GTG TGG ACT ACC GCT GC-3′) and reverse (5′-TCC GGA AAG
AGG AGA GAG G-3′); E-cadherin, forward (5′-ACA GCA CGT ACA
CAG ACCC TA-3′) and reverse (5′-GCA GAA GTG TCC CTG TCC CAG-3′);
N-cadherin, forward (5′-TTG GAT CAA TGT CAT AAT CAA GTG CTG
TA -3′) and reverse (5′-CTC CTA TGA GTG GAA CAG GAA CG -3′);
Vimentin, forward (5′-AGC CGA AAA CAC CCT GCA AT-3′) and
reverse (5′-CGT TCA AGG GTC AAG ACG TGC-3′).
Immunoblot analysis
Cell lysates were prepared from the cells and
treated tumors using a whole cell lysis buffer supplemented with
Halt protease and phosphatase inhibitor cocktail (Mammalian Protein
Extraction Reagent; Thermo Scientific, Rockford, IL, USA). Protein
samples were processed using standard western immunoblot
procedures. Membranes were incubated overnight at 4°C with primary
antibodies in Can Get Signal® Immunoreaction Enhancer
Solution 1 (Toyobo, Osaka, Japan): anti-human-HIF-1α antibody
(1:1000) (Cell Signaling Technology, Danvers, MA, USA), anti-human
Snail antibody (1:1000) (Cell Signaling Technology), anti-human
Slug antibody (1:1000) (Cell Signaling Technology), anti-human
E-cadherin antibody (1:1000) (Cell Signaling Technology),
anti-human N-cadherin antibody (1:1000) (Cell Signaling
Technology), anti-human Vimentin antibody (1:1000) (Cell Signaling
Technology) and anti-human α-tubulin antibody (1:2000)
(Sigma-Aldrich). After washing, the membranes were incubated with
the appropriate secondary antibody conjugated to horseradish
peroxidase (GE Healthcare Bio-Sciences, Piscataway, NJ, USA) in Can
Get Signal Immunoreaction Enhancer Solution 2 (Toyobo), and exposed
using ECL Prime Plus Western Blotting Detection System Reagent (GE
Healthcare Bio-Sciences). The signals were detected using a
Chemilumino analyzer LAS-3000 mini (Fujifilm, Tokyo, Japan).
Immunohistochemical analysis
The formalin-fixed and paraffin-embedded tumor
sections were pretreated with citrate buffer for 40 min at 95°C,
quenched with 0.05% H2O2, and incubated
overnight at 4°C with the following primary antibodies in Can Get
Signal Immunostain Solution A (Toyobo): rabbit anti-human Snail
antibody (1:1000) (Cell Signaling Technology), anti-human Slug
antibody (1:1000) (Cell Signaling Technology), anti-human
E-cadherin antibody (1:1000) (Cell Signaling Technology),
anti-human N-cadherin antibody (1:1000) (Cell Signaling
Technology), and anti-human Vimentin antibody (1:1000) (Cell
Signaling Technology). Following the treatment, the sections were
incubated with horseradish peroxidase (HRP)-conjugated goat
anti-rabbit IgG polyclonal antibody (Nichirei Bioscience, Tokyo,
Japan) for 30 min at room temperature. Signals were developed as a
brown reaction product using peroxidase substrate
3,3′-diaminobenzidine (Nichirei Bioscience). The sections were
counterstained with hematoxylin and examined with a BZ-8000
confocal microscope (Keyence, Osaka, Japan).
Statistical analysis
Data are presented as the mean value ± standard
error. The results of the two groups were analyzed using the
Mann-Whitney U test. The level of statistical significance was set
at p<0.05.
Results
Improving hypoxic conditions decreases
the expression levels of HIF-1α and EMT factors in HSC-3 cells in
vitro
We first examined the in vitro effects of
changing oxygen levels on HSC-3 cells by culturing the cells in two
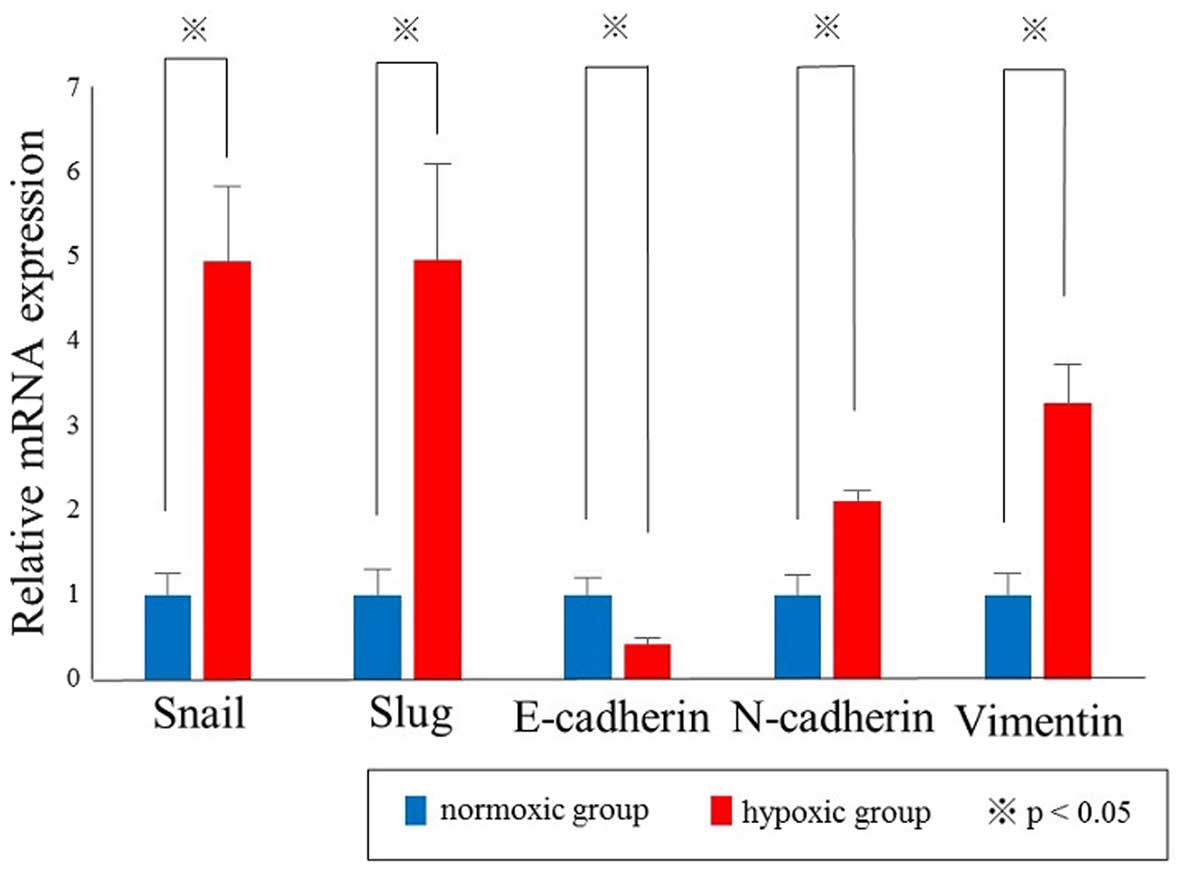
different oxygen conditions. Quantitative real-time PCR analyses
revealed that the relative mRNA expression levels of Snail,
Slug, N-cadherin and Vimentin were significantly
increased in hypoxic group compared with those in normoxic group,
whereas the expression of E-cadherin was significantly
decreased in hypoxic group compared with normoxic group (p<0.05,
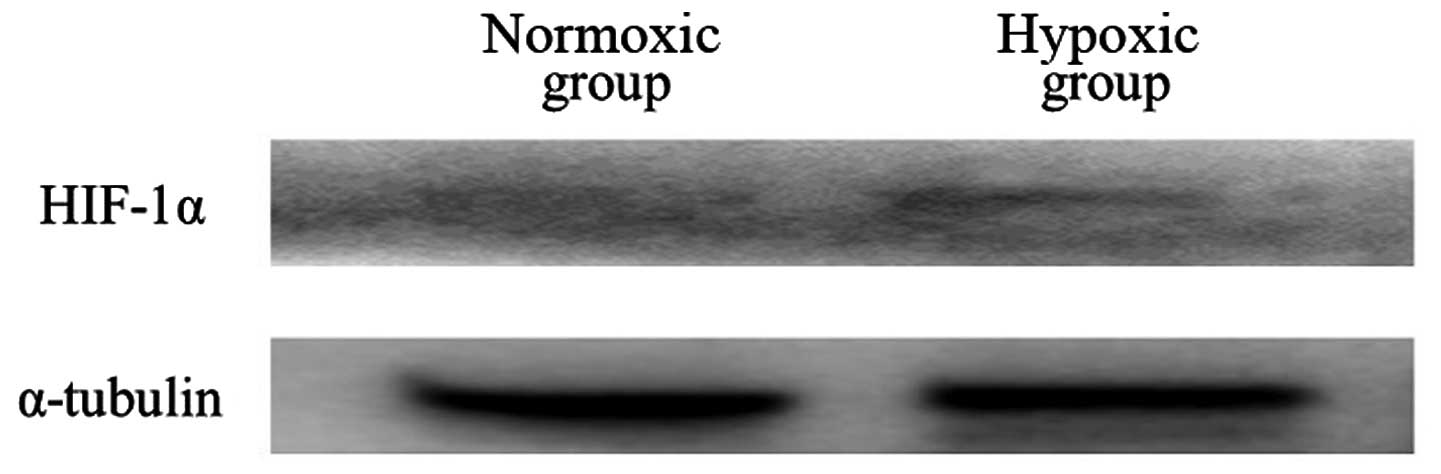
Fig. 2). Immunoblot analysis
showed that increased protein expression of HIF-1α was observed in
hypoxic group, but not in normoxic group (Fig. 3).
Transcutaneous CO2 suppresses
the expression levels of HIF-1α and EMT factors in HSC-3 cells in
vivo
We examined the in vivo effects of
transcutaneous CO2 on the expression levels of HIF-1 α
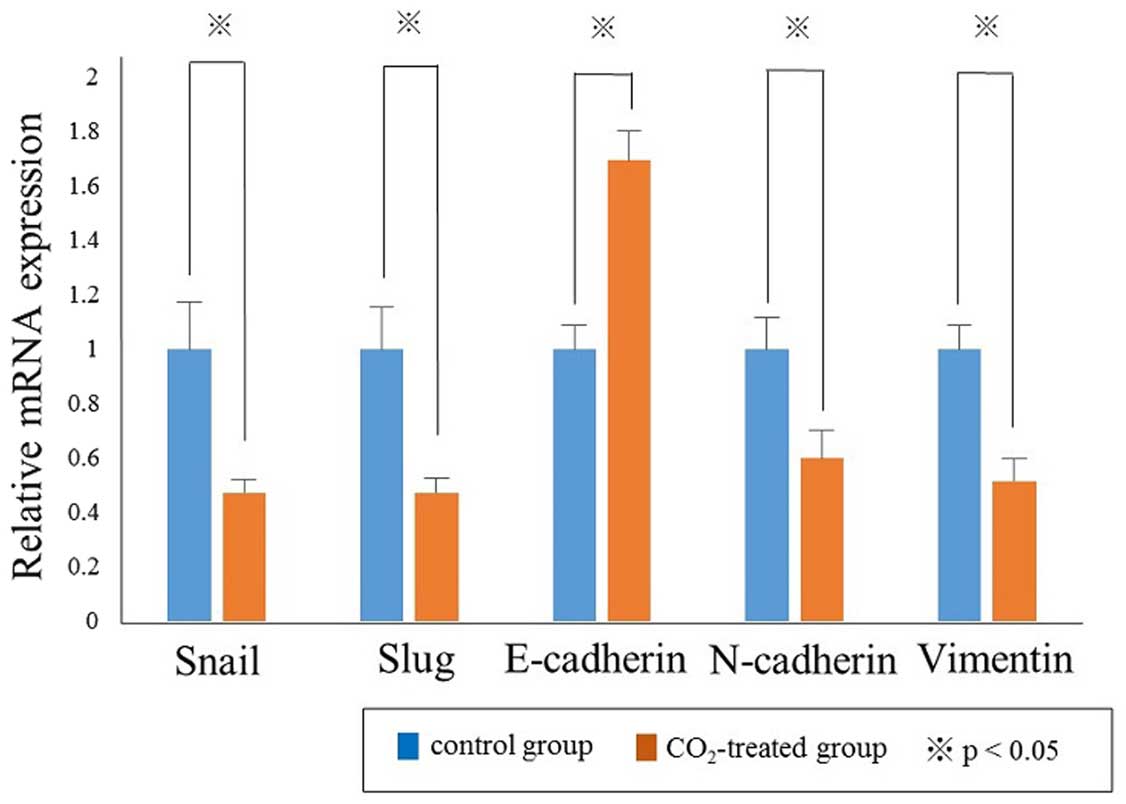
and EMT factors in HSC-3 cells. Quantitative real-time PCR analyses
revealed that the relative mRNA expression levels of Snail,
Slug, N-cadherin and Vimentin were significantly
decreased in the CO2-treated group compared with the
control group, in contrast the expression of E-cadherin was
significantly increased in the CO2-treated group
compared with the control group (p<0.05, Fig. 4). Consistent with the results from
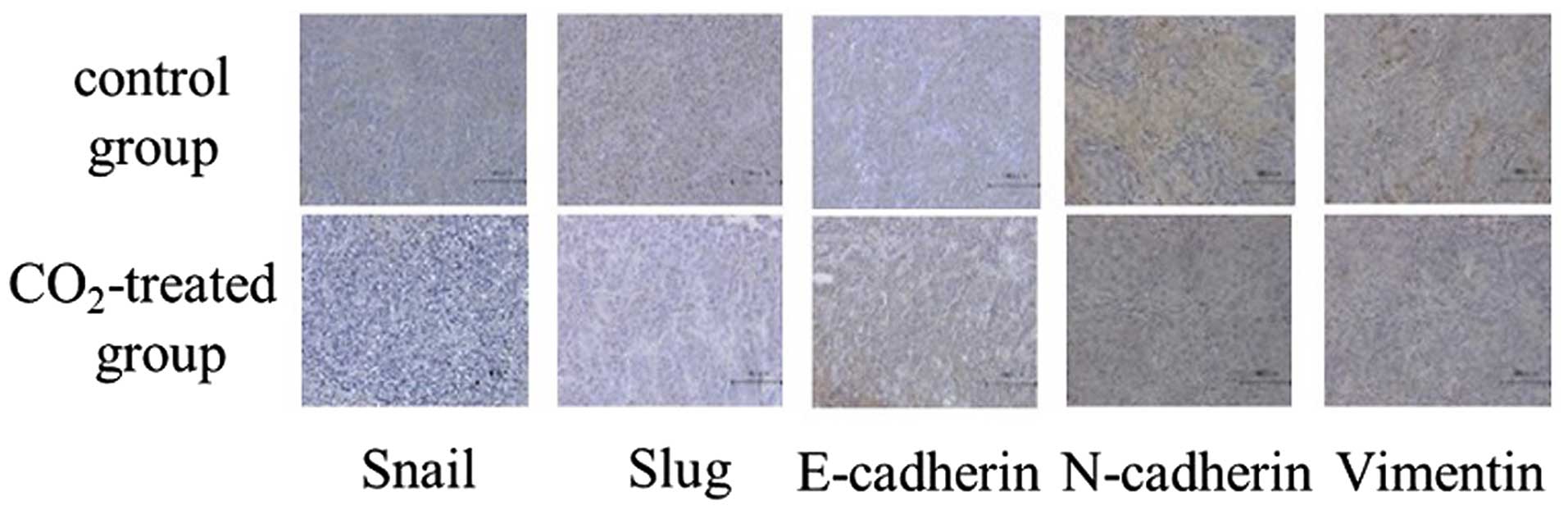
quantitative real-time PCR, immunohistochemical analysis revealed
that decreased expression levels in Snail, Slug, N-cadherin and
Vimentin expression with an increased E-cadherin expression was
observed in the CO2-treated tumors compared with the
control group (Fig. 5). In
addition, a western blot analysis showed that increased protein
expression of HIF-1α, Snail, Slug, N-cadherin and Vimentin were
observed in the control group, but not in the
CO2-treated group. In contrast, increased protein
expression of E-cadherin was observed in the CO2-treated
group (Fig. 6).
Discussion
OSCC is the most common malignancy in the oral
cavity, and is associated with more non-desirable outcomes than any
other oral disease because of the highly invasive nature of the
tumors and of the cervical lymph node metastasis (30).
Recent studies have demonstrated that the malignant
transformation of various carcinomas, including OSCC, is associated
with the loss of epithelial differentiation and the gain of the
mesenchymal phenotype, a process known as EMT (31,32).
EMT is a physiological phenomenon that involves a
loss of polarity and cell-cell adhesion, and that epithelial cells
gain characteristics of the mesenchymal cells, which include
enhanced motility and matrix resolution (32). EMT itself is an essential cellular
function which is involved in tissue construction during normal
developmental stages (33,34). Generally, cells undergoing EMT show
decreased expression of epithelial markers such as E-cadherin, and
increased expression of mesenchymal biomarkers, such as Snail,
Slug, N-cadherin and Vimentin (35–38).
E-cadherin is one of the major adhesion molecules of epithelial
cells. The decreased expression of E-cadherin is an indication of
invasion and metastasis, because it liberates carcinoma cells from
the primary site and enhances invasion and metastasis (39–41).
Thus during the EMT process, E-cadherin is the most important
factor, and its expression is a symbol of the occurrence of EMT.
Snail is a zinc finger protein and the first member of a
superfamily identified in Drosophila (42–44).
Snail 2, known as Slug, has been shown to function in a manner
similar to that of Snail; however, Slug displays a distinct tissue
distribution (45,46). Both Snail and Slug are considered
to be the major transcription factors involved in the modulation of
EMT in various types of cancer. They modulate EMT by repressing
E-cadherin transcription via binding to the E-cadherin promoter
(37). N-cadherin is one of the
mesenchymal cadherins, and N-cadherin upregulation has been
observed in many types of poorly differentiated tumors (47). In EMT process, ‘cadherin
switching’, the loss of E-cadherin expression and the gain of
N-cadherin expression, occurs, upon Snail/Slug expression.
Vimentin, which is also promoted by Snail and Slug, is a major
protein constituent of the intermediate filaments in normal and
neoplastic mesenchymal cells (48). Through the degeneration and
production of these factors, the conditions required to generate
EMT are established.
Many studies have shown the relationship between the
expression levels of EMT factors and the survival rates of patients
with SCC (including patients with OSCC) (9–19).
The decreased expression of E-cadherin was found to be
significantly associated with the survival rates of patients with
various cancers, including OSCC (9,10),
ovarian cancer (11), gastric
cancer (12), prostate cancer
(13), colon cancer (14), and lung cancer (15). The expression levels of other
important EMT factors, Snail (16), Slug (17), N-cadherin (18), and Vimentin (19), are also closely correlated with the
prognosis of patients with OSCC. Moreover, the expression levels of
these EMT factors are said to be closely associated with tumor
differentiation, tumor stage, lymph node metastasis and recurrence
in patients with OSCC (9,10,16–19,49–51).
Hence, there is a possibility that EMT suppression may improve the
prognosis of patients with OSCC.
Hypoxia, which is a common condition in solid
tumors, can increase the invasiveness and the metastatic ability of
tumor cells (52). Under hypoxic
conditions, HIF-1α is activated in tumor cells; the activation
promotes tumor growth, whereas the loss of HIF-1α activity markedly
decreases tumor growth, angiogenesis, and cellular energy
metabolism (53). Recent studies
have shown that HIF-1α is a crucial microenvironmental factor that
induces the expression levels of EMT factors such as Snail and
Vimentin (22–26).
Based on these facts, we hypothesized that oxygen
conditions may affect the expression levels of HIF-1α and EMT
factors in OSCC. We first examined the in vitro effects of
oxygen levels on HSC-3 cells by culturing the cells in two
different oxygen conditions. We revealed that HIF-1α expression was
increased in HSC-3 cells under hypoxic conditions, and in addition,
the increased expression of mesenchymal biomarkers with the
decreased expression of epithelial markers was observed in hypoxic
HSC-3 cells. Consistent with this, Zhang et al have reported
that HIF-1α expression was increased under hypoxic conditions in
comparison the expression under normoxic conditions in OSCC in
vitro (54). These results
indicate that hypoxic condition could affect the expression of EMT
factors in OSCC cells in vitro.
We previously reported that the transcutaneous
CO2 system improves hypoxia in treated tissue,
potentially resulting in an ‘artificial Bohr effect’ (55). Moreover, we have demonstrated that
transcutaneous CO2 suppresses human OSCC growth and
metastasis to the regional lymph nodes by both improving hypoxia in
treated tissue and increasing mitochondrial apoptosis (27). However, the molecular mechanism of
decreased metastatic potential by transcutaneous CO2 has
not been addressed.
Based on previous studies, we hypothesized that
transcutaneous CO2 could decrease the metastatic
potential to lymph nodes by EMT suppression via improving hypoxia
in treated tissue. We examined the in vivo effects of
transcutaneous CO2 on the expression levels of HIF-1α
and EMT factors using human OSCC xenograft models. Our results
showed that transcutaneous CO2 increased the expression
levels of epithelial markers with decreased expression levels of
HIF-1α and mesenchymal biomarkers, resulting in EMT
suppression.
In the present study, we found that hypoxic
conditions increased the expression levels of HIF-1α and EMT
factors in OSCC in vitro, and that oxygenation using
transcutaneous CO2 suppressed EMT by improving hypoxia
in OSCC in vivo. These results suggest that transcutaneous
CO2 could suppress EMT by improving hypoxia and that it
may reduce the metastatic potential of OSCC. Our present findings
strongly indicate that transcutaneous CO2 may be able to
improve the prognosis of patients with OSCC through the suppression
of EMT. In conclusion, this is the first report to show that
transcutaneous CO2 suppresses the expression levels of
EMT factors in human OSCC. Our findings indicate that
transcutaneous CO2 suppresses EMT by improving hypoxia
and that it could reduce the metastatic potential of OSCC.
Acknowledgements
We would like to thank Minako Nagata, Maya Yasuda
and Kyoko Tanaka for their expert technical assistance.
References
|
1
|
Warnakulasuriya S: Global epidemiology of
oral and oropharyngeal cancer. Oral Oncol. 45:309–316. 2009.
View Article : Google Scholar
|
|
2
|
Kume K, Haraguchi M, Hijioka H, Ishida T,
Miyawaki A, Nakamura N and Ozawa M: The transcription factor Snail
enhanced the degradation of E-cadherin and desmoglein 2 in oral
squamous cell carcinoma cells. Biochem Biophys Res Commun.
430:889–894. 2013. View Article : Google Scholar
|
|
3
|
Sasahira T, Kirita T, Yamamoto K, Ueda N,
Kurihara M, Matsushima S, Bhawal UK, Bosserhoff AK and Kuniyasu H:
Transport and Golgi organisation protein 1 is a novel tumour
progressive factor in oral squamous cell carcinoma. Eur J Cancer.
50:2142–2151. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Savagner P: Leaving the neighborhood:
Molecular mechanisms involved during epithelial-mesenchymal
transition. BioEssays. 23:912–923. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Voulgari A and Pintzas A:
Epithelial-mesenchymal transition in cancer metastasis: Mechanisms,
markers and strategies to overcome drug resistance in the clinic.
Biochim Biophys Acta. 1796:75–90. 2009.PubMed/NCBI
|
|
7
|
Grille SJ, Bellacosa A, Upson J,
Klein-Szanto AJ, van Roy F, Lee-Kwon W, Donowitz M, Tsichlis PN and
Larue L: The protein kinase Akt induces epithelial mesenchymal
transition and promotes enhanced motility and invasiveness of
squamous cell carcinoma lines. Cancer Res. 63:2172–2178.
2003.PubMed/NCBI
|
|
8
|
Ikenouchi J, Matsuda M, Furuse M and
Tsukita S: Regulation of tight junctions during the
epithelium-mesenchyme transition: Direct repression of the gene
expression of claudins/occludin by Snail. J Cell Sci.
116:1959–1967. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
da Silva SD, Morand GB, Alobaid FA, Hier
MP, Mlynarek AM, Alaoui-Jamali MA and Kowalski LP:
Epithelial-mesenchymal transition (EMT) markers have prognostic
impact in multiple primary oral squamous cell carcinoma. Clin Exp
Metastasis. 32:55–63. 2015. View Article : Google Scholar
|
|
10
|
Fan CC, Wang TY, Cheng YA, Jiang SS, Cheng
CW, Lee AY and Kao TY: Expression of E-cadherin, Twist, and p53 and
their prognostic value in patients with oral squamous cell
carcinoma. J Cancer Res Clin Oncol. 139:1735–1744. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Faleiro-Rodrigues C, Macedo-Pinto I,
Pereira D and Lopes CS: Prognostic value of E-cadherin
immunoexpression in patients with primary ovarian carcinomas. Ann
Oncol. 15:1535–1542. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou YN, Xu CP, Han B, Li M, Qiao L, Fang
DC and Yang JM: Expression of E-cadherin and beta-catenin in
gastric carcinoma and its correlation with the clinicopatho-logical
features and patient survival. World J Gastroenterol. 8:987–993.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Richmond PJ, Karayiannakis AJ, Nagafuchi
A, Kaisary AV and Pignatelli M: Aberrant E-cadherin and
alpha-catenin expression in prostate cancer: Correlation with
patient survival. Cancer Res. 57:3189–3193. 1997.PubMed/NCBI
|
|
14
|
Bondi J, Bukholm G, Nesland JM, Bakka A
and Bukholm IR: An increase in the number of adhesion proteins with
altered expression is associated with an increased risk of cancer
death for colon carcinoma patients. Int J Colorectal Dis.
21:231–237. 2006. View Article : Google Scholar
|
|
15
|
Tseng RC, Lee SH, Hsu HS, Chen BH, Tsai
WC, Tzao C and Wang YC: SLIT2 attenuation during lung cancer
progression deregulates beta-catenin and E-cadherin and associates
with poor prognosis. Cancer Res. 70:543–551. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li YY, Zhou CX and Gao Y: Snail regulates
the motility of oral cancer cells via RhoA/Cdc42/p-ERM pathway.
Biochem Biophys Res Commun. 452:490–496. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wushou A, Pan HY, Liu W, Tian Z, Wang LZ,
Shali S and Zhang ZY: Correlation of increased twist with lymph
node metastasis in patients with oral squamous cell carcinoma. J
Oral Maxillofac Surg. 70:1473–1479. 2012. View Article : Google Scholar
|
|
18
|
Zhao D, Tang XF, Yang K, Liu JY and Ma XR:
Over-expression of integrin-linked kinase correlates with aberrant
expression of Snail, E-cadherin and N-cadherin in oral squamous
cell carcinoma: Implications in tumor progression and metastasis.
Clin Exp Metastasis. 29:957–969. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou J, Tao D, Xu Q, Gao Z and Tang D:
Expression of E-cadherin and vimentin in oral squamous cell
carcinoma. Int J Clin Exp Pathol. 8:3150–3154. 2015.PubMed/NCBI
|
|
20
|
Hockel M, Schlenger K, Aral B, Mitze M,
Schaffer U and Vaupel P: Association between tumor hypoxia and
malignant progression in advanced cancer of the uterine cervix.
Cancer Res. 56:4509–4515. 1996.PubMed/NCBI
|
|
21
|
Jing SW, Wang YD, Chen LQ, Sang MX, Zheng
MM, Sun GG, Liu Q, Cheng YJ and Yang CR: Hypoxia suppresses
E-cadherin and enhances matrix metalloproteinase-2 expression
favoring esophageal carcinoma migration and invasion via hypoxia
inducible factor-1 alpha activation. Dis Esophagus. 26:75–83. 2013.
View Article : Google Scholar
|
|
22
|
Krishnamachary B, Zagzag D, Nagasawa H,
Rainey K, Okuyama H, Baek JH and Semenza GL: Hypoxia-inducible
factor-1-dependent repression of E-cadherin in von Hippel-Lindau
tumor suppressor-null renal cell carcinoma mediated by TCF3,
ZFHX1A, and ZFHX1B. Cancer Res. 66:2725–2731. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Krishnamachary B, Berg-Dixon S, Kelly B,
Agani F, Feldser D, Ferreira G, Iyer N, LaRusch J, Pak B, Taghavi
P, et al: Regulation of colon carcinoma cell invasion by
hypoxia-inducible factor 1. Cancer Res. 63:1138–1143.
2003.PubMed/NCBI
|
|
24
|
Lv L, Yuan J, Huang T, Zhang C, Zhu Z,
Wang L, Jiang G and Zeng F: Stabilization of Snail by HIF-1α and
TNF-α is required for hypoxia-induced invasion in prostate cancer
PC3 cells. Mol Biol Rep. 41:4573–4582. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang F, Chang M, Shi Y, Jiang L, Zhao J,
Hai L, Sharen G and Du H: Down-regulation of hypoxia-inducible
factor-1 suppresses malignant biological behavior of
triple-negative breast cancer cells. Int J Clin Exp Med.
7:3933–3940. 2014.
|
|
26
|
Wang N, Dong CR, Jiang R, Tang C, Yang L,
Jiang QF, Chen GG and Liu ZM: Overexpression of HIF-1α,
metallothionein and SLUG is associated with high TNM stage and
lymph node metastasis in papillary thyroid carcinoma. Int J Clin
Exp Pathol. 7:322–330. 2013.
|
|
27
|
Takeda D, Hasegawa T, Ueha T, Imai Y,
Sakakibara A, Minoda M, Kawamoto T, Minamikawa T, Shibuya Y, Akisue
T, et al: Transcutaneous carbon dioxide induces mitochondrial
apoptosis and suppresses metastasis of oral squamous cell carcinoma
in vivo. PLoS One. 9:e1005302014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Matsui T, Ota T, Ueda Y, Tanino M and
Odashima S: Isolation of a highly metastatic cell line to lymph
node in human oral squamous cell carcinoma by orthotopic
implantation in nude mice. Oral Oncol. 34:253–256. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Okada Y, Akisue T, Hara H, Kishimoto K,
Kawamoto T, Imabori M, Kishimoto S, Fukase N, Onishi Y and Kurosaka
M: The effect of bevacizumab on tumour growth of malignant fibrous
histiocytoma in an animal model. Anticancer Res. 30:3391–3395.
2010.PubMed/NCBI
|
|
30
|
Jordan RC and Daley T: Oral squamous cell
carcinoma: new in sights. J Can Dent Assoc. 63:517–518. 521–525.
1997.
|
|
31
|
Christiansen JJ and Rajasekaran AK:
Reassessing epithelial to mesenchymal transition as a prerequisite
for carcinoma invasion and metastasis. Cancer Res. 66:8319–8326.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Smith A, Teknos TN and Pan Q: Epithelial
to mesenchymal transition in head and neck squamous cell carcinoma.
Oral Oncol. 49:287–292. 2013. View Article : Google Scholar :
|
|
33
|
Acloque H, Adams MS, Fishwick K,
Bronner-Fraser M and Nieto MA: Epithelial-mesenchymal transitions:
The importance of changing cell state in development and disease. J
Clin Invest. 119:1438–1449. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huber MA, Kraut N and Beug H: Molecular
requirements for epithelial-mesenchymal transition during tumor
progression. Curr Opin Cell Biol. 17:548–558. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wheelock MJ, Shintani Y, Maeda M, Fukumoto
Y and Johnson KR: Cadherin switching. J Cell Sci. 121:727–735.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Peinado H, Olmeda D and Cano A: Snail, Zeb
and bHLH factors in tumour progression: An alliance against the
epithelial phenotype? Nat Rev Cancer. 7:415–428. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Peinado H, Portillo F and Cano A:
Transcriptional regulation of cadherins during development and
carcinogenesis. Int J Dev Biol. 48:365–375. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mattijssen V, Peters HM, Schalkwijk L,
Manni JJ, van ‘t Hof-Grootenboer B, de Mulder PH and Ruiter DJ:
E-cadherin expression in head and neck squamous-cell carcinoma is
associated with clinical outcome. Int J Cancer. 55:580–585. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Thomas PA, Kirschmann DA, Cerhan JR,
Folberg R, Seftor EA, Sellers TA and Hendrix MJ: Association
between keratin and vimentin expression, malignant phenotype, and
survival in postmenopausal breast cancer patients. Clin Cancer Res.
5:2698–2703. 1999.PubMed/NCBI
|
|
41
|
Yokoyama K, Kamata N, Hayashi E, Hoteiya
T, Ueda N, Fujimoto R and Nagayama M: Reverse correlation of
E-cadherin and snail expression in oral squamous cell carcinoma
cells in vitro. Oral Oncol. 37:65–71. 2001. View Article : Google Scholar
|
|
42
|
Tang CH and Tsai CC: CCL2 increases MMP-9
expression and cell motility in human chondrosarcoma cells via the
Ras/Raf/ MEK/ERK/NF-κB signaling pathway. Biochem Pharmacol.
83:335–344. 2012. View Article : Google Scholar
|
|
43
|
Barrallo-Gimeno A and Nieto MA:
Evolutionary history of the Snail/Scratch superfamily. Trends
Genet. 25:248–252. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nieto MA: The snail superfamily of
zinc-finger transcription factors. Nat Rev Mol Cell Biol.
3:155–166. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
45
|
Parent AE, Newkirk KM and Kusewitt DF:
Slug (Snai2) expression during skin and hair follicle development.
J Invest Dermatol. 130:1737–1739. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Murray SA, Oram KF and Gridley T: Multiple
functions of Snail family genes during palate development in mice.
Development. 134:1789–1797. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li G, Satyamoorthy K and Herlyn M:
N-cadherin-mediated intercellular interactions promote survival and
migration of melanoma cells. Cancer Res. 61:3819–3825.
2001.PubMed/NCBI
|
|
48
|
Vuoriluoto K, Haugen H, Kiviluoto S,
Mpindi JP, Nevo J, Gjerdrum C, Tiron C, Lorens JB and Ivaska J:
Vimentin regulates EMT induction by Slug and oncogenic H-Ras and
migration by governing Axl expression in breast cancer. Oncogene.
30:1436–1448. 2011. View Article : Google Scholar
|
|
49
|
Hashimoto T, Soeno Y, Maeda G, Taya Y,
Aoba T, Nasu M, Kawashiri S and Imai K: Progression of oral
squamous cell carcinoma accompanied with reduced E-cadherin
expression but not cadherin switch. PLoS One. 7:e478992012.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mandal M, Myers JN, Lippman SM, Johnson
FM, Williams MD, Rayala S, Ohshiro K, Rosenthal DI, Weber RS,
Gallick GE, et al: Epithelial to mesenchymal transition in head and
neck squamous carcinoma: Association of Src activation with
E-cadherin down-regulation, vimentin expression, and aggressive
tumor features. Cancer. 112:2088–2100. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhang S, Zhou X, Wang B, Zhang K, Liu S,
Yue K, Zhang L and Wang X: Loss of VHL expression contributes to
epithelial-mesenchymal transition in oral squamous cell carcinoma.
Oral Oncol. 50:809–817. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Teppo S, Sundquist E, Vered M, Holappa H,
Parkkisenniemi J, Rinaldi T, Lehenkari P, Grenman R, Dayan D,
Risteli J, et al: The hypoxic tumor microenvironment regulates
invasion of aggressive oral carcinoma cells. Exp Cell Res.
319:376–389. 2013. View Article : Google Scholar
|
|
53
|
Wu XY, Fu ZX and Wang XH: Effect of
hypoxia-inducible factor 1-α on Survivin in colorectal cancer. Mol
Med Rep. 3:409–415. 2010.
|
|
54
|
Zhang Q, Zhang ZF, Rao JY, Sato JD, Brown
J, Messadi DV and Le AD: Treatment with siRNA and antisense
oligonucleotides targeted to HIF-1alpha induced apoptosis in human
tongue squamous cell carcinomas. Int J Cancer. 111:849–857. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sakai Y, Miwa M, Oe K, Ueha T, Koh A,
Niikura T, Iwakura T, Lee SY, Tanaka M and Kurosaka M: A novel
system for transcutaneous application of carbon dioxide causing an
‘artificial Bohr effect’ in the human body. PLoS One. 6:e241372011.
View Article : Google Scholar
|