Gastric cancer, one of the most common malignancies
and causes almost one million new cases each year in the world
(1). To make matters worse,
despite decreasing incidence over the recent decades, GC remains
the third leading cause of global cancer-related death preceded by
lung and liver cancer and maintains the highest morbidity in
Eastern Asia (2,3). The main reasons for failure and
mortality of gastric cancer are cell infiltration and metastasis.
Metastatic cancer cells fractionally retain their epithelial
properties and obtain mesenchymal characteristics which give them
the ability to invade or distract. E-cadherin is a major protein
prominently required for cell-cell junctions and polarity, whose
loss has been established to be the principal event in metastatic
progression and tumor invasion. Moreover, downregulation of
E-cadherin might induce the occurrence of EMT, a multistage process
giving rise to the transformation of polar epithelial cells to
mesenchymal phenotype and authorizes cell migration as well
(4,5).
Two decades ago, Heuberger and Birchmeier made the
first ascertainment to expound that the presence of
E-cadherin/β-catenin complex allows for steady cell junctions
(6). β-catenin, one key protein in
Wnt signaling, separates from the degradation complex and then
accumulates in the cytoplasm, translocates into nucleus
subsequently to sensitize the Wnt pathway during carcinoma
progression. In the presence of downregulation of E-cadherin and
Wnt signals activation, both cytoplasmic and nuclear levels of
β-catenin increase where it interacts with cell growth and EMT
(7,8). MicroRNAs are small non-coding
molecules with 19–25 nucleotides that bind to 3′UTR of target mRNAs
with complementary base pairing to inhibit their translation or
speed up decomposition. Latest experiments investigated either
suppressive or promotive function of crosstalk between miRNAs and
certain signaling pathways during EMT and metastasis, in spite of
this, little attention has been paid to the interplay between
Wnt/β-catenin signaling and miRNAs in regulating EMT in gastric
cancer (9). Taking efforts to
comprehend the mechanism of Wnt adjustment by miRNAs may benefit
our perspectives of reversing the course of EMT or potential
therapeutic targets in GC. Therefore, this review focuses on
crosstalk between microRNAs and Wnt pathway in the course of
modulating EMT in gastric cancer.
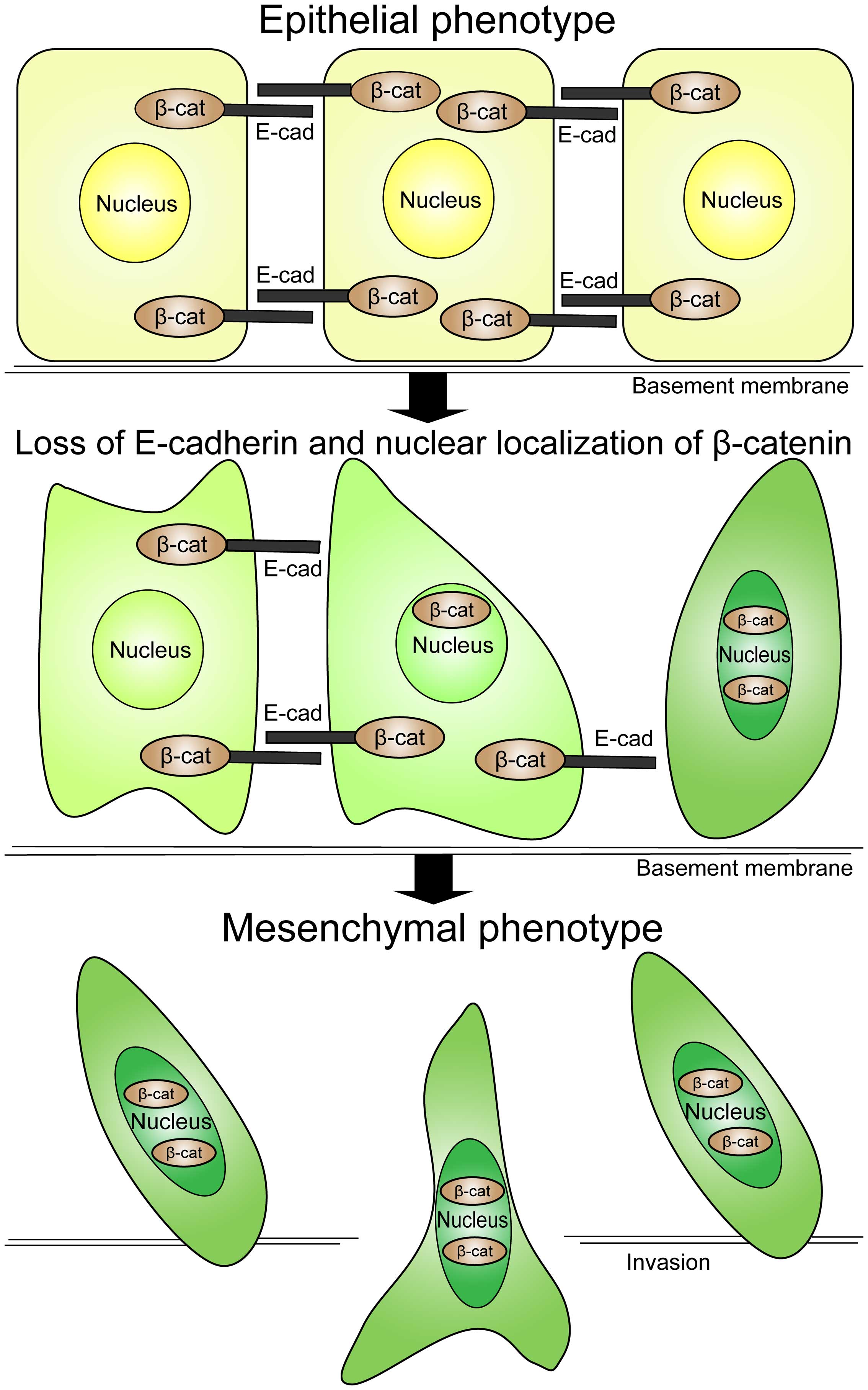
Epithelial-mesenchymal transition, firstly put
forward by Greenburg and Hay in 1982 (10), has been considered to be the
feature of embryonic development. However, emerging evidence has
indicated that the occurrence of EMT is critical for the initiation
and progression of diverse tumors. An EMT process is distinguished
by loss of E-cadherin and it creates profound phenotypic
alterations through which epithelial cells with apico-basal
polarity convert into front-rear polarity to gain mesenchymal
characteristics as well as the capacity of migration, invasion and
apoptotic resistance (11)
(Fig. 1). The epithelial cells
going through EMT become invasive and then migrate into distant
tissues where they experience a reverting mechanism
mesenchymal-epithelial transition (MET), afterward these cells
retrieve membrane junctions and form metastatic focus (12). EMT is characterized by the mutative
expression of three distinct families of protein consist of
cadherins (cell membrane surface protein), Vimentin (cytoskeletal
protein) and transcription factors including Snail, Twist, and ZEB
(13,14).
E-cadherin, encoded by CDH1, interacts with α, β, γ
and P120-catenin to constitute E-cadherin/catenin complex that
plays a critical role in adherens junctions between epithelial
cells (15). Loss of E-cadherin on
account of promoter hypermethylation has been widely considered to
correlate with GC aggressiveness and metastasis as well as
unfavorable prognosis of patients (16,17).
Hansford et al (18)
reported that HDGC families, particularly over the age of 80, may
be best defined by mutations in CDH1 and different clinical
ramifications, and that CDH1 mutation might provide more accurate
estimations of age-connected risk of GC. Nevertheless, Li et
al (19) demonstrated that
CDH1 promoter hypermethylation probably contributes to E-cad
reduction in sporadic instead of hereditary gastric cancer. It is
widely acknowledged that in the presence of deviant EMT, E-cadherin
switches to N-cadherin which is generally distributed in neural and
mesenchymal cells and has ability to expedite the degeneration of
stroma as well as cell movement. Jun et al suggested that
median survival is significantly correlated with N-cadherin
expression in cases of gastric cancer with brain metastasis
(20). Similarly, Kamikihara et
al (21) performed a genetic
association study in 146 gastric cancer patients to analyze the
relationship between N-cadherin expression and clinicopathological
features. The result exhibited that N-cadherin expression
positively correlates to hematogenous relapse while negatively
connects to patients' postoperative outcome independent of
E-cadherin expression, even if N-cadherin has no association with
tumor histology or lymphatic invasion.
Vimentin, a hallmark of interstitial cells, is known
to be tightly associated with the potential of tumor cell
invasiveness and migration. Vimentin expresses obviously higher
diffuse type than intestinal in patients with gastric
adenocarcinoma and it might be a feasible mark of relapse, far
metastasis or even decreased survival (22). Shirahata et al (23) recommended we use quantitative
methylation-specific PCR (qMSP), a sensitive technique that can
catch as few as one methylated gene copy out from 1,000
unmethylated ones, to detect promoter methylation in the serum of
GC patients. In addition, they recorded a high correlation between
Vim methylation and gastric carcinogenesis.
The zinc finger protein Snail consists of three
members containing Snail1, Snail2 (Slug) and Snail3 (Smuc), among
which Snail1 and Snail2 act synergistically in cases of gastric
tumor. Snail is initially recognized to facilitate EMT by way of
distinguishing E-box motifs in CDH1 to inhibit E-cadherin, whereas
growing indication has inferred the mechanism involves not only
downregulation of claudins, occludins, cytokeratins but also
upregulation of fibronectin and vitrinectin (24). Besides, the involvement of
NF-κB-Snail-E-cadherin axis or CCR7 pathway in induction of EMT
through upregulating Snail signaling proved in GC both implied the
decisive role of Snail explicitly (25,26).
Furthermore, Shin et al (27) reported that Snail could be a
prognostic predictor of gastric cancer due to its strong
interrelationship with tumor progression, lymph node metastasis,
lymphovascular or perineural invasion. Twist protein is a member of
the basic/helix-loop-helix transcription factor family and binds
selectively to the E-box consensus sequence to regulate target
genes such as E-cadherin (28). A
series of experiments have illustrated the considerable function of
Twist as an oncogene in vivo, suggestive of its potency to
be a target for the treatment of gastric carcinoma. Liu et
al (29) discovered that Twist
is expressed much higher in gastric cancer tissues compared to
adjacent normal tissues at mRNA and protein levels by means of
RT-PCR and western blotting. Likewise, abnormal Twist expression
related strongly to lower 5-year survival rates in patients of GC
in the Ru et al study, yet, they emphasized that the
significance disappears in cases of stage IV (30). Various examinations in vitro
collaboratively implied the carciogenesis of Twist. GC cells
transfected with Twist1 accelerated cell cycle progression by
recruiting p300 to enhance FoxM1 gene expression, whereas those
treated with Twist siRNA reversely experienced obvious cell cycle
arrest at G0/G1 phase, apoptosis induction as well as decreased
ability of proliferation and invasion (31,32).
Apart from this, Twist was proved to be a pivotal gene of
Helicobacter pylori-triggered GC cell stemness while
decreased after H. pylori eradication (33), reminding us to excavate therapeutic
targets against gastric cancer via Twist-dependent mechanism. ZEB
family members, ZEB1 and ZEB2, are transcriptional factors
suppressing E-cadherin, and they play a decisive role in tumor
metastasis. Substantial evidence showed that intense expression of
ZEB1 associates with both primary gastric cancer and peritoneal
diffusion which predicted earlier recurrence as well as lower
disease-free survival rate (34,35).
Murai et al (36)
classified 116 GC patients into epithelial or mesenchymal group
through Vimentin/E-cadherin ratio (V/E), as they pointed, the
mesenchymal group with high V/E ratio indicated an EMT status and
carried anomalous ZEB1 expression, suggesting aggressive
clinicopathological characteristics and poorer survival. Moreover,
less AGS cells passed through the Transwell chamber which signified
reduced capability of invasion in response to transfection with
ZEB1 siRNA (37). ZEB2 can also
bind to the E-box of E-cadherin gene promoter similarly to the
Snail family. In light of the exploration by Dai et al, ZEB2
was closely tied to poor clinicopathological parameters such as
depth of invasion, lymph node metastasis, TNM stage along with
higher expression of matrix metalloprotease (MMP) family (38). A recent study from Korea on brain
metastatic gastric adenocarcinoma argued that four of eight primary
GC cases represent evident nuclear ZEB2 expression. Nevertheless, 3
cases showed decreased nuclear ZEB2 in brain metastatic gastric
cancer samples corresponding to the consequence of brain metastatic
adenocarcinoma cell research, which might result from the complex
mechanisms of brain metastasis or small sample size due to low
incidence (39). However, this
finding might support the theory of MET process that anchors
circulating cancer cells to metastatic organs, although the effect
of this in metastatic dissemination is still under debate (40).
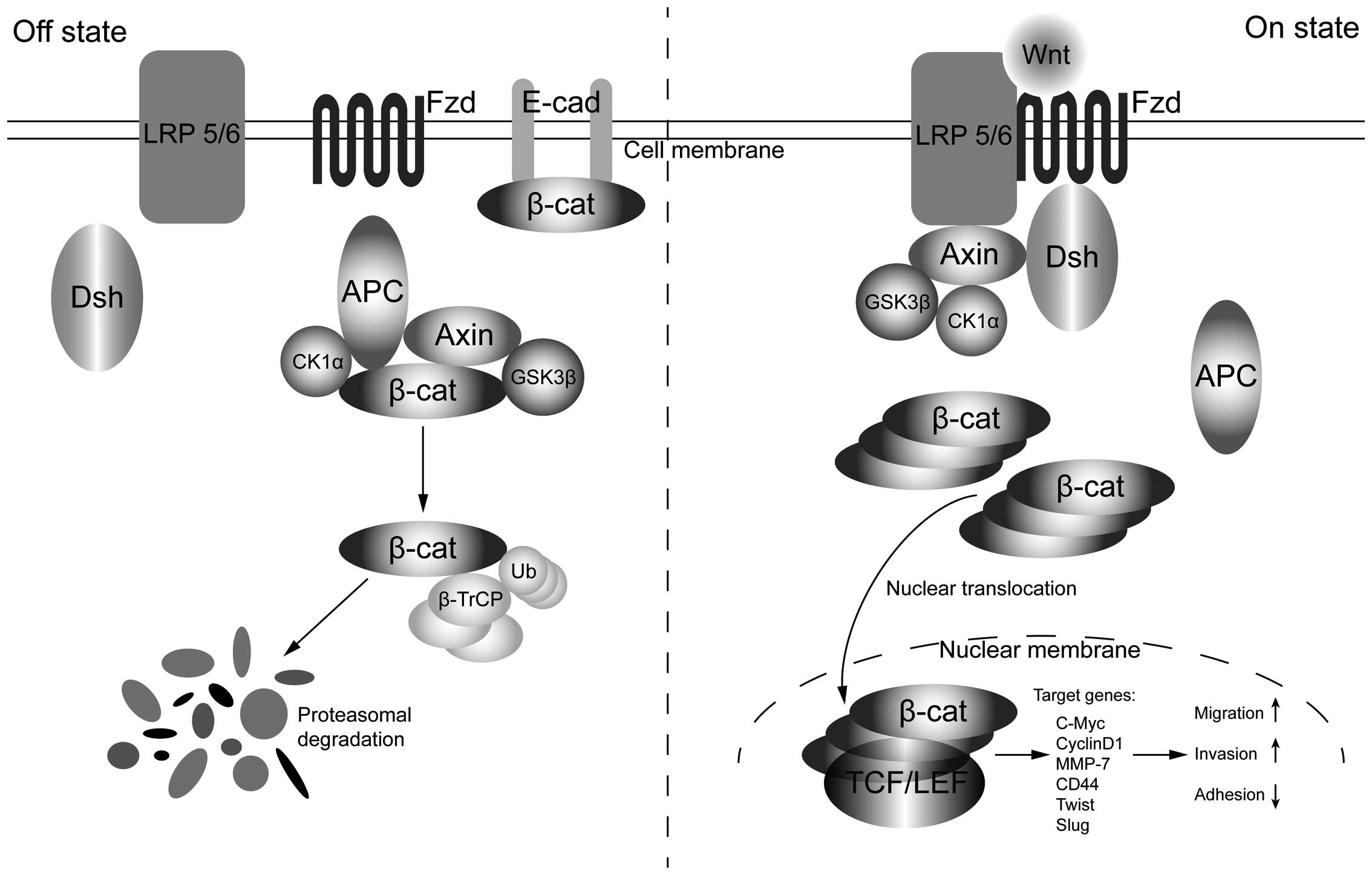
The Wnt pathway named from wingless and Int-1, plays
a foremost role in cell proliferation, differentiation, adhesion,
migration and stem cell self-renewal. This signaling could be
divided into two categories through whether it is β-catenin
dependent (canonical) or β-catenin independent (non-canonical).
Normally, cytoplasmic β-catenin is trussed up by the destruction
complex made up of Axin, adenomatous polyposis coli (APC), casein
kinase 1α (CK1α) and glycogen synthase kinase 3β (GSK3β), which is
distinguished by the E3 ubiquitin ligase β-TrCP and the proteosomal
degradation ultimately (51). The
steady state is destroyed when secreted Wnt ligands bind to
receptors on the surface of cell membrane mainly refer to as
Frizzled (Fz)/low-density lipoprotein receptor-related protein 5/6
(LRP5/6) receptor compound. Afterwards, Dishevelled (Dsh) protein
gets phosphorylated and forms a complex with Axin, which
subsequently binds GSK-3β so as to block its activation (52). This association further results in
decomposition of the degradation complex and β-catenin gathers in
the cytoplasm (53). Cumulative
β-catenin translocates into the nucleus, which has been generally
acknowledged to be the principle event of canonical Wnt pathway
activation, next interacts with TCF/LEF to compose a complex of
activators and accordingly set off the transcription of target
genes of Wnt/β-catenin signaling including c-Myc, cyclinD1, and
MMP-7 (54) (Fig. 2).
Several mutated components among the members of
canonical Wnt signaling, which further result in aberrant
activation of the Wnt/β-catenin pathway, play a paramount role in
malignant transformation and invasiveness of gastric cancer. For
instance, human Wnts has been proved to participate in stomach
tumor progression in autocrine or paracrine manner. Upregulation of
Wnt-1 ligand was reported to be capable of either maintaining the
stemness of GCSC or promoting the advanced stages of gastric cancer
(55,56). In addition, augmented Wnt-2 might
simultaneously result in cytoplasmic β-catenin accumulation and
nuclear localization in both intestinal- and diffuse-type gastric
carcinoma, which was positively associated with stomach cancer
formation as well as tumor invasion or dissemination (57). Kurayoshi et al (58) argued that Wnt-5a gives rise to
stimulating cell migration and invasion in gastric cancer cells by
means of activating focal adhesion kinase and the small GTP-binding
protein Rac. Previous studies stated that upregulation of Wnt10A
induced by tumor necrosis factor α (TNFα) and Helicobacter
pylori (H. pylori) infection might shed light on gastric
carcinogenesis (59). Saitoh et
al (60) confirmed the truth
of the ability of Wnt10B to trigger the development of gastric
cancer. Moreover, both Wnt10A and Wnt10B in GC was tied up with
activation of the β-catenin-TCF signaling pathway. CDH1 gene was
verified to encode E-cadherin so as to sustain epithelial cell-cell
adhesion and suppress tumor invasion while germline gene
alterations of CDH1 had a causative role in ~30–50% of hereditary
diffuse gastric cancer (HDGC) (61,62).
In particular, non-mutated (second) CDH1 allele resulting chiefly
from promoter hypermethylation was likely to disintegrate cell
adherens junctions and was privy to loss of cell polarity,
subsequent to which, β-catenin was activated and HDGC initiated
(63,64). Intestine-specific transcription
factor CDX2 has been found responding to the generation of gastric
adenocarcinoma from intestinal metaplasia, which revealed nuclear
staining of β-catenin and APC gene mutations simultaneously
(65,66).
Exon 3 of CTNNB1, where almost all the β-catenin
mutations reported in human cancers are localized, is widely
accepted to encode serine-threonine phosphorylation sequence for
GSK3β that regulates destruction of β-catenin through the
ubiquitin-proteasome pathway. Multiple experiments provided
evidence indicating that mutations in exon 3 of CTNNB1 precipitates
not only continuous activation of Wnt pathway but also multistep
stomach carcinogenesis (67,68).
Galectin-3, whose mutation locates at position 191, has been widely
recognized equipped to substitute proline to histidine (gal-3H64),
followed by increased nuclear accumulation of β-catenin as well as
promotion of TCF transcription during gastric cancer evolvement
(69). RNF43, a transmembrane E3
ligase, has been shown to eliminate Wnt receptors on cell surface
in order to restrain Wnt signaling (70,71).
Of note, according to a study from Korea, RNF43 gene might harbor
mutational regional intratumoral heterogeneity (ITH), which could
be accountable for tumorigenesis of GC (72). Lu et al (73) discussed that enhancer of zeste
homolog 2 (EZH2) has ectopic expression in gastric cancer tissues.
In a mechanistic manner, repressing CXXC finger protein 4 (CXXC4),
a protein stabilize the degradation complex of β-catenin, was
necessary for the contribution of EZH2 to GC. Many investigations
in GC cases identified crucial association between Actin-binding
protein anillin (ANLN) expression and clinical features while
transcription factor analysis demonstrated the possession of TCF
binding sequence in ANLN gene promoter. Interestingly, ANLN
manifested higher expression among proliferative type gastric
tumors compared to invasive or metabolic types (74). Overexpression of cadherin-17
(CDH17), frizzled (FzE3), Yin Yang 1 (YY1), porcupine (PPN) has all
been discovered to account for Wnt motivation, together with cell
growth in gastric cancer (75–78).
Besides, loss of Wnt repressor function has been
authenticated answerable to gastric cancer conformation. Dickkopf
(DKK) family has been shown to bring about Wnt signal inhibition by
way of binding to LRP5/6 while epigenetic silence of DKK genes is
frequently detected in GC specimens. As an example, DKK-1 was found
to be capable of antagonizing Wnt/β-catenin pathway as well as
weakening the self-renewing ability of cancer stem-like cells
(CSLCs) isolated from MKN-45 cell line (79). Furthermore, DKK-3 is known as
negative regulator of Wnt, whose reduced expression primarily is
due to promoter methylation involved in poor outcome of gastric
carcinoma (80). Than et al
(81) reported that administration
of adenovirus vector carrying REIC/DKK-3 (Ad-REIC/DKK-3) can
inhibit scirrhous gastric carcinoma (SGC) both in vitro and
in nude mice by boosting apoptosis and recruitment of NK cells. On
the contrary, despite DKK-1 or DKK-3 being Wnt suppressors, these
protein have been detected upregulated in GC as well as
significantly correlated to pT-stage and UICC stage (82,83),
revealing that DKKs could have potential in oncogenesis of gastric
cancer rather than as tumor inactivators, which might be explained
either as a negative feedback loop in response to initial
Wnt/β-catenin signaling activation or mediation by non-canonical
Wnt pathways (84). Shin et
al (85) sought to assess the
role of secreted frizzled-related proteins (SFRPs) as antagonist of
Wnt pathway and confirmed that aberrant methylation of SFRPs is the
major mechanism by which Wnt signaling is activated in human GC
cells. Intestinal-type gastric cancer patients exhibit somatic
mutation in germline APC, a pivotal Wnt inhibitor, whose locus at
the chromosome sequence 5q21-22 is frequently lost in
heterozygosity (86). APC gene
deletion was chiefly collected in advanced GCs, suggesting that it
might take place in the progression but not initiation of tumors
and had profound influence on aggressiveness (87). Axin is the scaffold protein in
β-catenin degradation compound while both germline and somatic
alterations in Axin1 or Axin2 genes have been affirmed in
gastrointestinal cancers. Pan et al (88) set forth five SNPs (334 C>T, 874
C>T, 1,396 G>A, 1,690 C>T and 1,942 T>G) and frameshift
mutations in Axin1 involved in GC, in the meantime, a highly
mutable G mononucleotide repeat sequence on exon 7 of Axin2
continually had a frameshift mutation (1-bp deletion) in gastric
cancer with microsatellite instability (MSI) and nuclear β-catenin
stabilization (89). Thus,
devoting our efforts to the regulation on Wnt/β-catenin pathway in
GC will notably benefit anti-metastatic approaches in the
clinic.
E-cadherin plays a crucial role in negatively
regulating Wnt signaling in addition to the proverbial ingredients
of β-catenin degradation complex. β-catenin makes up a direct
contact between cadherins and α-catenin that interacts with the
actin cytoskeleton to create tight cell-cell junctions.
Accumulating testimony of the significance of cadherin in
catenin-mediated cell adherence junctions has confirmed that
cytoplasmic carboxyl-terminus of E-cadherin could bind to β-catenin
so as to suppress its nuclear localization (6). On the contrary, Howard et al
(90) reported that the capability
of β-catenin to combine with cadherins is necessary for its
transcriptional activity since that cadherins might stabilize
β-catenin at membrane by outcompeting its degradation machinery
during EMT. Collectively, it appears that crosstalk between
cadherins and β-catenin may be responsible to the process of
epithelial-mesenchymal transition. Czyzewska et al (91) proved a statistical connection
between not only invasion depth and abnormal β-catenin expression
but also postoperative survival time and expression of E-cadherin.
Additionally, notable significance was observed between E-cadherin
along with β-catenin expression in both main mass of tumor and
lymph nodes involved. Moreover, Yoshii et al (92) detected that combined loss of
membranous E-cadherin and β-catenin expression in original mass
remarkably correlates to lymph node diversion in patients with
intestinal type early gastric cancer. It is noteworthy that, Silva
et al (93) studied a total
of 515 gastric adenocarcinoma patients and separated them into two
groups on the basis of their age. In their series, the young group
(age ≤40 years) presented with higher percentage of
E-cadherin/β-catenin membranous expression compared with the old
group (age >40 years), which could be attributed to the
distinguish between diffuse-type and intestinal-type tumors or the
mutual effect between E-cadherin and other proteins of Wnt
signaling such as APC, suggesting that young GC patients might
advance carcinomas through diversified genetic pathways.
Various metastasis molecules, such as leucine zipper
transcription factor-like 1 (LZTFL1) (94), paired-related homeobox 1 (PRRX1)
(95), Vestigial-like 4 (VGLL4)
(96), high mobility group protein
A2 (HMGA2) (97),
erythropoietin-producing hepatocellular A2 (EphA2) (98), FAT4 (a member of the cadherin
superfamily) (99), have been
showed to regulate EMT via modulating nuclear location of β-catenin
or target genes of Wnt signaling in gastric cancer. Secreted
frizzled-related protein 1 (SFRP1) is classically considered as a
conditioner of Wnt pathway by binding Wnt ligands and its ectopic
transcription has eventful clinical relevance in gastric cancer
development. Qu et al (100) reported that overexpressed sFRP1
is correlated with EMT induction in GC cells as well as lymph node
migration and reduced survival time. Han et al (101) established an EMT model by
treating BGC-823 cells with doxorubicin and elaborated upregulation
of β-catenin, LEF1, c-Myc which indicates activated β-catenin
signaling. Thereafter, noticeable reversion of EMT biomarkers was
found when applying indomethacin or siRNA to stamp down β-catenin.
Given that β-catenin is critical to cadherin-mediated cell adhesion
as the reciprocity between β-catenin and cadherin could promote EMT
in gastric cancer, Zhao et al (102) assessed luciferase reporter
activity of BGC823 cells where theoretical binding site of
β-catenin on the Vimentin promoter was knocked down by mutagenesis,
as a result, Vimentin transcription failed to be regulated. Thus,
we may safely draw a conclusion that intervention of EMT
progression could be achieved only if β-catenin operates steadily.
Another member of collagen genes, the collagen type I (COL1) was
investigated to enhance GC cell motility by means of prompting
disconnection of the E-cadherin/catenin compound. Moreover,
researchers declared that the dissociation of β-catenin from
E-cadherin and actin cytoskeleton is attributable to its tyrosine
phosphorylation, which modifies its chemical attraction to
cadherins and induces β-catenin nuclear trans-situation (103).
Besides, complementary affiliation between EMT and
Wnt pathway in regulation of gastric cancer can be referred to as
microRNA-dependent post-transcriptional modification. Past evidence
is suggestive of the concern of certain miRNAs in different phases
of EMT through Wnt signaling and elaboration of which might furnish
novel diagnostic or therapeutic options for human gastric cancer
hereafter.
MicroRNAs have been reported either as anomalously
expressed or mutated in various tumors. Specifically, above half of
miRNAs locate on tumor associated genomic regions. Analysis of
expression patterns of various miRNAs in different gastric cancer
tissues has showed the diversity of particular miRNA expression in
tumor mass relative to normal mucus. Abundant evidence has verified
that a number of miRNAs act as oncogenes or anti-oncogenes while
taking part in controlling EMT through targeting at constituents of
Wnt pathway (Table I), which
indicats the intimate connection between GC evolution and
miRNAs.
MicroRNA-200 family comprises five members including
miR-200a, miR-200b, miR-200c, miR-141, miR-429 and this family has
been extensively acknowledged to be representative of regulatory
factors of EMT by means of terminating Wnt/β-catenin pathway in
gastric cancer. In an effort to determine the suppressive role of
miR-200a in gastric cancer, Su and colleagues (104) employed TOP/FOP flash luciferase
assay to elucidate the collaboration beiween miR-200a and Wnt
signaling in influencing metastasis potency on GC cells. As a
result, they displayed an inverse relationship between miR-200a
expression and luciferase activity of CTNNB1, indicating that
β-catenin might be a direct target of miR-200a. Another mechanism
by which miR-200a hinders EMT as Cong et al (105) explained was that miR-200a
facilitates E-cadherin through upregulating its transcriptional
antagonists ZEB1/2 and impacting E-cadherin/β-catenin productive
competence. Besides, negative correlation of miR-200a to WHO grades
as well as EMT proteins via Wnt/β-catenin pathway was demonstrated
in gastric adenocarcinoma. Furthermore, a cohort of 90 gastric
cancer cases was investigated by Song et al (106) to ascertain two microRNA subtypes
in accordance with distinct prognosis and the poor-prognosis
subtype was identified to possess deregulation of EMT markers. A
further identification of the contribution of three pivotal miRNAs
(miR-200a, miR-200b, and miR-125b) to poor survival by targeting
EMT gene network was conducted while miR-200b was peculiarly
confirmed to be able to confront ZEB1 and strengthen E-cadherin
in vitro. miR-200b was also validated to act on the 3′UTR of
ZEB2 mRNA to antagonize it and localize E-cadherin to the plasma
membrane (107). Tang et
al (108) explored obvious
loss of miR-200b expression in five gastric cancer cells (HGC-27,
AGS, MGC-803, BGC-823, SGC-7901 and MKN-28) as well as 27 gastric
cancer tissues by qRT-PCR, and the 3′UTR of Wnt-1 was confirmed to
contain the binding regions of miR-200b and they showed restraining
expression of Wnt1, β-catenin and TCF-4 in MGC-803 transfected with
miR-200b mimics.
Many other microRNAs have been implied to be
significant in modulating EMT through Wnt signaling in GC over and
above miR-200 family. Zhang et al (109) recently introduced miR-27 as GC
promoter on account of its incremental level in gastric cancer
specimens. They stated that overexpressing miR-27 brings about
increased ZEB1, ZEB2, Slug, and Vimentin as well as decreased
E-cadherin expression level. APC was eventually identified as the
immediate target of miR-27 that in turn facilitated Wnt/β-catenin
pathway and EMT. Serum response factor (SRF) functions as a
pro-metastatic factor in the context of stomach cancer formation,
current research performed by Zhao et al (110) provided proof that miR-199a-5p is
transactivated when SRF binds to CArG elements in its promoter. The
authors reported that exceptional activation of miR-199a-5p leads
to not only inhibition of CDH1 at posttranscriptional level but
also import of β-catenin from adherens junctions to the cytoplasm
and nucleus, forcefully indicating that the adjustive effect of
miR-199a-5p on EMT may be based on Wnt signaling in GC. miR-544a
potentially conferred a role during EMT abduction in gastric cancer
cell line, Yanaka and colleagues (111) verified it by way of transfecting
MKN1 with a cell-based reporter system which comprises a promoter
sequence of Vim. Deeper investigation made clear that miR-544a
alters expression of Snail and ZEB1 by directly aiming at the 3′UTR
in CDH1 and AXIN2 which successively accelerates nuclear shift of
β-catenin. Xing et al (112) furnished insight into poor
clinicopathological parameters arisen from catenin-δ1 (CTNND1)
perversion by inspecting 126 human GC samples utilizing
immunohistochemistry. Since Wnt signaling assumes one of the
downstream effectors of CTNND1, luciferase assay was implemented to
identify the complementariness of miR-145 to the 3′UTR element of
CTNND1 mRNA, where enables the diversion of CTNND1-E-cadherin
complex from cytoplasm to membrane by downregulating N-cadherin. To
sum up, microRNAs keep an intimate connection with EMT and Wnt
related genes in gastric cancer as the key regulatory factors in
post-transcription network. Clarification of the participation of
miRNAs in reversing EMT process may exert profound influence on
gastric cancer diagnosis, therapy or prognostic evaluation.
Epithelial-mesenchymal transition has established
itself as a foundational process through which epithelial cells
turn into mesenchymal accompanied by apparent genetic and
phenotypic modifications. Assertive evidence is mounting that
Wnt/β-catenin signaling is one of the considerable pathways
orchestrating EMT mainly due to the catenin/cadherin complex. Owing
to the mutuality among EMT and Wnt/β-catenin pathway in GC
pathogenesis, researchers are making efforts to excavate potential
molecules as therapeutic targets for more precise treatment.
The Runt-domain transcription factor 3 (Runx3) has a
defending effect on gastric epithelium as it mediates cellular
proliferation and apoptosis while as many as 80% of GC patients are
observed to carry loss of Runx3. Gastric epithelial cells become
more plastic and susceptible to spontaneous EMT in absence of Runx3
and perverse Wnt signaling fuels the immortal cells to become
carcinogenic, while both could be abolished by endogenous Runx3
inversely, signifying that Runx3 could be deemed as a capable
treatment targets (113). Sox10
is a member of the SRY-related-HMG-box family that has been
documented to be silenced by promoter methylation in GC while its
overexpression remarkably attenuates distinct organ metastasis in
nude mice. Mechanistically, competition with TCF4 for β-catenin
binding consensus element is required for the EMT restraint of
Sox10 (114). Cai et al
(99) examined the repressive role
of FAT4, a cadherin-associated protein, in gastric cancer and the
underlying theory. Knocking out FAT4 boosted pulmonary and hepatic
metastatic lesions in vivo as well as the upregulation of
EMT biomarkers in vitro. Notably, this tumorous promotion
was abrogated on the condition of siRNA-mediated silencing of
β-catenin. Furthermore, many other molecules serve as agitators via
motivating Wnt/β-catenin pathway durung GC progress. EphA2 is a
member of the RTK family whose upregulated level is usually
connected to higher aggressiveness in gastric cancer. Huang et
al (98) found that
overexpression of EphA2 contributes to upregulation of N-cadherin
and Snail while knockdown of EphA2 elicits inverted effect. More
significantly, the result of EphA2 enhancement was neutralized when
applying XAV939 to terminate Wnt signaling, whereas agitation of
Wnt pathway by LiCl remedied the influence of EphA2 silencing on
EMT in the GC cell lines AGS and SGC7901. Zha et al
(97) verified the oncogenic
character of HMGA2 in nude mice subcutaneously transplanted with
MKN-28 and further study conclusively elucidated HMGA2 as a
contributor to EMT by preventing β-catenin from phosphorylation
degradationin in order to activate Wnt signaling. Analogously,
PRRX1 was proven to be positively related to EMT and Wnt/β-catenin
signaling downstream the molecule markers in GC specimens, while
the reinforced EMT and cell mobility was offset by inhibition of
Wnt pathway in BGC823 and SGC7901 lentiviral overexpression of
PRRX1 (95). Since a great number
of similar molecules are playing a part in EMT through modulating
Wnt/β-catenin pathway during GC progression, further insight into
directing these potential targets to reverse EMT can be applicable
to gastric cancer clinically.
In brief, numerous potential molecules act the part
of inducers or suppressors of EMT by coordinating Wnt/β-catenin
pathway in gastric cancer. MicroRNAs at the same time serve as a
link between EMT and Wnt signaling in human GC and miRNA-based
therapeutics such as antisense oligonucleotides, Locked nucleic
acid (LNA) antimiR constructs, miRNAs Sponges and miR-Mask, have
been investigated and developed (115). Nonetheless, elaborate
pharmacodynamic and pharmacokinetic investigation in vivo
models as well as rigorous clinical trials in GC patients are urged
to eliminate off target side effects and ensure regional effective
concentration as far as possible. We firmly believe that
reformative understanding of the impact of related molecules on
catenin/cadherin compound would contribute to advanced
therapeutical applications and better prognosis for human gastric
cancer.
This study was supported by the National Natural
Science Foundation of China (no. 81473605), State Administration of
Traditional Chinese Medicine of the People's Republic of China
(JDZX2012087), the Special Grants for Leading Principal
Investigators from Jiangsu Administration of Traditional Chinese
Medicine (LJ200908), Key Discipline Construction Program from
Jiangsu Administration of Traditional Chinese Medicine (JS1305),
Jiangsu Provincial Commission of Health and Family Planning
(BJ14013), the Priority Academic Program Development of Jiangsu
Higher Education Institutions (PAPD), Jiangsu Provincial Special
Program of Medical Science (BL2014100), and Scientific Research
Innovation for Graduates from Jiangsu Higher Education Institutions
(SJLX15_0440).
|
1
|
Guimarães RM and Muzi CD: Trend of
mortality rates for gastric cancer in Brazil and regions in the
period of 30 years (1980–2009). Arq Gastroenterol. 49:184–188.
2012. View Article : Google Scholar
|
|
2
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
3
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shamir ER, Pappalardo E, Jorgens DM,
Coutinho K, Tsai WT, Aziz K, Auer M, Tran PT, Bader JS and Ewald
AJ: Twist1-induced dissemination preserves epithelial identity and
requires E-cadherin. J Cell Biol. 204:839–856. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pasquier J, Abu-Kaoud N, Al Thani H and
Rafii A: Epithelial to mesenchymal transition in a clinical
perspective. J Oncol. 2015:7921822015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Heuberger J and Birchmeier W: Interplay of
cadherin-mediated cell adhesion and canonical Wnt signaling. Cold
Spring Harb Perspect Biol. 2:a0029152010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhao JH, Luo Y, Jiang YG, He DL and Wu CT:
Knockdown of β-catenin through shRNA cause a reversal of EMT and
metastatic phenotypes induced by HIF-1α. Cancer Invest. 29:377–382.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chang YW, Su YJ, Hsiao M, Wei KC, Lin WH,
Liang CL, Chen SC and Lee JL: Diverse targets of β-catenin during
the epithelial-mesenchymal transition define cancer stem cells and
predict disease relapse. Cancer Res. 75:3398–3410. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Moyret-Lalle C, Ruiz E and Puisieux A:
Epithelial-mesenchymal transition transcription factors and miRNAs:
‘Plastic surgeons’ of breast cancer. World J Clin Oncol. 5:311–322.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Greenburg G and Hay ED: Epithelia
suspended in collagen gels can lose polarity and express
characteristics of migrating mesenchymal cells. J Cell Biol.
95:333–339. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Steinestel K, Eder S, Schrader AJ and
Steinestel J: Clinical significance of epithelial-mesenchymal
transition. Clin Transl Med. 3:172014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Costabile V, Duraturo F, Delrio P, Rega D,
Pace U, Liccardo R, Rossi GB, Genesio R, Nitsch L, Izzo P, et al:
Lithium chloride induces mesenchymal-to-epithelial reverting
transition in primary colon cancer cell cultures. Int J Oncol.
46:1913–1923. 2015.PubMed/NCBI
|
|
13
|
Logullo AF, Nonogaki S, Pasini FS, Osório
CA, Soares FA and Brentani MM: Concomitant expression of
epithelial-mesenchymal transition biomarkers in breast ductal
carcinoma: Association with progression. Oncol Rep. 23:313–320.
2010.PubMed/NCBI
|
|
14
|
Nozato M, Kaneko S, Nakagawara A and
Komuro H: Epithelial-mesenchymal transition-related gene expression
as a new prognostic marker for neuroblastoma. Int J Oncol.
42:134–140. 2013.
|
|
15
|
Nilsson GM, Akhtar N, Kannius-Janson M and
Baeckström D: Loss of E-cadherin expression is not a prerequisite
for c-erbB2-induced epithelial-mesenchymal transition. Int J Oncol.
45:82–94. 2014.PubMed/NCBI
|
|
16
|
Moghbeli M, Moaven O, Memar B, Raziei HR,
Aarabi A, Dadkhah E, Forghanifard MM, Manzari F and Abbaszadegan
MR: Role of hMLH1 and E-cadherin promoter methylation in gastric
cancer progression. J Gastrointest Cancer. 45:40–47. 2014.
View Article : Google Scholar
|
|
17
|
Yu QM, Wang XB, Luo J, Wang S, Fang XH, Yu
JL and Ling ZQ: CDH1 methylation in preoperative peritoneal washes
is an independent prognostic factor for gastric cancer. J Surg
Oncol. 106:765–771. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hansford S, Kaurah P, Li-Chang H, Woo M,
Senz J, Pinheiro H, Schrader KA, Schaeffer DF, Shumansky K,
Zogopoulos G, et al: Hereditary diffuse gastric cancer syndrome:
CDH1 mutations and beyond. JAMA Oncol. 1:23–32. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li XJ, Zhao Y and Ren H: E-cadherin
expression and CDH1 promoter methylation in sporadic and hereditary
gastric cancer]. Nan Fang Yi Ke Da Xue Xue Bao. 35:125–127.
2015.(In Chinese). PubMed/NCBI
|
|
20
|
Jun KH, Lee JE, Kim SH, Jung JH, Choi HJ,
Kim YI, Chin HM and Yang SH: Clinicopathological significance of
N-cadherin and VEGF in advanced gastric cancer brain metastasis and
the effects of metformin in preclinical models. Oncol Rep.
34:2047–2053. 2015.PubMed/NCBI
|
|
21
|
Kamikihara T, Ishigami S, Arigami T,
Matsumoto M, Okumura H, Uchikado Y, Kita Y, Kurahara H, Kijima Y,
Ueno S, et al: Clinical implications of N-cadherin expression in
gastric cancer. Pathol Int. 62:161–166. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Otsuki S, Inokuchi M, Enjoji M, Ishikawa
T, Takagi Y, Kato K, Yamada H, Kojima K and Sugihara K: Vimentin
expression is associated with decreased survival in gastric cancer.
Oncol Rep. 25:1235–1242. 2011.PubMed/NCBI
|
|
23
|
Shirahata A, Sakuraba K, Kitamura Y,
Yokomizo K, Gotou T, Saitou M, Kigawa G, Nemoto H, Sanada Y and
Hibi K: Detection of vimentin methylation in the serum of patients
with gastric cancer. Anticancer Res. 32:791–794. 2012.PubMed/NCBI
|
|
24
|
Zeisberg M and Neilson EG: Biomarkers for
epithelial-mesenchymal transitions. J Clin Invest. 119:1429–1437.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hu Z, Liu X, Tang Z, Zhou Y and Qiao L:
Possible regulatory role of Snail in NF-κB-mediated changes in
E-cadherin in gastric cancer. Oncol Rep. 29:993–1000. 2013.
|
|
26
|
Zhang J, Zhou Y and Yang Y: CCR7 pathway
induces epithelial-mesenchymal transition through up-regulation of
Snail signaling in gastric cancer. Med Oncol. 32:4672015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shin NR, Jeong EH, Choi CI, Moon HJ, Kwon
CH, Chu IS, Kim GH, Jeon TY, Kim DH, Lee JH, et al: Overexpression
of Snail is associated with lymph node metastasis and poor
prognosis in patients with gastric cancer. BMC Cancer. 12:5212012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Barnes RM and Firulli AB: A twist of
insight - the role of Twist-family bHLH factors in development. Int
J Dev Biol. 53:909–924. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu AN, Zhu ZH, Chang SJ and Hang XS:
Twist expression associated with the epithelial-mesenchymal
transition in gastric cancer. Mol Cell Biochem. 367:195–203. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ru GQ, Wang HJ, Xu WJ and Zhao ZS:
Upregulation of Twist in gastric carcinoma associated with tumor
invasion and poor prognosis. Pathol Oncol Res. 17:341–347. 2011.
View Article : Google Scholar
|
|
31
|
Qian J, Luo Y, Gu X, Zhan W and Wang X:
Twist1 promotes gastric cancer cell proliferation through
up-regulation of FoxM1. PLoS One. 8:e776252013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang H, Gong J, Kong D and Liu HY:
Anti-proliferation effects of Twist gene silencing in gastric
cancer SGC7901 cells. World J Gastroenterol. 21:2926–2936. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Choi YJ, Kim N, Chang H, Lee HS, Park SM,
Park JH, Shin CM, Kim JM, Kim JS, Lee DH, et al: Helicobacter
pylori-induced epithelial-mesenchymal transition, a potential role
of gastric cancer initiation and an emergence of stem cells.
Carcinogenesis. 36:553–563. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Okugawa Y, Toiyama Y, Tanaka K, Matsusita
K, Fujikawa H, Saigusa S, Ohi M, Inoue Y, Mohri Y, Uchida K, et al:
Clinical significance of Zinc finger E-box Binding homeobox 1
(ZEB1) in human gastric cancer. J Surg Oncol. 106:280–285. 2012.
View Article : Google Scholar
|
|
35
|
Yabusaki N, Yamada S, Murai T, Kanda M,
Kobayashi D, Tanaka C, Fujii T, Nakayama G, Sugimoto H, Koike M, et
al: Clinical significance of zinc-finger E-box binding homeobox 1
mRNA levels in peritoneal washing for gastric cancer. Mol Clin
Oncol. 3:435–441. 2015.PubMed/NCBI
|
|
36
|
Murai T, Yamada S, Fuchs BC, Fujii T,
Nakayama G, Sugimoto H, Koike M, Fujiwara M, Tanabe KK and Kodera
Y: Epithelial-to-mesenchymal transition predicts prognosis in
clinical gastric cancer. J Surg Oncol. 109:684–689. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jia B, Liu H, Kong Q and Li B:
Overexpression of ZEB1 associated with metastasis and invasion in
patients with gastric carcinoma. Mol Cell Biochem. 366:223–229.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dai YH, Tang YP, Zhu HY, Lv L, Chu Y, Zhou
YQ and Huo JR: ZEB2 promotes the metastasis of gastric cancer and
modulates epithelial mesenchymal transition of gastric cancer
cells. Dig Dis Sci. 57:1253–1260. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Minn YK, Lee H, Hyung WJ, Kim JE, Choi J,
Yang SH, Song H, Lim BJ and Kim SH: MicroRNA-200 family members and
ZEB2 are associated with brain metastasis in gastric
adenocarcinoma. Int J Oncol. 45:2403–2410. 2014.PubMed/NCBI
|
|
40
|
Brabletz T: To differentiate or not -
routes towards metastasis. Nat Rev Cancer. 12:425–436. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang C, Li C, He F, Cai Y and Yang H:
Identification of CD44+CD24+ gastric cancer
stem cells. J Cancer Res Clin Oncol. 137:1679–1686. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Han ME, Jeon TY, Hwang SH, Lee YS, Kim HJ,
Shim HE, Yoon S, Baek SY, Kim BS, Kang CD, et al: Cancer spheres
from gastric cancer patients provide an ideal model system for
cancer stem cell research. Cell Mol Life Sci. 68:3589–3605. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jiang J, Zhang Y, Chuai S, Wang Z, Zheng
D, Xu F, Zhang Y, Li C, Liang Y and Chen Z: Trastuzumab (herceptin)
targets gastric cancer stem cells characterized by CD90 phenotype.
Oncogene. 31:671–682. 2012. View Article : Google Scholar
|
|
45
|
Chen T, Yang K, Yu J, Meng W, Yuan D, Bi
F, Liu F, Liu J, Dai B, Chen X, et al: Identification and expansion
of cancer stem cells in tumor tissues and peripheral blood derived
from gastric adenocarcinoma patients. Cell Res. 22:248–258. 2012.
View Article : Google Scholar :
|
|
46
|
Yoshida K, Tsujimoto H, Matsumura K,
Kinoshita M, Takahata R, Matsumoto Y, Hiraki S, Ono S, Seki S,
Yamamoto J, et al: CD47 is an adverse prognostic factor and a
therapeutic target in gastric cancer. Cancer Med. 4:1322–1333.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yang L, Ping YF, Yu X, Qian F, Guo ZJ,
Qian C, Cui YH and Bian XW: Gastric cancer stem-like cells possess
higher capability of invasion and metastasis in association with a
mesenchymal transition phenotype. Cancer Lett. 310:46–52.
2011.PubMed/NCBI
|
|
48
|
Bessède E, Staedel C, Acuña Amador LA,
Nguyen PH, Chambonnier L, Hatakeyama M, Belleannée G, Mégraud F and
Varon C: Helicobacter pylori generates cells with cancer stem cell
properties via epithelial-mesenchymal transition-like changes.
Oncogene. 33:4123–4131. 2014. View Article : Google Scholar
|
|
49
|
Ryu HS, Park J, Kim HH, Kim WH and Lee HS:
Combination of epithelial-mesenchymal transition and cancer stem
cell-like phenotypes has independent prognostic value in gastric
cancer. Hum Pathol. 43:520–528. 2012. View Article : Google Scholar
|
|
50
|
Xu GF, Zhang WJ, Sun Q, Xu X, Zou X and
Guan W: Combined epithelial-mesenchymal transition with cancer stem
cell-like marker as predictors of recurrence after radical
resection for gastric cancer. World J Surg Oncol. 12:3682014.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jang YS, Sim JJ, Ji E, Jeong KY and Kim
HM: Investigation of lactate calcium salt-induced β-catenin
destabilization in colorectal cancer cells. Life Sci. 139:160–165.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Liu X, Yun F, Shi L, Li ZH, Luo NR and Jia
YF: Roles of signaling pathways in the epithelial-mesenchymal
transition in cancer. Asian Pac J Cancer Prev. 16:6201–6206. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhang X and Hao J: Development of
anticancer agents targeting the Wnt/β-catenin signaling. Am J
Cancer Res. 5:2344–2360. 2015.
|
|
54
|
Yoshida GJ and Saya H: Inversed
relationship between CD44 variant and c-Myc due to oxidative
stress-induced canonical Wnt activation. Biochem Biophys Res
Commun. 443:622–627. 2014. View Article : Google Scholar
|
|
55
|
Mao J, Fan S, Ma W, Fan P, Wang B, Zhang
J, Wang H, Tang B, Zhang Q, Yu X, et al: Roles of Wnt/β-catenin
signaling in the gastric cancer stem cells proliferation and
salinomycin treatment. Cell Death Dis. 5:e10392014. View Article : Google Scholar
|
|
56
|
Zhang H and Xue Y: Wnt pathway is involved
in advanced gastric carcinoma. Hepatogastroenterology.
55:1126–1130. 2008.PubMed/NCBI
|
|
57
|
Cheng XX, Wang ZC, Chen XY, Sun Y, Kong
QY, Liu J and Li H: Correlation of Wnt-2 expression and
beta-catenin intracellular accumulation in Chinese gastric cancers:
Relevance with tumour dissemination. Cancer Lett. 223:339–347.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kurayoshi M, Oue N, Yamamoto H, Kishida M,
Inoue A, Asahara T, Yasui W and Kikuchi A: Expression of Wnt-5a is
correlated with aggressiveness of gastric cancer by stimulating
cell migration and invasion. Cancer Res. 66:10439–10448. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kirikoshi H, Sekihara H and Katoh M:
Up-regulation of WNT10A by tumor necrosis factor alpha and
Helicobacter pylori in gastric cancer. Int J Oncol. 19:533–536.
2001.PubMed/NCBI
|
|
60
|
Saitoh T, Kirikoshi H, Mine T and Katoh M:
Proto-oncogene WNT10B is up-regulated by tumor necrosis factor
alpha in human gastric cancer cell line MKN45. Int J Oncol.
19:1187–1192. 2001.PubMed/NCBI
|
|
61
|
Figueiredo J, Söderberg O, Simões-Correia
J, Grannas K, Suriano G and Seruca R: The importance of E-cadherin
binding partners to evaluate the pathogenicity of E-cadherin
missense mutations associated to HDGC. Eur J Hum Genet. 21:301–309.
2013. View Article : Google Scholar :
|
|
62
|
Garziera M, Canzonieri V, Cannizzaro R,
Geremia S, Caggiari L, De Zorzi M, Maiero S, Orzes E, Perin T,
Zanussi S, et al: Identification and characterization of CDH1
germline variants in sporadic gastric cancer patients and in
individuals at risk of gastric cancer. PLoS One. 8:e770352013.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Humar B, Blair V, Charlton A, More H,
Martin I and Guilford P: E-cadherin deficiency initiates gastric
signet-ring cell carcinoma in mice and man. Cancer Res.
69:2050–2056. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Humar B and Guilford P: Hereditary diffuse
gastric cancer: A manifestation of lost cell polarity. Cancer Sci.
100:1151–1157. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Mutoh H, Sakurai S, Satoh K, Tamada K,
Kita H, Osawa H, Tomiyama T, Sato Y, Yamamoto H, Isoda N, et al:
Development of gastric carcinoma from intestinal metaplasia in
Cdx2-transgenic mice. Cancer Res. 64:7740–7747. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kim HS, Lee JS, Freund JN, Min KW, Lee JS,
Kim W, Juhng SW and Park CS: CDX-2 homeobox gene expression in
human gastric carcinoma and precursor lesions. J Gastroenterol
Hepatol. 21:438–442. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Clements WM, Wang J, Sarnaik A, Kim OJ,
MacDonald J, Fenoglio-Preiser C, Groden J and Lowy AM: β-catenin
mutation is a frequent cause of Wnt pathway activation in gastric
cancer. Cancer Res. 62:3503–3506. 2002.PubMed/NCBI
|
|
68
|
Yoda Y, Takeshima H, Niwa T, Kim JG, Ando
T, Kushima R, Sugiyama T, Katai H, Noshiro H and Ushijima T:
Integrated analysis of cancer-related pathways affected by genetic
and epigenetic alterations in gastric cancer. Gastric Cancer.
18:65–76. 2015. View Article : Google Scholar
|
|
69
|
Kim SJ, Shin JY, Cheong TC, Choi IJ, Lee
YS, Park SH and Chun KH: Galectin-3 germline variant at position
191 enhances nuclear accumulation and activation of β-catenin in
gastric cancer. Clin Exp Metastasis. 28:743–750. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Koo BK, Spit M, Jordens I, Low TY, Stange
DE, van de Wetering M, van Es JH, Mohammed S, Heck AJ, Maurice MM,
et al: Tumour suppressor RNF43 is a stem-cell E3 ligase that
induces endocytosis of Wnt receptors. Nature. 488:665–669. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Hao HX, Xie Y, Zhang Y, Charlat O, Oster
E, Avello M, Lei H, Mickanin C, Liu D, Ruffner H, et al: ZNRF3
promotes Wnt receptor turnover in an R-spondin-sensitive manner.
Nature. 485:195–200. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Jo YS, Kim MS, Lee JH, Lee SH, An CH and
Yoo NJ: Frequent frameshift mutations in 2 mononucleotide repeats
of RNF43 gene and its regional heterogeneity in gastric and
colorectal cancers. Hum Pathol. 46:1640–1646. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Lu H, Sun J, Wang F, Feng L, Ma Y, Shen Q,
Jiang Z, Sun X, Wang X and Jin H: Enhancer of zeste homolog 2
activates wnt signaling through downregulating CXXC finger protein
4. Cell Death Dis. 4:e7762013. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Pandi NS, Manimuthu M, Harunipriya P,
Murugesan M, Asha GV and Rajendran S: In silico analysis of
expression pattern of a Wnt/β-catenin responsive gene ANLN in
gastric cancer. Gene. 545:23–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Qiu HB, Zhang LY, Ren C, Zeng ZL, Wu WJ,
Luo HY, Zhou ZW and Xu RH: Targeting CDH17 suppresses tumor
progression in gastric cancer by downregulating Wnt/β-catenin
signaling. PLoS One. 8:e569592013. View Article : Google Scholar
|
|
76
|
To KF, Chan MW, Leung WK, Yu J, Tong JH,
Lee TL, Chan FK and Sung JJ: Alterations of frizzled (FzE3) and
secreted frizzled related protein (hsFRP) expression in gastric
cancer. Life Sci. 70:483–489. 2001. View Article : Google Scholar
|
|
77
|
Kang W, Tong JH, Chan AW, Zhao J, Dong Y,
Wang S, Yang W, Sin FM, Ng SS, Yu J, et al: Yin Yang 1 contributes
to gastric carcinogenesis and its nuclear expression correlates
with shorter survival in patients with early stage gastric
adenocarcinoma. J Transl Med. 12:802014. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Mo ML, Li MR, Chen Z, Liu XW, Sheng Q and
Zhou HM: Inhibition of the Wnt palmitoyltransferase porcupine
suppresses cell growth and downregulates the Wnt/β-catenin pathway
in gastric cancer. Oncol Lett. 5:1719–1723. 2013.PubMed/NCBI
|
|
79
|
Cai C and Zhu X: The Wnt/β-catenin pathway
regulates self-renewal of cancer stem-like cells in human gastric
cancer. Mol Med Rep. 5:1191–1196. 2012.PubMed/NCBI
|
|
80
|
Yu J, Tao Q, Cheng YY, Lee KY, Ng SS,
Cheung KF, Tian L, Rha SY, Neumann U, Röcken C, et al: Promoter
methylation of the Wnt/beta-catenin signaling antagonist Dkk-3 is
associated with poor survival in gastric cancer. Cancer. 115:49–60.
2009. View Article : Google Scholar
|
|
81
|
Than SS, Kataoka K, Sakaguchi M, Murata H,
Abarzua F, Taketa C, Du G, Yashiro M, Yanagihara K, Nasu Y, et al:
Intraperitoneal administration of an adenovirus vector carrying
REIC/Dkk-3 suppresses peritoneal dissemination of scirrhous gastric
carcinoma. Oncol Rep. 25:989–995. 2011.PubMed/NCBI
|
|
82
|
Gomceli I, Bostanci EB, Ozer I, Kemik AS,
Turhan N, Tez M, Kilic S, Demiriz B and Akoglu M: A novel screening
biomarker in gastric cancer: Serum Dickkopf-1.
Hepatogastroenterology. 59:1661–1664. 2012.
|
|
83
|
Mühlmann G, Untergasser G, Zitt M, Zitt M,
Maier H, Mikuz G, Kronberger IE, Haffner MC, Gunsilius E and Ofner
D: Immunohistochemically detectable dickkopf-3 expression in tumor
vessels predicts survival in gastric cancer. Virchows Arch.
456:635–646. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Hu Y, Wan R, Yu G, Shen J, Ni J, Yin G,
Xing M, Chen C, Fan Y, Xiao W, et al: Imbalance of Wnt/Dkk negative
feedback promotes persistent activation of pancreatic stellate
cells in chronic pancreatitis. PLoS One. 9:e951452014. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Shin H, Kim JH, Lee YS and Lee YC: Change
in gene expression profiles of secreted frizzled-related proteins
(SFRPs) by sodium butyrate in gastric cancers: Induction of
promoter demethylation and histone modification causing inhibition
of Wnt signaling. Int J Oncol. 40:1533–1542. 2012.PubMed/NCBI
|
|
86
|
Ebert MP, Fei G, Kahmann S, Müller O, Yu
J, Sung JJ and Malfertheiner P: Increased β-catenin mRNA levels and
mutational alterations of the APC and β-catenin gene are present in
intestinal-type gastric cancer. Carcinogenesis. 23:87–91. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Fang Z, Xiong Y, Li J, Liu L, Zhang W,
Zhang C and Wan J: APC gene deletions in gastric adenocarcinomas in
a Chinese population: A correlation with tumour progression. Clin
Transl Oncol. 14:60–65. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Pan KF, Liu WG, Zhang L, You WC and Lu YY:
Mutations in components of the Wnt signaling pathway in gastric
cancer. World J Gastroenterol. 14:1570–1574. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Mazzoni SM and Fearon ER: AXIN1 and AXIN2
variants in gastrointestinal cancers. Cancer Lett. 355:1–8. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Howard S, Deroo T, Fujita Y and Itasaki N:
A positive role of cadherin in Wnt/β-catenin signalling during
epithelial-mesenchymal transition. PLoS One. 6:e238992011.
View Article : Google Scholar
|
|
91
|
Czyzewska J, Guzińska-Ustymowicz K,
Ustymowicz M, Pryczynicz A and Kemona A: The expression of
E-cadherin-catenin complex in patients with advanced gastric
cancer: Role in formation of metastasis. Folia Histochem Cytobiol.
48:37–45. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Yoshii T, Miyagi Y, Nakamura Y, Kobayashi
O, Kameda Y and Ohkawa S: Pilot research for the correlation
between the expression pattern of E-cadherin-β-catenin complex and
lymph node metastasis in early gastric cancer. Tumori. 99:234–238.
2013.PubMed/NCBI
|
|
93
|
Silva EM, Begnami MD, Fregnani JH, Pelosof
AG, Zitron C, Montagnini AL and Soares FA: Cadherin-catenin
adhesion system and mucin expression: A comparison between young
and older patients with gastric carcinoma. Gastric Cancer.
11:149–159. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Wang L, Guo J, Wang Q, Zhou J, Xu C, Teng
R, Chen Y, Wei Q and Liu ZP: LZTFL1 suppresses gastric cancer cell
migration and invasion through regulating nuclear translocation of
β-catenin. J Cancer Res Clin Oncol. 140:1997–2008. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Guo J, Fu Z, Wei J, Lu W, Feng J and Zhang
S: PRRX1 promotes epithelial-mesenchymal transition through the
Wnt/β-catenin pathway in gastric cancer. Med Oncol. 32:3932015.
View Article : Google Scholar
|
|
96
|
Li H, Wang Z, Zhang W, Qian K, Liao G, Xu
W and Zhang S: VGLL4 inhibits EMT in part through suppressing
Wnt/β-catenin signaling pathway in gastric cancer. Med Oncol.
32:832015. View Article : Google Scholar
|
|
97
|
Zha L, Zhang J, Tang W, Zhang N, He M, Guo
Y and Wang Z: HMGA2 elicits EMT by activating the Wnt/β-catenin
pathway in gastric cancer. Dig Dis Sci. 58:724–733. 2013.
View Article : Google Scholar
|
|
98
|
Huang J, Xiao D, Li G, Ma J, Chen P, Yuan
W, Hou F, Ge J, Zhong M, Tang Y, et al: EphA2 promotes
epithelial-mesenchymal transition through the Wnt/β-catenin pathway
in gastric cancer cells. Oncogene. 33:2737–2747. 2014. View Article : Google Scholar
|
|
99
|
Cai J, Feng D, Hu L, Chen H, Yang G, Cai
Q, Gao C and Wei D: FAT4 functions as a tumour suppressor in
gastric cancer by modulating Wnt/β-catenin signalling. Br J Cancer.
113:1720–1729. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Qu Y, Ray PS, Li J, Cai Q, Bagaria SP,
Moran C, Sim MS, Zhang J, Turner RR, Zhu Z, et al: High levels of
secreted frizzled-related protein 1 correlate with poor prognosis
and promote tumourigenesis in gastric cancer. Eur J Cancer.
49:3718–3728. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Han R, Xiong J, Xiao R, Altaf E, Wang J,
Liu Y, Xu H, Ding Q and Zhang Q: Activation of β-catenin signaling
is critical for doxorubicin-induced epithelial-mesenchymal
transition in BGC-823 gastric cancer cell line. Tumour Biol.
34:277–284. 2013. View Article : Google Scholar
|
|
102
|
Zhao L, Li W, Zang W, Liu Z, Xu X, Yu H,
Yang Q and Jia J: JMJD2B promotes epithelial-mesenchymal transition
by cooperating with β-catenin and enhances gastric cancer
metastasis. Clin Cancer Res. 19:6419–6429. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Li A, Zhou T, Guo L and Si J: Collagen
type I regulates β-catenin tyrosine phosphorylation and nuclear
translocation to promote migration and proliferation of gastric
carcinoma cells. Oncol Rep. 23:1247–1255. 2010.PubMed/NCBI
|
|
104
|
Su J, Zhang A, Shi Z, Ma F, Pu P, Wang T,
Zhang J, Kang C and Zhang Q: MicroRNA-200a suppresses the
Wnt/β-catenin signaling pathway by interacting with β-catenin. Int
J Oncol. 40:1162–1170. 2012.PubMed/NCBI
|
|
105
|
Cong N, Du P, Zhang A, Shen F, Su J, Pu P,
Wang T, Zjang J, Kang C and Zhang Q: Downregulated microRNA-200a
promotes EMT and tumor growth through the wnt/β-catenin pathway by
targeting the E-cadherin repressors ZEB1/ZEB2 in gastric
adenocarcinoma. Oncol Rep. 29:1579–1587. 2013.PubMed/NCBI
|
|
106
|
Song F, Yang D, Liu B, Guo Y, Zheng H, Li
L, Wang T, Yu J, Zhao Y, Niu R, et al: Integrated microRNA network
analyses identify a poor-prognosis subtype of gastric cancer
characterized by the miR-200 family. Clin Cancer Res. 20:878–889.
2014. View Article : Google Scholar
|
|
107
|
Kurashige J, Kamohara H, Watanabe M,
Hiyoshi Y, Iwatsuki M, Tanaka Y, Kinoshita K, Saito S, Baba Y and
Baba H: MicroRNA-200b regulates cell proliferation, invasion, and
migration by directly targeting ZEB2 in gastric carcinoma. Ann Surg
Oncol. 19(Suppl 3): S656–S664. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Tang H, Kong Y, Guo J, Tang Y and Xie X,
Yang L, Su Q and Xie X: Diallyl disulfide suppresses proliferation
and induces apoptosis in human gastric cancer through Wnt-1
signaling pathway by up-regulation of miR-200b and miR-22. Cancer
Lett. 340:72–81. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Zhang Z, Liu S, Shi R and Zhao G: miR-27
promotes human gastric cancer cell metastasis by inducing
epithelial-to-mesenchymal transition. Cancer Genet. 204:486–491.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Zhao X, He L, Li T, Lu Y, Miao Y, Liang S,
Guo H, Bai M, Xie H, Luo G, et al: SRF expedites metastasis and
modulates the epithelial to mesenchymal transition by regulating
miR-199a-5p expression in human gastric cancer. Cell Death Differ.
21:1900–1913. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Yanaka Y, Muramatsu T, Uetake H, Kozaki K
and Inazawa J: miR-544a induces epithelial-mesenchymal transition
through the activation of WNT signaling pathway in gastric cancer.
Carcinogenesis. 36:1363–1371. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Xing AY, Wang YW, Su ZX, Shi DB, Wang B
and Gao P: Catenin-δ1, negatively regulated by miR-145, promotes
tumour aggressiveness in gastric cancer. J Pathol. 236:53–64. 2015.
View Article : Google Scholar
|
|
113
|
Voon DC, Wang H, Koo JK, Nguyen TA, Hor
YT, Chu YS, Ito K, Fukamachi H, Chan SL, Thiery JP, et al: Runx3
protects gastric epithelial cells against epithelial-mesenchymal
transition-induced cellular plasticity and tumorigenicity. Stem
Cells. 30:2088–2099. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Tong X, Li L, Li X, Heng L, Zhong L, Su X,
Rong R, Hu S, Liu W, Jia B, et al: SOX10, a novel
HMG-box-containing tumor suppressor, inhibits growth and metastasis
of digestive cancers by suppressing the Wnt/β-catenin pathway.
Oncotarget. 5:10571–10583. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Garzon R, Marcucci G and Croce CM:
Targeting microRNAs in cancer: Rationale, strategies and
challenges. Nat Rev Drug Discov. 9:775–789. 2010. View Article : Google Scholar : PubMed/NCBI
|