Introduction
Although the prevalence of gastric cancer has
decreased in most areas of the world (1), it is still the third leading cause of
cancer death in both genders worldwide (2). Despite recent advances in diagnostic
techniques, such as magnifying endoscopy with narrow band imaging
(NBI) (3), and in treatment
including target therapy (4,5),
there are still many gastric cancer patients with a poor prognosis.
The pathogenic mechanism contributing to the aggressive biological
feature in this cancer needs to be clarified.
Semaphorins are secreted and membrane-bound proteins
and were originally implicated in the control of axonal wiring
(6). They comprise a wide protein
family and are involved in a range of functions, from tissue
morphogenesis to the immune response. Recently they have also been
shown to function in the regulation of various phases of cancer
development and have become potential therapeutic targets in
various types of cancer (7).
The high-affinity functional receptor of semaphorin
3E (SEMA3E) is plexin D1 (PLXND1), and SEMA3E-PLXND1 signaling is
required in developmental angiogenesis (8). Moreover, PLXND1 expression is
enhanced in endothelial cells of tumor vessels and in cancer cells
(9,10). The activation of PLXND1 by SEMA3E
on tumor cells facilitates the invasive migration of cancer cells
through the complex formation and transactivation of the epidermal
growth factor receptor 2 (ErbB2) (11), or through regulation of the
subcellular location and activity of Snail, which is a
transcriptional repressor that acts during
epithelial-to-mesenchymal transition (EMT) (12). Luchino et al reported that
SEMA3E/PLXND1 signaling has a critical role in tumor cell
resistance to apoptosis via NR4A1 binding to PLXND1 receptor
(13).
Actually, the expression levels of SEMA3E appear to
be positively correlated with increased metastases in ovarian,
melanoma, and colon cancers and with poor patient survival in
colorectal and pancreatic cancers (11,12,14).
However, little is known about the expression and function of
SEMA3E and PLXND1 in the development or metastasis of gastric
cancer. In this study, we investigated the involvement of
SEMA3E/PLXND1 signaling in the development of gastric cancer.
Materials and methods
Tissues
A total of 124 gastric tissues, 62 matched normal
and carcinoma pairs, were obtained from patients who underwent
surgery at Miyagi Cancer Center (Natori, Japan), between 2007 and
2013. All samples were immediately frozen after resection in liquid
nitrogen and stored at −80°C or fixed in 10% buffered formalin and
embedded in paraffin wax.
The gastric cancers were histopathologically
classified as the intestinal type and diffuse type according to the
classification of the World Health Organization, and were
additionally classified according to the pathologic
tumor-node-metastasis (TNM) Classification (15). No patients received chemotherapy or
radiotherapy before surgery. For statistical analysis, overall
survival was defined by death from any cause, and Kaplan-Meier
survival curves were used.
Cell lines
The gastric cancer cell lines MKN74 (intestinal
type), GCIY (diffuse type) and HGC-27 (diffuse type) were obtained
from RIKEN BioResource Center (Tsukuba, Japan). MKN74 was
maintained in RPMI-1640 (Wako Pure Chemical Industries, Osaka,
Japan) and GCIY and HGC-27 were maintained in DMEM (Wako Pure
Chemical Industries), containing 10% inactivated FBS (EuroClone,
Milan, Italy) with 100 U/ml penicillin and 100 μg/ml streptomycin
(Nacalai Tesque, Kyoto, Japan) and were cultured in a humidified 5%
CO2 incubator at 37°C.
RNA preparation, reverse transcription,
and quantitative real-time PCR (qRT-PCR)
Total RNA was extracted from frozen samples and cell
lines using RNeasy Mini kit (Qiagen, Tokyo, Japan) according to the
manufacturer’s protocol. First-strand cDNAs from all samples were
synthesized from 1.0 μg of total RNA by PrimeScript® 1st
strand cDNA Synthesis kit (Takara Bio, Shiga, Japan) following the
manufacturer’s protocol. The expression of PLXND1 and SEMA3E was
quantified by LightCycler Brilliant SYBR Green qRT-PCR kit (Roche
Applied Science, IN, USA) according to the manufacturer’s protocol
with the specific primer sets (Table
I). The levels of PLXND1 and SEMA3E expression in each sample
were normalized to the respective GAPDH expression levels. The
specificity of each PCR reaction was confirmed by melting curve
analyses.
 | Table IPrimers used in this study. |
Table I
Primers used in this study.
| Gene | Sequence |
|---|
| PLXND1 | Forward:
cagcgctactacaagcagatca
Reverse: gctcaaacttgtgctgcagttgtgt |
| SEMA3E | Forward:
aagtcagattccatcactgtgacat
Reverse: agcaaagtactgttgttctctatgc |
| GAPDH | Forward:
tgaaggtcggagtcaacgg
Reverse: agagttaaaagcagccctggtg |
Phosphorylation of extracellular
signal-regulated kinase (Erk)
To assess the phosphorylation of Erk in MKN74 by
SEMA3E, MKN74 cells were plated in the culture medium without FBS
overnight. The culture medium was aspirated from the dish and cells
were washed using PBS. Culture medium with or without recombinant
SEMA3E was injected. Five minutes post-injection, the cells were
harvested and western blot analysis was performed using antibodies
of α-tubulin, Erk and phosphorylated Erk.
RNA interference
To knockdown SEMA3E in GCIY and HGC-27, we used
Knockout™ RNAi systems (Clontech Laboratories, Mountain View, CA,
USA) according to the manufacturer’s protocol. We designed seven
shRNA sequences targeting SEMA3E according to a previous study
(11). After annealing of the
complementary shRNA oligonucleotides, we ligated those
oligonucleotides into pSIREN vector (sh1 and 2). Then, we
transfected Platinum-A packaging cell lines (Provided by Professor
Kitamura) with shSEMA3E or pSIREN Vector (control) to produce
recombinant retroviruses. Stably infected GCIY and HGC-27 cells
with the recombinant retrovi-ruses were selected with puromycin.
The cells transduced with the sh1 and -2 vector were selected for
further study since these cells showed effective reduction of
SEMA3E by qRT-PCR. The sequences of shRNA targeted SEMA3E are shown
in Table II. All the experiments
in vitro were performed in triplicate, repeated three times
and representative data are shown.
| Table IISequences of shRNA inserts for the
shSEMA3E expressing vector. |
Table II
Sequences of shRNA inserts for the
shSEMA3E expressing vector.
| shRNA | Sequence |
|---|
| shSEMA3E-1 |
| Sense |
gatccGTGCTGAAAGTAATCACAATaCTC
GAGaATTGTGATTACTTTCAGCACttttttG |
| Antisense |
AATTCaaaaaaGTGCTGAAAGTAATCACA
ATtCTCGAGtATTGTGATTACTTTCAGCACg |
| shSEMA3E-2 |
| Sense |
gatccGCCACGATCTTTACAAGCGAAAaC
TCGAGaTTTCGCTTGTAAAGATCGTGGCttttttG |
| Antisense |
AATTCaaaaaaGCCACGATCTTTACAAGC
GAAAtCTCGAGtTTTCGCTTGTAAAGATCGTGGCg |
SEMA3E expression retroviral vector
Human SEMA3E cDNA (Kazusa DNA Research Institute,
Chiba, Japan) was amplified by PCR and inserted into the
pcDNA4.0/3xHA-HisA vector (pcDNA4.0/3xHA-HisA-SEMA3E). Then, human
SEMA3E cDNA with HA tag was amplified by PCR and inserted into the
pBabe puro vector (pBabe-SEMA3E-HA). Recombinant retroviruses were
produced with Platinum-A packaged cell lines. Briefly, Plat-A cells
were transfected with pBabe-SEMA3E-HA or pBabe-puro Vector (EV).
FuGENE-6 (Roche Applied Science) and Opti-MEM I (Gibco/Life
Technologies Co.) were added following the manufacturer’s protocol.
Forty-eight hours after transfection, the retrovirus-containing
supernatant was collected and passed through a 0.45-μm filter.
MKN74 cells were infected with the recombinant retroviruses and
then selected with puromycin.
Antibodies and recombinant protein
Antibodies used in this study were obtained from the
following sources: α-tubulin (Santa Cruz Biotechnology, CA, USA):
Hemagglutinin (HA) (Roche); Erk, and phosphorylated Erk (Cell
Signaling Technology, MA, USA). Recombinant SEMA3E was purchased
from R&D Systems (R&D Systems, MN, USA).
Western blot analysis
Cells were harvested in RIPA buffer supplemented
with protease and phosphatase inhibitors (Roche). The lysates were
pre-cleared by centrifugation (10,000 × g) for 15 min at 4°C, and
supernatants were subjected to 5–20% sodium dodecyl sulfate
polyacrylamide gel electrophoresis and transferred onto
polyvinyliden difluoride membranes. After blocking with 5% non-fat
milk in Tris-buffered saline (TBS) containing 0.1% Tween-20, the
membranes were incubated overnight at 4°C with the primary
antibody. After washing with TBS buffer, they were incubated with
the appropriate horseradish peroxidase-conjugated secondary
antibodies. Reactive bands were detected using SuperSignal West
Pico Chemiluminescent Substrate (Thermo Fisher Scientific Inc., MA,
USA).
Cell growth assay
The cell growth rate was evaluated using Cell
Counting Kit-8 (Dojindo, Kumamoto, Japan). Cells (3×103)
of GCIY, HGC-27 and MKN74 per well were seeded in 96-well plates in
normal cell growth medium. After incubation for 24, 48 or 72 h,
Cell Counting Kit-8 solution (10 μl) was added and the cells were
further incubated for 2 h at 37°C. Then, measurements of absorbance
at 450 nm by VersaMax (Molecular Devices, CA, USA) were measured
after 24-, 48- and 72-h incubation. The index evaluated at 48 and
72 h was normalized to that at 24 h.
Soft agar colony formation assay
Soft agar assays were conducted in 6-well plates.
The base layer of each well consisted of 1 ml with final
concentrations of 1X medium (DMEM or RPMI plus 10% FBS) and 0.6%
low melting point agarose. Plates were cooled at room temperature
until solid, at which point a 1 ml growth agar layer was poured,
consisting of 5×104 cells of GCIY and 1×104
cells of MKN74 suspended in 1X media and 0.3% low melting point
agarose. Plates were again cooled at room temperature until the
growth layer congealed. A further 1 ml of 1X medium without agarose
was added on top of the growth layer on day 0 and 0.5 ml was
applied once a week. Cells were allowed to grow at 37°C for 4 weeks
and total colonies were counted.
Animal experiments
Six-week-old female
NOD/Shi-scid-IL2Rγnull (NOG) mice were purchased from
Central Institute for Experimental Animals (CIEA, Kawasaki, Japan)
and used in this experiment. They were allowed to acclimate for a
week before the experiments. They were housed in a clean room of
the animal care facility at Miyagi Cancer Research Center and kept
under standard temperature, humidity and timed lighting conditions
and provided mouse chow/water ad libitum. The approval of
the Animal Welfare Committee at the Institute was obtained for all
studies.
To investigate the effect of SEMA3E expression in
tumor formation and growth in vivo, 1×105 cells
of GCIY-control (n=6) or GCIY-sh1 (n=6) were injected
subcutaneously with 100 μl PBS into NOG mice. Then we measured the
volume of the subcutaneous tumors formed once a week for 8 weeks.
The tumor volume was calculated by the formula: [(long diameter] ×
(short diameter)2] × 1/2.
To assess the effect of SEMA3E expression on
peritoneal metastasis, we injected 1.0×106 cells of
GCIY-control (n=6) or GCIY-sh1 (n=6) with 200 μl PBS into the
peritoneum of mice. After 5.5 weeks, the mice were sacrificed, and
the presence of nodules and ascites in the peritoneal cavity was
investigated macroscopically.
Statistical analysis
The correlation of PLXND1 and SEMA3E expression with
the patient’s clinicopathological variables was analyzed by
Fisher’s exact test. Statistical significance of differences
between 2 groups was determined using Student’s t-test or Wilcoxon
rank sum test and p-value (<0.05) was regarded as statistically
significant. Statistical difference between 3 or 4 groups was
evaluated using Tukey’s test and a p-value <0.05 was regarded as
statistically significant. All statistical analyses were performed
using JMP® 10 (SAS Institute Inc., Cary, NC, USA).
Results
SEMA3E and PLXND1 expression in human
gastric cancer samples
The SEMA3E and PLXND1 expressions of cancerous and
adjacent non-cancerous tissues from the gastric cancer patients
were examined by qRT-PCR and normalized by the GAPDH expression
levels, and we evaluated the ratios of cancer/non-cancer expression
levels of SEMA3E and PLXND1. The human gastric cancer samples were
obtained from patients with tumors localized to the stomach (N0) or
with any lymph node involvement or distant metastases at diagnosis
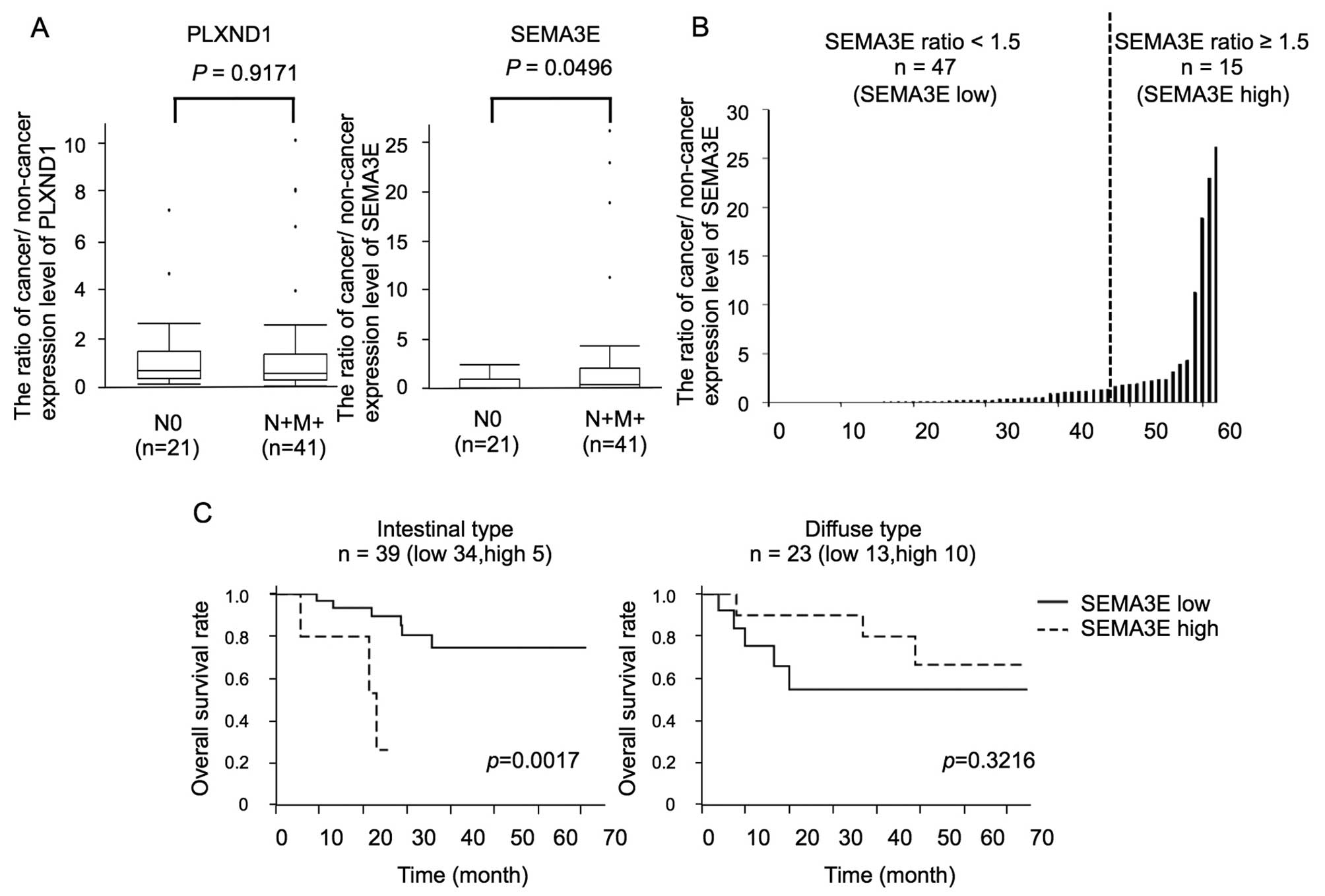
(N+M+). The mean expression ratio of PLXND1 was 1.27±1.73 and
1.50±2.40 (cancer/non-cancer of PLXND1 ± standard deviation) in N0
(n=21) and N+M+ (n=41) patients, respectively. The mean expression
ratio of SEMA3E was 0.52±0.72 and 2.73±6.05 (cancer/non-cancer of
SEMA3E ± standard deviation) in N0 (n=21) and N+M+ (n=41) patients,
respectively. While there were no significant differences in the
PLXND1 expression levels between N0 and N+M+ patients (p=0.9171),
the SEMA3E expression levels in N+M+ patients were significantly
higher than those in N0 patients (p=0.0496) (Fig. 1A).
Moreover, the human gastric cancer tissues were
classified according to the SEMA3E expression ratio into a
high-SEMA3E group (cancer/non-cancer ≥1.5, n=15) and low-SEMA3E
group (cancer/non-cancer <1.5, n=47) (Fig. 1B). The association between the
SEMA3E expression ratio and the clinicopathological features is
summarized in Table III. A
significant association was found between the SEMA3E expression
ratio and the diffuse type gastric cancer. The PLXND1 expression
ratio was higher in the high-SEMA3E group than in the low-SEMA3E
group, but there was no significant difference.
| Table IIIThe association between the SEMA3E
expression ratio and the clinicopathological features. |
Table III
The association between the SEMA3E
expression ratio and the clinicopathological features.
| SEMA3E low
(n=47) | SEMA3E high
(n=15) | P-value |
|---|
| Gender |
| Male | 32 | 10 | |
| Female | 15 | 5 | 1 |
| Histopathology |
| Intestinal | 34 | 5 | |
| Diffuse | 13 | 10 | 0.012 |
| pT |
| T1 | 7 | 1 | |
| T2–4 | 40 | 14 | 0.6665 |
| pN |
| N0 | 18 | 4 | |
| N1–3 | 29 | 11 | 0.5409 |
| pM |
| M0 | 40 | 13 | |
| M1 | 7 | 2 | 1 |
| ly |
| ly0 | 20 | 5 | |
| ly1–3 | 27 | 10 | 0.5638 |
| v |
| v0 | 21 | 10 | |
| v1–3 | 26 | 5 | 0.2351 |
| PLXND1 ratio |
| Mean ± SD | 0.99±1.33 | 2.78±3.51 | 0.1127 |
In addition, in the intestinal type gastric cancer
patients, the high-SEMA3E group (n=5) showed shorter survival than
the low-SEMA3E (n=32) group (p=0.0017, Fig. 1C).
Activation of Erk by SEMA3E in gastric
cancer cells
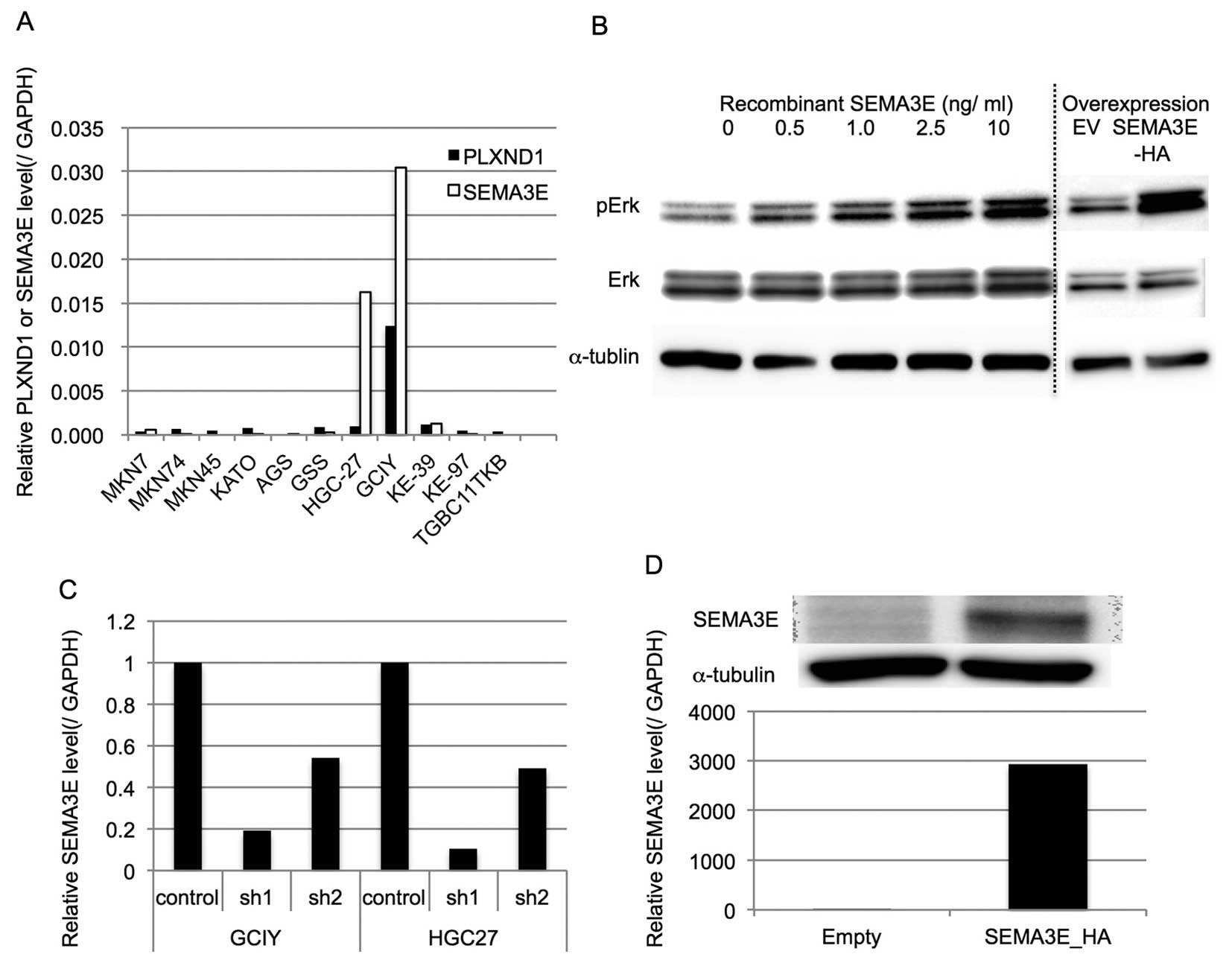
At first, the PLXND1 and SEMA3E expression levels of
several gastric cancer cell lines were examined by qRT-PCR and
normalized by the GAPDH expression level (Fig. 2A). Since PLXND1 expression was not
associated with the biological aggressiveness of gastric cancer, we
examined whether SEMA3E could activate Erk, which is the one of the
representative growth promoting oncogenic signals, in gastric
cancer cells with low level expression of PLXND1. As shown in
Fig. 2B, SEMA3E could
phosphorylate Erk in MKN74 cells, which endogenously express very
low level of PLXND1, depending on the concentration of recombinant
SEMA3E.
Generation of SEMA3E up- and
downregulated cells
To assess the functional role of SEMA3E in gastric
cancer, we generated stably shSEMA3E-expressing GCIY and HGC-27
cells, which express very high levels of endogenous PLXND1 as well
as SEMA3E. The qRT-PCR revealed that the SEMA3E expression levels
were effectively reduced in the knockdown cells (Fig. 2C).
MKN74, a gastric cancer cell line derived from
intestinal adenocarcinoma, expressed low levels of PLXND1 and
SEMA3E. Therefore, we generated stably SEMA3E-overexpressing MKN74
cells (MKN74-SEMA3E-HA). The qRT-PCR and western blot analysis
revealed that the SEMA3E expression levels were effectively
increased in MKN-SEMA3E-HA (Fig.
2D). Consistent with activation of Erk by SEMA3E in MKN74,
MKN74-SEMA3E-HA cells phosphorylated Erk more than MKN74-EV
(Fig. 2B).
Cell growth
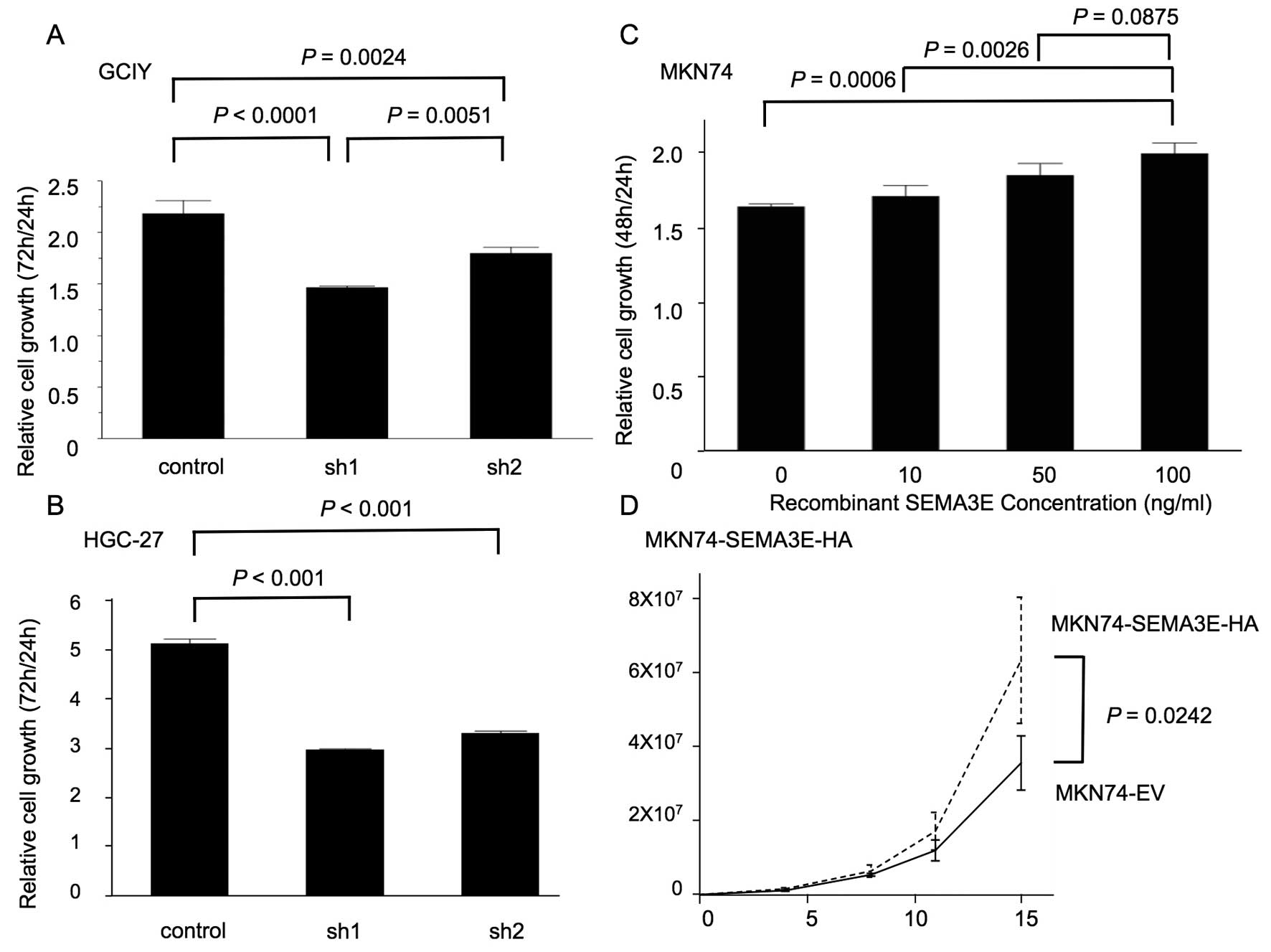
To assess the functional role of SEMA3E in gastric
cancer development, we utilized Cell Counting Kit-8 assays. The
results of these assays showed that cell growth was significantly
reduced in SEMA3E knockdown GCIY and HGC-27 cells (sh1 and sh2)
(Fig. 3A and B). In addition, the
cellular proliferation of MKN74 cells was significantly increased
in a SEMA3E-dependent manner (Fig.
3C) and MKN74-SEMA3E-HA significantly proliferated more than
MKN74-EV (Fig. 3D).
Soft agar colony formation assay
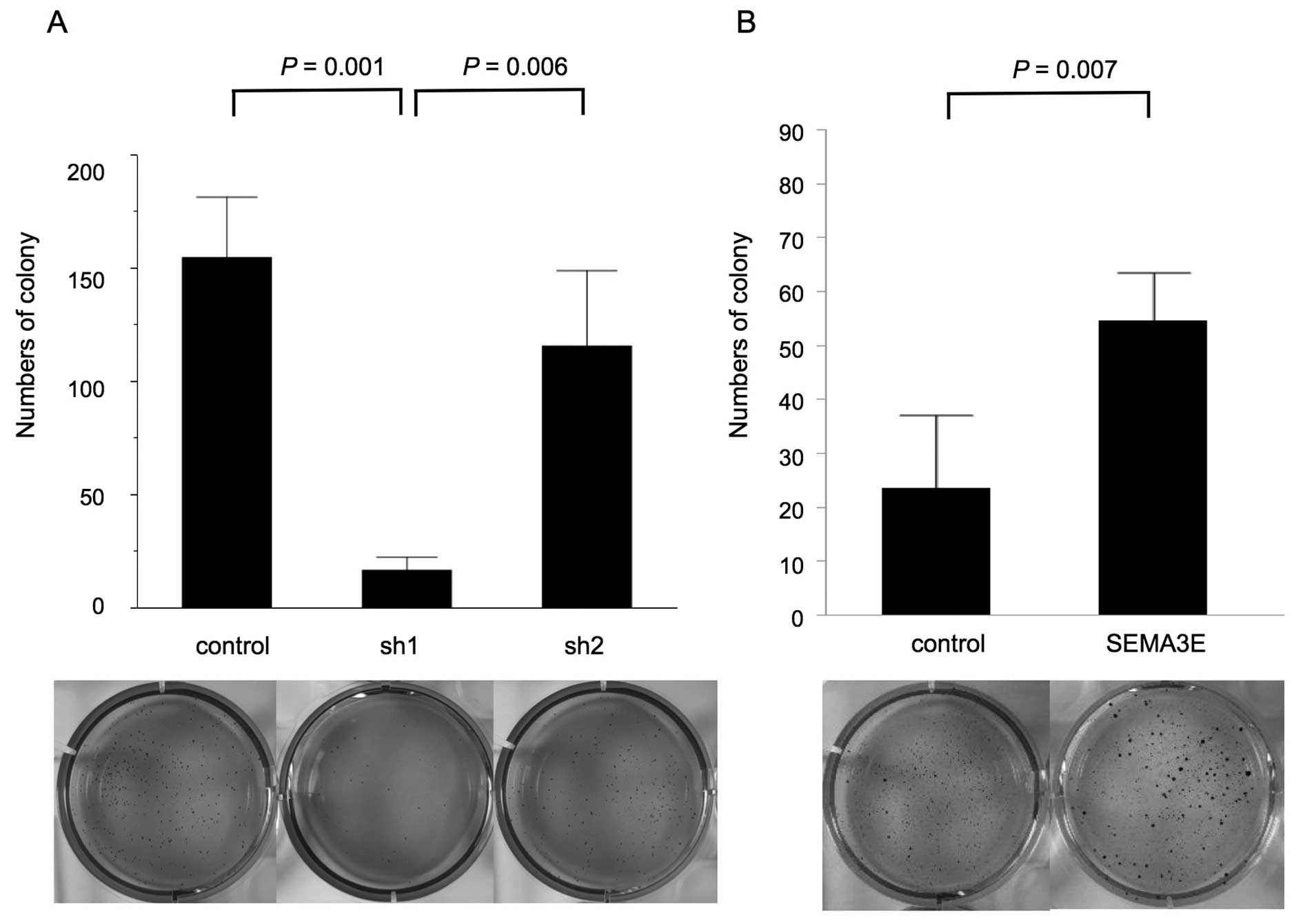
To evaluate the function of SEMA3E in the
anchorage-independent cell growth, we utilized a soft agar assay.
GCIY-sh1 cells formed a smaller numbers of colonies than respective
control cells (p=0.001, Fig. 4A).
Consistent with this finding, MKN74-SEMA3E-HA cells formed larger
numbers of colonies than MKN74-EV in anchorage-independent cell
growth (p=0.007, Fig. 4B).
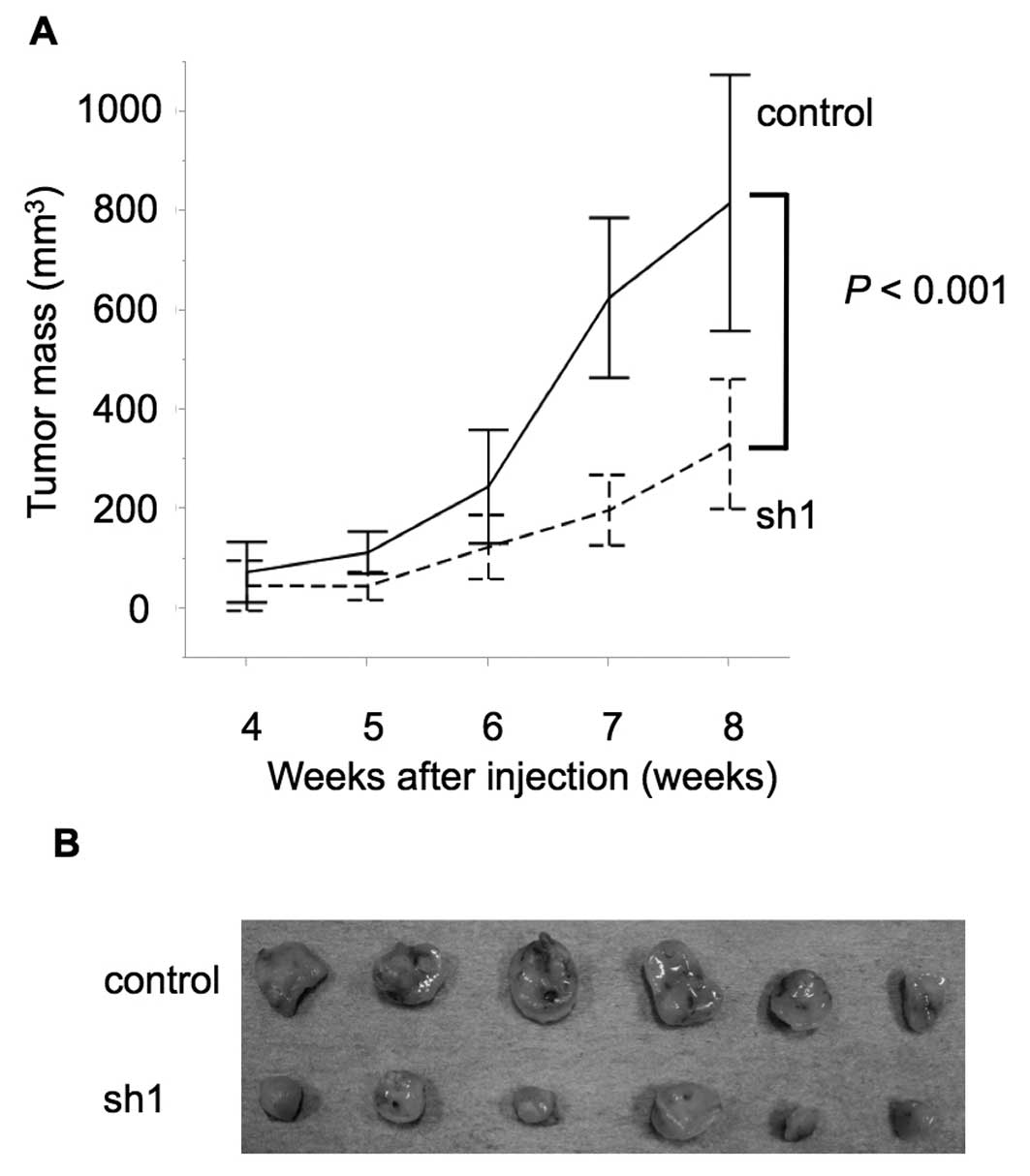
The effects of SEMA3E expression on gastric cancer
growth and peritoneal dissemination in vivo. We examined the
functional role of SEMA3E in gastric cancer development by
subcutaneous injection assay of NOG mice. The tumor growth in NOG
mice was significantly impaired in SEMA3E knockdown GCIY cells
(sh1) (p<0.001) (Fig. 5). Next,
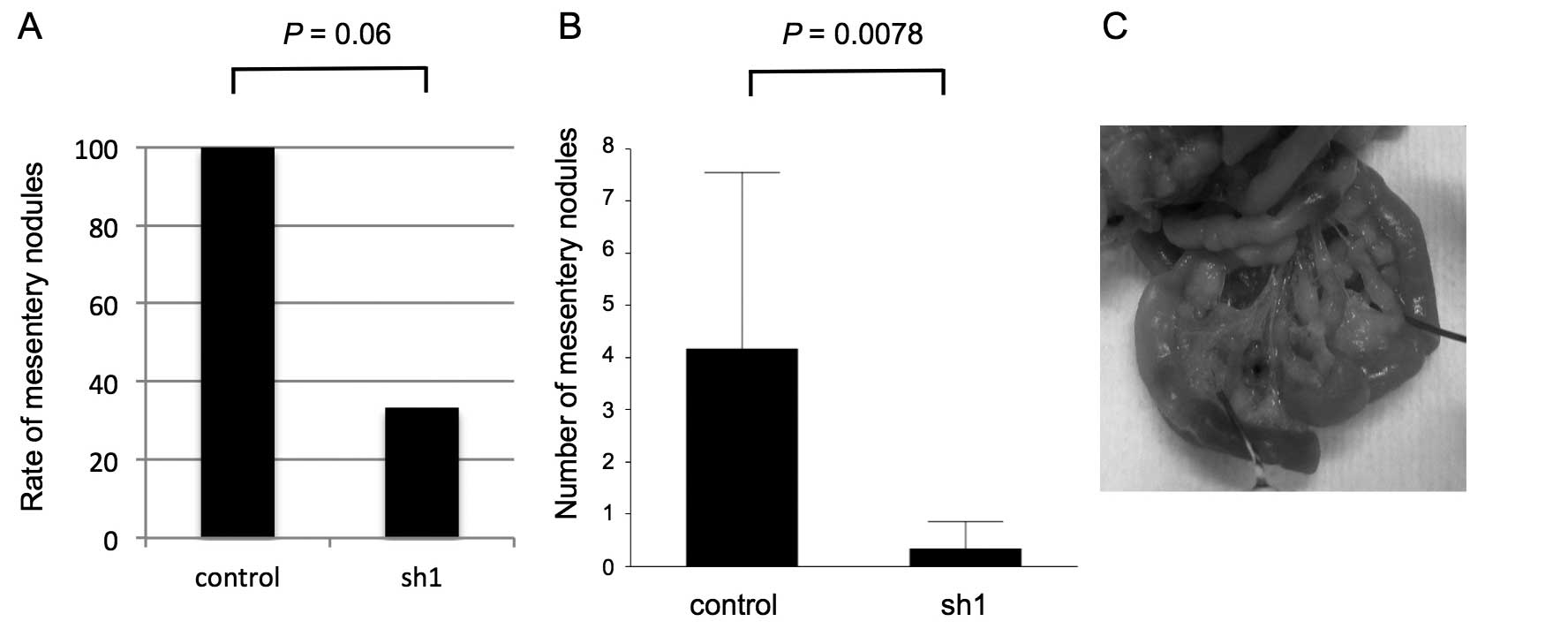
we examined the metastatic capacity of SEMA3E knockdown gastric
cancer cells by intraperitoneal administration assay. GCIY-sh1
cells macroscopically metastasized more slowly (Fig. 5A) and in smaller numbers (Fig. 5B) of mesentery nodules in the
peritoneal cavity compared to control cells (p=0.06, 0.008)
(Fig. 6).
Discussion
We investigated the involvement of SEMA3E-PLXND1
signaling in gastric cancer development and clearly revealed that:
i) the expression level of SEMA3E but not PLXND1 was significantly
associated with lymph node involvement and metastatic progression;
ii) the high level expression of SEMA3E was associated with poor
differentiation and poor survival in the intestinal type of gastric
cancer; iii) SEMA3E could activate Erk signalling even when the
PLXND1 expression level was very low in gastric cancer; iv) SEMA3E
knockdown in gastric cancer cells attenuated cell proliferation and
metastatic ability in vitro and in vivo; v) SEMA3E
enhanced cell proliferation and the anchorage-independent cell
growth in the intestinal type of gastric cancer. These results
suggest that SEMA3E plays a pivotal role in gastric cancer
progression and metastasis.
In several types of human cancers such as colon
cancer, melanoma and breast cancer, elevated expression levels of
SEMA3E correlate with increased malignancy (11,13).
Moreover, SEMA3E is associated with poor patient survival in
pancreatic cancer (14).
Therefore, our present findings are consistent with previous
studies indicating that the expression of SEMA3E is involved in the
enhanced aggressiveness of various types of carcinoma. In this
study, the high-SEMA3E group of intestinal type gastric cancer
patients showed significantly shorter survival than the low-SEMA3E
group. In addition, forced expression of SEMA3E in intestinal
gastric cancer cells enhanced both the anchorage-dependent and
-independent growth of these cells, suggesting that the
over-expression of SEMA3E accelerated the aggressiveness of this
type of gastric cancer cell via the induction of cellular growth.
On the other hand, no relation between the SEMA3E expression level
and the overall survival was found in the diffuse type of gastric
cancer. However, knockdown of this molecule in this type of gastric
cancer cells caused a reduction of the cellular proliferation in
vitro and in vivo, and suppressed peritoneal
disseminations. These findings indicate that SEMA3E is involved in
the development of both types of gastric cancer.
In this study, no association was found between
SEMA3E expression and gastric cancer cell migration. In ovarian,
colon, lung and breast cancer and melanoma, SEMA3E expression
promoted trans-endothelial migration (11,12,16).
In particular the SEMA3E-induced activation of MAPK promoted
epithelial-mesenchymal transition (EMT) and cancer cell migration
in a human ovarian endometrioid carcinoma cell line (12). Although SEMA3E could not affect
gastric cancer cell migration, SEMA3E expression enhanced the
anchorage-dependent as well as -independent cell growth,
subcutaneous tumor growth and peritoneal dissemination in NOG mice,
suggesting that SEMA3E is involved in gastric cancer development
through enhancing cell growth rather than cell migration.
Recently, it was reported that the expression level
of SEMA3E was inversely associated with tumor progression in
gastric cancer and that SEMA3E overexpression inhibited the
migration and invasion of gastric cancer in vitro and in
vivo (17). These results
conflict with our current findings. We cannot explain the reason
for this contradiction, but our results are not an artifact since
enhanced expression of SEMA3E in both clinical samples and cultured
cells was clearly associated with aggressive biological behavior.
In addition to the cancer promoting effect, SEMA3E showed
inhibition of the PDGF-mediated proliferation and migration of
human smooth muscle cells (18),
suggesting that this molecule has multifunctional effects that
differ according to the cellular type. Semaphorins were shown to
have both promoting and suppressive effects in gastric cancer
development. For example, SEMA3A was shown to be significantly
decreased as gastric cancer progressed and metastasized (19). In contrast, SEMA3C and SEMA5A were
demonstrated to be involved in the progression of gastric cancer
(20,21). Therefore, it is not surprising that
SEMA3E has both promoting and suppressive effects on gastric cancer
cells because cancer cells consist of heterogenetic cells.
Accordingly, SEMA3E might give rise to distinct results since the
clinical samples and cultured cells used in our studies and those
by Chen et al (17) were
quite different. Further study will be required to reveal the
function of SEMA3E in gastric cancer.
In conclusion, enhanced SEMA3E expression in the
intestinal type of gastric cancer was associated with a high rate
of the diffuse type of gastric cancer and poor prognosis. SEMA3E
knockdown in gastric cancer cells attenuated the cell proliferation
and metastatic ability in vitro and in vivo, and
SEMA3E promoted gastric cancer cell growth in the intestinal type
of gastric cancer. These results indicate that SEMA3E is likely to
be involved in the development of gastric cancer and might also be
a therapeutic target in the treatment of gastric cancer as well as
breast cancer.
Acknowledgements
We thank Professor Kitamura (The University of
Tokyo, Tokyo, Japan) for providing Platinum-A cells. This study was
supported by Grants-in-Aid for Scientific Research
(KAKENHI)(15K19080) and (24591022) to M.Y. and K.S.,
respectively.
Abbreviations:
|
EMT
|
epithelial-to-mesenchymal
transition
|
|
ErbB2
|
epidermal growth factor receptor 2
|
|
Erk
|
extracellular signal-regulated
kinase
|
|
NBI
|
narrow band imaging
|
|
NOG
|
NOD/Shi-scid-IL2Rγnull
|
|
qRT-PCR
|
quantitative real-time PCR
|
|
PLXND1
|
plexin D1
|
|
SEMA3E
|
semaphorin 3E
|
References
|
1
|
Nagini S: Carcinoma of the stomach: A
review of epidemiology, pathogenesis, molecular genetics and
chemoprevention. World J Gastrointest Oncol. 4:156–169. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar
|
|
3
|
Yagi K, Nozawa Y, Endou S and Nakamura A:
Diagnosis of early gastric cancer by magnifying endoscopy with NBI
from viewpoint of histological imaging: Mucosal patterning in terms
of white zone visibility and its relationship to histology. Diagn
Ther Endosc. 2012:9548092012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boku N: Past and present achievements, and
future direction of the Gastrointestinal Oncology Study Group
(GIOSG), a Division of Japan Clinical Oncology Group (JCOG). Jpn J
Clin Oncol. 41:1315–1321. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Boku N: HER2-positive gastric cancer.
Gastric Cancer. 17:1–12. 2014. View Article : Google Scholar :
|
|
6
|
Zhou Y, Gunput RA and Pasterkamp RJ:
Semaphorin signaling: Progress made and promises ahead. Trends
Biochem Sci. 33:161–170. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rehman M and Tamagnone L: Semaphorins in
cancer: Biological mechanisms and therapeutic approaches. Semin
Cell Dev Biol. 24:179–189. 2013. View Article : Google Scholar
|
|
8
|
Gu C, Yoshida Y, Livet J, Reimert DV, Mann
F, Merte J, Henderson CE, Jessell TM, Kolodkin AL and Ginty DD:
Semaphorin 3E and plexin-D1 control vascular pattern independently
of neuropilins. Science. 307:265–268. 2005. View Article : Google Scholar
|
|
9
|
Moriya J, Minamino T, Tateno K, Okada S,
Uemura A, Shimizu I, Yokoyama M, Nojima A, Okada M, Koga H, et al:
Inhibition of semaphorin as a novel strategy for therapeutic
angiogenesis. Circ Res. 106:391–398. 2010. View Article : Google Scholar
|
|
10
|
Roodink I, Raats J, van der Zwaag B,
Verrijp K, Kusters B, van Bokhoven H, Linkels M, de Waal RM and
Leenders WP: Plexin D1 expression is induced on tumor vasculature
and tumor cells: A novel target for diagnosis and therapy? Cancer
Res. 65:8317–8323. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Casazza A, Finisguerra V, Capparuccia L,
Camperi A, Swiercz JM, Rizzolio S, Rolny C, Christensen C, Bertotti
A, Sarotto I, et al: Sema3E-Plexin D1 signaling drives human cancer
cell invasiveness and metastatic spreading in mice. J Clin Invest.
120:2684–2698. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tseng CH, Murray KD, Jou MF, Hsu SM, Cheng
HJ and Huang PH: Sema3E/plexin-D1 mediated
epithelial-to-mesenchymal transition in ovarian endometrioid
cancer. PLoS One. 6:e193962011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Luchino J, Hocine M, Amoureux MC, Gibert
B, Bernet A, Royet A, Treilleux I, Lécine P, Borg JP, Mehlen P, et
al: Semaphorin 3E suppresses tumor cell death triggered by the
plexin D1 dependence receptor in metastatic breast cancers. Cancer
Cell. 24:673–685. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Biankin AV, Waddell N, Kassahn KS, Gingras
MC, Muthuswamy LB, Johns AL, Miller DK, Wilson PJ, Patch AM, Wu J,
et al; Australian Pancreatic Cancer Genome Initiative. Pancreatic
cancer genomes reveal aberrations in axon guidance pathway genes.
Nature. 491:399–405. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Japanese Gastric Cancer Association.
Japanese classification of gastric carcinoma: 3rd English edition.
Gastric Cancer. 14:101–112. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Christensen C, Ambartsumian N, Gilestro G,
Thomsen B, Comoglio P, Tamagnone L, Guldberg P and Lukanidin E:
Proteolytic processing converts the repelling signal Sema3E into an
inducer of invasive growth and lung metastasis. Cancer Res.
65:6167–6177. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen H, Xie GH, Wang WW, Yuan XL, Xing WM,
Liu HJ, Chen J, Dou M and Shen LS: Epigenetically downregulated
Semaphorin 3E contributes to gastric cancer. Oncotarget.
6:20449–20465. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Movassagh H, Shan L, Halayko AJ, Roth M,
Tamm M, Chakir J and Gounni AS: Neuronal chemorepellent Semaphorin
3E inhibits human airway smooth muscle cell proliferation and
migration. J Allergy Clin Immunol. 133:560–567. 2014. View Article : Google Scholar
|
|
19
|
Tang C, Gao X, Liu H, Jiang T and Zhai X:
Decreased expression of SEMA3A is associated with poor prognosis in
gastric carcinoma. Int J Clin Exp Pathol. 7:4782–4794.
2014.PubMed/NCBI
|
|
20
|
Pan G, Zhu Z, Huang J, Yang C, Yang Y,
Wang Y, Tuo X, Su G, Zhang X, Yang Z, et al: Semaphorin 5A promotes
gastric cancer invasion/metastasis via urokinase-type plasminogen
activator/phosphoinositide 3-kinase/protein kinase B. Dig Dis Sci.
58:2197–2204. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Miyato H, Tsuno NH and Kitayama J:
Semaphorin 3C is involved in the progression of gastric cancer.
Cancer Sci. 103:1961–1966. 2012. View Article : Google Scholar : PubMed/NCBI
|