Introduction
Asbestos exposure causes lung fibrosis known as
asbestosis, one of the most typical forms of pneumoconiosis, as
well as pleural plaque (PP), benign pleural effusion, and diffuse
pleural thickening (1–3). In addition to these benign diseases,
the occurrence of malignant tumors such as lung cancer and
malignant mesothelioma (MM) is an important consideration for the
understanding and development of disease prevention methods, early
diagnosis and treatment (4–6). The
carcinogenic potential of asbestos fibers has been recognized.
Initial DNA damage caused by production of reactive oxygen species
(ROS) mainly derived from iron-including asbestos such as
crocidolite (CRD) and amosite is considered the dominant cause of
asbestos-induced carcinogenesis (7,8).
Next, physiological DNA damage caused by asbestos fibers to
pulmonary epithelial cells and pleural mesothelial cells is the
other cause of malignant transformation because of the rigid and
thin physical features of fibers with more than a 3 aspect ratio
for fibers defined as a 'fiber' (9,10).
In addition to these causes, the easy absorbance of other
carcinogenic substances inhaled into the lung, such as tobacco
smoke and air pollutants on the surface of asbestos fibers, may
enhance the synergistic effects for malignant transformation,
especially for lung epithelial cells, since the odds ratio for lung
cancer caused by asbestos inhalation and tobacco smoking showed
synergistic effects according to various epidemiological studies
(9–11).
We have been investigating the immunological effects
of asbestos fibers because asbestos is a mineral silicate
possessing Si and O as the core chemicals, and silica
(SiO2) has been shown to affect immune competent cells.
The evidence of dysregulation of the immune system caused by silica
was confirmed epidemiologically by frequent complications of
autoimmune diseases, as well as cell biological investigations
which revealed chronic activation of responder and regulatory T
cells (Treg; CD4+, CD25+ and forkhead box P3
(FoxP3) positive inhibitory cells) caused by silica particles
(12–14). Considering the malignant
complications in asbestos-exposed patients, it is possible that
asbestos exposure promotes the reduction of tumor immunity in these
patients (15,16). It is on the basis of this viewpoint
that we examined the effects of asbestos on human immune cells.
Investigations involving human natural killer (NK) cells, a cell
line and freshly isolated NK cells derived from healthy donors (HD)
and stimulated ex vivo, showed reduction of NK cell killing
activity with decreased expression of some NK cell-activating
receptors such as NKG2D and 2B4, with the suppression of the
mitogen-activated protein kinase (MAPK) signaling pathway (17,18).
In addition, the most remarkable reduction among these activating
receptors was NKp46 when comparing its expression to freshly
isolated NK cells derived from HD, PP and MM patients (17,18).
The expression of NKp46 on the surface of NK cells was well
correlated with NK cell killing activities in these patients.
Differentiation and clonal expansion of CD8+ cytotoxic T
lymphocytes (CTLs) were inhibited when chrysotile asbestos was
co-cultured in a mixed lymphocyte reaction (MLR) assay with
reduction of cell attacking molecules such as granzyme B and
perforin (19,20).
Experiments involving T cells have been developed
using cell line models with continuous exposure to chrysotile A
(ChA), chrysotile B (ChB) or CRD for more than one year in
vitro, with cells derived from the human T cell
leukemia/lymphoma (HTLV)-1 virus immortalized human polyclonal cell
line, MT-2. The original MT-2 cells (Org) showed apoptosis with
activation of pro-apoptotic MAPK, mitochondrial apoptotic pathway
and ROS production, when exposed transiently to ChA, ChB or CRD
with relatively high doses of these fibers (21–24).
However, when Org cells (MT-2 cells that never encountered asbestos
fibers) were exposed continuously with relatively low doses of
fibers (doses that do not yield apoptosis in less than half of the
cells) for more than one year, these sublines (designated as CB1 to
3, CA1 to 3, and CR1 exposed to ChB, ChA and CRD, respectively, and
established independently) exhibited changes in cell features
(15,22,24).
The cells showed acquisition of asbestos-induced apoptosis,
alteration of cytokine production, excess production of interleukin
(IL)-10 and transforming growth factor (TGF)-β, reduced production
of interferon (IFN)-γ, resistance to TGF-β-induced growth
inhibition, and enhanced phosphorylation and expression of β-actin
on their cell surface (15,22–25).
In addition to these alterations, the cell surfaces in these
sublines showed reduction of C-X-C motif chemokine receptor 3
(CXCR3), which is known to be important to attract IFN-γ-producing
and tumor attacking T cells near tumor cells (26–28).
Coupled with reduced secretion of IFN-γ, these sublines were
characterized by reduced anti-tumor immunity. These findings were
confirmed in freshly isolated CD4+ T cells derived from
patients with PP or MM (26–28).
Thus, considering these results and those obtained from NK cells
and CTLs indicated that asbestos exposure causes reduction of
antitumor immunity in asbestos-exposed individuals.
Since it was reported that MT-2 cells possess Treg
function (29) and exhibit excess
production of typical soluble factors such as Treg, IL-10 and TGF-β
in CB1-3, CA1-3 and CR sublines continuously exposed to asbestos
fibers such as ChB, ChA and CRD (15,22–25),
it was thought that asbestos exposure of the MT-2 cell line causes
enhanced Treg function. This possibility was investigated together
with the increase in Treg function of sublines through cell cell
contact, as well as the reported excess production of soluble
factors (30).
It was also found that sublines continuously exposed
to asbestos showed remarkably reduced expression of forkhead
transcriptional factor 1 (FoxO1) (31). We reported that this reduced FoxO1
caused reduction of pro-apoptotic molecules such as Puma, Bim and
Fas ligand in exposed sublines (CB1-3, CA1-3 and CR1), in addition
to Bcl-2 overexpression induced by phosphorylation of signal
tranducer and activator of transcription 3 (STAT3) caused by
autocrine usage of overproduced IL-10 (31). FoxO1 is known to regulate various
cell cycle regulators such as cyclins and cyclin-dependent
kinase-inhibitors (CDK-Is) (32–36).
Therefore, in this study we analyzed the expression of these cell
cycle regulators and the status of cell cycle progression in
sublines continuously exposed to asbestos, and compared the results
with those obtained for Org cells.
Materials and methods
Cell lines and asbestos
Details of MT-2, Org and the asbestos-induced
apoptosis resistant sublines (CA1-3, CB1-3 and CR1) have been
reported previously (22–24). These cells were maintained in a
humidified atmosphere of 5% CO2 at 37°C in RPMI-1640
medium supplemented with 10% fetal calf serum (FCS), streptomycin
and penicillin. Seven asbestos-resistant sublines, CA1-3, CB1-3 and
CR1, were generated by continuous exposure to ChA, ChB, and CRD,
respectively. As previously reported (22–24),
the doses of asbestos fibers for continuous exposure were 5-10
µg/ml. These doses induced less than half of the cells to
proceed to apoptosis when transiently exposed (22–24).
The International Union Against Cancer standard ChA and ChB were
kindly provided by the Department of Occupational Health at the
National Institute for Occupational Health, South Africa. In
addition, ChA, ChB and CRD were kindly provided as standard fibers
from the Japan Association for the Study of Fiber Materials. The
mineralogical features of fibers used have been reported previously
(37).
Real-time RT-PCR
The expression levels of various cell cycle
regulators such as CDK-Is including p21Cip1,
p27Kip1, p57Kip2, p16ink4a,
p15ink4b, p18ink4c, and p19ink4d,
and cyclins including cyclin A, B, D1, D2 and e, were analyzed in
Org and continuously exposed sublines (CA1-3, CB1-3 and CR) using
the real-time RT-PCR method. All the primers used are listed in
Table I. Total cellular mRNA from
ORG, CA1-3. CB1-3 and CR1 cells was extracted using the RNeasy Mini
kit (Qiagen GmbH, Hilden, Germany). After synthesis of the first
strand of cDNA, real-time RT-PCR was performed using the SYBeR
Green method (Takara Bio Inc., Kusatsu, Japan) with the Mx3000P
QPCR System (Agilent Technologies, Inc., Santa Clara, CA, USA)
according to the manufacturer's instructions.
 | Table IPrimers used for real-time
RT-PCR. |
Table I
Primers used for real-time
RT-PCR.
| Gene | Sequences
|
|---|
| Forward | Reverse |
|---|
| GAPDH |
5′-GAGTCAACGGATTTGGTCGT -3′ |
5′-TTGATTTTGGAGGGATCTCG-3′ |
| Cyclin A |
5′-ATGTGTGCAGAAGGAGGTCC-3′ |
5′-GAAGGTCCATGAGACAAGGC-3′ |
| Cyclin B |
5′-CGAAGATCAACATGGCAGG-3′ |
5′-CTTGGAGAGGCAGTATCAACC-3′ |
| Cyclin D1 |
5′-ATGTGTGCAGAAGGAGGTCC-3′ |
5′-CCTTCATCTTAGAGGCCACG-3′ |
| Cyclin D2 |
5′-TGCAGAAGGACATCCAACC-3′ |
5′-AGGAACATGCAGACAGCACC-3′ |
| Cyclin E |
5′-TAAATGTCCCGCTCTGAGCC-3′ |
5′-ACGTTTGCCTTCCTCTTCCT-3′ |
| p21Cip1 |
5′-AGCAGAGGAAGACCATGTGG-3′ |
5′-AGGCAGAAGATGTAGAGCGG-3′ |
| p27Kip1 |
5′-AACGTGCGAGTGTCTAACGG-3′ |
5′-CTTCCATGTCTCTGCAGTGC-3′ |
| p57Kip2 |
5′-AGAGATCAGCGCCTGAGAAG-3′ |
5′-TTGCTGCTACATGAACGGTC-3′ |
|
p16ink4a |
5′-ACCAGAGGCAGTAACCATGC-3′ |
5′-CACATGAATGTGCGCTTAGG-3′ |
|
p15ink4b |
5′-CGTTAAGTTTACGGCCAACG-3′ |
5′-CATCATCATGACCTGGATCG-3′ |
|
p18ink4c |
5′-AGTTCCTGGTGAAGCACACG-3′ |
5′-GGCTAACAACCTCATTCCTCC-3′ |
|
p19ink4d |
5′-ATGTCAACGTGCCTGATGG-3′ |
5′-GGAGATCAGATTCAGCTGCC-3′ |
Western blotting
The procedures for western blotting were performed
according to the previously reported methods (31). Briefly, MT-2Org cells and cells
from asbestos continuously exposed sublines were lysed in 20 mM
Tris-HCl (pH 7.5) containing 1 mM EDTA, 1 mM EGTA, 10 mM
2-mercaptoethanol, 1% Triton X-100, 1% sodium deoxycholate, 0.1%
sodium lauryl sulfate, 150 mM NaCl and 1% protease inhibitor
cocktail (Sigma) and briefly sonicated. After centrifugation at
18,000 × g for 10 min, the supernatant was collected and measured
using a BCA protein assay kit (Pierce, Rockford, IL, USA). Cell
lysate containing 50 µg protein was boiled in SDS-sample
buffer and then subjected to SDS-PAGe separation. The resolved
proteins were subsequently electrotransferred onto Immobilon P
membranes (Millipore, Bedford, MA, USA). After initial blocking
with Tris-buffered saline containing 0.2% Tween-20 (TBS-T)
supplemented with 5% BSA for 2 h membranes were then incubated with
each primary antibody in TBS-T containing 1% BSA at dilutions
recommended by the manufacturers for 1–2 h at room temperature
(RT). Thereafter, the membrane was gently rinsed with TBS-T and
then incubated with horseradish peroxidase-conjugated anti-mouse or
anti-rabbit secondary antibodies in TBS-T at dilutions recommended
by the manufacturers for 1 h at RT. After a final set of rinsing
with TBS-T, the presence of the proteins of interest was evaluated
using a chemiluminescence reaction mediated by an ECL Plus
chemiluminescence detection kit (GE Healthcare, Little Chalfont,
UK) and each was then visualized with Chemi-Stage (Toyobo, Osaka,
Japan).
Western blotting for some of the cell cycle
regulators was performed using mouse anti p21Cip1 (F-5)
(Santa Cruz Biotechnology), rabbit anti p16ink4a (C-20)
(Santa Cruz Biotechnology), rabbit anti-cyclin D1 (M-20) (Santa
Cruz Biotechnology), and rabbit anti-β-actin as the control (Cell
Signaling Technology, Danvers, MA, USA).
Flow cytometric analysis of the cell
cycle
The individual cell cycle phases in logarithmically
proliferating ORG, CA1-3, CB1-3 and CR1 cells were analyzed using
flow cytometry. For the continuously exposed sublines CA1-3, CB1-3
and CR1, supplemented asbestos fibers were removed using density
gradient centrifugation and cells without fibers were then cultured
for one week before the analysis. All of these sublines and the Org
MT-2 cells were cultured with bromodeoxyuridine (BrdU) for 30 min,
and after being washed twice with PBS the cells were incubated with
fluorescence-labelled anti-BrdU antibody and 7-amino-actinomycin D
(7AAD) for detection of DNA indices. The G1 phase in the cell cycle
was then determined as BrdU-negative, DNA indices = 2n fraction,
the S phase was BrdU-positive, and DNA indices = 2n< <4n
fraction, while the G2/M phases were BrdU-negative and DNA indices
= 4n fraction.
Knock-down of FoxO1 in the MT-2Org cell
line
Procedures regarding knock-down of FoxO1 in MT-2Org
with lentivirus have been described (31). Lentivirus plasmid vectors
pLKO.1-puro-Control having scramble shRNA sequence and pLKO.1-puro
containing shRNA sequence targeting human FoxO1 (TRCN0000039579 and
TRCN0000039580), and the packaging plasmids kit of pLP1, pLP2 and
pLP/VSVG, were obtained from Sigma and Invitrogen (Carlsbad, CA,
USA), respectively. Recombinant lentivirus was produced in HEK293T
cells and MT-2Org cells were infected with recombinant lentivirus.
Sublines having the shRNA expression cassette with puromycin
resistance were established after culture with medium containing 1
µg/ml puromycin (Sigma) for 2 weeks. Resultant cell lines
were designated Org-Ctrl (Scramble), Org-KD#1 (TRCN0000039579) or
Org-KD#2 (TRCN0000039580). The amount of FoxO1 protein was
determined with immunoblot analysis using anti-FoxO1 monoclonal
antibody (Cell Signaling Technology).
Results
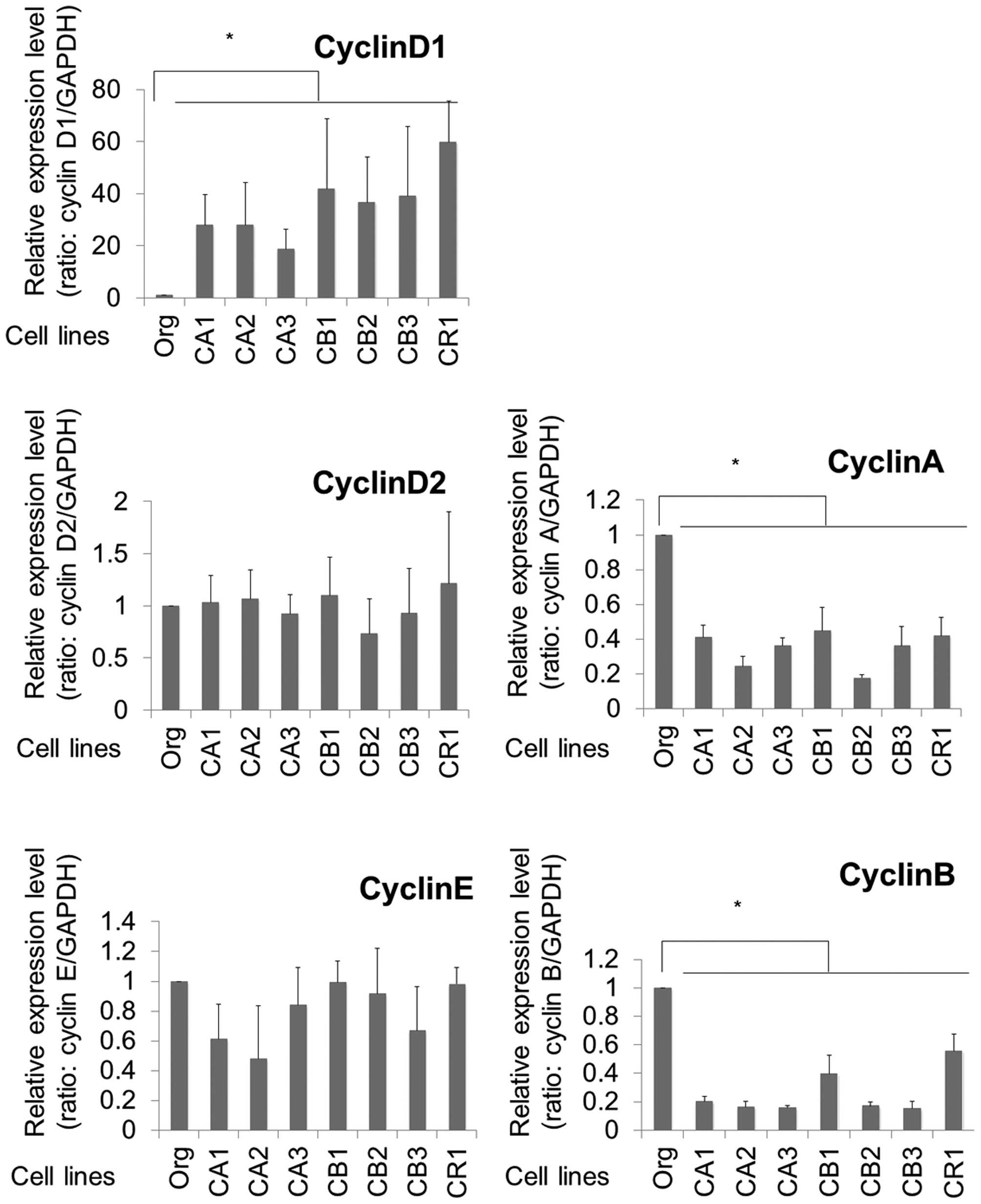
mRNA expression of cell cycle
regulators
As shown in Figs. 1
and 2, various cell cycle
regulators in the cells of Org, CA1-3, CB1-3 and CR1 were analyzed.
FoxO1 transcription factor is known to negatively regulate cyclin
D1 among the various cyclins. In addition, the sublines
continuously exposed to asbestos (MT-2 Org, CA1-3, CB1-3 and CR1)
showed remarkably reduced expression of the FoxO1 gene. As
expected, cyclin D1 expression was remarkably enhanced in all the
sublines by ~20–60-fold when compared with that of Org cells. For
other cyclins, expressions of cyclin D2 and E showed no differences
between Org and sublines. Although cyclins A and B showed decreased
expression in sublines compared to Org cells, the reduction rate
was 0.2–0.5-fold in the sublines. The excess expression of cyclin
D1 in the sublines was a remarkable finding among the cyclins
examined.
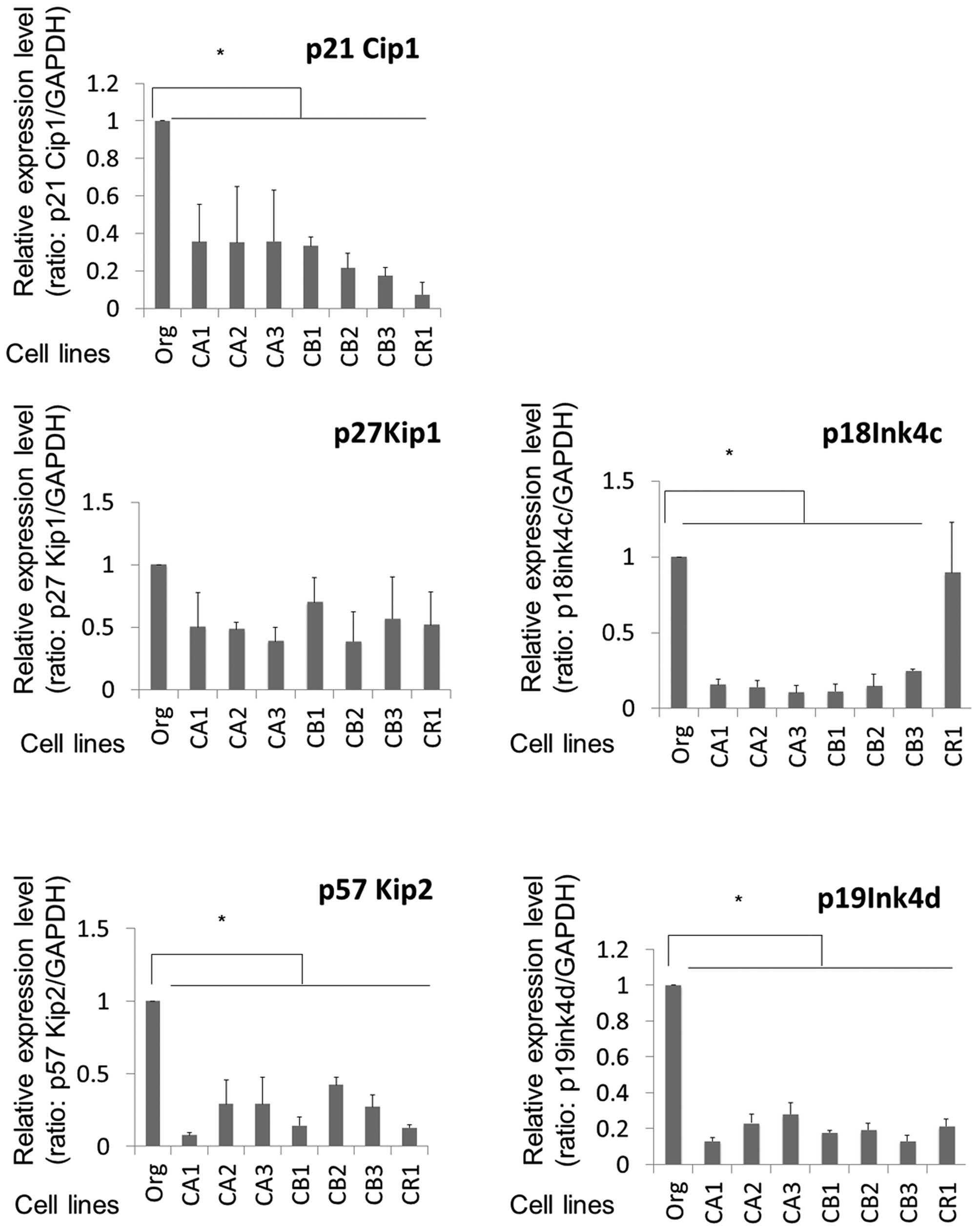
In contrast to results for the cyclins, various
CDK-Is exhibited reduced expression in sublines compared to MT-2
Org cells. As shown in Fig. 2,
p21Cip1, p57Kip2, p18ink4c and
9\p19ink4d showed significantly reduced expression in
the sublines.
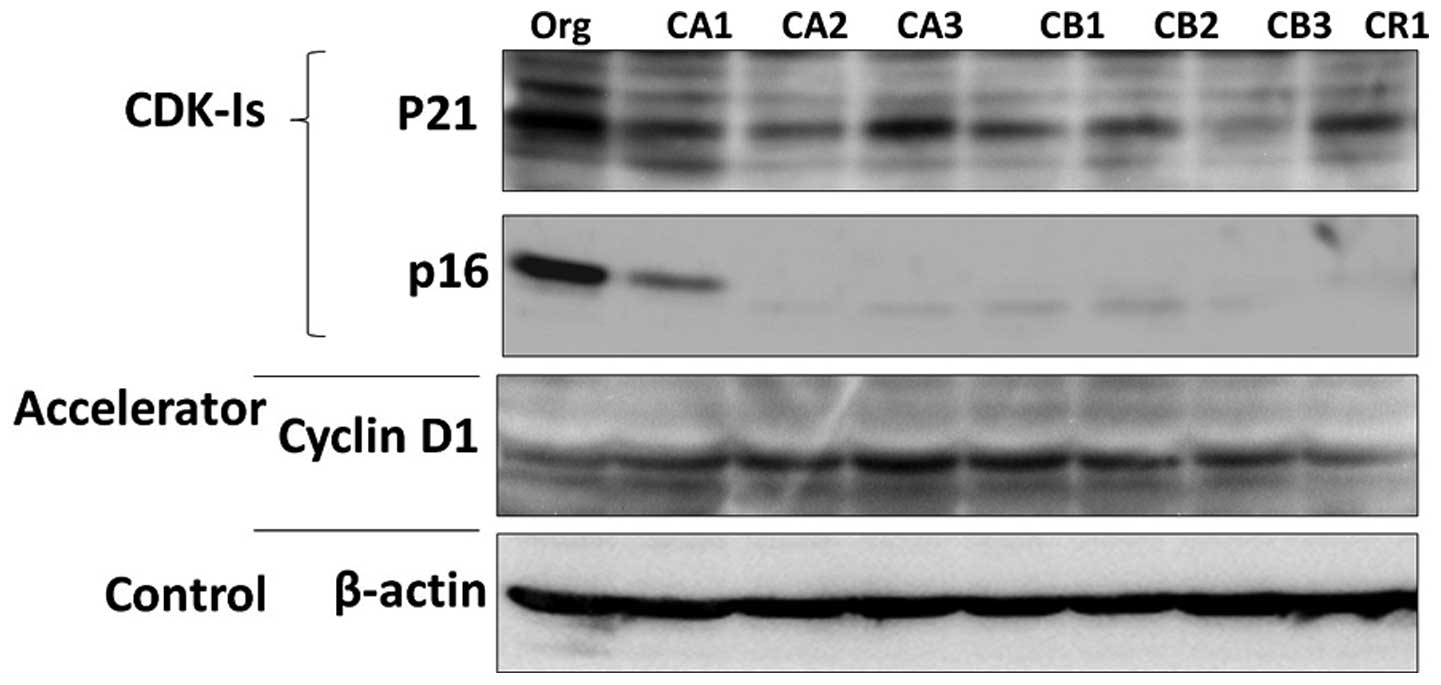
Protein expression of cyclin D1 and
CDK-Is
The representative protein expression of
p21Cip1, p16ink4a and cyclin D1 are shown in
Fig. 3. Similar to findings for
mRNA expression, cyclin D1 was expressed highly in sublines when
compared to that in MT-2 Org cells. In addition, expressions of
p21Cip1 and p16ink4a were reduced in the
sublines. Although p16ink4a mRNA was not representative
because of the lower quality of real-time RT-PCR, it was clear that
the expression of p16ink4a decreased remarkably in
sublines at the protein level.
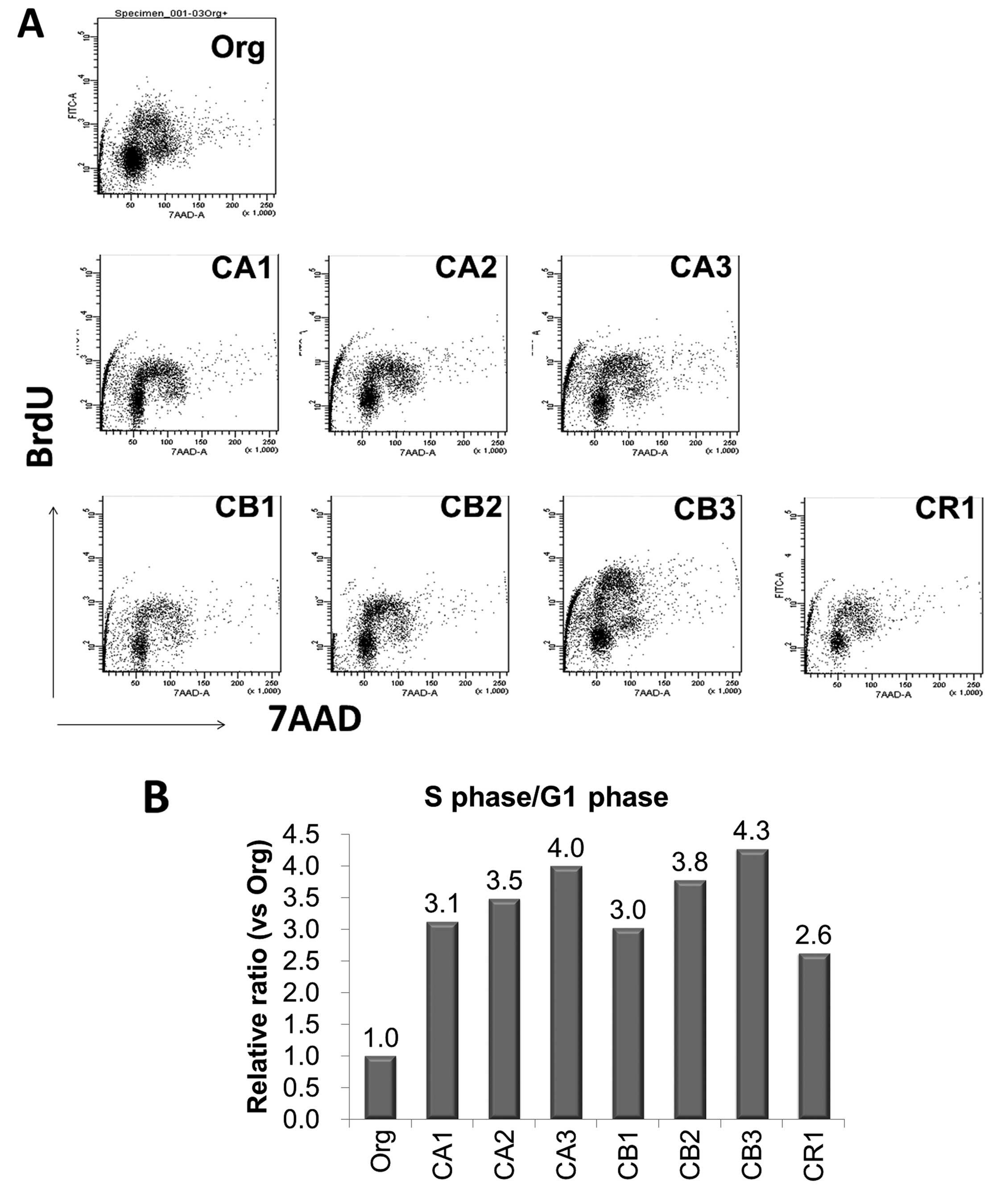
Cell cycle phases in MT-2Org and sublines
continuously exposed to asbestos fibers
As shown in Fig.
4A, cell cycle phases in MT-2 Org cells and the seven sublines
continuously exposed to asbestos fibers (CA1-3, CB1-3 and CR1) were
analyzed using staining with anti-Brdu antibody and DNA indices
using 7AAD. The percentages of the S phase (cells with
BrdU-positive and 2n< <4n in DNA indices) were then divided
by the percentage of the G1 phase (cells with BrdU-negative and 2n
in DNA indices). The results for all cell lines and MT-2 Org are
shown in Fig. 4B, and S/G1
phase-population ratios are relative to a ratio of 1.0 for MT-2
Org. All sublines showed a higher ratio ranging from 2.6 to 4.3.
This indicated that the cell cycle in sublines had progressed
rapidly compared to that of MT-2 Org cells, as suggested from the
data of cell cycle regulator expression, the remarkable excess
expression of cyclin D1, and the reduced expression of CDK-Is.
Cyclin D1 expression in MT-2 Org knocked
down forcibly by FoxO1
The sublines of MT-2 Org, which were continuously
exposed to asbestos fibers, showed a remarkably decreased
expression of FoxO1 as we reported previously. In addition, cyclin
D1, which is negatively regulated by FoxO1, showed recovered
expression in the sublines, and a similar pattern was observed for
CDK-Is, which are positively regulated by FoxO1 and showed reduced
expression in the sublines. To confirm the direct effects of
reduced FoxO1 in MT-2 Org for the expression of cyclin D1,
lentivirus-derived shRNA for FoxO1 was induced in MT-2Org, and the
expression of cyclin D1 in MT-2 Org and the knocked-down subline
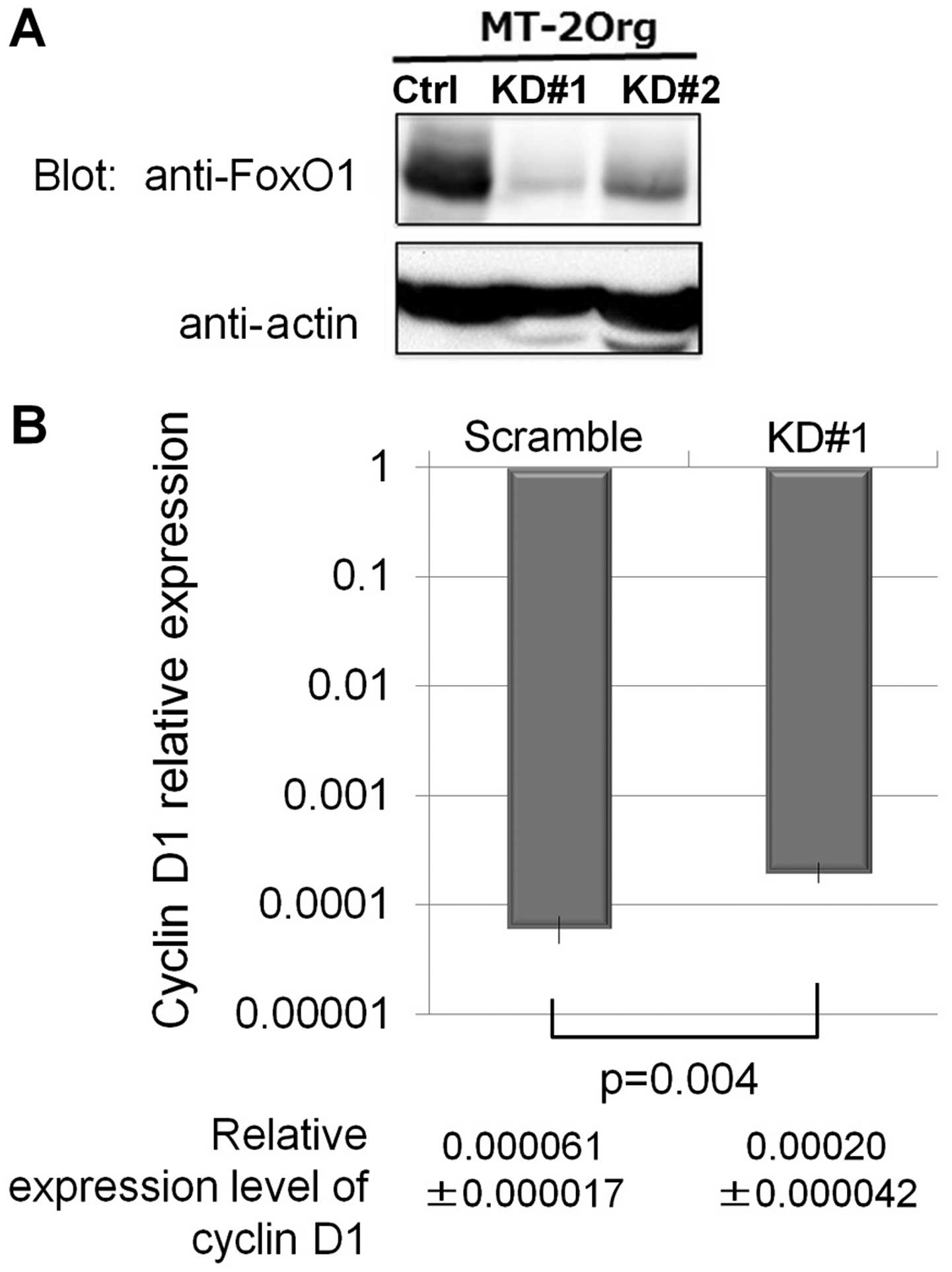
was analyzed. As shown in Fig. 5A,
two of the knocked-down cells (KD#1 and KD#2) showed reduced
expression of FoxO1. Additionally, the mRNA expression of cyclin D1
was assayed using KD#1 cells and scrambled sequence transfected
cells. Although the expression level in KD#1 was not enhanced
remarkably, as we observed in sublines continuously exposed to
asbestos fibers for more than one year, the expression of cyclin D1
was significantly upregulated in KD#1 compared to that in scrambled
transfected cells.
Discussion
Asbestos-exposed patients show complications
comprising malignant tumors such as lung cancer and malignant
mesothelioma (4–6). In addition, these individuals might
have an increased risk of developing other tumors such as
laryngeal, gastrointestinal and bladder cancers (38,39).
Pulmonary regions are also affected by carcinogenic activities such
as ROS production, physical impairment of cellular DNA due to the
mineralogical features of the asbestos fibers, and absorption of
other carcinogenic substances inhaled into the lung, all of which
may lead to the development of malignant tumors. However, a
consideration of other cancers and the long latent period (30–50
years) for pulmonary and pleural occurrence of cancers suggest that
reduction of antitumor immunity may be an important factor in the
development of asbestos-induced tumors (4–6). It
is from this viewpoint that we have been investigating the
immunological effects of asbestos fibers on immune competent cells.
Our results showed that NK cells exhibited reduced killing activity
and expression of various activating receptors, as well as a
decrease of MAPK signaling in experiments involving an
asbestos-exposed human NK cell line, freshly isolated NK cells
exposed to asbestos in vitro, and in NK cells derived from
asbestos-exposed patients with PP or MM (17,18).
CTLs showed reduced differentiation and clonal expansion when
CD8+ T cells were put into the MLR assay with reduction
of cell attacking molecules such as granzyme B and perforin
(19,20). Investigation of T helper cells
showed that CXCR3 expression and IFN-γ production were reduced in
trials involving the cell line model, freshly isolated cells from
HD and continuously exposed to asbestos in vitro, and in
CD4+ cells derived from patients with PP or MM (15,26,27).
The other immune cell that plays an important role
in anti-tumor immunity is Treg. If the function and volume of Treg
are enhanced, the activity of tumor-killing T cells is suppressed,
particularly in the area surrounding tumor cells, and antitumor
immunity is subsequently reduced (40–42).
To investigate the possibility of enhanced Treg function following
its continuous exposure to asbestos, we investigated Treg function
using MT-2 cells, which were reported to possess Treg function
(29,30). Our findings indicated that MT-2
cells exposed continuously to asbestos with a relatively low dose
did not exhibit apoptosis when exposed transiently, showed
enhancement of Treg function via cell-cell contact, and revealed an
increase of soluble factors to function as Treg, namely, IL-10 and
TGF-β, as we reported previously (22,24,30).
The next goal was to determine how asbestos alters
cell proliferation or cell cycle progression in Treg. A key fact
for this investigation was the remarkably reduced expression of
FoxO1 transcription factor found in MT-2 sublines exposed
continuously to asbestos fibers such as ChA, ChB, and CRD. These
sublines showed resistance against the asbestos-induced apoptosis
via upregulation of IL-10, increased phosphorylation of STAT3 by
autocrine usage of IL-10, and subsequent upregulation of Bcl-2
located downstream of STAT3 (22).
Moreover, our recent study revealed that reduced FoxO1 caused
reduction of other apoptosis-related molecules such as Bim, Puma,
and Fas ligand, which are known to be regulated by FoxO1 (31).
FoxO1 is known to regulate various cell cycle
regulators, such as cyclin D1 and CDK-Is. Since cyclin D1 is
negatively regulated and CDK-Is are positively regulated, it seems
that FoxO1 control of the cell cycle does not proceed quickly.
However, if continuous asbestos exposure causes massive
downregulation of FoxO1, the cell proliferating activity might be
accelerated in such cells. Therefore, the expression of cell cycle
regulators in MT-2 Org and its sublines was examined. Results
showed an overall tendency toward acceleration of cell cycle
progression, since cyclin D1 was highly expressed, CDK-Is showed
reduced expression, and the S phase in sublines increased in
comparison to MT-2 Org cells. The increased expression of cyclin D1
was regulated by FoxO1, since artificially silenced cells from MT-2
Org showed increased cyclin D1 expression.
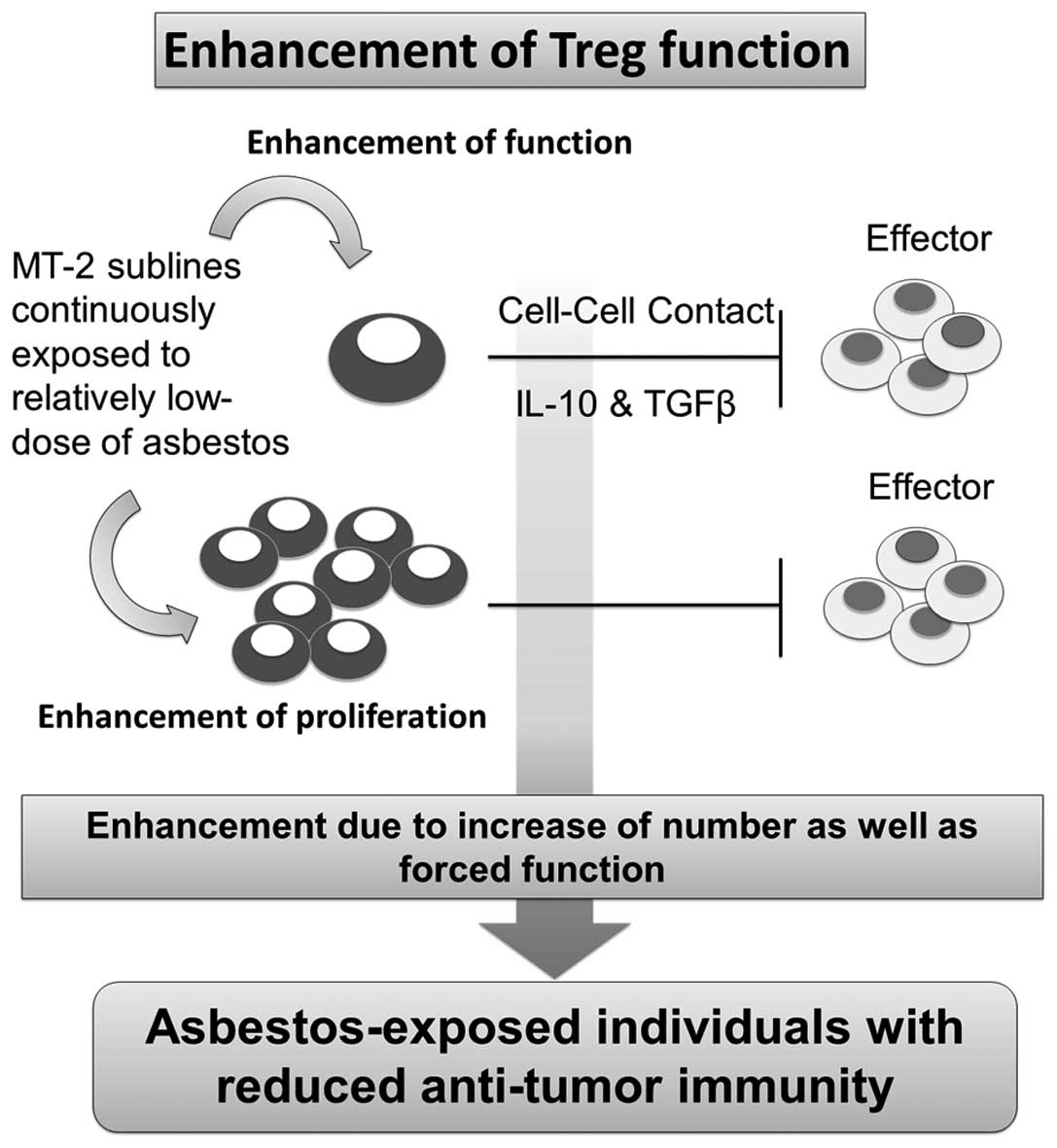
A consideration of the overall results and our
previous findings reveals that MT-2 sublines continuously exposed
to asbestos fibers such as ChA, ChB and CRD showed enhanced Treg
function via cell-cell contact and excess secretion of typical two
soluble factors, namely, IL-10 and TGF-β, and examination of Treg
in the asbestos-exposed population may reflect enhancement of
function and proliferation, quality and volume (Fig. 6). However, it is difficult to
examine these facts in asbestos-exposed patients because the
function and volume of Treg needs to be analyzed in the
tumor-surrounding area instead of peripheral blood. This evaluation
therefore needs time. Notwithstanding these considerations, our
cell line model clearly showed that asbestos causes a reduction of
antitumor immunity in asbestos-exposed individuals and makes them
more sensitive to the development of tumors after a long-term
latent period following the initial exposure to asbestos.
Future studies are needed to evaluate Treg function
and volume in the surrounding areas of asbestos-induced tumors, and
to develop preventive procedures to neutralize asbestos-induced
enhancement of Treg through the use of natural products in foods or
plants. These approaches may be helpful for chemoprevention of
asbestos-induced tumorigenesis.
Acknowledgments
This study was supported by the Private University
Strategic Research Base Formation Support Project (2011 to 2016),
Kakenhi 15K08788, the Japanese Society for the Promotion of
Science, and Research Grants from Kawasaki Medical School (27B058,
26B53, 25B67, 24S4 and 23S6).
References
|
1
|
Parkes WR: Asbestos-related disorders. Br
J Dis Chest. 67:261–300. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mossman BT and Gee JB: Asbestos-related
diseases. N Engl J Med. 320:1721–1730. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Peacock C, Copley SJ and Hansell DM:
Asbestos-related benign pleural disease. Clin Radiol. 55:422–432.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Morinaga K, Kishimoto T, Sakatani M, Akira
M, Yokoyama K and Sera Y: Asbestos-related lung cancer and
mesothelioma in Japan. Ind Health. 39:65–74. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
O'Reilly KM, Mclaughlin AM, Beckett WS and
Sime PJ: Asbestos-related lung disease. Am Fam Physician.
75:683–688. 2007.PubMed/NCBI
|
|
6
|
Lazarus A, Massoumi A, Hostler J and
Hostler DC: Asbestos-related pleuropulmonary diseases: Benign and
malignant. Postgrad Med. 124:116–130. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kamp DW, Graceffa P, Pryor WA and Weitzman
SA: The role of free radicals in asbestos-induced diseases. Free
Radic Biol Med. 12:293–315. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shukla A, Gulumian M, Hei TK, Kamp D,
Rahman Q and Mossman BT: Multiple roles of oxidants in the
pathogenesis of asbestos-induced diseases. Free Radic Biol Med.
34:1117–1129. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Toyokuni S: Mechanisms of asbestos-induced
carcinogenesis. Nagoya J Med Sci. 71:1–10. 2009.PubMed/NCBI
|
|
10
|
Toyokuni S: Role of iron in
carcinogenesis: Cancer as a ferrotoxic disease. Cancer Sci.
100:9–16. 2009. View Article : Google Scholar
|
|
11
|
Lemen RA, Dement JM and Wagoner JK:
Epidemiology of asbestos-related diseases. Environ Health Perspect.
34:1–11. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee S, Matsuzaki H, Kumagai-Takei N,
Yoshitome K, Maeda M, Chen Y, Kusaka M, Urakami K, Hayashi H,
Fujimoto W, et al: Silica exposure and altered regulation of
autoimmunity. Environ Health Prev Med. 19:322–329. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hayashi H, Miura Y, Maeda M, Murakami S,
Kumagai N, Nishimura Y, Kusaka M, Urakami K, Fujimoto W and Otsuki
T: Reductive alteration of the regulatory function of the CD4(+)
CD25(+) T cell fraction in silicosis patients. Int J Immunopathol
Pharmacol. 23:1099–1109. 2010.
|
|
14
|
Lee S, Hayashi H, Maeda M, Chen Y,
Matsuzaki H, Takei-Kumagai N, Nishimura Y, Fujimoto W and Otsuki T:
Environmental factors producing autoimmune dysregulation - chronic
activation of T cells caused by silica exposure. Immunobiology.
217:743–748. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Otsuki T, Matsuzaki H, Lee S,
Kumagai-Takei N, Yamamoto S, Hatayama T, Yoshitome K and Nishimura
Y: Environmental factors and human health: Fibrous and particulate
substance-induced immunological disorders and construction of a
health-promoting living environment. Environ Health Prev Med.
21:71–81. 2016. View Article : Google Scholar
|
|
16
|
Kumagai-Takei N, Maeda M, Chen Y,
Matsuzaki H, Lee S, Nishimura Y, Hiratsuka J and Otsuki T: Asbestos
induces reduction of tumor immunity. Clin Dev Immunol.
2011:4814392011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nishimura Y, Miura Y, Maeda M, Kumagai N,
Murakami S, Hayashi H, Fukuoka K, Nakano T and Otsuki T: Impairment
in cytotoxicity and expression of NK cell- activating receptors on
human NK cells following exposure to asbestos fibers. Int J
Immunopathol Pharmacol. 22:579–590. 2009.PubMed/NCBI
|
|
18
|
Nishimura Y, Maeda M, Kumagai N, Hayashi
H, Miura Y and Otsuki T: Decrease in phosphorylation of ERK
following decreased expression of NK cell-activating receptors in
human NK cell line exposed to asbestos. Int J Immunopathol
Pharmacol. 22:879–888. 2009.
|
|
19
|
Kumagai-Takei N, Nishimura Y, Maeda M,
Hayashi H, Matsuzaki H, Lee S, Hiratsuka J and Otsuki T: Effect of
asbestos exposure on differentiation of cytotoxic T lymphocytes in
mixed lymphocyte reaction of human peripheral blood mononuclear
cells. Am J Respir Cell Mol Biol. 49:28–36. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kumagai-Takei N, Nishimura Y, Maeda M,
Hayashi H, Matsuzaki H, Lee S, Kishimoto T, Fukuoka K, Nakano T and
Otsuki T: Functional properties of CD8 (+) lymphocytes in patients
with pleural plaque and malignant mesothelioma. J Immunol Res.
2014:6701402014. View Article : Google Scholar
|
|
21
|
Hyodoh F, Takata-Tomokuni A, Miura Y,
Sakaguchi H, Hatayama T, Hatada S, Katsuyama H, Matsuo Y and Otsuki
T: Inhibitory effects of anti-oxidants on apoptosis of a human
polyclonal T-cell line, MT-2, induced by an asbestos, chrysotile-A.
Scand J Immunol. 61:442–448. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Miura Y, Nishimura Y, Katsuyama H, Maeda
M, Hayashi H, Dong M, Hyodoh F, Tomita M, Matsuo Y, Uesaka A, et
al: Involvement of IL-10 and Bcl-2 in resistance against an
asbestos-induced apoptosis of T cells. Apoptosis. 11:1825–1835.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Maeda M, Yamamoto S, Chen Y, Kumagai-Takei
N, Hayashi H, Matsuzaki H, Lee S, Hatayama T, Miyahara N, Katoh M,
et al: Resistance to asbestos-induced apoptosis with continuous
exposure to crocidolite on a human T cell. Sci Total Environ.
429:174–182. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maeda M, Chen Y, Hayashi H, Kumagai-Takei
N, Matsuzaki H, Lee S, Nishimura Y and Otsuki T: Chronic exposure
to asbestos enhances TGF-β1 production in the human adult T cell
leukemia virus-immortalized T cell line MT-2. Int J Oncol.
45:2522–2532. 2014.PubMed/NCBI
|
|
25
|
Maeda M, Chen Y, Kumagai-Takei N, Hayashi
H, Matsuzaki H, Lee S, Hiratsuka J, Nishimura Y, Kimura Y and
Otsuki T: Alteration of cytoskeletal molecules in a human T cell
line caused by continuous exposure to chrysotile asbestos.
218:1184–1191. 2013.
|
|
26
|
Maeda M, Nishimura Y, Hayashi H, Kumagai
N, Chen Y, Murakami S, Miura Y, Hiratsuka J, Kishimoto T and Otsuki
T: Reduction of CXC chemokine receptor 3 in an in vitro model of
continuous exposure to asbestos in a human T-cell line, MT-2. Am J
Respir Cell Mol Biol. 45:470–479. 2011. View Article : Google Scholar
|
|
27
|
Maeda M, Nishimura Y, Hayashi H, Kumagai
N, Chen Y, Murakami S, Miura Y, Hiratsuka J, Kishimoto T and Otsuki
T: Decreased CXCR3 expression in CD4+ T cells exposed to
asbestos or derived from asbestos-exposed patients. Am J Respir
Cell Mol Biol. 45:795–803. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Matsuzaki H, Maeda M, Lee S, Nishimura Y,
Kumagai-Takei N, Hayashi H, Yamamoto S, Hatayama T, Kojima Y,
Tabata R, et al: Asbestos-induced cellular and molecular alteration
of immunocompetent cells and their relationship with chronic
inflammation and carcinogenesis. J Biomed Biotechnol.
2012:4926082012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hamano R, Wu X, Wang Y, Oppenheim JJ and
Chen X: Characterization of MT-2 cells as a human regulatory T
cell-like cell line. Cell Mol Immunol. 12:780–782. 2015. View Article : Google Scholar
|
|
30
|
Ying C, Maeda M, Nishimura Y,
Kumagai-Takei N, Hayashi H, Matsuzaki H, Lee S, Yoshitome K,
Yamamoto S, Hatayama T, et al: Enhancement of regulatory T
cell-like suppressive function in MT-2 by long-term and low-dose
exposure to asbestos. Toxicology. 338:86–94. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Matsuzaki H, Lee S, Maeda M, Kumagai-Takei
N, Nishimura Y and Otsuki T: FoxO1 regulates apoptosis induced by
asbestos in the MT-2 human T-cell line. J Immunotoxicol.
13:620–627. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Arden KC: FoxO: Linking new signaling
pathways. Mol Cell. 14:416–418. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Accili D and Arden KC: FoxOs at the
crossroads of cellular metabolism, differentiation, and
transformation. Cell. 117:421–426. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang H and Tindall DJ: CDK2 and FOXO1: A
fork in the road for cell fate decisions. Cell Cycle. 6:902–906.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu P, Kao TP and Huang H: CDK1 promotes
cell proliferation and survival via phosphorylation and inhibition
of FOXO1 transcription factor. Oncogene. 27:4733–4744. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Eijkelenboom A and Burgering BM: FOXOs:
Signalling integrators for homeostasis maintenance. Nat Rev Mol
Cell Biol. 14:83–97. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kohyama N, Shinohara Y and Suzuki Y:
Mineral phases and some reexamined characteristics of the
International Union Against Cancer standard asbestos samples. Am J
Ind Med. 30:515–528. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gao FF and Oury TD: Other neoplasia.
Pathology of Asbestos-Associated Diseases. Roggi VL, Oury TD and
Sporn TA: 3rd ed. Springer; Berlin: pp. 177–192. 2014, View Article : Google Scholar
|
|
39
|
Craighead Je: Nonthoracic cancers possibly
resulting from asbestos exposure. Asbestos and its Diseases.
Craighead JE and Gibbs AR: Oxford University Press; New York, NY:
pp. 230–252. 2008, View Article : Google Scholar
|
|
40
|
Sakaguchi S, Ono M, Setoguchi R, Yagi H,
Hori S, Fehervari Z, Shimizu J, Takahashi T and Nomura T:
Foxp3+ CD25+ CD4+ natural
regulatory T cells in dominant self-tolerance and autoimmune
disease. Immunol Rev. 212:8–27. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nishikawa H and Sakaguchi S: Regulatory T
cells in tumor immunity. Int J Cancer. 127:759–767. 2010.PubMed/NCBI
|
|
42
|
Yamaguchi T and Sakaguchi S: Regulatory T
cells in immune surveillance and treatment of cancer. Semin Cancer
Biol. 16:115–123. 2006. View Article : Google Scholar
|