Introduction
Colorectal cancer is one of the most common
malignant tumors and the fourth leading cause of cancer-related
death around the world, with roughly 1.4 million new cases and
nearly 700,000 deaths per year (1). According to research prediction,
colon cancer cases will increase to 2.2 million, and the number of
deaths may reach 1.1 million by 2030 (1). To date, the pathogenesis of
colorectal cancer is not completely elucidated. Many factors
including age, family heredity, diet, inflammation, and obesity may
all contribute to the development and progression of colorectal
cancer (2). Besides surgery, the
current means of treating colorectal cancer mainly are
radiotherapy, chemotherapy and immunotherapy, which are associated
with unwanted side effects and leading to decreased life quality of
patients. Therefore, the development of safe and effective novel
anticolorectal cancer agents from nature is urgently needed.
In recent years, natural products have been widely
studied by researchers for their remarkable effects and less side
effect in the prevention and treatment of colorectal cancer. For
example, xanthones extracted from Garcinia mengostana
inhibited colon cancer tumorigenesis both in vivo and in
vitro (3). Resveratrol, mainly
found in grapes, has been shown to prevent the development and
progression of colorectal tumors by downregulating Kras expression
(4). Besides results from
laboratory studies, a number of clinical trials also demonstrated
that nature compounds along or as supplements have potential in
treating colorectal cancer (5,6).
These studies suggest that nature compounds may serve as promising
anticancer agents in the prevention and treatment of colorectal
cancer.
Ganoderma lucidum (G. lucidum), also known as
Lingzhi or Reshi, is a traditional Chinese medicine that has been
used in East Asian countries for promoting health and longevity for
over thousands of years (7). In
the past few decades, many researchers studied the biological
function and examined the responsible active components of G.
lucidum. Many studies reported that G. lucidum has
numerous pharmacological effects, including immunomodulatory,
antiallergenic, anti-oxidative, cardiovascular protective,
antidiabetic, and antitumor effects (8,9).
Modern uses of G. lucidum include treatment of coronary
heart diseases, arteriosclerosis, hepatitis, arthritis, nephritis,
asthma, bronchitis and cancer (9).
A diverse group of active compounds including polysaccharide,
triterpenoids, alkaloids, fatty acids, lactones, steroids and
others were identified and isolated from G. lucidum
(10). In recent years, much
attention has been focused on G. lucidum polysaccharide
(GLP), a structurally diverse class of biological macromolecules,
which contribute to numerous pharmacological effects of G.
lucidum. In particular, the antitumor effects of GLP have
attracted much attention.
A substantial number of studies demonstrated that
GLP inhibited carcinogenesis in many types of cancer, including
liver (11), breast (12,13),
leukemia (14), ovarian (15), lung (16,17),
and colorectal cancer (18). Early
studies suggest that the anti-carcinogenic effects of GLP may be
due to its immunomodulatory activity (19). In addition, the study of anticancer
mechanisms of GLP was mainly restricted to GLP extracted from
fruiting body of G. lucidum or mycelia cultivated in liquid
culture medium. Recently, with the advance in sporoderm-breaking
technology, much attention has been paid to chemical components of
the sporoderm-broken spores of G. lucidum (BSGL) and their
versatile biological activities. One study found that the spores of
G. lucidum contain a large amount of bioactive substances
and have a higher bioactivity than the fruiting bodies of G.
lucidum (20). Another study
showed the amount of polysaccharide of sporoderm-broken spores is
1.7 times that of unbroken ones (21). Research has suggested that the
growth inhibition rate of BSGL on HepG2 cells was significantly
higher than unbroken ones (22).
These studies suggest that BSGL may serve as promising anticancer
agent for cancer chemoprevention and therapy.
In this study, we examined the effects and mechanism
of BSGL water extract (BSGLWE), which mainly contains GLP, on
colorectal cancer development and progression in vitro and
in vivo. To our knowledge, this is the first study to
examine the effects of BSGLWE on colon cancer. Our data suggest
that BSGLWE is effective against colorectal cancer development
through regulating cell cycle, apoptosis, proliferation and
necrosis.
Materials and methods
Materials
Hoechst 33342 was purchased from Invitrogen
(Carlsbad, CA, USA). [3-(4, 5-dimethylthia-zol-2-yl)-2,
5-diphenyltetrazolium bromide] (MTT) was obtained from HXBIO
(Hangzhou, China). FITC Annexin V Apoptosis Detection Kit I and
propidium iodide (Pi)/RNase staining kit were purchased from BD
Pharmingen (San Diego, CA, USA). Polyclonal β-actin (#4967s), PARP
(#9542), Bcl-2 (#2876s) antibodies; monoclonal caspase-3 (#9665),
caspase-9 (#9508), Foxo3a (75D8) (#2497p) and horseradish
peroxidase-conjugated secondary antibody were obtained from Cell
Signaling Technology (Danvers, MA, USA). The NAG-1 polyclonal
rabbit antibody was generated in this laboratory as described
before (23). Human GDF15/NAG-1
ELISA kit (NAG-1 also known as growth differentiated factor 15
GDF15) was from R&D Systems (Minneapolis, MN, USA). RNA
extraction kit was from Aidlab Biotech (Beijing, China). The
iScript cDNA synthesis kit and SYBR master mix were purchased from
Bio-Rad (Hercules, CA, USA). The bicinchoninic acid (BCA) assay kit
was from Pierce (Rockford, IL, USA). The Western Lightening™
Plus-ECL Enhanced chemiluminescence Substrate assay kit was from
Perkin-Elmer (Waltham, MA, USA).
Water extract preparation of BSGLWE
The powder of sporodum-broken spores of Ganoderma
lucidum was purchased from Taian Zhenxin LLC (Shandong, China).
The polysaccharides from the powder of sporoderm-broken spores of
G. lucidum were extracted by hot water extraction method.
Briefly, 5 g of sporoderm-broken spores of G. lucidum powder
was placed in 100 ml of double distilled water, and stirred (300
rpm) at 70°C for 12 h. The solution was centrifuged at 4000 rpm for
15 min to remove insoluble materials. The supernatant was
concentrated and freeze-dried using H051 freeze dryer, ScanVac
(LaboGene, Lynge, Denmark). For subsequent cell culture experiment,
the powder of sporoderm-broken spores of G. lucidum water
extract (BSGLWE) was dissolved in Dulbecco's modified Eagle's
medium (DMEM) from Gibco (Gaithersburg, ML, USA) with 10% fetal
bovine serum (FBS) as stock solution of 10 mg/ml, and then passed
through 0.22-µm filter and diluted to required
concentrations.
Cell culture
Human colorectal cancer cell line HCT116 was
purchased from the American Type Culture Collection (ATCC, VA,
USA). Cells were maintained in a humidified atmosphere with 5%
CO2 at 37°C in DMEM supplemented with 10% FBS. For
experiments, cells were seeded in serum-containing medium and at
60–80% confluence, cells were treated with BSGLWE at different
concentrations and treated for certain time adjusted by each
experiment.
Cell viability assay
Cell viability was assessed by MTT assay. Briefly,
cells (1×104) were seeded in 96-well plates and
incubated in DMEM medium containing 10% FBS until cells reach 50%
confluence. HCT116 cells were than treated with 0, 1.25, 2.5, 5,
and 7.5 mg/ml BSGLWE for 24, 48 and 72 h. Then MTT solution (5
mg/ml) was added and incubated for an additional 4 h. After
incubation, supernatants were removed and the remaining
water-insoluble formazan crystals were dissolved in 150 µl
DMSO for 10 min with gentle shacking. The optical density was
measured at 490 nm using a multi-well plate reader (Bio-Tek
Instruments Inc., Winooski, VT, USA). The percentage of viability
was calculated and compared with that of the control cells without
BSGLWE treatment. The 50% inhibitory concentration
(IC50) of BSGLWE was calculated as the 50% decrease in
the optical density compared to untreated controls.
Hoechst 33342 staining
Nuclear fragmentation was examined by Hoechst 33342.
Briefly, HCT116 cells treated with BSGLWE at 0, 1.25, 2.5, 5, 7.5
mg/ml for 24 h, and cells were stained with Hoechst 33342 (10
µg/ml) for 15 min at room temperature. Cells were observed
using a fluorescence microscope (Olympus).
Flow cytometric analysis of cell cycle
and apoptosis
Flow cytometric assay was used for cell cycle and
apoptosis assays. Briefly, for cell cycle determination, equal
numbers of HCT116 cells (2×105) were seeded in 6-well
plate per well and incubated with different concentration of BSGLWE
(0, 5, 7.5 mg/ml) for 36 h. The cells were washed with PBS and then
collected, and fixed in 70% ice cold ethanol, and storage at −20°C
for at least 2 h. Cells were then washed with PBS twice, and
centrifuged for 10 min at 1200 rpm and aspirate the supernatant,
and resuspended cells in 0.5 ml of PI/RNase staining buffer (BD
Pharmingen) for 15 min at room temperature, and DNA content was
immediately analyzed using Guava Easycyte HT flow cytometry system
(Guava Technologies, Merck KGaA, Darmstadt, Germany), and
quantified using ModFit 3.2 software (Verity Software House,
Topsham, ME, USA). For apoptosis examination, approximately
2×105 cells per well were also seeded in 6-well plates
and treated with 0, 1.25, 2.5, 5, and 7.5 mg/ml BSGLWE for 24, 36
and 48 h. Cells were then collected and stained with Annexin
V-FITC/PI at room temperature in the dark and then analyzed by flow
cytometer (Guava Technologies, Merck KGaA). The percentage of
Annexin V−/PI+ (necrosis), Annexin
V+/PI− (early apoptosis) and Annexin
V+/PI+ (late apoptosis) cells were calculated
according to manufacturer's instruction (BD Pharmingen). Samples
were subsequently analyzed by flow cytometer (Merck Millipore
Corp., Darmstadt, Germany).
In vivo tumor xenograft study
All the experimental procedures were conducted
following the Guide for the Use and Care of Laboratory Animals of
the National Institutes of Health. This study was approved by the
Committee on the Ethics of Animal Experiments of Zhejiang Chinese
medical university (Permit Number: SYXK 2012–0002). All procedures
in this protocol are in compliance with the Animal Welfare Act
Regulations. Four-week-old male BALB/C nude mice were kept in
Specific Pathogen Free (SPF) environment. After 2 weeks of
adaptation, mice were randomly divided by weight. HCT116 cells were
injected subcutaneously (s.c.) into the left flank of each nude
mouse (5×106 cells in 200 µl PBS). The mice were
randomized into treatment and control groups: control group
(saline, n=18), low dose (150 mg/kg, n=18), high dose (300 mg/kg,
n=18), 5-FU treatment (20 mg/kg, n=8).
The day after injection of tumor cells, the high and
low dose group mice were treated with BSGLWE intraperitoneally per
day, while the control group mice were injected saline per day, and
5-FU was given every two days through intraperitoneal
administration. However, 5-FU administration was reduced to every
four days after a dramatic weight loss at 2 weeks after HCT116 cell
injection. Tumor volume and body weights were measured twice a week
and palpable tumors were measured in two dimensions (length and
width) using a digital vernier caliper (0.01 mm). Tumor volume was
calculated using the equation V (mm3) = length × width
2/2. Six weeks after injection, all mice were
sacrificed. At necropsy, the xenograft tumors were carefully
excised and weighed. Half of all tumors were fixed in 10% neutral
formalin and processed for hematoxylin and eosin (H&E) and
immunohistochemical staining. The rest of the tumor tissue was
snap-frozen in liquid nitrogen and stored at −80°C for subsequent
analysis. Terminal blood was collected by cardiac puncture for the
analysis of the circulating level of NAG-1/GDF15 secreted from
HCT116 xenografts.
RNA exaction and quantitative real-time
PCR
Total RNA was extracted from HCT116 cells and HCT116
xenograft tumors by RNA extraction kit as described by Aidlab
Biotech. Both the quantity and quality of total RNA were analyzed
by the Agilent Bioanalyzer 2100 system. Total RNA (1 µg) was
reverse transcribed with an iScript cDNA synthesis kit (Bio-Rad).
Real-time PCR was performed to determine the expression of listed
genes using SYBR PCR master mix (Bio-Rad) on CFX96 Real-time PCR
system (Bio-Rad). β-actin was used as the reference gene for all
samples. The PCR conditions consisted of 40 cycles, with 5 sec
denaturation at 95°C, 30 sec annealing at 60°C and 5 sec extension
at 65°C. The relative expression of mRNA for each sample was
calculated as follows: ΔCt = Ct (sample) - Ct (β-actin), ΔΔCt
(sample) = ΔCt (sample) −ΔCt (calibrator). The fold change in mRNA
was calculated through relative quantification (2−ΔΔCt).
Table I shows the primer sequences
of all the primers used in this experiment.
 | Table IPrimers used in qRT-PCR. |
Table I
Primers used in qRT-PCR.
| Primers | Forward | Reverse |
|---|
| β-actin |
CTGGAACGGTGAAGGTGACA |
AAGGAACTTCCTTGAACAATGCA |
| Bcl-2 |
AAGAGCAGACGGATGGAAAAAGG |
GGGCAAAGAAATGCAAGTGAATG |
| Surviving |
GCATGGGTGCCCCGACGT |
TGGCTCCGGCCAGAGGCCTCAA |
| NAG-1 |
CTCCAGATTCCGAGAGTTGC |
AGAGATACGCAGGTGCAGGT |
| NF-κB |
ATGGCTTCTATGAGGCTGAG |
GTTGTTGTTGGTCTGGATGC |
| TNF-α |
AGAGGGAGAGAAGCAACTACA |
GGGTCAGTATGTGAGAGGAAGA |
| c-FOS |
GTGGCTTCCCTTGATCTGACTG |
AACAGGAAGTCATCAAAGGGCT |
| FOXO3 |
GCAAACCCTCTCGGACTCTC |
CCCACGTTCAAACCAACAAC |
| Cyclin A2 |
ATGTCACCGTTCCTCCTTG |
GGGCATCTTCACGCTCTATT |
| Cyclin B1 |
GCCAATAAGGAGGGAGCAGT |
ACCTACACCCAGCAGAAACC |
| Cyclin D1 |
GTGGCCTCTAAGATGAAGGAGA |
GGAAGTGTTCAATGAAATCGTG |
| P16 |
ACCAGAGGCAGTAACCATGC |
TGATCTAAGTTTCCCGAGGTTT |
| P21 |
TTAGCAGCGGAACAAGGAGT |
CGTTAGTGCCAGGAAAGACA |
| WEE1 |
TGTGGTGGTGTGCTGCTTAT |
TTCAAAGGGAGGGTATGTCTG |
| E2F 1 |
CCAACTCCCTCTACCCTTGA |
GTCTCCCTCCCTCACTTTCC |
| RB1 |
ACTCTCACCTCCCATGTTGC |
TGCACTCCTGTTCTGACCTC |
| HIF-1α |
GGGCAATCAATGGATGAAAG |
AGTAATTCTTCACCCTGCAG |
| FADD |
ACGCTTCGGAGGTAGATG |
CCTGGTACAAGAGGTTCA |
| TRAF2 |
CACCGGTACTGCTCCTTCTG |
TGAACACAGGCAGCACAGTT |
| Caspase-8 |
CCAGAGACTCCAGGAAAAGAGA |
GATAGAGCATGACCCTGTAGGC |
Western blotting
Total protein from HCT116 cells and xenograft tumors
were extracted using standard methods and protein concentrations
were determined by BCA protein assay kit. A total of 40 µg
of protein were loaded onto a 10 or 12% SDS-polyacrylamide gel and
electrophoresed at 100 V for 2 h. Separated proteins were
transferred onto a PVDF (polyvinylidene difluoride) membrane at 100
V for 2 h on ice. After transfer, membranes were blocked with 5%
nonfat dry milk in 1X TBST (Tris-buffered saline with Tween; 50
mmol/l of Tris, pH 7.5, 150 mmol/l of NaCl, 0.1% Tween-20) at room
temperature for 1 h. The membrane was probed with NAG-1, Bcl-2,
PARP, Foxo3a or β-actin primary antibodies overnight at 4°C and
then secondary antibody at RT for 1 h according to manufacturer's
instructions. The membrane was stripped using Restore Western Blot
Stripping Buffer according to manufacturer's instruction. After
stripping, the membrane was re-probed for β-actin as a loading
control. The signals were detected using the Western Lightning Plus
ECL-enhanced chemiluminescence substrate according to
manufacturer's instruction. ImageJ 1.41 software (Bethesda, MD,
USA) was used for the calculation of the optical density.
Enzyme-linked immune sobert assay
HCT116 cells (2×105 cells/well) were
plated in 6-well plate and incubated at 37°C. After treatment with
BSGLWE for 48 h, cell culture medium was collected for ELISA
analysis. Total protein concentrations were determined by BCA assay
in cell lysates. The quantification of GDF15/NAG-1 protein was
determined using the Quantikine GDF15/NAG-1 ELISA kit (R&D).
The concentration of NAG-1 in cell culture medium and serum of nude
mice were determined by comparing their optical density to standard
curve. Data were presented as normalized by protein concentration
in cell culture medium or by tumor weights in serum of nude
mice.
Immunohistochemistry
Formalin-fixed tumor tissues were embedded in
paraffin and cut into 4-µm sections. The sections were then
stained with H&E for pathological evaluation. The rest of the
paraffin blocks were cut into 4-µm sections. The sections
were deparaffinized using citric acid buffer (pH 6.0) and incubated
for 8 min at 100°C. The slides were treated with 3% hydrogen
peroxide to block endogenous peroxidase activity and then incubated
with 1% bovine serum albumin (BSA) for 25 min. Next, the slides
were incubated overnight at 4°C with anti-human primary antibodies:
Bcl-2 (GB12008, 1/100 diluted in 1% BSA), Ki67 (GB13030-2, 1/1000
diluted in 1% BSA), PCNA (GB11010, 1/500 diluted in 1% BSA). All
primary antibodies were obtained from Wuhan Goodbio Technology Co.,
Ltd. (Wuhan, China). The slides were then incubated with 5
µg/ml biotinylated anti-goat IgG secondary antibody (Dako,
Carpinteria, CA, USA) for 50 min at room temperature. After
washing, slides were stained with 3,3-diaminobenzidine (DAB)
(Dako), washed and counter-stained with hematoxylin, dehydrated,
and then mounted with a coverslip. All images were captured using
an inverted fluorescence microscope (Nikon, Japan).
Statistical analysis
GraphPad Prism 5 (GraphPad Software, Inc., La Jolla,
CA, USA) was used for all statistical analysis. Data are expressed
as mean ± standard error (SE). Differences between groups were
examined for statistical significance by t-test. Two-sided P-values
were calculated, and a value of P<0.05 was considered to be
statistically significant.
Results
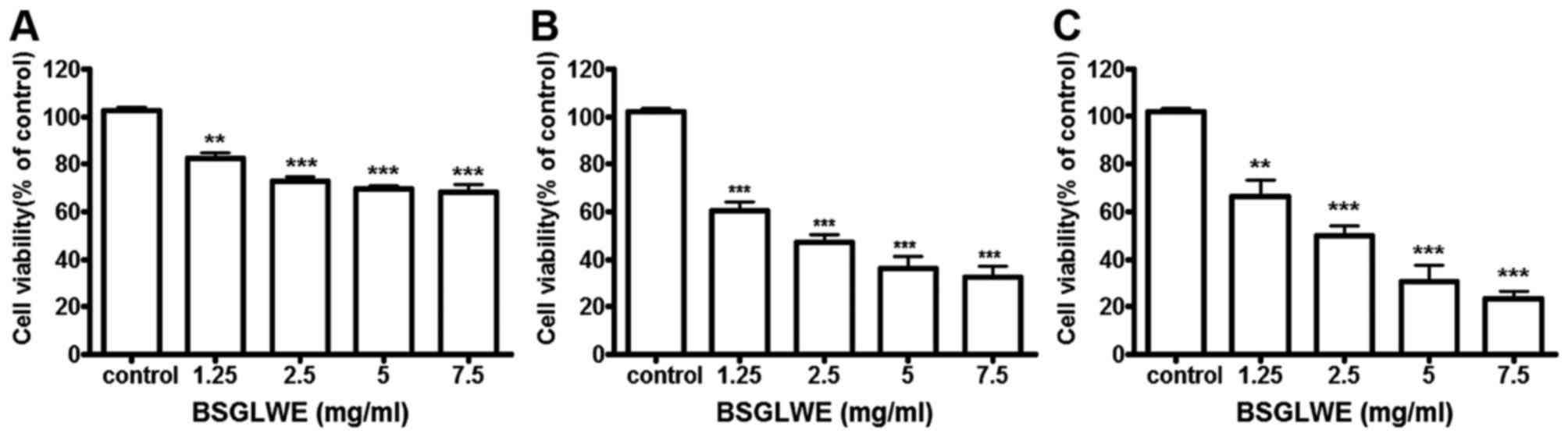
BSGLWE inhibits cell viability in a dose-
and time-dependent manner
To explore the growth inhibitory potential of BSGLWE
against colon cancer, HCT116 cells were treated with BSGLWE at
various concentrations for 24, 48 and 72 h. MTT assays revealed
that cell viability was significantly decreased upon BSGLWE
treatment in a time- and dose-dependent manner in HCT116 cells
(Fig. 1). Specifically, at 24 h,
with the increase of BSGLWE from 0 to 7.5 mg/ml, cell viability
decreased from 100 to 68.36% in HCT116 cells (p<0.001, Fig. 1A). This inhibitory effect was
further enhanced upon 48 and 72 h of treatments (p<0.001,
Fig. 1B and C). Using highest
concentration as example, HCT116 cells treated with 7.5 mg/ml
BSGLWE reduced cell proliferation to 68.36±3.02, 32.66±4.66 and
23.59±2.81% at 24, 48 and 72 h, respectively (p<0.001),
suggesting that longer incubation with BSGLWE could markedly
increase cytotoxicity of BSGLWE to HCT116 cells. The
IC50 was determined to be 2.24 mg/ml and 2.40 mg/ml at
48 and 72 h, respectively (data not shown).
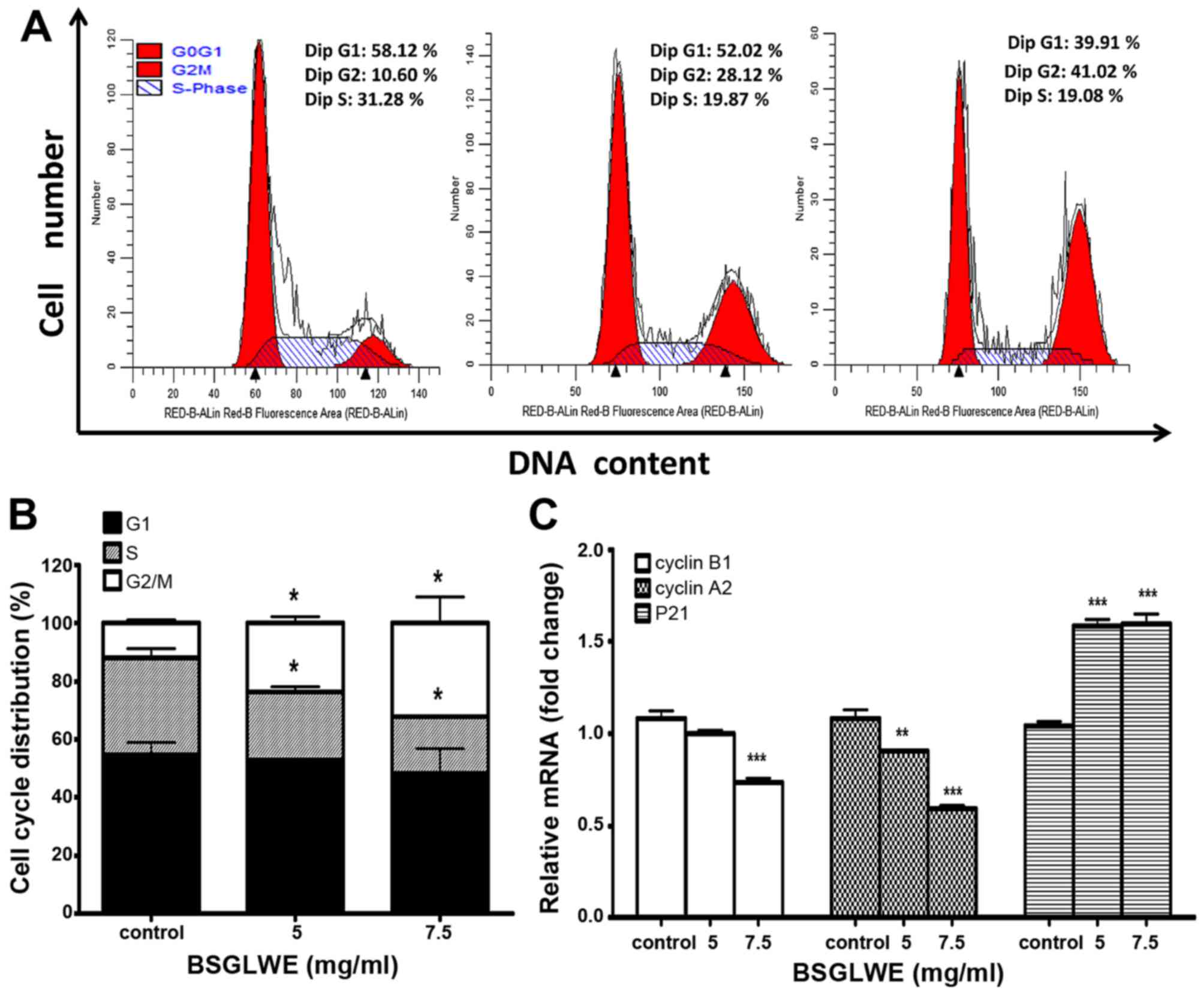
BSGLWE induces cell cycle arrest at G2/M
phase
In order to investigate the anticancer mechanism of
BSGLWE in HCT116 cells, we determined the effects of BSGLWE on cell
cycle distribution by flow cytometry analysis. As shown in Fig. 2A and 2B, the ratio of HCT116 cells in G0/G1
phase slightly decreased upon BSGLWE treatments at non-significant
level, while the ratio of cell population in S phase significantly
decreased (p<0.05). Compared to control cells, BSGLWE
significantly increased percentage of G2/M phase from 12.00±1.09 to
23.75±2.21% and to 32.20±8.85% upon 5 and 7.5 mg/ml of treatments,
respectively (p<0.05, Fig.
2B).
To confirm this result, the expression levels of
G2/M checkpoint regulators such as cyclin B1 and cyclin A2 were
examined. As shown in Fig. 2C, the
expression of cyclin B1 and cyclin A2 at mRNA levels were
significantly downregulated by BSGLWE treatments (p<0.001). In
addition, the mRNA level of P21, a cell cycle arresting protein,
was significantly upregulated (p<0.001, Fig. 2C). Taken together, our results
indicate that BSGLWE may inhibit HCT116 cell proliferation through
regulating key genes involved in arresting cell cycle progression
at G2/M phase.
BSGLWE induces apoptosis in vitro
To further elucidate the mechanism of cell death
induced by BSGLWE in HCT116 cells, Hoechst 33342 staining was
carried out. The changes in the cell nuclei were observed under the
fluorescence microscope. As shown in Fig. 3A, the nuclei of HCT116 cells had
blue fragmentation compared with control cells as indicated by
distinct features of condensation, coagulation, and fragmentation
of the nuclear chromatin, as well as typical apoptotic bodies as
highlighted in circles (Fig. 3A).
We also used flow cytometry analysis to further study the ability
of BSGLWE in inducing apoptosis in HCT116 cells. As shown in
Fig. 3B, we observed that the
amount of Annexin V+/PI− (early apoptosis)
and Annexin V+/PI+ (late apoptosis) stained
cells were both increased significantly upon BSGLWE (1.25, 2.5, 5,
and 7.5 mg/ml) treatments dose-dependently, and in a time-dependent
manner at 24, 36 and 48 h (p<0.001). The rate of early and late
apoptotic cells were quantified and depicted in Fig. 3C.
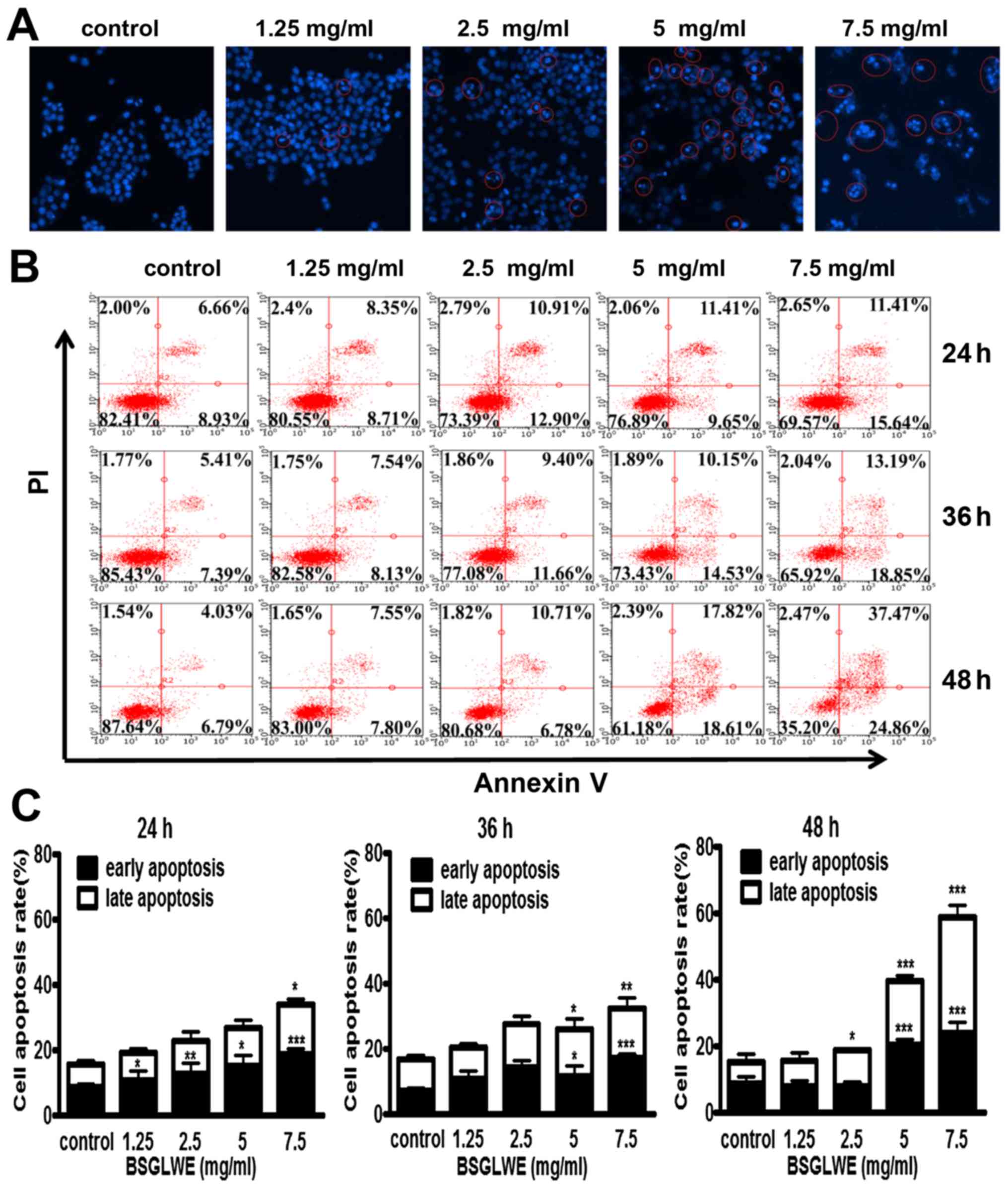 | Figure 3BSGLWE induces apoptosis in HCT116
cells. (A) Cells were treated with different concentration of
BSGLWE for 24 h. Hoechst 33342 staining was used to analyze the
apoptotic cells. (B) BSGLWE (0, 1.25, 2.5, 5, 7.5 mg/ml) induced
apoptosis in HCT116 at 24, 36 and 48 h. Apoptosis was quantified by
Annexin V/FITC flow cytometry. (C) The apoptosis ratio of HCT116
cells increased in dose-/time-dependent manner with BSGLWE
treatment. (D and E) Quantification of the mRNA levels of
apoptosis-related genes after treatment with BSGLWE. (F) The
expression of apoptosis-associated proteins, Bcl-2, PARP, NAG-1,
caspase-3 and caspase-9 were assessed by western blotting. β-actin
used as an internal control. (G) Concentration of NAG-1 in cell
culture medium was examined by ELISA assay, data presented was
normalized by concentration of protein lysates. Data are present as
mean ± SE of three independent experiments, *p<0.05;
**p<0.01; ***p<0.001, as compared with
control group. |
The relative mRNA expression levels of different
regulatory genes involved in apoptosis were then determined by
qRT-PCR. Treatment with different concentrations of BSGLWE
(1.25–7.5 mg/ml) upregulated the expression of survivin and reduced
the expression of Bcl-2 (p<0.001, Fig. 3D). However, the mRNA level of bax
was not changed upon BSGLWE treatment (data not shown).
Additionally, the expression of Bcl-2 at protein level was also
reduced by BSGLWE at dose-dependent manner upon 48 and 72 h of
treatments as determined by western blotting (Fig. 3F). In particular, higher
concentrations of BSGLWE (5 and 7.5 mg/ml) seemed to have more
effects then lower doses in inducing apoptosis in HCT116 cells.
In addition, BSGLWE treatment downregulated the
pro-caspase-3 and pro-caspase-9 expression at protein levels as
determined by western blotting in a time-dependent manner (Fig. 3F), suggesting caspase activation.
We also found total PARP was cleaved and cleaved-PARP was
significantly increased by BSGLWE in HCT116 cells at 48 and 72 h
(Fig. 3F).
Nonsteroidal anti-inflammatory drug-activated gene
(NAG-1) or growth differentiated factor 15 (GDF15), a pro-apoptotic
gene, is a divergent member of the transforming growth factor β
(TGF-β) superfamily (24).
Previous studies have reported that NAG-1 plays an important role
in inhibiting tumor growth (25–28).
It has been well studied that many natural products demonstrate
their anticancer effects through upregulating NAG-1 expression
(24). Our previous studies and
results from many other laboratories suggest that NAG-1 plays a key
role in inhibiting cancer cell proliferation through inducing
apoptosis (24,25). For example, Piyanuch et al
demonstrated that berberine-induced apoptosis of colon cancer cells
was through upregulating the expression of NAG-1 (29). Therefore, we also determined
effects of BSGLWE on NAG-1 induction in HCT116 cells. As shown in
Fig. 3E and F, BSGLWE
significantly induced the expression of NAG-1 at both mRNA and
protein levels (p<0.001). Since NAG-1 is a secreted protein, we
used ELISA to quantify NAG-1 concentration in cell culture medium.
After normalized by total protein concentration in cell lysates,
our results showed a positive correlation between the secretion of
NAG-1 in cell culture medium and the concentration of BSGLWE at 5
and 7.5 mg/ml (p<0.01) (Fig.
3G). However, low doses (1.25 and 2.5 mg/ml) of BSGLWE seemed
not potent enough to induce NAG-1 secretion in HCT116 cells. Taken
together, these results suggest that BSGLWE significantly induced
apoptosis in colorectal cancer HCT116 cells through regulating key
molecules involved in apoptosis cascades. NAG-1 may play an
important role in BSGLWE-induced apoptosis and growth inhibition in
HCT116 cells.
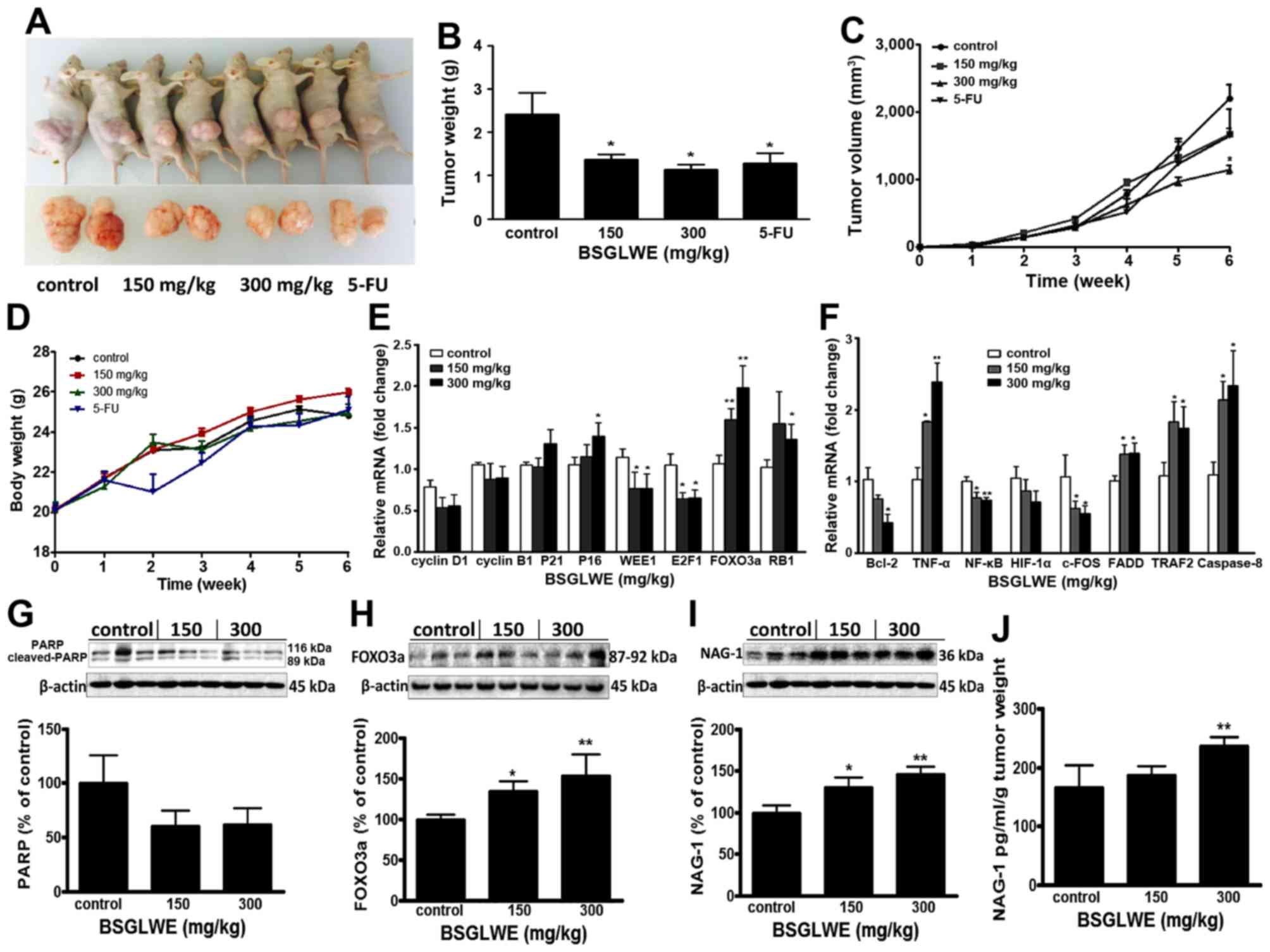
BSGLWE reduces tumor growth in colon
cancer xenograft in nude mice
To evaluate the antitumor effects of BSGLWE in
vivo, we examined the effects of low dose and high dose of
BSGLWE (150 mg/kg vs. 300 mg/kg) by oral gavage on tumor growth in
a mouse tumor xenograft model. 5-FU was used as positive control.
Fig. 4A shows two representative
photograph of the average size of the tumor volume from each group,
which indicates reduced tumor volume in BSGLWE treated group
compared with control group. Both lower dose and higher dose of
BSGLWE inhibited HCT116 xenograft tumor growth and decreased the
final tumor volume in dose-dependent manner by 23.8 and 47.8%
(P<0.05), respectively (Fig.
4C). The final tumor weights at necropsy of the two doses were
all significantly lower than control group (p<0.05, Fig. 4B). Final tumor weights of the four
groups were 2.22±0.11 g (control), 1.27±0.19 g (150 mg/kg),
1.00±0.21 g (300 mg/kg) and 1.28±0.23 g (5-FU) (p<0.05)
(Fig. 4B).
Compared with the control group, the body weight of
BSGLWE treated mice did not change significantly, while the body
weights of 5-FU treated group decreased significantly two weeks
after injection (Fig. 4D).
Therefore, we had to reduce the frequency of 5-FU administration to
nude mice. In addition, no other adverse effects such as skin
ulcerations or toxic death were observed in BSGLWE groups. This
result suggests that BSGLWE may have less toxicity to mice compared
to 5-FU.
In order to study how BSGLWE inhibited tumor
development in vivo, we examined the expression of related
genes and proteins that regulate the cell cycle, proliferation, and
apop-tosis by qRT-PCR and western blotting in xenograft tumor
samples. As shown in Fig. 4E, we
found the relative expression of cell cycle inhibitors, P16 and the
retinoblastoma gene (RB1) at mRNA levels, were significantly
increased in 300 mg/kg BSGLWE treated tumors compared to controls,
while the expression of P21, also a cell cycle inhibitor, was
increased but at non-significant level. In addition, WEE1, E2F1
mRNAs were significantly decreased in HCT116 xenograft tumors upon
BSGLWE treatments (p<0.05) (Fig.
4E). It was reported that WEE1 is upregulated in several types
of cancer, and inhibition of WEE1 in tumor could cause mitotic
catastrophe in glioblastoma cancer (30–32).
E2F1 transcription factor plays a positive role in G1/S phase
progression (33). Moreover, both
mRNA and protein levels (as depicted in histograph) of FOXO3a, a
cycle progression inhibitor, were significantly upregulated in
xenograft tumors by BSGLWE treatment in a dose-dependent manner
(Fig. 4E and H). The high
expression of FOXO3a not only causes cell cycle stagnation in the
mitotic phase, but also make the cells more susceptible to injury
and apoptosis (34). We also
examined the expression of cyclin D1 and cyclin B1, and found they
were reduced upon BSGLWE treatment, but at non-significant levels
(Fig. 4E). These results indicate
that BSGLWE may inhibit HCT116 xenograft tumor development through
inhibiting cell cycle progression.
In addition, we examined the expression of apoptosis
related genes such as Bcl-2, TNF-α, NF-κB, HIF-1α, FADD, TRAF2,
caspase-8 and c-FOS by qRT-PCR. As shown in Fig. 4F, BSGLWE significantly reduced the
expression of anti-apoptosis genes including Bcl-2, NF-κB, and
c-FOS in xenograft tumors compared to controls in a dose-dependent
manner (p<0.05) (Fig. 4F),
while the pro-apoptotic gene TNF-α, caspase-8, TRAF2, and FADD were
significantly induced by BSGLWE. We also found PARP expression was
downregulated slightly as determined by western blotting
(p>0.05, Fig. 4G).
To further examine whether NAG-1 may also play a
role in the antitumor effects of BSGLWE in vivo, we examined
the expression of NAG-1 in xenograft tumors and serum samples.
Interestingly, we found that NAG-1 protein expression was
significantly upregulated in a dose-dependent manner in HCT116
xenograft tumors upon BSGLWE treatment (p<0.01, Fig. 4I). We then measured the
concentration of NAG-1 in serum of tumor-bearing mice by ELISA, and
found that after normalized by tumor weights, the relative
concentration of NAG-1 protein was increased upon BSGLWE treatment
with a significant upregulation by 300 mg/kg treatment (p<0.01,
Fig. 4J).
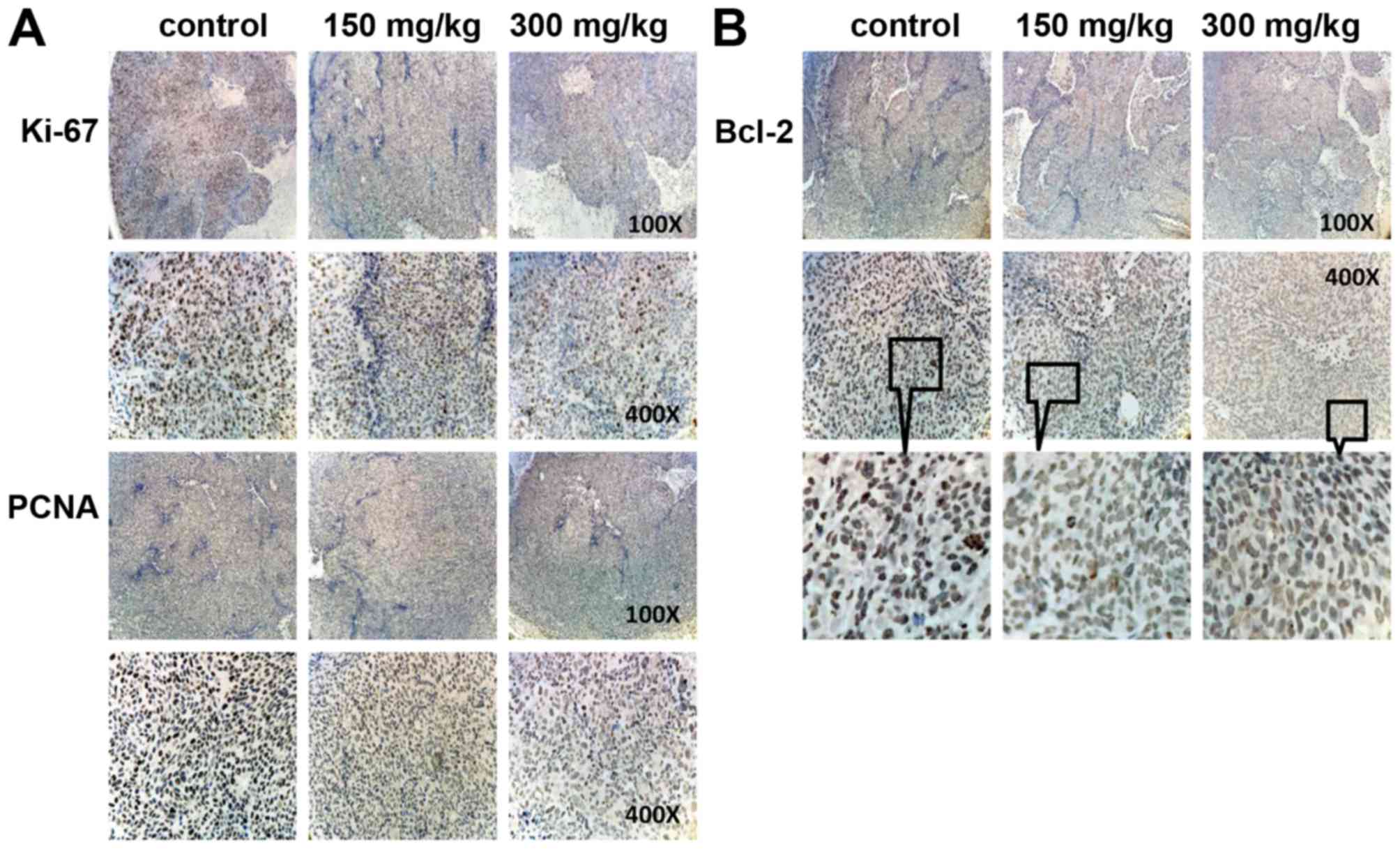
Immunohistochemistry was done to determine
expression of PCNA and Ki67, which are important cell nuclear
proliferation markers. The expression of PCNA and Ki67 were
markedly decreased in BSGLWE treatment groups dose-dependently
compared with control group (Fig.
5A), indicating that the BSGLWE effectively inhibit
proliferation of colon tumor cells in vivo. In addition, we
found anti-apop-totic marker Bcl-2 was markedly reduced in tumor
sections of BSGLWE treatment groups (Fig. 5B). Taken together, these data
suggest that the inhibitory effects of BSGLWE on colorectal
tumorigenesis in xenograft model may be through regulating key
molecules involved in inhibition of cell proliferation, increase of
cell cycle arrest and induction of apoptosis.
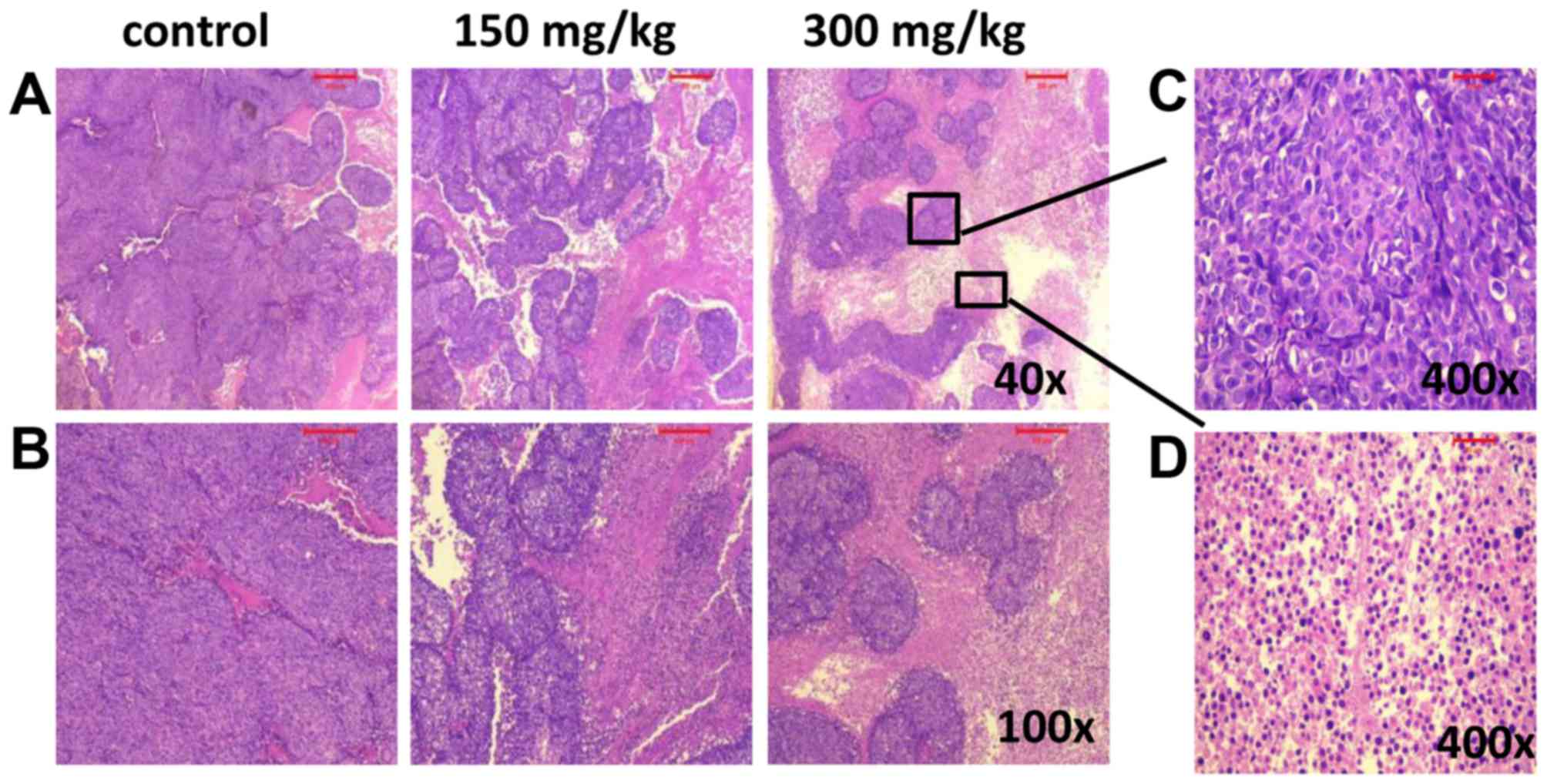
BSGLWE induces necrosis in nude mouse
xenograft tumors
H&E was used to observe the pathological changes
in xenograft tumors after different treatments. Compared with
control group, the tumor necrosis area increased in the xenograft
tumors treated with BSGLWE. The pictures indicate that BSGLWE
obviously induced necrosis in a dose-dependent manner in the
xenograft tumors compared with control group (Fig. 6A and B). The zoomed-in image from
300 mg/kg treated (Fig. 6A upper
right image), necrotic area was mainly characterized by large
blurred, massive, unstructured red-stained material, mixed with
blue-stained nucleus fragments in the tumor tissue as compared with
dark purple stained living cells in non-necrotic area (Fig. 6C and D).
Discussion
In recent years, the interest in natural compounds
for prevention and treatment of cancer has increased. Various
classes of anticancer agents derived from plants including
alkaloids, saponins, polysaccharides, terpenoids and flavonoids
have been extensively studied in laboratories and clinical
investigations. The anticancer activities of the water-soluble
extract of G. lucidum, which mainly contains GLP, has also
been recognized. Available studies suggest that the anticancer
mechanism of GLP may include immunomodulation (35,36),
suppression of tumor angiogenesis (16), inhibition of cancer cell invasion
and metastasis (37), inhibition
of tumor cell proliferation and induction of tumor cell apoptosis
(38), as well as reduction of the
anticancer drug resistance (39).
However, GLP is a mixture of peptidoglycan, glucose,
heteropolysaccharide and other polysaccharides (8,10).
Due to the complex structure of GLP and the limitation of
separation and analysis, the composition of GLP has not yet been
fully identified. Bioactive GLP has been isolated from the fruiting
bodies of G. lucidum, from the mycelia cultivated in liquid
culture medium, and from the spores of G. lucidum (19). Recently, studies found that the
BSGL possesses more bioactive GLP than fruiting bodies of G.
lucidum or mycelia and also showed more potent ability in
inhibiting cancer cell growth than unbroken spores (20–22).
In this study, we demonstrate that BSGLWE (mainly
contains GLP) is potent in inhibiting colorectal cancer cell growth
and tumor development through causing cell cycle arrest, inhibiting
cell proliferation and inducing apoptosis both in vitro and
in vivo. NAG-1 gene, which plays an important role in
inhibiting carcinogenesis in many cancers, may be a key target of
BSGLWE for its chemopreventive activity in colorectal cancer. To
our knowledge, this is the first study to examine the
anticarcinogenic effects and mechanisms exerted by BSGLWE in
colorectal cancer, and first to show NAG-1 could be induced by
BSGLWE in cancer cells.
Cancer is caused by the imbalance of cell
proliferation and cell death, and is considered as a genetic
disease that occurs with increased genetic instability involved in
regulation of many cellular processes (13). These processes include
proliferation, apoptosis, angiogenesis, cell cycle progression and
invasion (40). Cell cycle
disorder is one of the main contributors to carcinogenesis
(41,42). The normal cells operate in the
exact chronological order through G1-S-G2-M phase under the precise
control by cell cycle molecular network system (41,42).
Deregulation of cell cycle would cause imbalance between cell
proliferation and apoptosis, which may eventually lead to cancer.
Our results suggest that BSGLWE could arrest cell cycle at G2/M
phase as determined by flow cytometry in HCT116 cells. BSGLWE
significantly decreased mRNA levels of cyclin A2, cyclin B1 and
increased the expression of P21 in HCT116 cells. Both
downregulation of cyclin A2 and cyclin B1 induced cell cycle arrest
at G2/M phase (43). P21 has a
wide range of kinase inhibitory activity inhibiting the activity of
various cyclin-CDK complexes that cause primarily G1 arrest
(44,45). However, some studies suggest that
upon upregulation, P21 also plays an important role in G2/M arrest
(46). These results suggest that
BSGLWE may cause HCT116 cell G2/M phase arrest through upregulating
P21 and downregulating cyclin A1 and cyclin B2. Consistent with our
study, Zhao et al found that G. lucidum whole extract
induced cell cycle arrest at G2/M phase in ovarian cancer cells
(47). Wang et al found
that GLP extracted from fruiting body of G. lucidum could
inhibit breast cancer cell proliferation through G2/M phase cell
cycle arrest (48). However, at
present, no study examined effects of GLP extracted from
sporoderm-broken spores on cell cycle arrest.
We also found that the expression of WEE1 and E2F
were significantly downregulated while the expression of P16,
FOXO3a and RB1 were significantly upregulated in the xenograft
tumors of the nude mice upon BSGLWE treatment. P16 can compete with
cyclin D1 binding to CDK6 or CDK4 and thus specifically inhibit
CDK4 or CDK6 activity, eventually leading to cell cycle arrest at
G1 phase (49,50). WEE1 is a key gene for G2/M phase
arrest, inhibition or downregulation of WEE1 kinase could lead to
mitotic catastrophe in glioblastoma cancer cells (31,32).
E2F transcription factor and Rb gene family play an important role
in regulating cell cycle progression from the G1 phase to the S
phase (51). These results suggest
that BSGLWE may cause cell cycle arrest at different check points
in HCT116 xenograft tumors.
GLP has also been reported to induce apoptosis in
several cancer cells (13,18). Apoptosis is regulated by a number
of genes and is a precise process. In our study, BSGLWE
downregulated the expression of Bcl-2, a key anti-apoptosis
molecule, both in HCT116 cells and xenograft tumors as determined
by western blotting, qRT-PCR and immunostaining. The
poly(ADP-ribose) polymerase (PARP), an apoptosis marker, was
cleaved both in HCT116 cells and xenograft tumors upon BSGLWE
treatment. The transcription factors, Forkhead box O (FOXO) genes,
are involved in multiple signaling pathways and play critical roles
in a number of physiological and pathological processes, including
cancer (52). One family member,
FOXO3a, has been well defined to be involved in cell cycle arrest
and apoptosis induction, and FOXO3a induction is important for
tumor suppression (53). In our
study, BSGLWE treatment significantly increased FOXO3a expression
both at mRNA and total protein levels in xenograft tumors,
suggesting FOXO3a may be a potential molecular target of BSGLWE.
Study by Dey et al demonstrated that both up regulation of
mRNA and total protein levels of FOXO3a, but not the phosphorylated
form, play a key role in 3β-Adiol-induced apoptosis in prostate
cancer cells (54). As mentioned
earlier, transcription factor NF-κB and cytokine TNF-α both play a
key role in cellular processes including inflammation, apoptosis
and cell cycle arrest.
Several studies have demonstrated that there is a
signaling interplay between FOXO3a, TNF-α, and NF-κB in regulating
physiological and pathological diseases (55–57).
For example, Lee et al found that FOXO3a promoted remarkable
apoptosis in human endothelial cells (HUVECs) through upregulating
of TNF-α and suppression of NF-κB (58). Correlating with these observations,
we found BSGLWE also increased TNF-α expression and decreased NF-κB
expression in xenograft tumors, suggesting that the
FOXO3a-TNF-α-NF-κB network may also be important molecular targets
for BSGLWE induced apoptosis and cell cycle arrest in colorectal
cancer. However, apoptosis is a complex biological process which
can be divided into two paths: the extrinsic and the intrinsic
pathways, function via mitochondrial and death receptor,
respectively (59). In this study,
we found that BSGLWE also induced FADD, TRAF2, and caspase-8
expression, key regulators of death-receptor-induced apoptosis
pathway, suggesting a more complex regulation of apoptosis by
BSGLWE during colorectal cancer carcinogenesis. More studies are
needed to further differentiate the two pathways and the exact
regulating mechanisms elicited by BSGLWE.
In the xenograft tumor study, we found both high and
low doses of BSGLWE treatment significantly reduced xenograft tumor
size and tumor weights compared to control group. At 0–4 weeks, the
difference of tumor size was not quite obvious between treatment
and control group. However, the growth rate of tumors in BSGLWE
groups, especially the 300 mg/kg group, was significantly slowed
down at the fifth week until end of the study. These results
suggest that BSGLWE may be more effective in inhibiting colorectal
tumor progression at late stage but not being quite effective in
inhibiting the initiation and early development of colorectal
cancer in this study. This observation may also suggest a long-term
treatment with BSGLWE might be necessary for colorectal cancer
prevention and therapeutic purpose in clinical practices. In
agreement with our study, Shi and Qing (60) found that long-term treatment with
G. lucidum significantly reduced death rate in late phase
cancer patients as compared with short-term treatment (9). Few clinical studies in general
suggest that G. lucidum is safe to either normal population
or patients with disease (61,62).
However, the safety and potential toxicity of long-term use of
G. lucidum for therapeutic purpose or prevention purpose
need to be evaluated in future studies.
Cell death has been mainly categorized into
apoptosis, autophagy and necrosis based on morphological features
and biochemical characteristics. Recently, a growing number of
studies indicate necroptosis, distinct from apoptosis and necrosis,
as a new mechanism of cell death (63). Cells can readily switch from one
form of death to another (64). In
this study, we found that the xenograft tumor of mice treated with
BSGLWE showed more necrosis in a dose-dependent manner than the
control group as depicted in Fig.
6. Necrosis is often observed in the internal regions of tumors
where nutrient and oxygen supplies are limited (65). Necrosis is considered to be a form
of cell death characterized by early plasma membrane
permeabilization and organelle swelling and is accompanied by a
significant inflammatory response (66). Traditionally, apoptosis and
autophagy were considered as the most prominent cell death or cell
death-related mechanisms (67). By
now, multiple other cell death modalities such as necrosis were
described and necrosis is recognized to be most likely involved in
the response to chemotherapeutic treatments (67). Other than inducing apoptosis or
autophagy as their anticancer mechanisms, many natural compounds
including genistein, curcumin and Berberine have been reported to
induce necrosis in cancer cells as reviewed by Gali-Muhtasib et
al (68). It is reported that
approximately 90% of cancer patients die of metastasis due to the
resistance of cancer cells to apoptosis inducing drugs (69). Kim et al reported that for
prognosis, the higher the rate of necrosis area adjusted by tumor
volume after chemotherapy the better the prediction of
metastasis-free survival of localized osteosarcoma patients
(70). Therefore, it is critical
to discover anticancer drugs that induce alternative modes of cell
death such as necrosis for cancer chemoprevention and therapy. Our
results suggest that BSGLWE could induce both apoptosis and
necrosis in HCT116 xenograft tumors and thus slow down the rate of
tumor growth.
Body weight change is important in evaluating
toxicity and side effects of a chemotherapy drug or nature product
in animal studies. Our study indicates that BSGLWE had no negative
effects on body weights of nude mice. During the study,
tumor-bearing mice in control group began to lose weight (a
hallmark of cachexia) at the fifth week and body weights of control
group were lowest among all groups at the end of study. Cachexia is
common in cancer patients and is characterized by weight loss due
to skeletal muscle wasting and fat depletion (71). Cachexia causes weakness,
immobility, low tolerance to anticancer therapy, and poor quality
of life, which significantly contributes to cancer-related deaths
in cancer patients (71). In
contrast, the body weights of 150 and 300 mg/kg BSGLWE treated mice
maintained a continuous increase of body weight throughout the
whole experiment, suggesting BSGLWE may have a beneficial effect on
attenuating cancer-induced cachexia in nude mice.
It is worth noting that lower dose of BSGLWE (150
mg/kg) may even have a protective effect since the body weight of
this group was maintained at the highest level among all groups
throughout the study. On the contrary, 5-FU, a commonly used
chemotherapy agent, markedly induced weight loss and severe side
effects in nude mice two weeks after injection of HCT116 cells.
Therefore, we had to reduce the frequency of 5-FU administration to
nude mice at week 2. Surprisingly, we found that after reducing
5-FU administration, tumor growth rate sharply accelerated at week
5, suggesting relapse may occur upon reduction of 5-FU treatment.
However, BSGLWE at either low or high doses did not show any of the
side effects or signs of weight loss, indicating that BSGLWE might
be a safer anticancer agent compared to 5-FU.
The combination of benefit without toxicity
represents the desired end result in the development of effective
therapeutic agents. G. lucidum has been used for thousands
of years as a health promotion and treatment strategy. Other than
studies from laboratories, there are also some published reports of
human trials in the assessment of G. lucidum for treating
several diseases, including cancer. In the process of clinical
treatment of cancer, G. lucidum spore powder is combined
with anticancer drugs to reduce reaction with radiotherapy and
chemotherapy in patients. G. lucidum used as one of the
component of a Chinese medicine herb complex which significantly
improved immune function, overall health and ability to fight
cancer in patients receiving chemotherapy or radiotherapy
medications (72).
A recent meta-analysis which included five
randomized controlled trials (RTC) with a total of 373 cancer
patients showed that patients who had been given G. lucidum
alongside chemo/radiotherapy were more likely to respond positively
compared to chemo/radiotherapy alone (RR 1.50; 95% CI 0.90–2.51,
P=0.02) (73). Among these five
RTCs, four studies showed that patients treated with G.
lucidum had relatively improved quality of life compared to
controls (73). Our study also
showed that BSGLWE protected tumor-bearing mice from weight loss
and reduced the degree of malignancy compared to 5-FU or control
groups. However, evidence from well-designed human clinical trials
is still scarce. In addition, one question is whether antitumor
effects of G. lucidum is a direct activity or is mediated
through effects on immune system modulation, which still is a key
question to be addressed in future studies (19). Therefore, more clinical human
trials are needed to better understand the bioactivity of G.
lucidum, especially GLP from the sporoderm-broken spores.
Investigation can progress in order to use G. lucidum as new
nutraceutical or drug for the prevention and treatment of
colorectal cancer in near future.
NAG-1, a pro-apoptotic gene, is a divergent member
of TGF-β superfamily. Many studies have shown that NAG-1 acts as a
tumor suppressor protein by inhibiting tumor growth and inducing
apoptosis in the early stages of cancer (24–27).
NAG-1 induction may be associated with cell cycle arrest and
apoptosis in a variety of cancer cells (24–27).
NAG-1 is up regulated in human colorectal cancer cells by several
NSAIDs, as well as by dietary compounds including resveratrol,
genistein, diallyl disulfide, conjugated linoleic acid, green tea
catechins, epigallocatechin-3-gallate (EGCG), indole-3-carbinol,
capsaicin and other anticancer agents (24). In this study, BSGLWE upregulated
the expression of NAG-1 both in vitro and in vivo
which have never been reported before.
In HCT116 cell culture medium, NAG-1 concentration
was increased dose-dependently upon BSGLWE treatment as compared to
control cells. In nude mice, the source of serum NAG-1 was derived
from human HCT116 cells. We found NAG-1 protein secreted into serum
of nude mice was also induced upon BSGLWE treatment. Laboratory
studies overall suggest that NAG-1 has an anticancer effect in many
types of cancer primarily through induction of apoptosis (24). However, clinical studies have shown
that the expression of NAG-1 in cancer patient, including colon
cancer, were elevated, especially positively correlated with tumor
stage and grade (24,74). Unfortunately, the exact role of
NAG-1 during tumorigenesis in cancer patients is unknown. In
agreement with results from laboratory studies, our study suggests
that NAG-1 induction by BSGLWE may play a partial role in
BSGLWE-induced apoptosis and cell death in colorectal cancer.
However, more studies are needed to definitively determine whether
the BSGLWE-induced cell death in HCT116 cells is through NAG-1
induction.
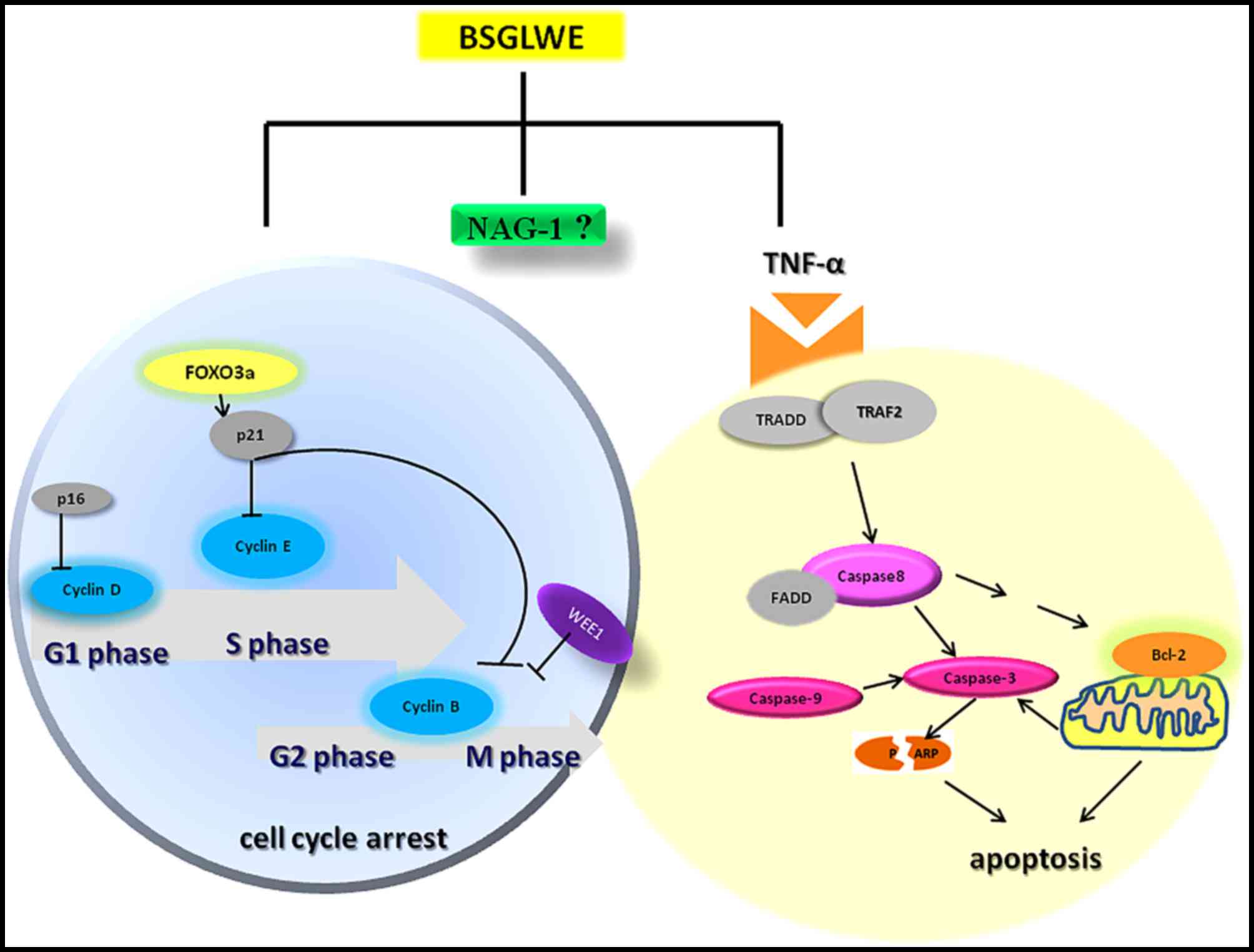
In conclusion, the present study demonstrated that
BSGLWE significantly inhibited colorectal cancer cell proliferation
and tumor growth through deregulating expression of the key
molecules of cell cycle, apoptosis and proliferation as depicted in
Fig. 7. BSGLWE induced NAG-1
expression in HCT116 cancer cells and NAG-1 induction may be
closely related to reduced cell viability and increased apoptosis
and possibly cell cycle arrest upon BSGLWE treatment as modeled in
Fig. 7. To our knowledge this is
the first study showing that BSGLWE could inhibit cell
proliferation in colorectal cancer cells, which may involve NAG-1
induction. However, definitive role of NAG-1 in BSGLWE induced
cytotoxicity in colorectal cancer need to be further elucidated in
future studies. Our results also indicate that BSGLWE may serve as
a novel anticancer agent for colorectal cancer chemoprevention and
therapy.
Abbreviations:
|
NAG-1
|
non-steroidal anti-inflammatory
drug-activated gene-1
|
|
GDF15
|
growth differentiation factor 15
|
|
BSGLWE
|
sporoderm-broken spores of
Ganoderma lucidum water extracts
|
|
GLP
|
Ganoderma lucidum
polysaccharides
|
Acknowledgments
We thank Dr Thomas Eling at National Institute of
Environmental Health Sciences (NIEHS) for critical reading of this
manuscript. We wish to thank Yu Huang at Zhejiang Medical
University animal facility for his kind help in animal care and
technique support. This study was supported by National Natural
Science Foundation of China (grant no. 81473397).
References
|
1
|
Arnold M, Sierra MS, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global patterns and trends in
colorectal cancer incidence and mortality. Gut. 66:683–691. 2017.
View Article : Google Scholar
|
|
2
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar
|
|
3
|
Aisha AFA, Abu-Salah KM, Ismail Z and
Majid AMSA: in vitro and in vivo anti-colon cancer effects of
Garcinia mangostana xanthones extract. BMC Complement Altern Med.
12:1042012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Saud SM, Li W, Morris NL, Matter MS,
Colburn NH, Kim YS and Young MR: Resveratrol prevents tumorigenesis
in mouse model of Kras activated sporadic colorectal cancer by
suppressing oncogenic Kras expression. Carcinogenesis.
35:2778–2786. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Afrin S, Giampieri F, Gasparrini M,
Forbes-Hernandez TY, Varela-López A, Quiles JL, Mezzetti B and
Battino M: Chemopreventive and therapeutic effects of edible
berries: a focus on colon cancer prevention and treatment.
Molecules. 21:1692016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shehzad A, Wahid F and Lee YS: Curcumin in
cancer chemo-prevention: Molecular targets, pharmacokinetics,
bioavailability, and clinical trials. Arch Pharm (Weinheim).
343:489–499. 2010. View Article : Google Scholar
|
|
7
|
Dan X, Liu W, Wong JH and Ng TB: A
ribonuclease isolated from wild Ganoderma lucidum suppressed
autophagy and triggered apoptosis in colorectal cancer cells. Front
Pharmacol. 7:2172016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bishop KS, Kao CH, Xu Y, Glucina MP,
Paterson RR and Ferguson LR: From 2000 years of Ganoderma lucidum
to recent developments in nutraceuticals. Phytochemistry.
114:56–65. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Boh B, Berovic M, Zhang J and Zhi-Bin L:
Ganoderma lucidum and its pharmaceutically active compounds.
Biotechnol Annu Rev. 13:265–301. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wasser SP: Reishi or Ling Zhi (Ganoderma
lucidum). Encyclopedia of Dietary Supplements. Coates PM, Blackman
MR, Cragg GM, Levine M, Moss J and White JD: Marcel Dekker; New
York: pp. 603–622. 2005
|
|
11
|
Li A, Shuai X, Jia Z, Li H, Liang X, Su D
and Guo W: Ganoderma lucidum polysaccharide extract inhibits
hepatocellular carcinoma growth by downregulating regulatory T
cells accumulation and function by inducing microRNA-125b. J Transl
Med. 13:1002015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Loganathan J, Jiang J, Smith A, Jedinak A,
Thyagarajan-Sahu A, Sandusky GE, Nakshatri H and Sliva D: The
mushroom Ganoderma lucidum suppresses breast-to-lung cancer
metastasis through the inhibition of pro-invasive genes. Int J
Oncol. 44:2009–2015. 2014.PubMed/NCBI
|
|
13
|
Shang D, Li Y, Wang C, Wang X, Yu Z and Fu
X: A novel polysaccharide from Se-enriched Ganoderma lucidum
induces apoptosis of human breast cancer cells. Oncol Rep.
25:267–272. 2011.
|
|
14
|
Yang G, Yang L, Zhuang Y, Qian X and Shen
Y: Ganoderma lucidum polysaccharide exerts anti-tumor activity via
MAPK pathways in HL-60 acute leukemia cells. J Recept Signal
Transduct Res. 36:6–13. 2016. View Article : Google Scholar
|
|
15
|
Hsieh TC and Wu JM: Suppression of
proliferation and oxidative stress by extracts of Ganoderma lucidum
in the ovarian cancer cell line OVCAR-3. Int J Mol Med.
28:1065–1069. 2011.PubMed/NCBI
|
|
16
|
Cao QZ and Lin Z-B: Ganoderma lucidum
polysaccharides peptide inhibits the growth of vascular endothelial
cell and the induction of VEGF in human lung cancer cell. Life Sci.
78:1457–1463. 2006. View Article : Google Scholar
|
|
17
|
Sun LX, Li WD, Lin ZB, Duan XS, Li XF,
Yang N, Lan TF, Li M, Sun Y, Yu M, et al: Protection against lung
cancer patient plasma-induced lymphocyte suppression by Ganoderma
lucidum polysaccharides. Cell Physiol Biochem. 33:289–299. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liang Z, Yi Y, Guo Y, Wang R, Hu Q and
Xiong X: Chemical characterization and antitumor activities of
polysaccharide extracted from Ganoderma lucidum. Int J Mol Sci.
15:9103–9116. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ferreira IC, Heleno SA, Reis FS, Stojkovic
D, Queiroz MJ, Vasconcelos MH and Sokovic M: Chemical features of
Ganoderma polysaccharides with antioxidant, antitumor and
antimicrobial activities. Phytochemistry. 114:38–55. 2015.
View Article : Google Scholar
|
|
20
|
Guo L, Xie J, Ruan Y, Zhou L, Zhu H, Yun
X, Jiang Y, Lü L, Chen K, Min Z, et al: Characterization and
immunostimulatory activity of a polysaccharide from the spores of
Ganoderma lucidum. Int Immunopharmacol. 9:1175–1182. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang XL, Hui-Qin WU, Fang H and Lin XS:
Analysis of polysaccharide from broken cellular wall and unbroken
spore of Ganoderma lucidum. Chin Tradit Herbal Drugs. 37:813–816.
2006.
|
|
22
|
Zhao J, Qiu C, Li J, Fan J and Chen K:
inhibitive effect of selenium-enriching wall-broken Ganoderma
lucidum spore powder on HepG2 cells. Life Sci. 79:11292006.
|
|
23
|
Baek SJ, Kim KS, Nixon JB, Wilson LC and
Eling TE: Cyclooxygenase inhibitors regulate the expression of a
TGF-beta superfamily member that has proapoptotic and
antitumorigenic activities. Mol Pharmacol. 59:901–908.
2001.PubMed/NCBI
|
|
24
|
Wang X, Baek SJ and Eling TE: The diverse
roles of nonsteroidal anti-inflammatory drug activated gene
(NAG-1/GDF15) in cancer. Biochem Pharmacol. 85:597–606. 2013.
View Article : Google Scholar :
|
|
25
|
Kang SU, Shin YS, Hwang HS, Baek SJ, Lee
SH and Kim CH: Tolfenamic acid induces apoptosis and growth
inhibition in head and neck cancer: Involvement of NAG-1
expression. PLoS One. 7:e349882012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang X, Chrysovergis K, Bienstock RJ, Shim
M and Eling TE: The H6D variant of NAG-1/GDF15 inhibits prostate
xenograft growth in vivo. Prostate. 72:677–689. 2012. View Article : Google Scholar
|
|
27
|
Arafat K, Iratni R, Takahashi T, Parekh K,
Al Dhaheri Y, Adrian TE and Attoub S: Inhibitory effects of
salinomycin on cell survival, colony growth, migration, and
invasion of human non-small cell lung cancer A549 and LNM35:
Involvement of NAG-1. PLoS One. 8:e669312013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liggett JL, Zhang X, Eling TE and Baek SJ:
Anti-tumor activity of non-steroidal anti-inflammatory drugs:
Cyclooxygenase-independent targets. Cancer Lett. 346:217–224. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Piyanuch R, Sukhthankar M, Wandee G and
Baek SJ: Berberine, a natural isoquinoline alkaloid, induces NAG-1
and ATF3 expression in human colorectal cancer cells. Cancer Lett.
258:230–240. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guertin AD, Martin MM, Roberts B, Hurd M,
Qu X, Miselis NR, Liu Y, Li J, Feldman I, Benita Y, et al: unique
functions of CHK1 and WEE1 underlie synergistic anti-tumor activity
upon pharmacologic inhibition. Cancer Cell Int. 12:452012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mir SE, De Witt Hamer PC, Krawczyk PM,
Balaj L, Claes A, Niers JM, Van Tilborg AA, Zwinderman AH, Geerts
D, Kaspers GJ, et al: In silico analysis of kinase expression
identifies WEE1 as a gatekeeper against mitotic catastrophe in
glioblastoma. Cancer Cell. 18:244–257. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vitale I, Galluzzi L, Castedo M and
Kroemer G: Mitotic catastrophe: A mechanism for avoiding genomic
instability. Nat Rev Mol Cell Biol. 12:385–392. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ogawa H, Ishiguro K, Gaubatz S, Livingston
DM and Nakatani Y: A complex with chromatin modifiers that occupies
E2F- and Myc-responsive genes in G0 cells. Science. 296:1132–1136.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Burgering BM and Medema RH: Decisions on
life and death: FOXO Forkhead transcription factors are in command
when PKB/Akt is off duty. J Leukoc Biol. 73:689–701. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li WJ, Chen Y, Nie SP, Xie MY, He M, Zhang
SS and Zhu KX: Ganoderma atrum polysaccharide induces anti-tumor
activity via the mitochondrial apoptotic pathway related to
activation of host immune response. J Cell Biochem. 112:860–871.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xu Z, Chen X, Zhong Z, Chen L and Wang Y:
Ganoderma lucidum polysaccharides: Immunomodulation and potential
anti-tumor activities. Am J Chin Med. 39:15–27. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu QP, Xie YZ, Li SZ, La Pierre DP, Deng
Z, Chen Q, Li C, Zhang Z, Guo J, Wong C-KA, et al: Tumour cell
adhesion and integrin expression affected by Ganoderma lucidum.
Enzyme Microb Technol. 40:32–41. 2006. View Article : Google Scholar
|
|
38
|
Sun Z, Huang K, Fu X, Zhou Z, Cui Y and Li
H: A chemically sulfated polysaccharide derived from Ganoderma
lucidum induces mitochondrial-mediated apoptosis in human
osteosarcoma MG63 cells. Tumour Biol. 35:9919–9926. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li WD, Zhang BD, Wei R, Liu JH and Lin ZB:
Reversal effect of Ganoderma lucidum polysaccharide on multidrug
resistance in K562/ADM cell line. Acta Pharmacol Sin. 29:620–627.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Borgs L, Beukelaers P, Vandenbosch R,
Belachew S, Nguyen L and Malgrange B: Cell 'circadian' cycle: New
role for mammalian core clock genes. Cell Cycle. 8:832–837. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee B, Sandhu S and McArthur G: Cell cycle
control as a promising target in melanoma. Curr Opin Oncol.
27:141–150. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hochegger H, Takeda S and Hunt T:
Cyclin-dependent kinases and cell-cycle transitions: Does one fit
all? Nat Rev Mol Cell Biol. 9:910–916. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Abbas T and Dutta A: p21 in cancer:
Intricate networks and multiple activities. Nat Rev Cancer.
9:400–414. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lim S and Kaldis P: Cdks, cyclins and
CKIs: Roles beyond cell cycle regulation. Development.
140:3079–3093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Niculescu AB III, Chen X, Smeets M, Hengst
L, Prives C and Reed SI: Effects of p21 (Cip1/Waf1) at both the
G1/S and the G2/M cell cycle transitions: pRb is a critical
determinant in blocking DNA replication and in preventing
endoreduplication. Mol Cell Biol. 18:629–643. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhao S, Ye G, Fu G, Cheng JX, Yang BB and
Peng C: Ganoderma lucidum exerts anti-tumor effects on ovarian
cancer cells and enhances their sensitivity to cisplatin. Int J
Oncol. 38:1319–1327. 2011.PubMed/NCBI
|
|
48
|
Wang J, Zhang L, Yu Y and Cheung PC:
Enhancement of antitumor activities in sulfated and
carboxymethylated polysaccharides of Ganoderma lucidum. J Agric
Food Chem. 57:10565–10572. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Soták M, Sumová A and Pácha J: Cross-talk
between the circadian clock and the cell cycle in cancer. Ann Med.
46:221–232. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Williams RT, Barnhill LM, Kuo HH, Lin WD,
Batova A, Yu AL and Diccianni MB: Chimeras of p14ARF and 16:
Functional hybrids with the ability to arrest growth. PLoS One.
9:e882192014. View Article : Google Scholar
|
|
51
|
Dyson N: The regulation of E2F by
pRB-family proteins. Genes Dev. 12:2245–2262. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fu Z and Tindall DJ: FOXOs, cancer and
regulation of apoptosis. Oncogene. 27:2312–2319. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Nho RS and Hergert P: FoxO3a and disease
progression. World J Biol Chem. 5:346–354. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Dey P, Ström A and Gustafsson JÅ: Estrogen
receptor β upregulates FOXO3a and causes induction of apoptosis
through PUMA in prostate cancer. Oncogene. 33:4213–4225. 2014.
View Article : Google Scholar
|
|
55
|
Hoesel B and Schmid JA: The complexity of
NF-κB signaling in inflammation and cancer. Mol Cancer. 12:862013.
View Article : Google Scholar
|
|
56
|
Chung HY, Lee EK, Choi YJ, Kim JM, Kim DH,
Zou Y, Kim CH, Lee J, Kim HS, Kim ND, et al: Molecular inflammation
as an underlying mechanism of the aging process and age-related
diseases. J Dent Res. 90:830–840. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Fluckiger A, Dumont A, Derangère V, Rébé
C, de Rosny C, Causse S, Thomas C, Apetoh L, Hichami A,
Ghiringhelli F, et al: Inhibition of colon cancer growth by
docosahexaenoic acid involves autocrine production of TNFα.
Oncogene. 35:4611–4622. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lee HY, Youn SW, Kim JY, Park KW, Hwang
CI, Park WY, Oh BH, Park YB, Walsh K, Seo JS, et al: FOXO3a turns
the tumor necrosis factor receptor signaling towards apoptosis
through reciprocal regulation of c-Jun N-terminal kinase and
NF-kappaB. Arterioscler Thromb Vasc Biol. 28:112–120. 2008.
View Article : Google Scholar
|
|
59
|
Dasgupta A, Nomura M, Shuck R and Yustein
J: Cancer's Achilles' Heel: Apoptosis and Necroptosis to the
Rescue. Int J Mol Sci. 18:1–20. 2016. View Article : Google Scholar
|
|
60
|
Shi K-G and Qing L-H: The follow-up
observation assessment of medium and late phases cancer treated by
Chinese Ganoderma lucidum essence (CGLE). Ganoderma: Genetics,
Chemistry, Pharmacology and Therapeutics - Proceedings of
International Symposium on Ganoderma Research; 2002, http://cstm.cnki.net/stmt/TitleBrowse/KnowledgeNet/ZGYS200210001028?db=STMi8515.
|
|
61
|
Zhao H, Zhang Q, Zhao L, Huang X, Wang J
and Kang X: Spore powder of Ganoderma lucidum improves
cancer-related fatigue in breast cancer patients undergoing
endocrine therapy: A pilot clinical trial. Evid Based Complement
Alternat Med. 2012:8096142012. View Article : Google Scholar
|
|
62
|
Suprasert P, Apichartpiyakul C, Sakonwasun
C, Nitisuwanraksa P and Phuackchantuck R: Clinical characteristics
of gynecologic cancer patients who respond to salvage treatment
with Lingzhi. Asian Pac J Cancer Prev. 15:4193–4196. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chen D, Yu J and Zhang L: Necroptosis: An
alternative cell death program defending against cancer. Biochimica
et Biophysica Acta. 1865:228–236. 2016.PubMed/NCBI
|
|
64
|
Feoktistova M, Wallberg F, Tenev T,
Geserick P, Leverkus M and Meier P: Techniques to distinguish
apoptosis from necroptosis. Cold Spring Harb Protoc.
2016:pdb.top070375. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zong WX and Thompson CB: Necrotic death as
a cell fate. Genes Dev. 20:1–15. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kanduc D, Mittelman A, Serpico R,
Sinigaglia E, Sinha AA, Natale C, Santacroce R, Di Corcia MG,
Lucchese A, Dini L, et al: Cell death: Apoptosis versus necrosis
(Review). Int J Oncol. 21:165–170. 2002.PubMed/NCBI
|
|
67
|
Diederich M and Cerella C: Non-canonical
programmed cell death mechanisms triggered by natural compounds.
Semin Cancer Biol. 40–41:4–34. 2016. View Article : Google Scholar
|
|
68
|
Gali-Muhtasib H, Hmadi R, Kareh M, Tohme R
and Darwiche N: Cell death mechanisms of plant-derived anticancer
drugs: Beyond apoptosis. Apoptosis. 20:1531–1562. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Wilson TR, Johnston PG and Longley DB:
Anti-apoptotic mechanisms of drug resistance in cancer. Curr Cancer
Drug Targets. 9:307–319. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kim MS, Lee SY, Cho WH, Song WS, Koh JS,
Lee JA, Yoo JY and Jeon DG: Tumor necrosis rate adjusted by tumor
volume change is a better predictor of survival of localized
osteosarcoma patients. Ann Surg Oncol. 15:906–914. 2008. View Article : Google Scholar
|
|
71
|
Fearon KC, Glass DJ and Guttridge DC:
Cancer cachexia: Mediators, signaling, and metabolic pathways. Cell
Metab. 16:153–166. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhuang SR, Chen SL, Tsai JH, Huang CC, Wu
TC, Liu WS, Tseng HC, Lee HS, Huang MC, Shane GT, et al: Effect of
citronellol and the Chinese medical herb complex on cellular
immunity of cancer patients receiving chemotherapy/radiotherapy.
Phytother Res. 23:785–790. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Jin X, Ruiz Beguerie J, Sze DM and Chan
GC: Ganoderma lucidum (Reishi mushroom) for cancer treatment.
Cochrane Database Syst Rev. 6:CD0077312012.
|
|
74
|
Khaled YS, Elkord E and Ammori BJ:
Macrophage inhibitory cytokine-1: A review of its pleiotropic
actions in cancer. Cancer Biomark. 11:183–190. 2012. View Article : Google Scholar : PubMed/NCBI
|