Introduction
Liver cancer is a malignant cancer (1), and radiotherapy is a common means of
prolonging the life of patients with middle- to late-stage cancer
(2). Radiogenic reactive oxygen
species (ROS) induce cancer cell death (3). The transcription factor nuclear
factor (erythroid-derived 2)-like 2 (Nrf2) is a master regulator of
oxidative and xenobiotic metabolism (4) that is sensitive to ROS (5). Nrf2 controls the transcription and
expression of ~1% of all human genes; Nrf2-regulated genes
participate in biotransformation reactions, redox status, energy
metabolism and proteostasis (6–9). The
expression products of Nrf2-targeted genes, such as antioxidant
enzymes, can relieve the oxidative stress generated by ionizing
radiation in cancer cells (10–12),
leading to resistance to cancer radiotherapy. Under normal
conditions, Nrf2 interacts with Kelch-like ECH-associated protein 1
(Keap1) in the cytoplasm (13).
KEAP1 interacts with two regions of NRF2 (amino-acid sequences DLG
and ETGE) located in the N-terminal Neh2 domain to direct
ubiquitination of NRF2 by the Cullin-3/Rbx1 complex and the
proteasomal degradation of NRF2 (14,15)
(Fig. 1). Upon oxidation of KEAP1,
NRF2 escapes degradation, enters the nucleus, and then binds to the
Maf protein, which targets antioxidant response element (ARE) genes
to initiate downstream antioxidant gene transcription (16–19).
For cancer radiotherapy, we aimed to inhibit the Nrf2-ARE signaling
pathway. We found that the Wnt inhibitor LGK-974 can hinder the
entrance of Nrf2 into the nucleus of HepG2 cells. Another report
indicated that NRF2 participates in the formation of an
Axin1/GSK-3/b-TrCP protein complex that is regulated by WNT-3A in
liver cells (20).
LGK-974 inhibits Wnt signaling pathways by blocking
secretion of the Wnt3A protein in HepG2 cells. The canonical Wnt
signaling pathway is initiated upon binding of secreted Wnt3A to
the receptors LRP5/6 and Frizzled, which are embedded in the cell
membrane (21,22). LRP5/6 has five contiguous PPPSPXS
motifs, and Axin can bind to a phosphorylated PPPSPXS motif, GSK-3β
and CK1 simultaneously (23,24).
The association between Axin and LRP5/6 ultimately contributes to
the degradation of Axin-GSK3β-CK1-APC protein complexes. Thus,
β-catenin avoids degradation by Axin-GSK3β-CK1-APC complexes. In
the cytoplasm, β-catenin accumulates and enters the nucleus to
activate different genes that regulate cellular proliferation and
differentiation (25).
LGK-974 inhibits porcupine (PORCN), an
O-acyltransferase in the endoplasmic reticulum required for the
secretion of Wnt, and therefore inhibits endogenous Wnt signaling
(26). In the presence of LGK-974,
Wnt3A palmitoylation does not occur, and the Wnt protein cannot
form its proper conformation (27–29).
Ultimately, Wnt3A becomes stranded in the cytoplasm. Given this
promising finding, we suspect that LGK-974 might enhance the
efficacy of radiotherapy by inhibiting Nrf2.
Materials and methods
Chemicals and reagents
High-glucose Dulbecco's modified Eagle's medium
(DMEM) and penicillin/streptomycin were purchased from HyClone
Laboratories, Inc., (Logan, UT, USA). Fetal bovine serum (FBS) was
obtained from Gibco-BRL (Grand Island, NY, USA). LGK-974 was
obtained from Selleck Chemicals (Houston, TX, USA). Trypsin,
DEPC-treated water, dimethyl sulfoxide (DMSO), RIPA lysis buffer,
Giemsa stain and other reagents were purchased from Beijing
Solarbio Life Sciences (Beijing, China). MTT and
H2DCF-DA were obtained from Sigma-Aldrich (St. Louis,
MO, USA). Primary antibodies including rabbit/mouse antibodies
against Wnt3A, Nrf2, β-catenin, APC-6, NQO-1, HO-1 and survivin
were purchased from Abcam (Cambridge, MA, USA), and the anti-Keap-1
antibody was purchased from Santa Cruz Biotechnology (Santa Cruz,
CA, USA). Horseradish peroxidase-conjugated anti-rabbit IgG and
anti-mouse IgG were obtained from ZSGB-Bio (Beijing, China).
Cell culture
Cells were cultured in high-glucose DMEM containing
10% FBS and 100 U/ml penicillin/streptomycin at 37°C in a
humidified CO2 incubator.
LGK-974 treatment and irradiation
The cells were divided into four groups: group a,
control; group b, LGK-974 alone; group c, irradiation alone; and
group d, LGK-974 + irradiation. The LGK-974 and LGK-974 +
irradiation groups were treated with 0.1, 1, 5 or 10 µM
LGK-974. After 24 h, the irradiation and LGK-974 + irradiation
groups were exposed to 2 Gy or 4 Gy doses of 137Cs γ-rays at a dose
rate of 1.02 Gy per minute in an exposure instrument (Atomic Energy
of Canada Ltd., Chalk River, ON, Canada). LGK-974 was dissolved in
the solvent DMSO according to the manufacturer's instructions. We
dissolved 5 mg of LGK-974 into 1.2612 ml of DMSO, obtaining a 10 mM
LGK-974 starting solution. The experimental concentrations, such as
0.1, 1, 5 and 10 µM, were generated by diluting the 10 mM
LGK-974 starting solution with culture medium.
HepG2 cell counts, growth curve and clone
formation assay
Cells were seeded in 12-well plates at a density of
3×104 cells/well in triplicate and then treated with the
indicated concentrations of LGK-974 and the indicated doses of
irradiation, followed by 6 days in culture. The medium was replaced
with the identical fresh media every 24 h, and the number of
surviving cells was counted daily using trypan blue and a
hemocytometer. HepG2 cells were plated at the same densities (100
cells/well) in 12-well plates in triplicate, treated with 1.0
µM LGK-974 for 24 h, and then exposed to the indicated doses
of 137Cs radiation (1.02 Gy/min). Two hours after irradiation, the
medium was replaced with fresh medium without any drug. The cells
were then incubated for ~8 days, with changes of medium every 2
days. Finally, the cells were subjected to Giemsa staining and
colonies containing >50 cells were counted.
Cell viability assay
HepG2 cell viability was determined using an MTT
assay, which involves the reduction of yellow MTT by mitochondrial
succinate dehydrogenase in viable cells. HepG2 cells were cultured
in a 96-well plate (5,000 cells/well) for 24 h. Then, the
experimental groups were starved in serum-free media overnight. A
series of concentrations of LGK-974 (0.1, 1 and 10 µM) were
added to each well for groups b and d, which were then cultured for
an additional 24 h. Group c and group d were exposed to 2 Gy or 4
Gy γ-irradiation and then incubated with 5 mg/ml MTT for 4 h in the
dark. The supernatant was removed, and the formazan crystals in
each well were dissolved in 150 µl of DMSO with shaking for
15 min at room temperature in the dark. The absorbance at 562 nm
was measured using a microplate reader. Cell viability was
expressed as a percentage relative to the vehicle-treated control
cells.
Flow cytometry for apoptosis assays and
detection of intracellular
ROS. HepG2 cells in the logarithmic growth
phase were treated with 0.1, 1 or 10 µM LGK-974 and exposed
to 2 Gy or 4 Gy γ-irradiation. Cells in both the control group and
the experimental groups were collected 12 h after the treatment.
HepG2 cells were prepared and evaluated according to the
instructions with the Annexin V-FITC/PI apoptosis detection kit.
Cell apoptosis assays were performed using flow cytometry (BD
Biosciences, San Jose, CA, USA). Intracellular ROS levels were
measured using H2DCFH-DA (Molecular Probes/Invitrogen,
Waltham, MA, USA). HepG2 cells were treated with or without 10
µM LGK-974 and 10 µl H2O2 6 h
prior to irradiation and then treated with 10 µM
H2DCFH-DA for 30 min. Next, the cells were washed with
phosphate-buffered saline (PBS), followed by trypsinization. After
detachment, the cells were collected, washed twice and resuspended
in 500 µl of PBS. We used a flow cytometer at
excitation/emission wavelengths of 488/525 nm to measure the
fluorescence.
Immunofluorescence
HepG2 cells (2.0×105 cells/well) were
cultured in 24-well plates on sterilized coverslips, treated with
or without 10 µM LGK-974/and exposed to 4 Gy. After 6 h,
these cells were washed with PBS and fixed with 4% paraformaldehyde
for 20 min at room temperature. Afterwards, the cells were washed 3
times with PBS for 5 min and permeabilized in 0.3% Triton X-100/PBS
for 15 min. Next, the cells were subjected to immunodetection. All
washes were performed 3 times with PBS for 5 min. The cells were
incubated with 10% goat serum/PBS for 30 min at room temperature
and then incubated with the polyclonal anti-Nrf2 antibody at
1:1,000 in 10% goat serum/PBS overnight at 4°C. Then, the cells
were washed and incubated with the FITC-conjugated secondary
anti-rabbit IgG antibody diluted 1:100 in 10% goat serum/PBS for 1
h. Finally, the cells were washed and incubated with 50 µl
of 0.5 µg/ml DAPI for 5 min at room temperature to stain the
nuclei. The fluorescence images were captured using a fluorescence
microscope.
Quantitative reverse-transcription
PCR
HepG2 cells were harvested, and total RNA was
isolated using TRIzol® reagent (Invitrogen/Life
Technologies). cDNA was reverse-transcribed with 5 µg of
total RNA using oligo(dT) primers and M-MLV reverse transcriptase
(Promega, Fitchburg, WI, USA). The primer sequences were as
follows: 5′-CCACCAGCAGCGAC TCTGA-3′ and 5′-GCAGAAGGTGATCCAGACTC-3′
as forward and reverse primers for C-myc, respectively; 5′-GTGG
CAGTGGCTCCATGTACTC-3′ and 5′-CTTGGAAGCCACAGAAATGCAG-3′ as forward
and reverse primers for NQO-1, respectively;
5′-TTGCCAGTGCCACCAAGTTC-3′ and 5′-TCAGCAGCTCCTGCAACTCC-3′ as
forward and reverse primers for HO-1, respectively; and
5′-CTGCACCACCAACTGCTTAG-3′ and 5′-AGGTCCACCACTGACACGTT-3′ as
forward and reverse primers for GAPDH, respectively.
Western blot analysis
Cancer cells were washed twice with ice-cold PBS (pH
7.4), harvested in cell lysis buffer at 4°C for 20–30 min and
centrifuged at 12,000 rpm at 4°C for 15 min to obtain total protein
lysates for immunoblot analysis. The protein concentrations were
quantified using the BCA protein assay reagent according to the
manufacturer's protocols. Equal amounts of protein were loaded onto
12% SDS-polyacrylamide gels for electrophoresis; then, the proteins
were transferred onto a PVDF membrane (Bio-Rad Laboratories,
Hercules, CA, USA) for 1 h using a semi-dry transfer system
(Bio-Rad Laboratories). The membrane was blocked with 5% non-fat
milk in PBST buffer (0.1% Tween-20 in PBS) for 2 h at room
temperature and then incubated with the appropriate primary
antibodies overnight. After hybridization with a primary antibody,
the membrane was washed three times with TBST and then incubated
with anti-mouse or anti-rabbit IgG horseradish
peroxidase-conjugated secondary antibodies for 50 min at room
temperature, followed by three washes with TBST. Finally, the
immunoreactive bands were visualized using enhanced
chemiluminescence reagents.
Single-cell gel assay (comet assay)
After exposure to 4 Gy irradiation, cells were
digested and collected immediately, washed twice with PBS, and
suspended in PBS at a concentration of 4×105 cells/ml.
The comet slides were coated with 0.75% normal-melting-point
agarose/PBS. Once the first layer of normal-melting-point agarose
was coagulated, a mixture of 70 µl low-melting-point
agarose/PBS and 30 µl of cell suspension was applied as the
second layer. The comet slides were then immersed in cold fresh
lysis solution (2.5 M NaCl, 10 mM Tris base, 1% N-sodium lauryl
sarcosinate, 30 mM Na2EDTA, 10% DMSO, 1% Triton X-100)
for 2.5 h at 4°C. Next, the comet slides were immersed in TBE
buffer for 20 min in a horizontal electrophoresis tank and
electrophoresis was performed at 30 V for 20 min in TBE buffer,
followed by neutralization for 20 min. The slides were then rinsed
twice with PBS and stained with ethidium bromide. Finally, the
comet slides were viewed under a fluorescence microscope, and data
were collected using a digital imaging system and analyzed with
CASP software (CaspLab, Wroclaw, Poland).
Statistical analysis
Each experiment was performed at least 3 times, and
the data are expressed as the means ± standard deviation (SD). For
comparisons between three or more groups, one-way analysis of
variance was used, followed by Tukey's multiple comparison tests.
Statistical analysis was performed using the GraphPad Prism
software, version 5.0 (GraphPad Software, Inc., San Diego, CA,
USA).
Results
Comparison of the radiosensitivity of
three human hepatic carcinoma cell lines
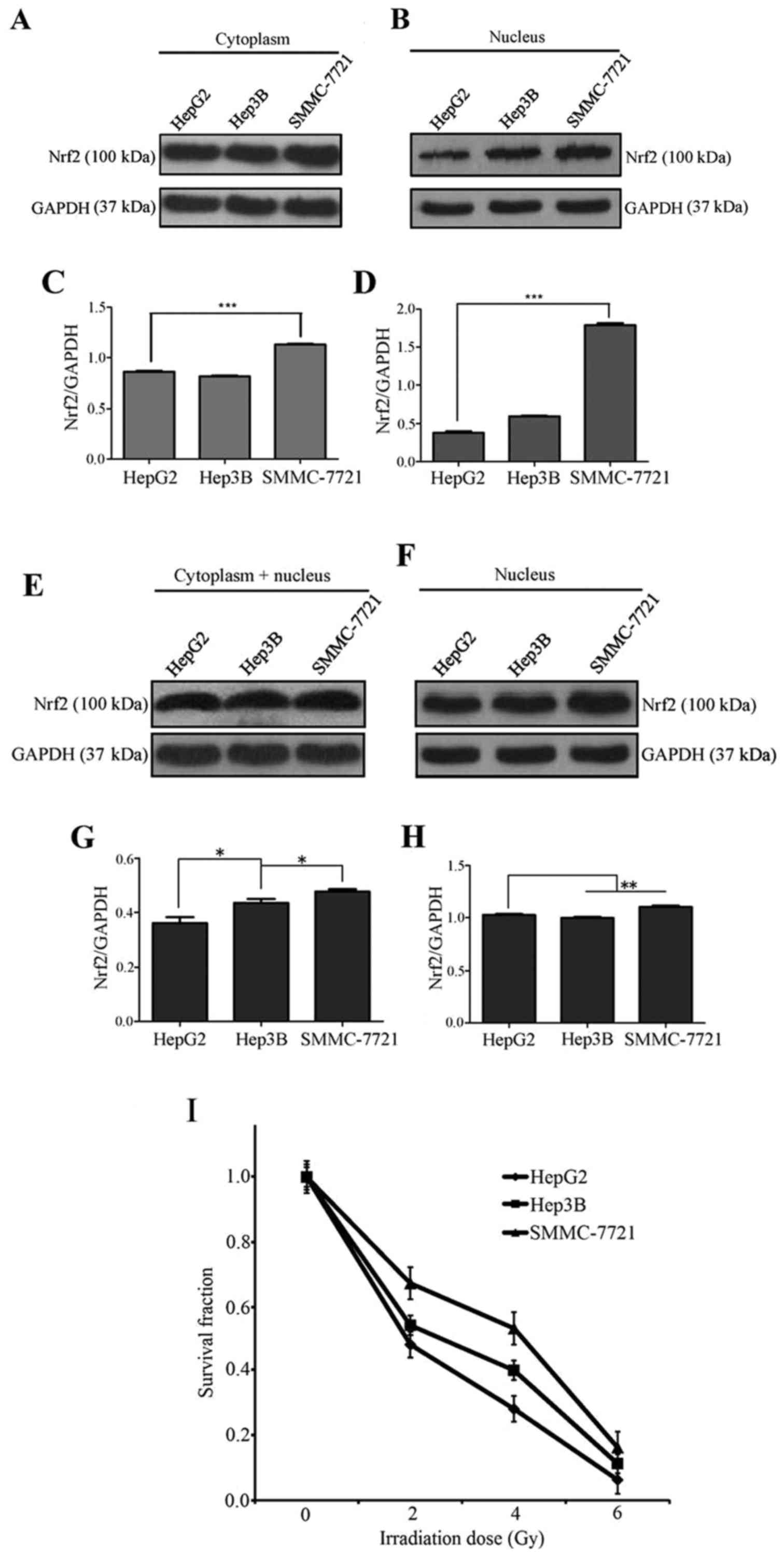
To evaluate the Nrf2 protein levels between three
liver cell lines, we performed western blot assays on nuclear and
cytoplasmic proteins. To test the effects of radiation on each cell
line, we performed
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assays. The Nrf2 levels in total protein samples were not notably
different between the three hepatoma cell types under normal
culture conditions, although SMMC-7721 cells had the highest Nrf2
protein levels (Fig. 1A and C).
However, in the nucleus, the Nrf2 level in HepG2 cells was the
lowest of the three cell lines, followed by Hep3B cells and then
SMMC-7721 cells (Fig. 1B and D).
This result showed that Nrf2 is primarily localized to the
cytoplasm in HepG2 cells, is equally distributed between the
cytoplasm and nucleus in Hep3B cells, and is predominantly
localized to the nucleus in SMMC-7721 cells. To test the effects of
irradiation on these three cell lines, we exposed HepG2, Hep3B and
SMMC-7721 cells to 2 Gy irradiation. Then, western blot assays were
performed to examine the Nrf2 content. The results showed that Nrf2
expression was increased in both total protein samples and nuclear
fractions (Fig. 1E and F); in
addition, the level of Nrf2 in the nucleus of HepG2 cells was added
by the greatest extent among the three cell lines (Fig. 1F and H). Based on the results
following exposure of cells to 0, 2, 4 or 6 Gy irradiation, HepG2
and Hep3B cells were clearly more sensitive to irradiation than
SMMC-7721 cells (Fig. 1I). The
above results show that SMMC-7721 is not sensitive to radiation and
that the antioxidative transcription factor Nrf2 is primarily
localized to the nucleus in SMMC-7721 cells. The radioresistant
effect of Nrf2 is primarily attributable to its function in the
nucleus. It is very likely that nuclear Nrf2 plays a role in the
protection against radiation once this protein sufficiently
accumulates in the nucleus. Irradiation had the greatest influence
on Nrf2 distribution in HepG2 cells and it can push Nrf2 into HepG2
cell nucleus. To establish whether the Wnt3A inhibitor LGK-974 can
prevent the entry of Nrf2 into the nucleus and to clearly
distinguish cytoplasmic Nrf2 from nuclear Nrf2, we selected HepG2
cells as the model for our further research.
LGK-974 increases the radiosensitivity of
HepG2 cells
To suppress interference caused by the solvent DMSO,
we performed growth curve assays and MTT assays on HepG2 cells
treated with LGK-974 solution or its solvent DMSO. We found that
DMSO had little impact on the cells. The results were comparable
between the untreated control and DMSO groups, and both showed
differences compared with the LGK-974 group (Fig. 2A–C). To clarify the
radiosensitizing effect of LGK-974 on HepG2 cells, we obtained cell
growth curves by performing MTT assays and apoptosis assays via
flow cytometry. To observe radiosensitivity of cells treated with
LGK-974 visually, we performed a clone formation assay. The results
of these experiments showed that LGK-974 increased the sensitivity
of HepG2 cells to irradiation. HepG2 cells were treated with
LGK-974 alone, ionizing radiation alone, or both the drug and the
radiation. We counted the number of cells before the treatment.
After 2 Gy, 4 Gy, 6 Gy or 8 Gy irradiation, the number of viable
cells decreased substantially, and the proliferation rate decreased
as the irradiation dose increased (Fig. 2D). LGK-974 suppresses HepG2 cell
growth and exerts cytotoxic effects (Fig. 2E). The cells grew until five days
after LGK-974 treatment; thereafter, the cell growth rate
decreased. We found that LGK-974 increased the radiosensitivity of
HepG2 cells. The cells treated with 10 µM LGK-974 and
exposed to 4 Gy irradiation grew the most slowly of all tested
cells (Fig. 2F). The results of
the clone formation assay showed that LGK-974 enhanced the
radiosensitivity of HepG2 cells (Fig.
2I and J). In apoptosis assays conducted via flow cytometry,
the HepG2 cells in the experimental groups were cultured in 1 or 10
µM LGK-974. After 24 h, the exposure groups received 4 Gy
irradiation; 4 h later, the cells were collected for apoptosis
assays. The HepG2 cells in the combined irradiation and LGK-974
group exhibited the highest apoptosis rate (Fig. 2G and H). In MTT assays, after
culturing the cells in different concentrations of LGK-974 for 24
h, we determined the proportion of surviving cells. The highest
rates of cell death occurred in HepG2 cells treated with 10 or 50
µM LGK-974. However, the proportion of surviving cells was
comparable between these two groups. The survival rate of the 50
µM LGK-974 group was very low, and the number of cells was
too low to perform any additional experiments. Given the above
results, we chose 10 µM LGK-974 for subsequent
experiments.
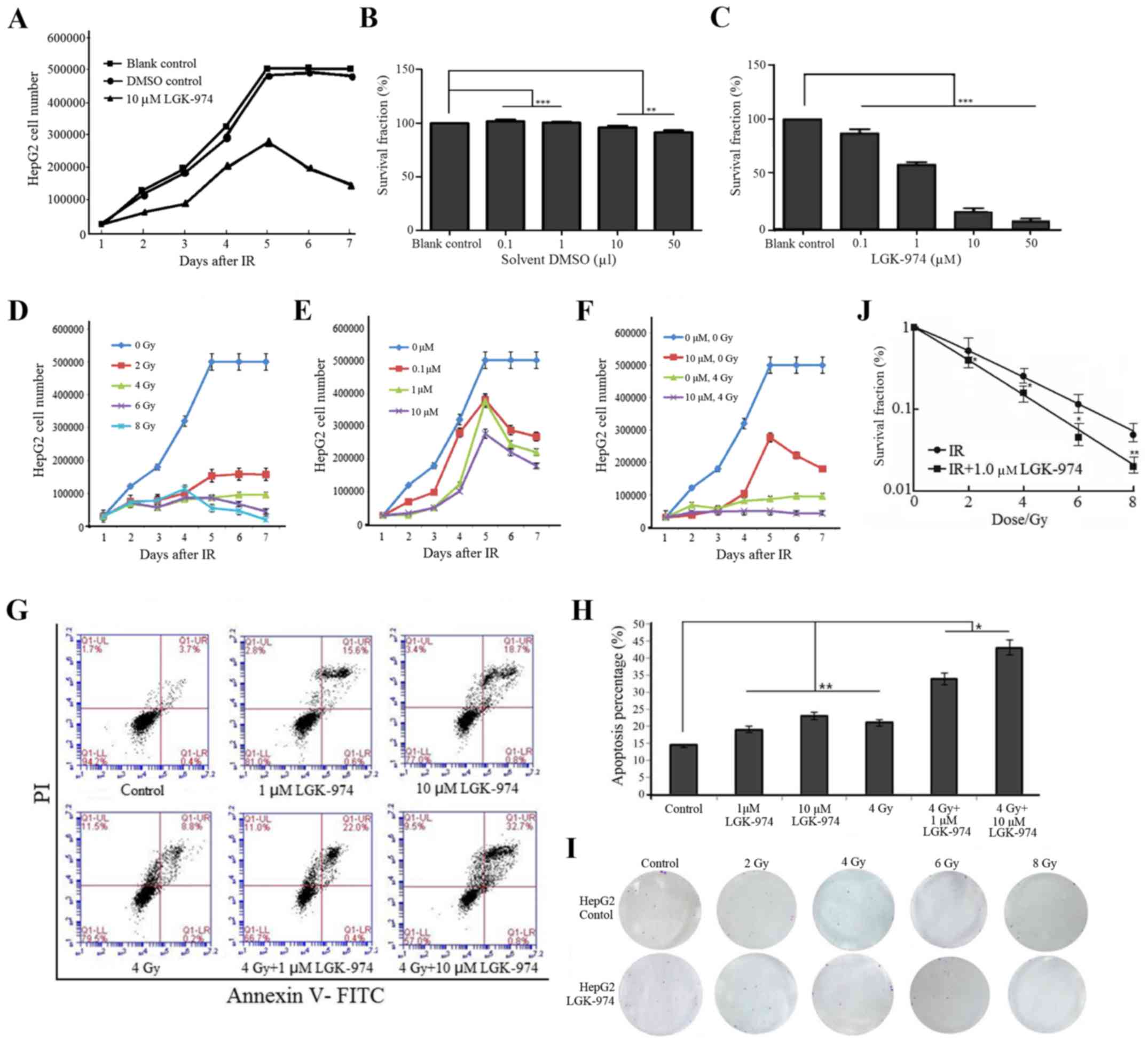 | Figure 2The Wnt3A inhibitor LGK-974 increases
the radiosensitivity of HepG2 cells. (A) Growth rate of HepG2 cells
treated with LGK-974 solution or its solvent DMSO. The number of
HepG2 cells was counted at 7 days. The results were identical
between the solvent DMSO group and the blank control group, and
HepG2 cell proliferation and survival rates decreased as the
LGK-974 concentration increased. (B and C) HepG2 cells were
incubated with different concentrations of LGK-974 or a
corresponding amount of solvent DMSO for 48 h; then, MTT assays
were performed. Survival rate of HepG2 cells under different
concentrations. The cell survival rate changed so little as the
DMSO concentration increased. The cell survival rate decreased as
the LGK-974 concentration increased. In the presence of 50
µM LGK-974, nearly all cells died. (D and E) Growth rate of
HepG2 cells after exposure to different doses of irradiation and
different concentrations of LGK-974. Radiation had a strong
influence on cell growth. Four days after 6 Gy or 8 Gy irradiation,
all HepG2 cells gradually died. Most cells survived after 2 Gy or 4
Gy irradiation. (E) HepG2 cell proliferation and survival
capability decreased as the LGK-974 concentration increased. The
cells gradually died after the fifth day. The HepG2 cells received
10 µM LGK-974 had the lowest growth rate and the maximum
mortality. (F) Growth rate of HepG2 cells after exposure to 4 Gy or
10 µM LGK-974. The cells without any processing had the
highest growth rate. The cells that received both 10 µM
LGK-974 and 4 Gy had the lowest growth rate. (G and H) Cell
apoptosis after different treatments. The cells in the LGK-974 and
4 Gy radiation treatment group exhibited the highest apoptosis
rate. (I and J) HepG2 cells were cultured in 12-well culture plates
at 100 cells/well, treated with LGK-974 solution for 24 h, and then
exposed to 2, 4, 6, or 8 Gy irradiation. At 8 days, the results
were analyzed. ***P<0.001, **P<0.01,
*P<0.05. |
LGK-974 inhibits the activation of Nrf2
in HepG2 cells after exposure to irradiation
To demonstrate the effects of LGK-974, we performed
western blot assays and RT-PCR assays. HepG2 cells were cultured in
different concentrations of LGK-974. After 24 h, we extracted total
protein samples for western blot assays (Fig. 3A). The expression of Nrf2 and its
downstream genes HO-1 and NQO-1 clearly decreased as the LGK-974
concentration increased (Fig. 3A, B, D
and E). However, the levels of Wnt3A and survivin increased
with increasing LGK-974 concentrations (Fig. 3A and C). This phenomenon explained
how LGK-974 could restrict Wnt3A to the cytoplasm, thus, preventing
the secretion of large amounts of Wnt3A from HepG2 cells. After the
cells were exposed to 4 Gy irradiation, the Wnt3A level declined.
LGK-974 treatment caused a gradual increase in the Wnt3A content
(Fig. 3G and J). This finding
confirmed the above hypotheses. The decrease in survivin expression
demonstrated the cytotoxicity of LGK-974 (Fig. 3A and F). To evaluate the effects of
LGK-974 and irradiation on HepG2 cells, we treated the cells with
different concentrations of LGK-974 for 24 h. Then, the cells were
exposed to 4 Gy radiation. After 8 h, we extracted total protein
samples for western blot assays (Fig.
3G). After 4 Gy irradiation, the Nrf2 level increased,
indicating that irradiation can increase oxidative stress in cells.
To handle this stress, oxidative activity and protein synthesis of
the transcription factor Nrf2 protein increased. However, the Nrf2
concentration decreased in the presence of LGK-974 (Fig. 3G and H). In cells treated with 1
µM LGK-974, the Nrf2 concentration was <0.1 µM;
however, this concentration remained lower than the Nrf2
concentrations in the control cells and the radiation-exposed
cells. The content of Nrf2 protein in HepG2 cells clearly
decreased. However, at low concentrations of LGK-974, this trend
was not apparent, or there was interference caused by other factors
in these cells. The NQO-1 level was strongly affected by
irradiation (Fig. 3A and I). The
level of β-catenin after ionizing radiation and treatment with 0.1
or 1 µM LGK-974 tended to be increased but was decreased by
treatment with 5 or 10 µM LGK-974 (Fig. 3A and K). This result verified that
ionizing radiation promotes the secretion of Wnt3A and that Wnt3A
stimulated the canonical Wnt pathway. As the LGK-974 concentration
increased, the canonical Wnt pathway was further inhibited. The
level of β-catenin decreased gradually with increasing LGK-974
concentrations. To reveal the effect of the solvent DMSO on the
experimental results, we performed western blot assays on cells
treated with DMSO alone. The results demonstrated that DMSO had
nearly no effect on Wnt3A and Nrf2 expression, and the results
between the blank control and DMSO control groups were identical
(Fig. 3L–N). Using DMSO as a
control, we also obtained similar results that LGK-974 decreased
Nrf2 levels and inhibited NQO-1 and HO-1 expression (Fig. 3O–R). The mRNA levels of HO-1 and
NQO-1 increased substantially after 4 Gy irradiation and then
decreased due to LGK-974 treatment (Fig. 3S). This result demonstrated that
irradiation can activate the Nrf2 pathway and that LGK-974 can
inhibit the Nrf2 pathway. The trends in the C-myc and survivin mRNA
levels were identical to those for the HO-1 and NQO-1 proteins
(Fig. 3T). Irradiation induces the
translocation of Nrf2 into the cell nucleus, but LGK-974 weakens
this phenomenon and inhibits Nrf2 expression (Fig. 3U). This result demonstrated that
irradiation can activate the Wnt pathway and that LGK-974 can
inhibit the Wnt pathway.
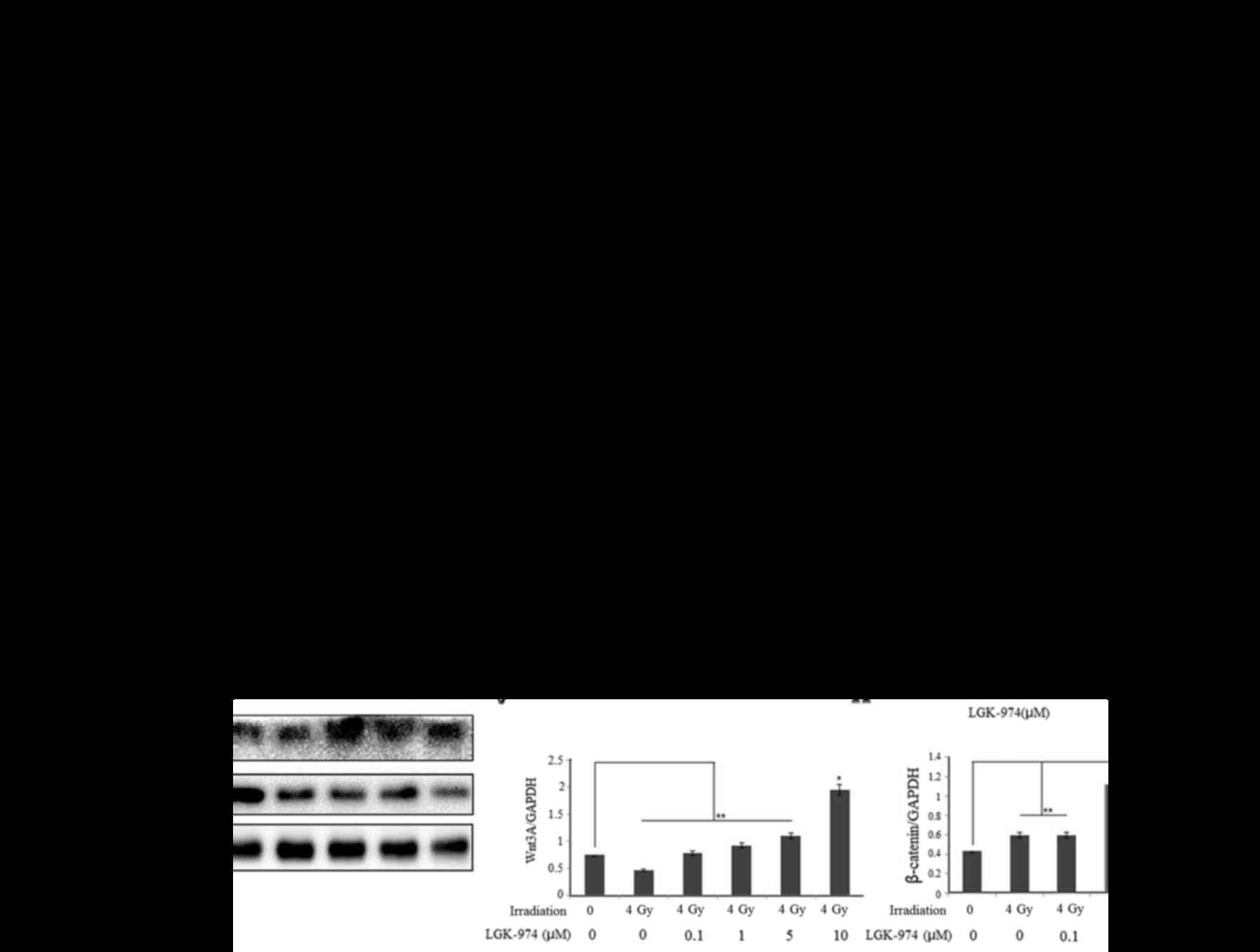 | Figure 3The Wnt3A inhibitor LGK-974 can
modulate the Nrf2 signaling pathway. (A) HepG2 cells were treated
with LGK-974 at the indicated concentrations and collected after 24
h. Then, we extracted total protein samples for western blot
analysis. (B–F) Relative levels of Nrf2, Wnt 3A, HO-1, NQO-1 and
survivin in HepG2 cells after treatment with different
concentrations of LGK-974. (G) HepG2 cells were treated with
different concentrations of LGK-974. After 20 h, the cells were
exposed to 4 Gy radiation. After 8 h, we collected the cells for
western blot analysis. (H–K) Nrf2, NQO-1, Wnt3A and β-catenin
levels in HepG2 cells under the different treatments described
above. (L) HepG2 cells were treated with 10 µM LGK-974 or
solvent DMSO alone. After 24 h, the cells were collected and
subjected to western blot analysis. (M and N) Nrf2 and Wnt3A levels
in HepG2 cells treated with 10 µM LGK-974 or solvent DMSO
alone. (O) HepG2 cells were treated with 10 µM LGK-974 or
exposed to 4 Gy radiation. After 24 h, we collected the cells for
western blot analysis. (P–R) Nrf2, HO-1 and NQO-1 levels in HepG2
cells under the different treatments as shown in (O). (S and T)
Relative mRNA levels of Nrf2, NQO-1, Wnt3A and β-catenin in HepG2
cells. HepG2 cells were treated with different concentrations of
LGK-974. After 12 h, the cells were exposed to 4 Gy radiation.
After an additional 6 h, we collected the cells for total RNA
extraction and PCR assays. (U) The Nrf2 levels in different treated
HepG2 cells. HepG2 cells were treated with or without 10 µM
LGK-974 for 24 h; then, the cells were exposed to 4 Gy irradiation.
After an additional 6 h, immunofluorescence analysis was
performed. |
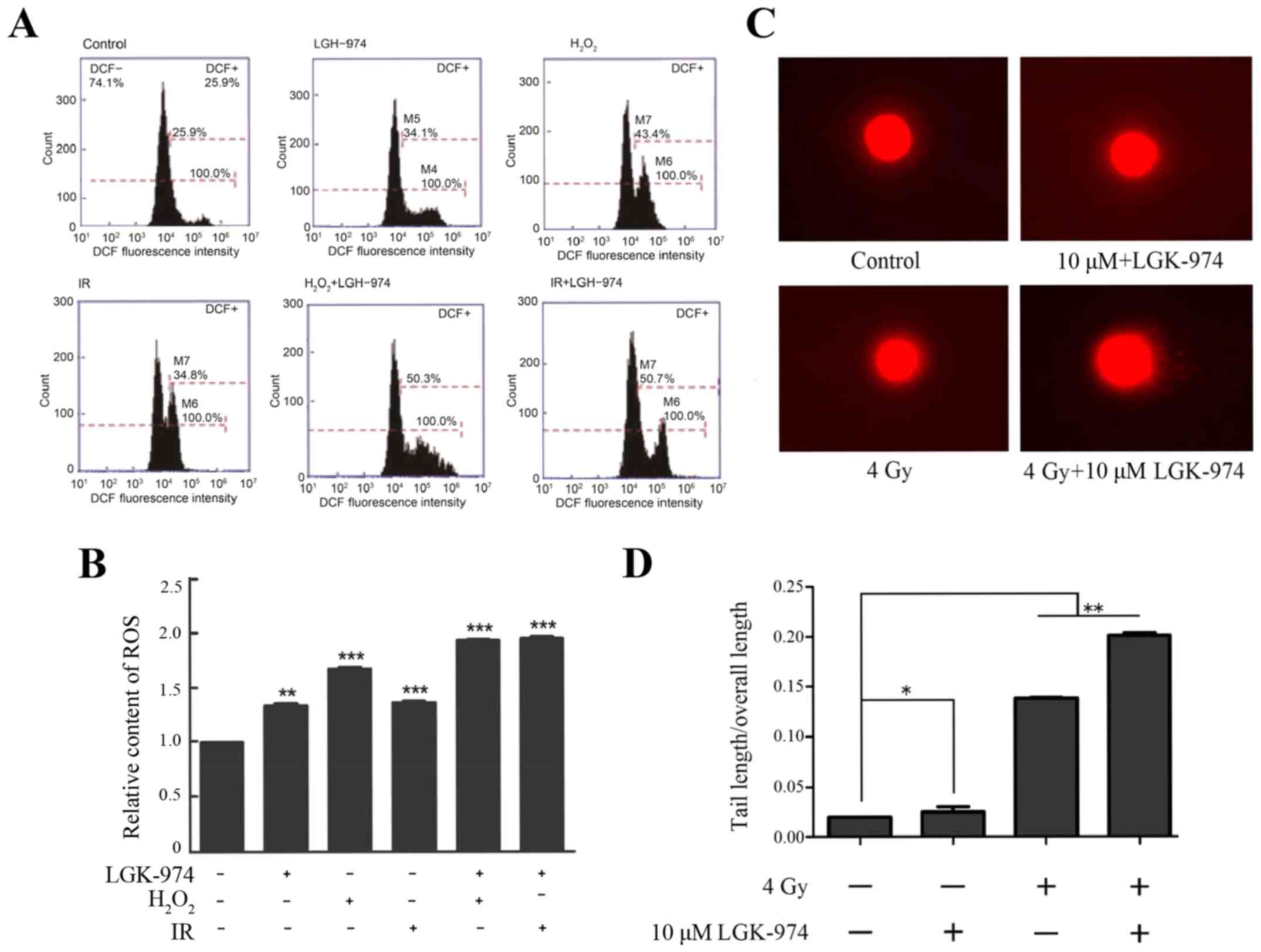
LGK-974 treatment increases the
radiosensitivity of HepG2 cells by increasing the level of ROS
To determine whether the decrease in Nrf2 protein
expression induced by LGK-974 contributes to increased ROS
accumulation in HepG2 cells, we measured the intracellular ROS
levels after ionizing radiation using the fluorescence indicator
2,7-dichlorodihydrofluorescein diacetate (H2DCFH-DA). We
chose H2O2 treatment as a positive control.
The oxidative effect of H2O2 is well known
(30). Irradiation increased the
ROS levels in HepG2 cells comparably to H2O2.
We found that the treatment of HepG2 cells with both 10 µM
LGK-974 and 4 Gy ionizing radiation clearly increased the levels of
ROS after 6 h. Fluorimetric quantification of the change in ROS
levels showed that the combination of ionizing radiation and
LGK-974 treatment caused an ~2.2-fold increase in the level of ROS
relative to the control treatment and a 1.57-fold increase relative
to ionizing radiation alone. Additionally, LGK-974 alone affected
ROS generation (Fig. 4A and B).
The increase in ROS levels in the groups treated with ionizing
radiation alone or with H2O2 alone
demonstrated that irradiation can increase oxidative stress in
cells. To establish whether irradiation or LGK-974 can damage
chromosomes, in addition to describing the effect of LGK-974 on
DNA, we performed comet assays. HepG2 cells were incubated with or
without 10 µM LGK-974 for 24 h and then exposed to 4 Gy
irradiation. The results demonstrated that irradiation damaged the
DNA of HepG2 cells and that additional treatment with LGK-974
exacerbated this phenomenon (Fig.
4C,D). In summary, the above results showed that LGK-974 can
amplify the effects of radiation on HepG2 cells.
Discussion
Liver cancer has high malignancy and an insidious
onset, is invasive and fast-growing, has high recurrence and
mortality rates and is more commonly found in men (31,32).
Radiation therapy is an effective treatment method to extend the
life of patients with liver cancer (33,34).
A major challenge in the attempt to cure cancer is the
radioresistance that is acquired during radiotherapy. Nrf2, a major
regulator of redox homeostasis, has previously been confirmed to
play an important role in the resistance to oxidative stress during
radiation therapy (35).
Inhibiting Nrf2 may potentiate cancer radiotherapy (36). Regarding the transcription factor
Nrf2, most studies have focused on the role of the electrophile and
redox sensor KEAP1 in modulating NRF2 protein levels to affect cell
functions and to exert antioxidative effects. In the present study,
we did not examine the Keap1-Nrf2 pathway and instead explored the
novel relationship between Wnt and Nrf2. We discovered that
LGK-974, a Wnt inhibitor, increases radiosensitivity inhibits Nrf2
signaling. Therefore, Wnt-Nrf2 may represent a novel signaling
pathway.
Recently, it has been found that the Wnt signaling
pathway and Nrf2 are associated, both in vivo and in
vitro (37). The downstream
protein complexes of Wnt and Axin1-GSK-3β interact with Nrf2, thus
holding Nrf2 in the cytoplasm. Under these conditions, the
transcription factor Nrf2 cannot enter the cell nucleus; instead,
NRF2 becomes phosphorylated, leading to b-TrCP-dependent
ubiquitination and proteasomal degradation. In this situation,
antioxidant genes, such as HO-1 and NQO-1, cannot be transcribed.
In hepatoma cells, the Wnt classical pathway is initiated when
secreted Wnt3A interacts with LRP5/6 and Frizzled embedded in the
cell membrane (38). LGK-974
limits the activity of PORCN, which is located in the endoplasmic
reticulum and catalyzes Wnt3A protein palmitoylation and secretion
(39). β-catenin is a downstream
cytoplasmic protein in the Wnt pathway. In the case of Wnt
signaling activation, Wnt escapes degradation, accumulates in the
cytoplasm and finally enters the nucleus to induce the
transcription of genes such as C-myc. We summarized the research
findings and utilized the Wnt inhibitor LGK-974 in liver cancer
radiotherapy. Preliminarily, we confirmed that LGK-974 has
radiosensitizing properties.
As a Wnt inhibitor, LGK-974 has been investigated in
several types of cancer. In pancreatic cancer cells, as an
inspection control, LGK-974 inhibited cell proliferation and
induced cell differentiation in RNF43-mutant pancreatic
adenocarcinoma xenograft models (40). In breast cancer cells, blocking the
autocrine Wnt signaling pathway using LGK-974 ameliorated NRBP1
overexpression-induced inhibition of cell proliferation (41). LGK974 decreased tumor cell growth
and metastasis in 8 head and neck squamous cell carcinoma cell
lines (42). LGK-974 also
inhibited the proliferation of mucin-producing intrahepatic
cholangiocarcinoma and mixed hepatocellular-cholangiocarcinoma
(43). All these findings show
that LGK-974 can inhibit cancer cell growth and metastasis;
however, no previous study has evaluated the effect of LGK-974 on
radiosensitivity.
In the present study, a liver cancer cell line was
cultured in vitro, and we used the Wnt inhibitor LGK-974
dissolved in DMSO in combination with ionizing radiation to study
the possibility that this drug might induce sensitization to
radiation. The amount of DMSO applied was very low. As a solvent,
DMSO has low toxicity (44), and
its effects on cells are far weaker than those of LGK-974. To more
clearly demonstrate the effect of LGK-974 on HepG2 cells, we
compared the experimental groups with a group of HepG2 cells
cultured under normal conditions. Our results showed that LGK-974
significantly inhibited the growth of HepG2 cells and promoted cell
apoptosis. LGK-974 inhibited C-myc gene expression downstream of
Wnt3A, NQO-1 gene expression downstream of Nrf2, and HO-1 gene and
protein expression. Analysis of the ROS levels in HepG2 cells
revealed that the antioxidant capacity of these cells decreased in
the presence of LGK-974.
At present, our studies were performed only at the
cellular level. In addition, only one cell line was used for
mechanistic studies. Further experiments should be performed on
more cell lines, animal models and subsequently, humans. To further
elucidate the mechanism of action of LGK-974, we will perform
additional assays, as the findings from the present study were not
comprehensive. Although we performed flow cytometry to examine cell
apoptosis, this single assay is inadequate. We will perform TUNEL
staining to obtain images of apoptotic cells in future research.
LGK-974 is an oral medicine that has already been accepted for
phase I clinical trials, although only as a tumor chemotherapy
drug, not for applications involving radiotherapy. This inhibitor
of Wnt can also block Nrf2 function in HepG2 cells, suggesting that
Wnt-Nrf2 may be a novel pathway related to radiation oncology. The
discovery of the radiosensitizing effect of LGK-974 has great
value. Once applied in the clinic, this treatment may be beneficial
to liver cancer patients.
LGK-974 is an effective inhibitor of the canonical
Wnt signaling pathway. We demonstrated that LGK-974 can inhibit
Nrf2 signaling by limiting the entry of Nrf2 into the nucleus.
LGK-974 has cytotoxicity and can increase the radiosensitivity of
HepG2 cells. These findings provide new ideas for novel potential
radiosensitizing drugs in cancer radiotherapy.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (nos. 31670859 and 81470098);
the Natural Science Foundation of Tianjin (no. 15KPXM01SF056); the
Fundamental Research Funds from CAMS and PUMC (no. 2016ZX310068);
and the Research Funds for the Innovation Team of IRM-CAMS (no.
1650).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Teltscharov L, Vlachov K and Marovski T:
Effect of radiotherapy on the liver function of cancer patients.
Radiobiol Radiother (Berl). 5:673–677. 1964.In German.
|
|
3
|
He L, Lai H and Chen T: Dual-function
nanosystem for synergetic cancer chemo-/radiotherapy through
ROS-mediated signaling pathways. Biomaterials. 51:30–42. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
DeNicola GM, Karreth FA, Humpton TJ,
Gopinathan A, Wei C, Frese K, Mangal D, Yu KH, Yeo CJ, Calhoun ES,
et al: Oncogene-induced Nrf2 transcription promotes ROS
detoxification and tumorigenesis. Nature. 475:106–109. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Merchant AA, Singh A, Matsui W and Biswal
S: The redox-sensitive transcription factor Nrf2 regulates murine
hematopoietic stem cell survival independently of ROS levels.
Blood. 118:6572–6579. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hayes JD and Dinkova-Kostova AT: The Nrf2
regulatory network provides an interface between redox and
intermediary metabolism. Trends Biochem Sci. 39:199–218. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kitteringham NR, Abdullah A, Walsh J,
Randle L, Jenkins RE, Sison R, Goldring CE, Powell H, Sanderson C,
Williams S, et al: Proteomic analysis of Nrf2 deficient transgenic
mice reveals cellular defence and lipid metabolism as primary
Nrf2-dependent pathways in the liver. J Proteomics. 73:1612–1631.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mitsuishi Y, Taguchi K, Kawatani Y,
Shibata T, Nukiwa T, Aburatani H, Yamamoto M and Motohashi H: Nrf2
redirects glucose and glutamine into anabolic pathways in metabolic
reprogramming. Cancer Cell. 22:66–79. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu KC, Cui JY and Klaassen CD: Effect of
graded Nrf2 activation on phase-I and -II drug metabolizing enzymes
and transporters in mouse liver. PLoS One. 7:e390062012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Saw CL, Wu Q and Kong AN: Anti-cancer and
potential chemo-preventive actions of ginseng by activating Nrf2
(NFE2L2) anti-oxidative stress/anti-inflammatory pathways. Chin
Med. 5:372010. View Article : Google Scholar
|
|
11
|
Seng S, Avraham HK, Jiang S, Yang S,
Sekine M, Kimelman N, Li H and Avraham S: The nuclear matrix
protein, NRP/B, enhances Nrf2-mediated oxidative stress responses
in breast cancer cells. Cancer Res. 67:8596–8604. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hong CC, Ambrosone CB, Ahn J, Choi JY,
McCullough ML, Stevens VL, Rodriguez C, Thun MJ and Calle EE:
Genetic variability in iron-related oxidative stress pathways
(Nrf2, NQ01, NOS3, and HO-1), iron intake, and risk of
postmenopausal breast cancer. Cancer Epidemiol Biomarkers Prev.
16:1784–1794. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zipper LM and Mulcahy RT: The Keap1
BTB/POZ dimerization function is required to sequester Nrf2 in
cytoplasm. J Biol Chem. 277:36544–36552. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nasiri HR, Linge S and Ullmann D:
Thermodynamic profiling of inhibitors of Nrf2:Keap1 interactions.
Bioorg Med Chem Lett. 26:526–529. 2016. View Article : Google Scholar
|
|
15
|
Leinonen HM, Kansanen E, Pölönen P,
Heinäniemi M and Levonen AL: Dysregulation of the Keap1-Nrf2
pathway in cancer. Biochem Soc Trans. 43:645–649. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
McMahon M, Thomas N, Itoh K, Yamamoto M
and Hayes JD: Dimerization of substrate adaptors can facilitate
cullin-mediated ubiquitylation of proteins by a 'tethering'
mechanism: A two-site interaction model for the Nrf2-Keap1 complex.
J Biol Chem. 281:24756–24768. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tong KI, Kobayashi A, Katsuoka F and
Yamamoto M: Two-site substrate recognition model for the Keap1-Nrf2
system: A hinge and latch mechanism. Biol Chem. 387:1311–1320.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang C, Li C, Peng H, Ye Z, Zhang J, Liu X
and Lou T: Activation of the Nrf2-ARE pathway attenuates
hyperglycemia-mediated injuries in mouse podocytes. Cell Physiol
Biochem. 34:891–902. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee LY, Köhler UA, Zhang L, Roenneburg D,
Werner S, Johnson JA and Foley DP: Activation of the Nrf2-ARE
pathway in hepatocytes protects against steatosis in nutritionally
induced non-alcoholic steatohepatitis in mice. Toxicol Sci.
142:361–374. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rada P, Rojo AI, Offergeld A, Feng GJ,
Velasco-Martín JP, González-Sancho JM, Valverde ÁM, Dale T,
Regadera J and Cuadrado A: WNT-3A regulates an Axin1/NRF2 complex
that regulates antioxidant metabolism in hepatocytes. Antioxid
Redox Signal. 22:555–571. 2015. View Article : Google Scholar :
|
|
21
|
MacDonald BT, Semenov MV, Huang H and He
X: Dissecting molecular differences between Wnt coreceptors LRP5
and LRP6. PLoS One. 6:e235372011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mi K and Johnson GV: Role of the
intracellular domains of LRP5 and LRP6 in activating the Wnt
canonical pathway. J Cell Biochem. 95:328–338. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zeng X, Tamai K, Doble B, Li S, Huang H,
Habas R, Okamura H, Woodgett J and He X: A dual-kinase mechanism
for Wnt co-receptor phosphorylation and activation. Nature.
438:873–877. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
MacDonald BT, Yokota C, Tamai K, Zeng X
and He X: Wnt signal amplification via activity, cooperativity, and
regulation of multiple intracellular PPPSP motifs in the Wnt
co-receptor LRP6. J Biol Chem. 283:16115–16123. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim M, Kim S, Lee SH, Kim W, Sohn MJ, Kim
HS, Kim J and Jho EH: Merlin inhibits Wnt/β-catenin signaling by
blocking LRP6 phosphorylation. Cell Death Differ. 23:1638–1647.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Morgan JT, Raghunathan VK, Chang YR,
Murphy CJ and Russell P: Wnt inhibition induces persistent
increases in intrinsic stiffness of human trabecular meshwork
cells. Exp Eye Res. 132:174–178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Abrami L, Kunz B, Iacovache I and van der
Goot FG: Palmitoylation and ubiquitination regulate exit of the Wnt
signaling protein LRP6 from the endoplasmic reticulum. Proc Natl
Acad Sci USA. 105:5384–5389. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Komekado H, Yamamoto H, Chiba T and
Kikuchi A: Glycosylation and palmitoylation of Wnt-3a are coupled
to produce an active form of Wnt-3a. Genes Cells. 12:521–534. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kurayoshi M, Yamamoto H, Izumi S and
Kikuchi A: Post-translational palmitoylation and glycosylation of
Wnt-5a are necessary for its signalling. Biochem J. 402:515–523.
2007. View Article : Google Scholar :
|
|
30
|
Rhee SG: Redox signaling: Hydrogen
peroxide as intracellular messenger. Exp Mol Med. 31:53–59. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics for Hispanics/Latinos, 2012. CA Cancer J Clin.
62:283–298. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hosoda Y, Kim Y, Nishino M, Okano M, Nagai
K, Yasui M and Tsujinaka T: A case of rectal cancer successfully
treated with stereotactic radiotherapy for liver and lung
metastases. Gan To Kagaku Ryoho. 42:2109–2111. 2015.In
Japanese.
|
|
34
|
Tono T, Hashimoto K, Yamada Y, Nishida K,
Yanagawa T, Danno K, Fujie Y, Fujita S, Fujita J, Yoshida T, et al:
Efficacy of stereotactic radiotherapy for primary and metastatic
liver cancer. Gan To Kagaku Ryoho. 40:1853–1855. 2013.In
Japanese.
|
|
35
|
Zhou S, Ye W, Shao Q, Zhang M and Liang J:
Nrf2 is a potential therapeutic target in radioresistance in human
cancer. Crit Rev Oncol Hematol. 88:706–715. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Menegon S, Columbano A and Giordano S: The
dual roles of NRF2 in cancer. Trends Mol Med. 22:578–593. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nault JC, Rebouissou S and Zucman Rossi J:
NRF2/KEAP1 and Wnt/β-catenin in the multistep process of liver
carcinogenesis in humans and rats. Hepatology. 62:677–679. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Go GW, Srivastava R, Hernandez-Ono A, Gang
G, Smith SB, Booth CJ, Ginsberg HN and Mani A: The combined
hyperlipidemia caused by impaired Wnt-LRP6 signaling is reversed by
Wnt3a rescue. Cell Metab. 19:209–220. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Madan B, Ke Z, Harmston N, Ho SY, Frois
AO, Alam J, Jeyaraj DA, Pendharkar V, Ghosh K, Virshup IH, et al:
Wnt addiction of genetically defined cancers reversed by PORCN
inhibition. Oncogene. 35:2197–2207. 2016. View Article : Google Scholar
|
|
40
|
Jiang X, Hao HX, Growney JD, Woolfenden S,
Bottiglio C, Ng N, Lu B, Hsieh MH, Bagdasarian L, Meyer R, et al:
Inactivating mutations of RNF43 confer Wnt dependency in pancreatic
ductal adenocarcinoma. Proc Natl Acad Sci USA. 110:12649–12654.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wei H, Wang H, Ji Q, Sun J, Tao L and Zhou
X: NRBP1 is down-regulated in breast cancer and NRBP1
overexpression inhibits cancer cell proliferation through
Wnt/β-catenin signaling pathway. Onco Targets Ther. 8:3721–3730.
2015.
|
|
42
|
Rudy SF, Brenner JC, Harris JL, Liu J, Che
J, Scott MV, Owen JH, Komarck CM, Graham MP, Bellile EL, et al: In
vivo Wnt pathway inhibition of human squamous cell carcinoma growth
and metastasis in the chick chorioallantoic model. J Otolaryngol
Head Neck Surg. 45:262016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Fraveto A, Cardinale V, Bragazzi MC,
Giuliante F, De Rose AM, Grazi GL, Napoletano C, Semeraro R, Lustri
AM, Costantini D, et al: Sensitivity of human intrahepatic
cholangiocarcinoma subtypes to chemotherapeutics and molecular
targeted agents: A study on primary cell cultures. PLoS One.
10:e01421242015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Galvao J, Davis B, Tilley M, Normando E,
Duchen MR and Cordeiro MF: Unexpected low-dose toxicity of the
universal solvent DMSO. FASEB J. 28:1317–1330. 2014. View Article : Google Scholar
|