Introduction
Thyroid cancer constitutes the most common endocrine
malignancy increasingly diagnosed worldwide (1–4).
Well-differentiated thyroid carcinomas, including papillary and
follicular carcinomas, accounts for approximately 95% of all
thyroid tumors (5). Respective
patients have an excellent prognosis, with 85–90% benefiting from
early detection and treatment. In contrast, undifferentiated
anaplastic thyroid carcinomas are rare and highly aggressive
tumors. The prognosis of these patients is very poor with a
mortality rate of nearly 100% (4–6).
The preoperative characterization of thyroid nodules
is still a challenge for the clinicians. Nowadays, thyroid
ultrasonography (USG) and thyroid fine needle aspiration (FNA) are
the most commonly pre-operative techniques used for the diagnosis
of malignant thyroid tumors. However, FNA is an invasive procedure
that showed inconclusive biopsy results in 10–20% of all cases,
when solely based on histopathological evaluation. Additional
immunohistochemical markers are thus required for accurate and
reliable diagnosis of thyroid lesions. To address this issue,
attention has been given to galectin-3 and cytokeratin-19 as
potential markers (7–17). However, both markers have some
limitations, because significant expression was reported in few
proportion of benign thyroid nodules (18–23),
suggesting that galectin-3, an adhesion/growth-regulatory tissue
lectin, and cytokeratin-19 are interesting but not completely
reliable immunohistochemical markers to distinguish benign from
malignant thyroid lesions. Thus, there is a need to identify novel
biomarkers that could be used to separate benign from cancer
thyroid nodules. Such biomarkers could also offer new insights into
the biology of thyroid cancers, with potential for innovations of
targeted therapies.
Following the traits of galectin-3, the emerging
network concept for this class of multifunctional effectors working
in the cell and extracellularly guides interest to further members
of this family (24,25). Several galectins can be expressed
by tumors, as documented for colon and head and neck cancer as well
as also in other types of diseases (26–28),
with marked impact on cancer progression by itself and by
functional cooperation between the proteins. Homodimeric galectin-1
is an apparent candidate for analysis, sharing counter-receptors
with galectin-3, and exhibits a tumor-type-specific impact on
progression (29–32). The expression of galectin-1 has
been shown to be upregulated in thyroid carcinoma-derived cell
lines (33–35), and our team has previously reported
for the first time a high serum level of galectin-1 in patients
diagnosed with a well-differentiated thyroid cancer (36).
In the present study, we compared the diagnostic
value of the galectin-1 to galectin-3 and cytokeratin-19 using a
panel of benign and malignant thyroid lesions in order to answer
the question whether galectin-1 may be viewed as a biomarker for
diagnosis of thyroid cancer. We also examined the implication of
galectin-1 depletion on proliferation and invasion of papillary and
anaplastic thyroid cancer cell lines as well as on tumor growth in
an orthotopic nude mouse model.
Materials and methods
Clinical data
The immunohistochemical detection of galectin-1 and
-3 was first studied in a clinical series of specimens of four
healthy patients [gender: 2 male/2 female, median age (range): 34
years (23–54), 4 total thyroidectomies], 40 patients with benign
thyroid lesions (10 colloid nodules, 16 follicular adenomas, 7
Hashimoto's thyroiditis and 7 Graves' disease), and 29 patients
with malignant thyroid lesions (17 papillary carcinomas and 12
follicular variant of papillary carcinomas). For these 69 cases,
the age range was 22–81 years with a median of 54; 9 patients were
male and 60 were female (36 hemi-thyroidectomies and 33 total
thyroidectomies). To extend this series, two tissue microarrays
(TMA) composed of 66 follicular adenomas and 66 papillary
carcinomas were processed accordingly (galectin-1, galectin-3 and
cytokeratin-19). Written informed consent was obtained from all the
patients to use the surgical specimens for scientific research.
Approval for any experiments was obtained from the institutional
ethics committee. The patient samples and clinical data were
collected retrospectively from the records of the pathology
departments of the CHU Erasme (Brussels, Belgium), the EpiCURA
Center (Baudour, Belgium), the Saint Pieter's Hospital-Bordet
Institute (Brussels, Belgium) and the Lille University Hospital
(Lille, France) selected by two pathologists (Professor M.
Remmelink and Professor E. Leteurtre), then analyzed
semi-quantitatively (blind scoring) using a light microscope
(Axiocam MRc5, Zeiss, Oberkochen, Germany). All samples were
collected with written informed consent and the study was approved
by the ethics committee of the University of Mons (Mons, Belgium)
(OM 004).
Immunohistochemistry on tissue specimens
and tissue microarrays
All tumor samples were routinely fixed for 24 h in
4% buffered formaldehyde, dehydrated and then embedded in paraffin.
Immunohistochemistry was performed on 5 µm sections that
were mounted on silane-coated glass sides. Before starting the
immunodetection, tissue sections were briefly exposed to microwave
pre-treatment in a 0.01M citrate buffer (pH 6.2) for 7 min at 900 W
and then for 10 min at 350W. The sections were then pretreated with
0.06% hydrogen peroxide solution (H2O2) for 4
min to block endogenous peroxidase activity, rinsed in
phosphate-buffered saline (PBS) and successively exposed for 5 min
to solutions containing avidin (0.1 mg/ml PBS) and biotin (0.1
mg/ml PBS) to avoid false-positive staining reaction from
endogenous biotin. After a washing step with PBS, the sections were
incubated for 15 min with a solution of 0.5% casein in PBS and
sequentially exposed at room temperature to solutions of i)
specific primary antibody (1 h): Galectin-1 (polyclonal rabbit
anti-human galectin-1: 1:100) (29,37),
Galectin-3 (polyclonal rabbit anti-human galectin-3: 1:200)
(29,37), CK19 (monoclonal mouse anti-human
cytokeratin 19: 1:50; M0772; Dako, Glostrup, Denmark); ii)
corresponding biotinylated secondary antibody (30 min): polyclonal
goat anti-rabbit/mouse IgG (1:50; BA-1000/BA-9200, Vector
Laboratories, Burlingame, CA, USA); and iii)
avidin-biotin-peroxidase complex (ABC Kit Vector Laboratories). The
specificity of galectin-1 and -3 antibodies was validated by
western blotting in different thyroid cancer cell lines (B-CPAP,
FTC133C and 8505C cell lines derived from papillary, follicular and
anaplastic thyroid carcinoma respectively) reporting immunoreactive
band for galectin-1 at 14 kDa and for galectin-3 at 26 kDa (data
not shown). The slides were thoroughly washed with PBS between
incubation steps. Immunocomplexes were finally visualized by
exposure to the chromogen diaminobenzidine (DAB, BioGenex, Fremont,
CA, USA) in the presence of H2O2. After
rinsing, the sections were counterstained with luxol fast blue and
mounted with a synthetic medium. To exclude antigen-independent
staining, the incubation step with primary antibodies was omitted
from the protocol as negative controls. In all cases, these
controls were negative (data not shown). Individual tissue
specimens were given a score (0–6) by adding the percent of
immunopositive cells (range 0–3: 0=0%, 1=1–33%, 2=34–66%, and
3=67–100%) to the intensity score (range 0–3, 0=no, 1=low,
2=moderate, and 3=high).
Cell culture conditions
Two cell lines from human thyroid cancers were
studied: TPC-1 derived from papillary carcinoma and 8505C derived
from anaplastic carcinoma (kindly gifted by Professor C. Maenhaut,
IRIBHM ULB, Brussels, Belgium). The HeLa cell line derived from a
cervical carcinoma (kindly gifted by Dr J. Martial, ULg, Liège,
Belgium) was used as the positive control of galectin-1 expression.
The TPC-1 and 8505C cell lines were grown in RPMI-1640 (Lonza,
Verviers, Belgium) supplemented with 10% fetal bovine serum (FBS,
Lonza, Verviers, Belgium) and 1% penicillin/streptomycin (PAA
Laboratories, Pasching, Austria). HeLa cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% FBS
and 1% penicillin/streptomycin. Cells were incubated at 37°C in a
humidified 95% air/5% CO2 atmosphere. The culture medium
was changed once every two days. For routine maintenance and
experimental studies, cells were detached by incubation with
accutase solution (PAA Laboratories), resuspended and counted using
an electronic cell counter (model Z1 Coulter counter, Beckman
Coulter, Fullerton, CA, USA) before plating.
Depletion of galectin-1
Galectin-1 knockdown (gal-1 KD) transfectant cells
derived from TPC-1 and 8505C, as well as matched control cells with
normal galectin-1 expression, were generated in our laboratory.
Knockdown of galectin-1 expression was achieved by using shRNA
lentiviral particles targeting galectin-1 specific mRNA
(sc-35441-v, Santa Cruz Biotechnology, Santa Cruz, CA, USA).
Control cells were obtained by transduction with shRNA lentiviral
particles encoding a scrambled shRNA sequence (gal-1 sc). TPC-1 and
8505C cells were processed according to the manufacturer's
instructions. Stable clones were selected by addition of 2
µg/ml puromycin (InvivoGen, San Diego, CA, USA) to the
culture medium, preliminary experiments showed that this
concentration was toxic on non-transduced cells.
Immunofluorescence staining
Twelve-well culture plates containing sterile round
glass coverslips were seeded at a density of
104–105 cells/well and grown for four days.
The cells were washed with PBS and fixed with 4% paraformaldehyde
(PFA) for 15 min at room temperature. The fixed cells were rinsed
in PBS, permeabilized with 0.1% Triton X-100 in PBS for 15 min and
blocked with 0.05% casein in PBS (blocking solution) for 20 min.
The cells were incubated overnight with specific primary antibodies
which were diluted in blocking solution. The following primary
antibodies were used: galectin-1 (polyclonal rabbit anti-human
galectin-1 at 1:100, 1 h) (29,37)
and matrix metalloproteinase 9 (MMP9) (polyclonal rabbit anti-MMP9
at 1:50, 1 h; sc-10737, Santa Cruz Biotechnology). Then, the cells
were washed with PBS containing 0.1% Triton X-100 and incubated for
30 min with Alexa Fluor 488-conjugated anti-rabbit IgG (A-11008,
Invitrogen, Gent, Belgium) diluted 1:50 in blocking solution. The
cells were washed with PBS containing 0.1% Triton X-100 for 15 min,
rinsed with distilled water for 5 min and mounted with Vectashield
Mounting Medium containing DAPI (Vector Laboratories), then
examined by confocal microscopy using an Olympus FV1000D laser
scanning inverted microscope (Olympus, Hamburg, Germany).
Western blotting
Cells were seeded in a T-75 culture flask and grown
at 37°C and 5% CO2 until confluence. The cells were
washed with PBS and recovered on ice with a scraper in PBS. The
pellet was resuspended in a detergent cocktail (M-PER Mammalian
Extraction buffer) supplemented with protease (Halt protease
inhibitor cocktail, Pierce, Rockford, IL, USA) and phosphatase
inhibitors (Halt phosphatase inhibitor cocktail, Pierce). Protein
concentrations were determined by the BCA Protein assay (Pierce)
using bovine serum albumin as standard. Extracted proteins (40
µg) were subjected to 10% SDS-PAGE and electrotransferred at
200 mA for 90 min on nitrocellulose membranes (iBlot®
Dry Blotting system, Life Technologies-Invitrogen, Gent, Belgium).
Immunodetection was performed using polyclonal rabbit anti-human
galectin-1 (o/n, 1:1,000) (29,37).
Peroxidase-labeled anti-rabbit IgG antibody (1 h, 1:5,000; RPN4301,
GE Healthcare, Diegem, Belgium) was used as secondary reagent.
Peroxidase activity was visualized using the
SuperSignal® West Pico Chemiluminescent Substrate
(Pierce) following the manufacturer's instructions. Molecular
weight markers were analyzed in parallel for internal calibration
(PAGE Ruler Prestained Proteins, Thermo Fisher Scientific, Waltham,
MA, USA). For semi-quantitative analysis, nitrocellulose membranes
were reloaded with a mouse monoclonal anti-β-actin antibody (clone
C4, 1:5,000; MAB1501, Millipore, Temecula, CA, USA) detected using
a peroxidase-labeled anti-mouse IgG antibody (1/5,000) (NA931,
Amersham Pharmacia Biotech, Roosendaal, The Netherlands) and
reagents of the Lumi-Light kit (Roche, Brussels, Belgium). Films
were scanned (Image Reader, Raytest®, Straubenhardt,
Germany) and the immunoreactive band intensities were quantified
using the software AIDA® Image Analyser 3.45
(Raytest).
RNA extraction and real-time PCR
Cells were plated in T-75 culture flasks and
cultured until confluence and lysed in RLT buffer supplemented with
β-mercaptoethanol (RNeasy mini kit, Qiagen, Venlo, The Netherlands)
at 4°C, and then centrifuged with RNeasy spin column. After washing
steps, RNA was collected in RNase-free water and subjected to DNase
treatment as described by the manufacturer. RNA concentration was
evaluated using a NanoDrop™ 1000 spectrophotometer (Thermo Fisher
Scientific, Wilmington, DE, USA). Then, expression profiles of
galectin-1 and 18S RNAs were quantified by real-time PCR. cDNA was
synthesised using a standard reverse transcription method (qScript
cDNA SuperMix, Quanta Biosciences, Gaithersburg, MD, USA).
Real-time PCR reactions were performed using the SYBR Green PCR
Master Mix (Applied Biosystems, Foster City, CA, USA) and
sequence-specific primer sets designed from PrimerBank (http://pga.mgh.harvard.edu/primerbank/)
for galectin-1 (forward: 5′-TCGCCAGCAACCTGAATCTC-3′, reverse:
5′-GCACGAAGCTCTTAGCGTCA-3′, amplicon size: 81) and from Primer 3
(http://frodo.wi.mit.edu) for 18S (forward:
5′-GCGGCGGAAAATAGCCTTTG-3′, reverse: 5′-GATCACACGTTCCACCTCATC-3′,
amplicon size: 139) (Life Technologies, Gent, Belgium). The
amplification was performed on a LightCycler® 480 System
(Roche Diagnostics GmbH, Mannheim, Germany) using an initial
activation step (95°C for 10 min) followed by 40 cycles of
amplification (95°C for 15 sec and 60°C for 60 sec). Melting curves
from 60°C to 99°C were assessed to evaluate PCR specificity. A
preliminary analysis demonstrated linear and similar amplification
efficacies. Relative quantification was determined by normalizing
the crossing threshold (CT) of galectin-1 with the CT of 18S
(loading control) using the 2−ΔCT method (38).
Measurement of cell culture growth by
cell counting
Transduced TPC-1 and 8505C cells were plated at a
density of 15000 cells/cm2 in 12-well dishes. The next
day, cell cultures were refed with fresh medium. Measurement of
cell culture density was performed over 7 days after plating. Cells
were detached by incubation with accutase solution (PAA
Laboratories). After vigorous pipetting, the amounts of cells in
suspension were determined using an electronic cell counter (model
Z1 Coulter counter, Beckman Coulter).
Cell migration assay
Cell migration assay was performed using a wound
healing assay (80209, Ibidi, Martinsried, Germany). The kit was
used according to the manufacturer's instructions. Transfectants of
the TPC-1 and 8505C lines were seeded at a density of
5×105 cells/well in a 6-well plate cultured with
RPMI-1640 complete medium and incubated for 24 h. Culture inserts
were removed creating a cell-free gap of 500 µm and cells
were washed with medium. Scratch wounds were observed under a
microscope (Carl Zeiss Axiovert 25C, Göttingen, Germany) and
residual scratch distances were measured on images captured at 0,
4, 6 and 21 h after wounding.
Cell invasion assay
Cell invasiveness was assessed using a Boyden
chamber assay consisting a 12-well plate (lower chambers) with cell
culture inserts (upper chambers). The two chambers were separated
by polycarbonate membrane (8-µm pore size) coated with an
artificial extracellular matrix (ECM550, Chemicon Cell Invasion
assay kit, Millipore, Billerica, MA, USA). The kit was used
according to manufacturer's instructions. Gal-1 sc and gal-1 KD
cells were seeded into the inserts (5×105 cells/insert)
in serum-free medium (RPMI-1640) and complete medium (RPMI-1640,
10% FBS) was added to the lower chamber. After 96 h, non-invading
cells on the upper side of the inserts were removed using a
cotton-tipped swab and invading cells on the bottom side of the
inserts were stained with crystal violet. Quantification was
performed by counting the stained cells under an optical microscope
in quadruplicate (Axiocam MRc5, Zeiss).
Mouse tumor models
Four-week-old male nude mice were used (n=18,
Charles River Laboratories, L'arbresle, France) and maintained
under pathogen-free conditions for one week. The orthotopic
transplantation was performed using 5×105 cells (8505C
control and Gal-1 knockdown cells) suspended in Hank's balanced
salt solution (HBSS) and, after cervicotomy under general
anesthesia (ketamine-xylazine, 0.1 ml/10 g, Sigma-Aldrich, St.
Louis, MO, USA), injected into the left thyroid gland during
microsurgery (Professor S. Saussez). Tumor growth was weekly
followed by magnetic resonance imaging (MRI) using a 7T scanner
(PharmaScan, Bruker, Ettlingen, Germany) equipped with a 40-mm
volume coil and controlled with the ParaVision software. Animals
were kept anesthetized with 1–1, 5% isoflurane delivered in oxygen.
Their respiratory rate was monitored during the entire imaging
session. A T2-weighted TurboRARE sequence was used: 19 axial slices
of 0.75-mm thickness with TR: 2000 msec, TE: 30 msec, 0.100×0.100
mm resolution, NEX: 20. Tumor volumes were calculated from MR
images using the VivoQuant software (InviCRO, Boston, MA, USA) by
manually contouring the tumor on each slice. Mice were sacrificed
50 days after tumor cell inoculation or when mouse weight decreased
to 20% under its initial weight (Nembutal, 0.1 ml/10 g, Ceva,
Brussels, Belgium). Tumors with adjacent tissues and lungs were
collected and analyzed by hematoxylin/eosin coloration searching
for tumor and metastases. All animal experiments were performed
according to the institutional guidelines and approved by the
ethics committee of the University of Mons (Mons, Belgium)
(LVE-01–01).
Statistical analysis
The non-parametric Mann-Whitney U test as well as
the Kruskal-Wallis test were used to investigate significant
differences between the groups. Statistical evaluations included
also Fisher's exact test. Cox regression and Kaplan-Meier method
were used to perform survival analysis. A P-value <0.05 was
considered statistically significant.
Results
Galectin-1, galectin-3 and cytokeratin-19
immunostaining profiles in benign and malignant thyroid
lesions
The first aim of our study was to assess
immunohistochemically presence of galectin-1 and -3, in 40 benign
thyroid lesions [including 16 adenomas (AD), 7 cases of Graves'
disease (GD), 7 Hashimoto's thyroiditis (HT), 10 colloid nodules
(CN)] and in 29 malignant thyroid lesions [including 17 papillary
carcinomas (PC) and 12 follicular variants of papillary carcinoma
(FVPC)] and in four normal thyroid cases (N). The analysis of the
cytoplasmic immunostaining revealed that both galectin-1 and
galectin-3 were significantly higher expressed in cancer cells of
malignant thyroid lesions (PC and FVPC) compared to epithelial
cells in benign lesions (AD, GD, HT, CN) and in normal controls (N)
(p=0.000002 and p=0.0002 for, respectively, galectin-1 and
galectin-3; Kruskall-Wallis test). Galectin-1 expression was
completely absent in the epithelial compartment of all benign
thyroid neoplasms (Fig. 1A and B),
whereas a weak to moderate expression was found in benign samples
for galectin-3 (Fig. 1A and C),
suggesting that galectin-1 can be more reliable to distinguish
malignant from benign lesions. Of note, considering the tumor
micro-environment, stromal cells were also positive for galectin-1
and galectin-3, and both galectins were immunodetected in both
benign and malignant lesions (Fig.
1A).
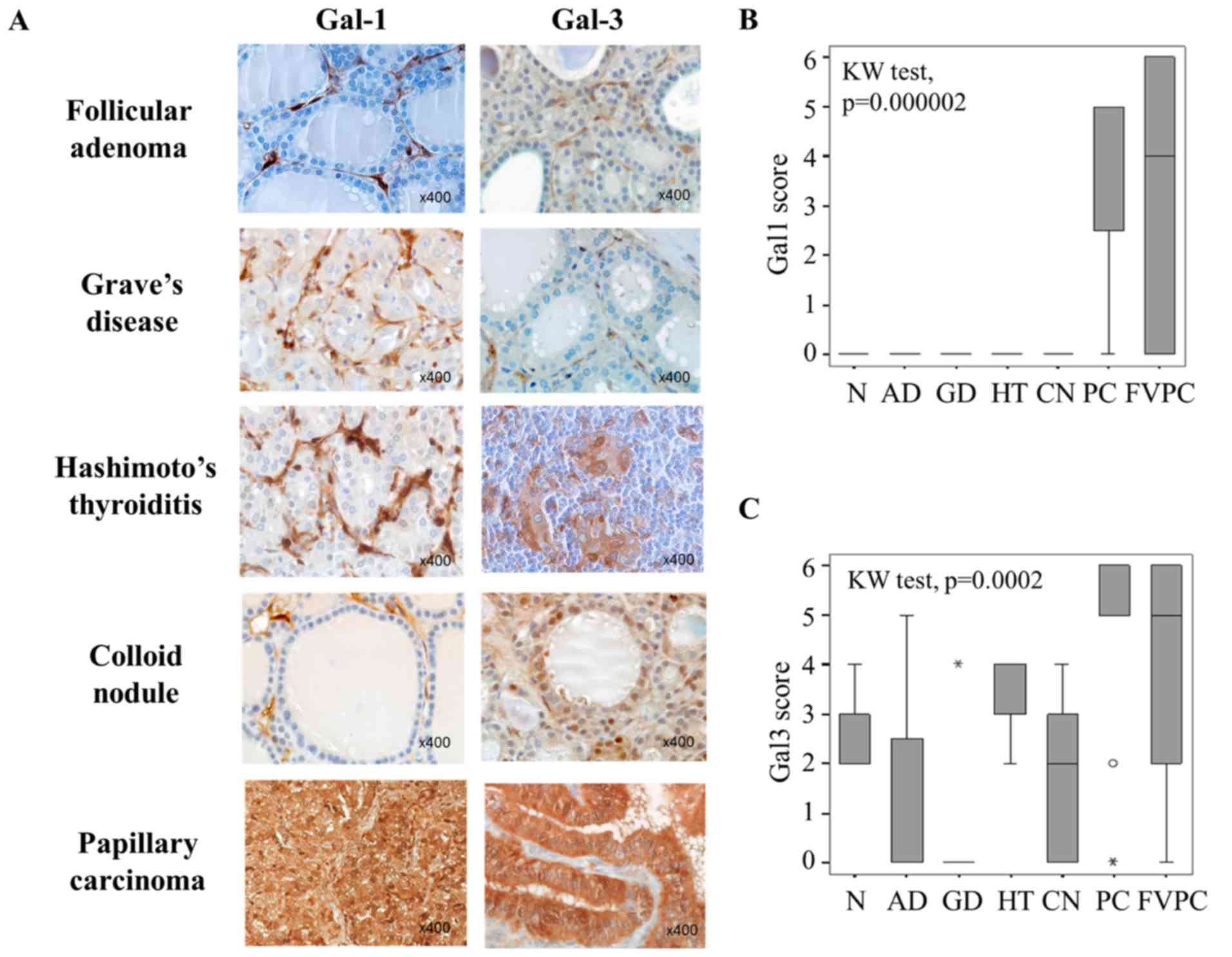 | Figure 1Evaluation of galectin-1 and
galectin-3 expression in benign and malignant thyroid lesions. (A)
Immunohistochemical expression of galectin-1 and galectin-3 in
benign (follicular adenoma, Graves' disease, Hashimoto's
thyroiditis, colloid nodule) and malignant (papillary carcinoma)
thyroid lesions. Magnification ×400. (B and C) Statistical analysis
of galectin-1 and galectin-3 expression in a series of 69 cases of
benign and malignant thyroid lesions (p<0.001, Kruskal-Wallis
test). NT, normal tissue; AD, follicular adenomas; GD, Graves'
diseases; HT, Hashimoto's thyroiditis; CN, colloid nodules; PC,
papillary carcinomas; FVPC, follicular variant of papillary
carcinomas. The immunostaining was semi-quantitatively assessed in
the cytoplasmic compartment and scored from 0 to 6 by combining
staining intensity (0–3) and percentage of positivity (0–3). Data
are presented as box plots indicating the 1st and the 3rd quartiles
centered on medians (thick lines) with whiskers for the minimum and
maximum non-outlier values. |
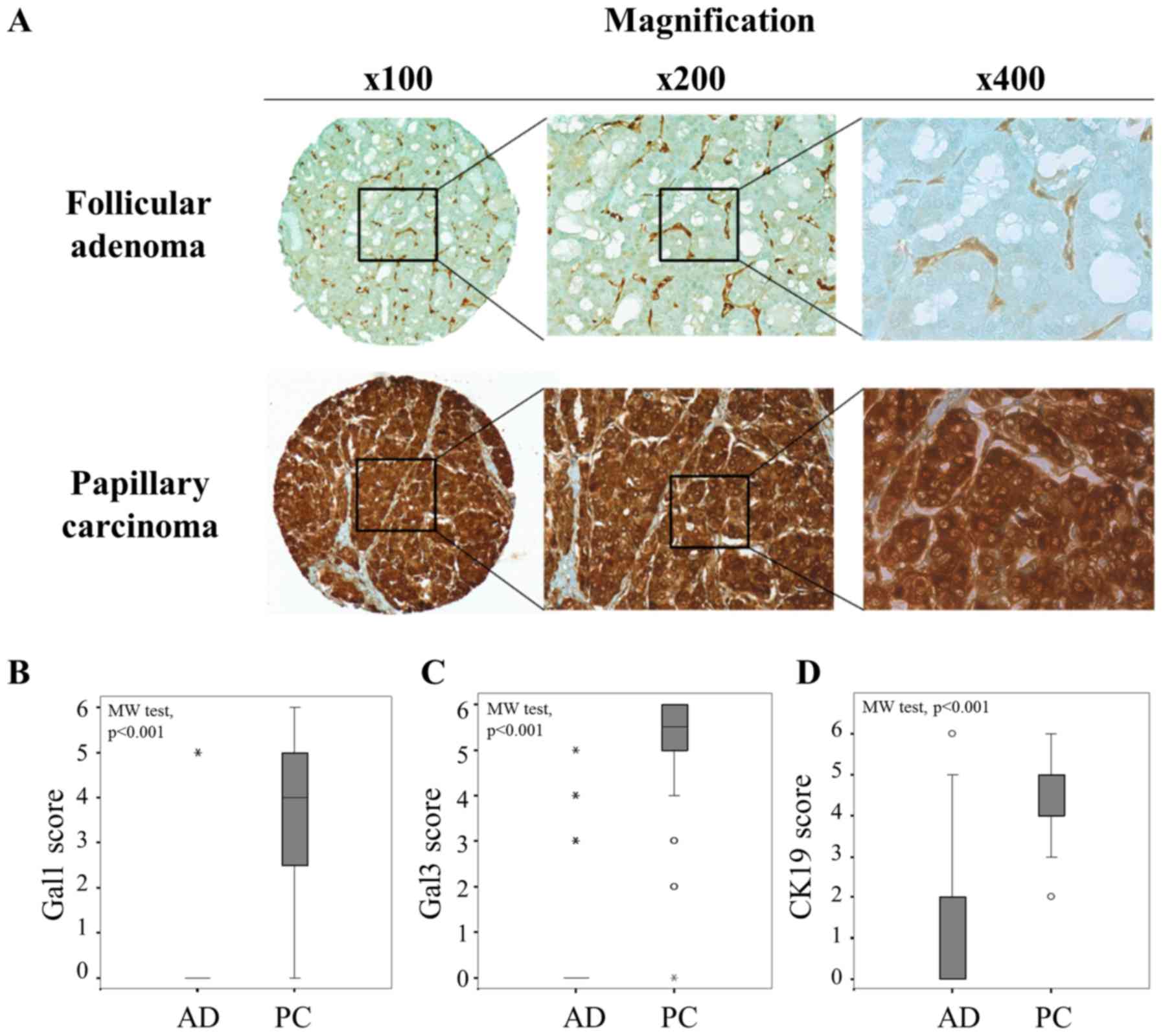
Extending these results by analysis of two series of
tissue microarrays (TMA) composed of 66 follicular adenomas and 66
papillary carcinomas, a significantly higher expression of
galectin-1, -3 and cytokeratin-19 was observed in papillary
carcinomas (PC) than in follicular adenomas (AD) (p<0.001,
Mann-Whitney test) (Fig. 2),
supporting that the diagnostic value of galectin-1 was as good as
those of galectin-3 and cytokeratine-19. Of note, the cutoffs that
we defined to separate negative/low versus positive immunostaining
were >0 for galectin-1 and 3, and >2 for cytokeratin-19
(Fig. 2B–D). From crosstab
analyses, the sensitivity, specificity and the positive/negative
predictive values (PPV and NPV) were calculated for each marker
(Table I). The data showed that
galectin-1 exhibited an excellent specificity (97%) to discriminate
malignant from benign thyroid lesions with a very good PPV (96%),
while the highest performances for galectin-3 and cytokeratin-19
were rather seen for the sensitivity (>97%) and the NPV
(>96%). Thus, analysis of galectin-1 could provide complementary
information refining the conventional diagnostic markers.
 | Table IDiagnostic value of galectin-1,
galectin-3 and CK19 in discrimination of malignant from benign
thyroid tumors. |
Table I
Diagnostic value of galectin-1,
galectin-3 and CK19 in discrimination of malignant from benign
thyroid tumors.
| Proteins | Specificity
(%) | Sensitivity
(%) | PPV (%) | NPV (%) | Fisher's exact
test |
|---|
| Galectin-1 | 97 | 80 | 96 | 83 | p<0.001 |
| Galectin-3 | 83 | 97 | 85 | 96 | p<0.001 |
| CK19 | 76 | 98 | 80 | 88 | p<0.001 |
Impact of galectin-1 depletion on
papillary and anaplastic thyroid cancer cell proliferation,
migration and invasion
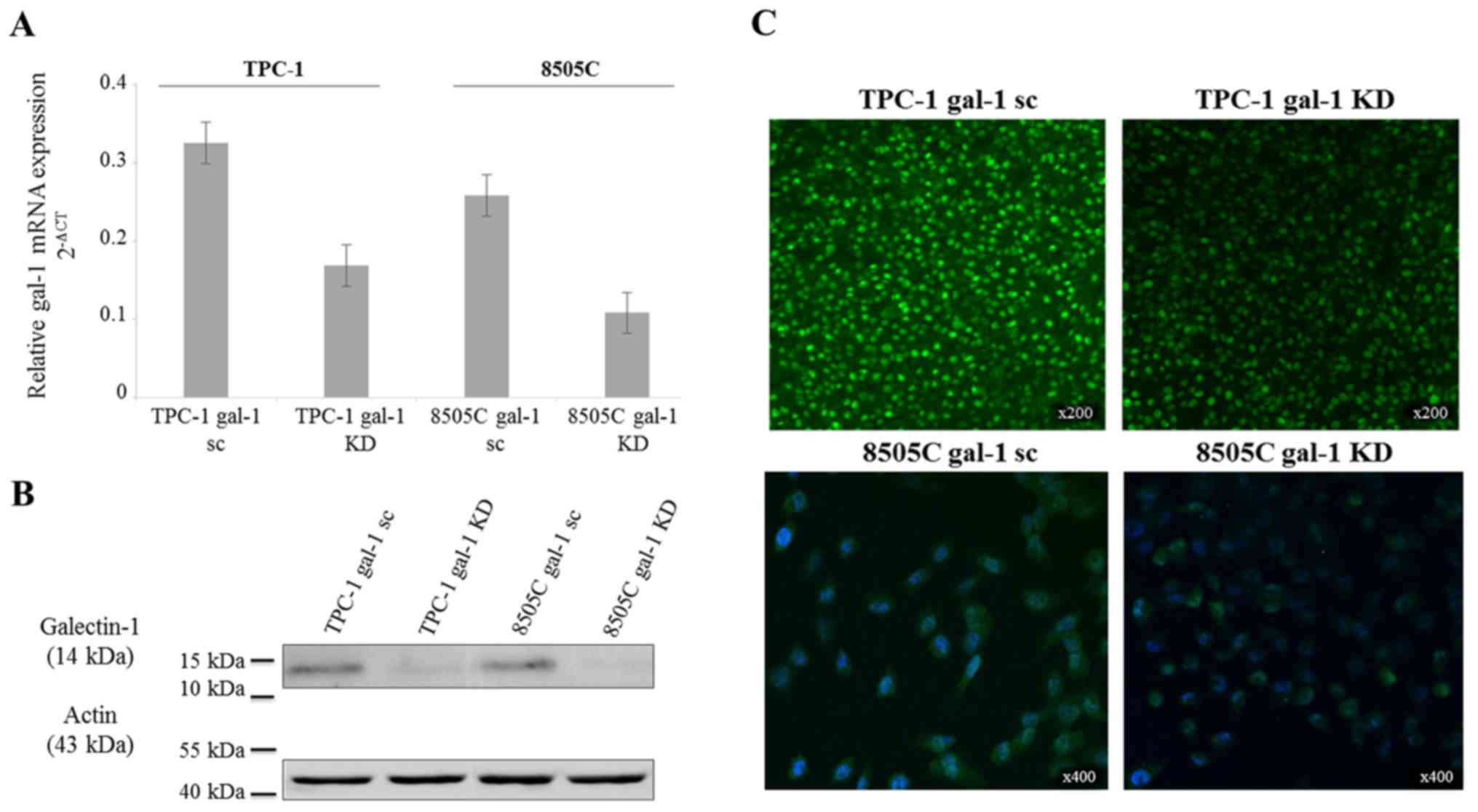
To explore a galectin-1 functionality in thyroid
cancer cells, we used a lentivirus-mediated shRNA to knock down
galectin-1 expression in TPC-1 and 8505C cell lines derived from
human papillary and anaplastic carcinoma, respectively. The
baseline expression level of galectin-1 mRNA was slightly lower in
8505C wild-type cells than in TPC-1 wild-type cells. RT-qPCR,
immunofluorescence and western blotting were realized to confirm
the effective reduction (approximately 50%) of galectin-1-specific
mRNA and protein expression (Fig.
3).
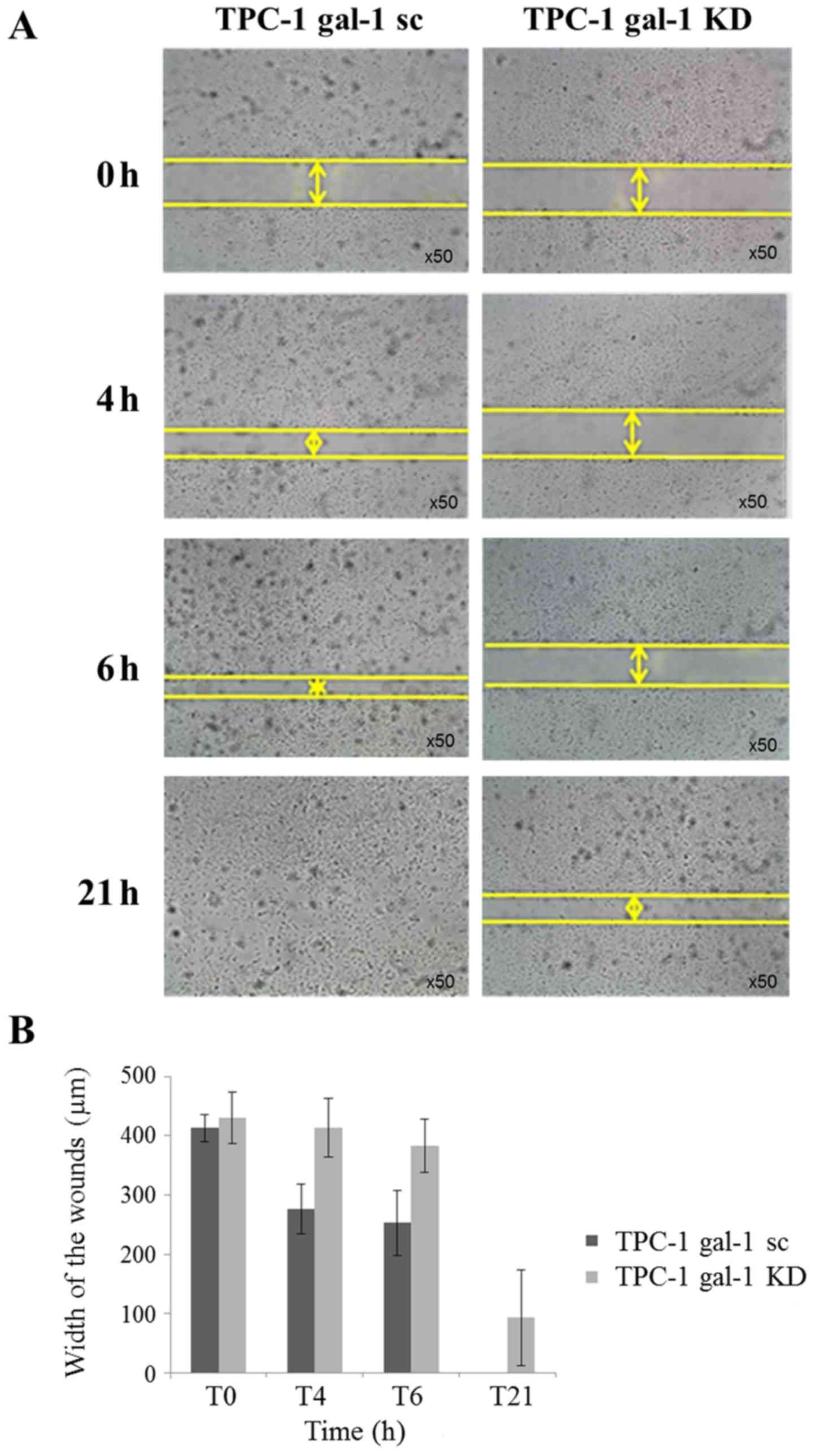
First, we evaluated the effect of galectin-1
depletion on TPC-1 cells and observed no effect on cell
proliferation and invasion (data not shown) but a significant
reduction of cell migration. Monolayer wound healing assays were
performed and the wound-closure distances have been measured
comparing scrambled (TPC-1 gal-1 sc) and knockdown (TPC-1 gal-1 KD)
TPC-1 cells (Fig. 4A).
Quantification of cell motility over 21 h after scratching clearly
showed that migration was significantly decreased in galectin-1
knockdown cells (Fig. 4A and
B).
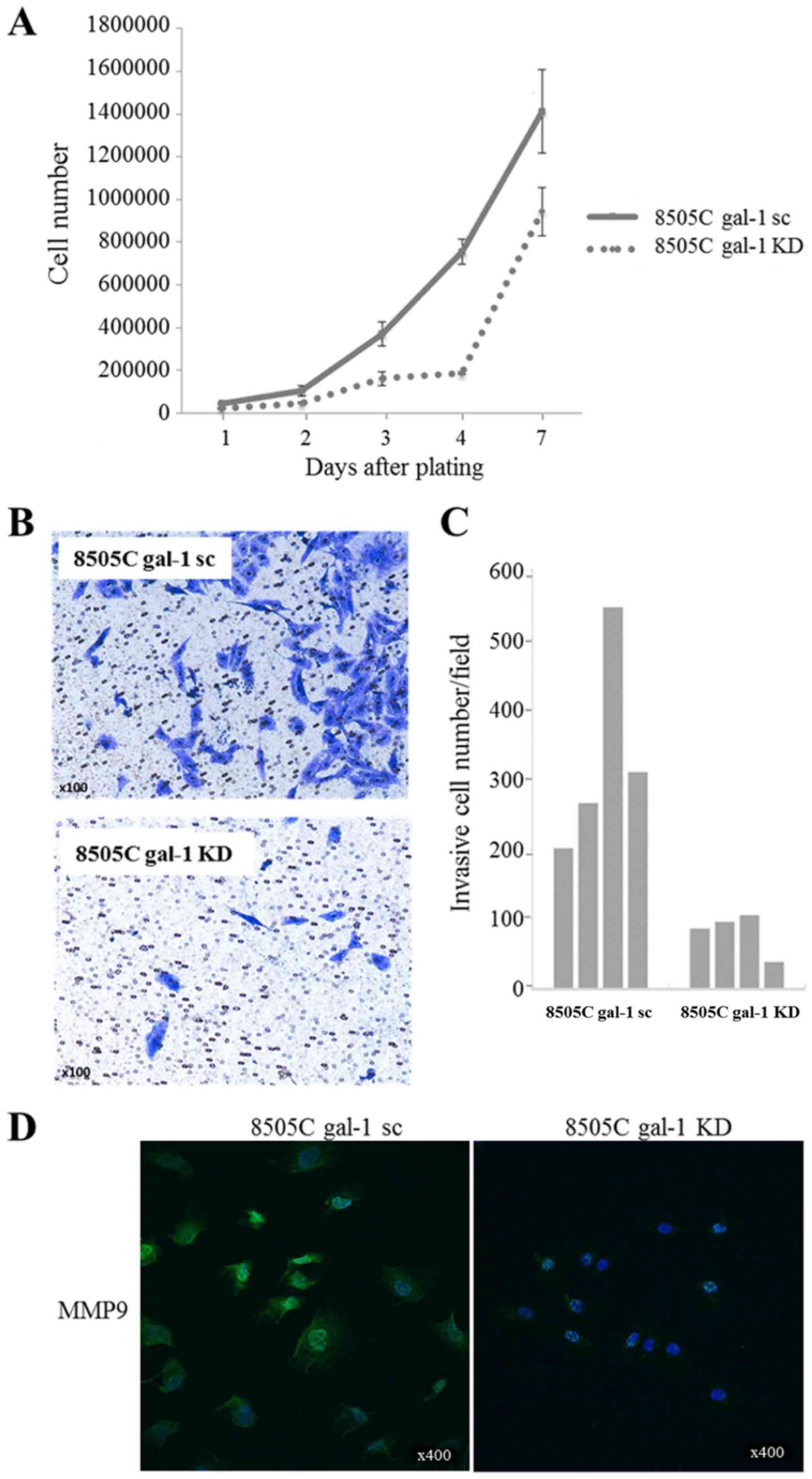
Then, we examined the effect of galectin-1 knockdown
on 8505C anaplastic cell proliferation (cell counting over 7 days).
As illustrated in Fig. 5A, cell
densities were significantly lower in galectin-1 knockdown cells
(8505C gal-1 KD) than in scrambled cells (8505C gal-1 sc),
indicating that the depletion of galectin-1 could potently reduce
the proliferation of thyroid cancer cells. Cell invasion assays in
Boyden chambers were also performed to assess the impact of
galectin-1 knockdown on 8505C cell invasiveness. In this case, the
depletion of galectin-1 resulted in a significant inhibition of
cell invasion compared to scrambled cells (Fig. 5B). Decrease in cell invasion was
confirmed by counting the cell number that migrated through the
membrane (Fig. 5C). Accordingly,
the expression of the matrix metalloproteinase MMP9 in 8505C cells
was significantly reduced as a result of galectin-1 knockdown
(Fig. 5D). In contrast to our
observations in TPC-1 cells, we did not observe any significant
effect of galectin-1 depletion on 8505C cell migration (data not
shown).
Effect of galectin-1 depletion on tumor
growth in an orthotopic nude mouse model
A murine orthotopic model of human anaplastic
thyroid carcinoma was developed to investigate the impact of
galectin-1 depletion on tumor growth and metastatic dissemination
in vivo. Control (CT, n=8) and galectin-1 knockdown (KD,
n=10) 8505C anaplastic tumor cells were injected into the thyroids
of nude mice. Primary tumor growth was followed in vivo
using an MRI imaging system. In this model, tumor volume analysis
revealed that all mice injected with 8505C control cells (n=8)
developed thyroid tumors within 22 days of injection (Fig. 6A top panel and C). In contrast, mice injected with 8505C
gal-1 KD cells showed slower (n=4) or absent (n=6) tumor
development at days 22 and 29 after inoculation (Fig. 6B top panel and C). At day 42 post-injection, final tumor
volumes were 53.1–1174.4 mm3 and 0–99.1 mm3;
for control and galectin-1 knockdown mice, respectively. The
Kaplan-Meier survival curve indicated that the downregulation of
galectin-1 was significantly associated with better survival rate
(univariate Cox regression, p=0.026) (Fig. 6D). Histological analysis at day 50
of control tumors showed histopathological pattern of anaplastic
thyroid carcinoma and revealed the presence of lung metastasis in
all control mice (Fig. 6A middle
and bottom panels). Of note, mice injected with galectin-1 KD cells
showed small tumors at inoculation sites and minimal number of lung
metastasis (Fig. 6B middle and
bottom panels). The number of lung metastases (counted on five
histological slides/mice) ranged from 4 to 1048 (median 50.5) and 0
to 14 (median 7) for control and galectin-1 knockdown mice,
respectively (p=0.001, Mann-Whitney test) (Fig. 6E). Collectively, these data suggest
that galectin-1 is required for tumor growth and lung metastasis
development.
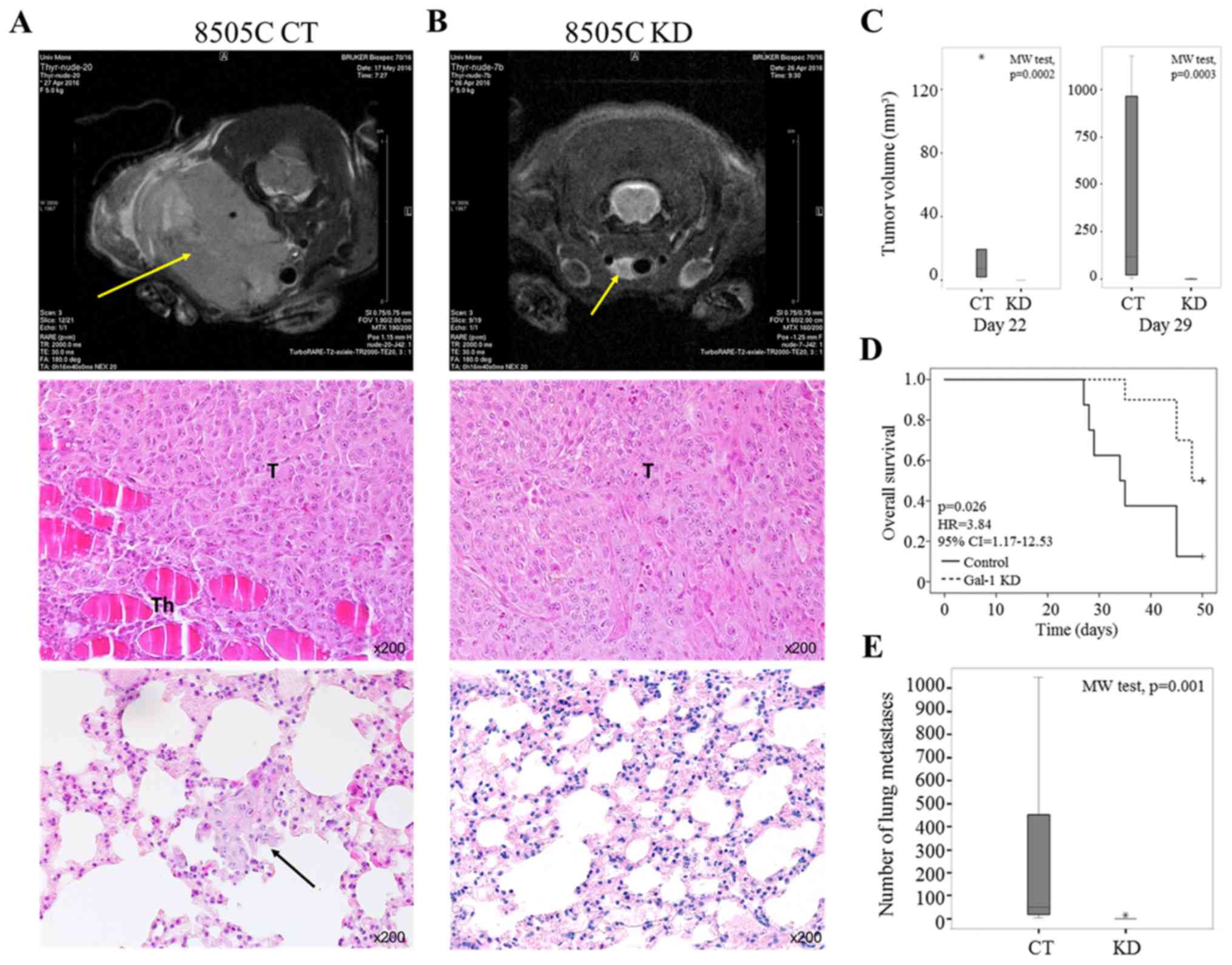 | Figure 6Orthotopic mouse model of anaplastic
thyroid carcinoma. (A and B) The upper panel shows the
representative axial sections of MRI recorded in nude mice at days
42 after 8505C control (CT) or knockdown (KD) tumor cell
inoculation. The yellow arrows indicate the tumor localization. The
middle and bottom panels represent histological examination of,
respectively, tumor in animal thyroid and lung invasion at day 42
after 8505C control (A) or knockdown (B) tumor cell inoculation.
The black arrow indicates lung metastasis in control mice. H&E
staining (magnification, ×200). T, Tumor; Th, Thyroid gland. (C)
Tumor volumes comparing control (CT) and galectin-1 knockdown (KD)
mice at days 22 (left panel; p=0.0002, Mann-Whitney test) and 29
(right panel; p=0.0003, Mann-Whitney test) after cancer cell
inoculation. Data are presented as box plot where thick line is the
median, bottom and top of the box are, respectively, the 1st and
the 3rd quartiles, whiskers indicate the minimum and maximum
non-outlier values, and * shows an extreme value. (D)
Kaplan-Meier survival curves for control (CT) and galectin-1
knockdown (KD) mice (Cox regression, p=0.026). (E). Graph
displaying the number of lung metastases in control (CT) and
galectin-1 knockdown (KD) mice (p=0.001, Mann-Whitney test). Data
are presented as box plot where thick line is the median, bottom
and top of the box are, respectively, the 1st and the 3rd
quartiles, whiskers indicate the minimum and maximum non-outlier
values, and * shows an extreme value. |
Discussion
Although conventional histology and FNA are
considered as gold standards, the pathologists are confronted with
difficulties in reaching an accurate differential diagnosis between
benign and malignant thyroid nodules. To improve disease
identification, immunohistochemical markers, such as cytokeratin-19
(CK19) and galectin-3, have been proposed and their efficiencies
for thyroid cancer diagnosis have been evaluated. CK19 is the
smallest member of cytokeratin family and belongs to the
intermediate filaments. Several studies reported that CK19
expression is strong and diffuse in papillary carcinoma and absent
or low in benign thyroid lesions (8,9,15–17).
Galectin-3, a structurally unique member of galectin family with an
N-terminal tail composed of nine collagen-like repeats and a
peptide with sites for Ser phosphorylation (39), is anti-apoptotic, associated with
the pathogenesis of well-differentiated thyroid carcinoma (7,10–14).
However, these markers have shown some limitations because
positivity was also reported in some benign cases (16–23).
Mehrotra et al showed that galectin-3 was expressed in a
large proportion of follicular adenomas, multinodular goiters and
Hashimoto's thyroiditis (19). In
the study of Mataraci et al, CK19 expression was found in
adenomatous nodular hyperplasia and follicular adenoma (16). Thus, the current challenge is to
find new immunohistochemical markers that might be more helpful to
refine diagnosing thyroid cancer, to avoid unnecessary
thyroidectomy. In this context, our data confirm previous studies
preconizing the use of galectin-3 and CK19 as routine biomarkers to
diagnose thyroid carcinomas (40,41).
More importantly, it provides supplementary
information on galectin-1 expression in both benign and malignant
thyroid lesions. Our data showed that the expression of these
markers is significantly higher in papillary carcinoma than in
follicular adenoma. Herein, the galectin-1 displayed a high
specificity of 97%, suggesting that galectin-1 can be used in
association with galectin-3 and CK19 as a complementary diagnostic
tool to improve discrimination between benign and malignant thyroid
neoplasms. Our observations validate and further support the
potential value of galectin-1 in the diagnosis of well
differentiated thyroid cancer as proposed by previous studies
(33–35).
Since its discovery, altered galectin expression was
frequently associated with many cellular processes and malignant
transformation. In particular, galectin-1 was reported to
contribute to pathogenesis, in a favorable or unfavorable manner
depending on the cell type (32).
Of relevance, a high serum level of galectin-1 occurs in patients
diagnosed with a well-differentiated thyroid cancer (36). However, mechanisms by which
galectin-1 can exert diverse effects in thyroid carcinogenesis and
cancer progression remain still unknown. In the present study, we
investigated the role of galectin-1 in the biology of thyroid
carcinoma using shRNA strategy in two thyroid cancer cell lines.
Our findings revealed that knockdown of galectin-1 resulted in the
inhibition of TPC-1 cell migration, 8505C cell proliferation and
invasion in vitro as well as in the remarkable decrease of
tumor (and metastasis) development in vivo. This body of
evidence suggests that galectin-1 can be involved in the
progression of thyroid carcinoma. Particularly, the impact of
galectin-1 knockdown on invasion of anaplastic cancer cells (8505C)
could probably be linked to the modulation of the matrix
metalloprotein 9 (MMP9) expression. In fact, our results
demonstrated that the down-regulation of galectin-1 led to the
decrease of MMP9 expression. That the lectin is known as inducer of
MMP9 and other pro-degradative/inflammatory effectors via NF-κB
(42) and also promotes breast
cancer cell metastasis by upregulating MMP9 expression (43) flanks the assumption for a
functional correlation. Due to the possible involvement of other
proteins, the context dependence of these observations deserve
attention (44).
Anaplastic thyroid carcinoma is one of the most
aggressive human malignancies. Several studies had proposed
orthotopic models of human thyroid cancer for testing novel
therapeutic strategies (45–47).
We evaluated the impact of galectin-1 depletion on tumor growth in
an orthotopic nude mouse model of human anaplastic thyroid
carcinoma. Our in vivo experiments revealed reduced or lack
of tumor growth and lung metastasis, as well as prolonged survival,
in galectin-1 KD mice compared to control mice. These findings
support that galectin-1 directly affects thyroid cancer development
and may represent a suitable target for intervention in anaplastic
thyroid carcinoma.
In summary, our data showed that galectin-1 is a
good immunohistochemical marker that could meaningfully complement
information by monitoring galectin-3 and cytokeratin-19 in the
quest to discriminate benign thyroid nodules from malignant tumors.
Its observed involvement in cell proliferation, migration and
invasion in vitro, as well as in tumor growth in vivo
also support that galectin-1 plays functional roles in tumor
progression and metastasis formation. Impact on NF-κB or hedgehog
signaling or Rho GTPases can underlie these activities (32,48,49),
targeting counter-receptor cross-linking offering an opportunity to
block these protumoral routes (50). In aggregate, these data indicate an
association of galectin-1 and -3 expression with likely functional
cooperation, encouraging to further study the galectin network.
Acknowledgments
A.W. was granted by the Télévie (F. R. S.-FNRS,
Belgian National Fund for Scientific Research, 22877711). The study
received financial support from the 'Fonds pour l'Action de
Recherche Concertée' (ARC-14/19 UMONS 3) (Belgium). The Center for
Microscopy and Molecular Imaging (CMMI) is supported by the
European Regional Development Fund and Wallonia. We thank Benoît
Bauduin for his help in MRI (acquisition and processing of
images).
References
|
1
|
Pellegriti G, Frasca F, Regalbuto C,
Squatrito S and Vigneri R: Worldwide increasing incidence of
thyroid cancer: Update on epidemiology and risk factors. J Cancer
Epidemiol. 2013:9652122013.PubMed/NCBI
|
|
2
|
Goyal N, Setabutr D, Abdulghani J and
Goldenberg D: Molecular and genetic markers of follicular-cell
thyroid cancer: Etiology and diagnostic and therapeutic
opportunities. Adv Exp Med Biol. 779:309–326. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nikiforov YE: Molecular diagnostics of
thyroid tumors. Arch Pathol Lab Med. 135:569–577. 2011.PubMed/NCBI
|
|
4
|
Grogan RH, Mitmaker EJ and Clark OH: The
evolution of biomarkers in thyroid cancer-from mass screening to a
personalized biosignature. Cancers (Basel). 2:885–912. 2010.
View Article : Google Scholar
|
|
5
|
Cabanillas ME, Patel A, Danysh BP, Dadu R,
Kopetz S and Falchook G: BRAF inhibitors: Experience in thyroid
cancer and general review of toxicity. Horm Cancer. 6:21–36. 2015.
View Article : Google Scholar :
|
|
6
|
Kim KB, Cabanillas ME, Lazar AJ, Williams
MD, Sanders DL, Ilagan JL, Nolop K, Lee RJ and Sherman SI: Clinical
responses to vemurafenib in patients with metastatic papillary
thyroid cancer harboring BRAF(V600E) mutation. Thyroid.
23:1277–1283. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu Z, Li X, Shi L, Maimaiti Y, Chen T, Li
Z, Wang S, Xiong Y, Guo H, He W, et al: Cytokeratin 19,
thyroperoxidase, HBME-1 and galectin-3 in evaluation of aggressive
behavior of papillary thyroid carcinoma. Int J Clin Exp Med.
7:2304–2308. 2014.PubMed/NCBI
|
|
8
|
Flanagan JN, Pineda P, Knapp PE, De Las
Morenas A, Lee SL and Braverman LE: Expression of cytokeratin 19 in
the diagnosis of thyroid papillary carcinoma by quantitative
polymerase chain reaction. Endocr Pract. 14:168–174. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Krzeslak A, Gaj Z, Pomorski L and Lipinska
A: Expression of cytokeratin 19 in the cytosolic fraction of
thyroid lesions: ELISA and western blot analysis. Mol Med Rep.
1:565–569. 2008.PubMed/NCBI
|
|
10
|
Sumana BS, Shashidhar S and Shivarudrappa
AS: Galectin-3 immunohistochemical expression in thyroid neoplasms.
J Clin Diagn Res. 9:EC07–EC11. 2015.PubMed/NCBI
|
|
11
|
Bartolazzi A, Gasbarri A, Papotti M,
Bussolati G, Lucante T, Khan A, Inohara H, Marandino F, Orlandi F,
Nardi F, et al Thyroid Cancer Study Group: Application of an
immunodiagnostic method for improving preoperative diagnosis of
nodular thyroid lesions. Lancet. 357:1644–1650. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Inohara H, Honjo Y, Yoshii T, Akahani S,
Yoshida J, Hattori K, Okamoto S, Sawada T, Raz A and Kubo T:
Expression of galectin-3 in fine-needle aspirates as a diagnostic
marker differentiating benign from malignant thyroid neoplasms.
Cancer. 85:2475–2484. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gasbarri A, Martegani MP, Del Prete F,
Lucante T, Natali PG and Bartolazzi A: Galectin-3 and CD44v6
isoforms in the preoperative evaluation of thyroid nodules. J Clin
Oncol. 17:3494–3502. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Carpi A, Rossi G, Coscio GD, Iervasi G,
Nicolini A, Carpi F, Mechanick JI and Bartolazzi A: Galectin-3
detection on large-needle aspiration biopsy improves preoperative
selection of thyroid nodules: A prospective cohort study. Ann Med.
42:70–78. 2010. View Article : Google Scholar
|
|
15
|
Liu Z, Yu P, Xiong Y, Zeng W, Li X,
Maiaiti Y, Wang S, Song H, Shi L, Liu C, et al: Significance of
CK19, TPO, and HBME-1 expression for diagnosis of papillary thyroid
carcinoma. Int J Clin Exp Med. 8:4369–4374. 2015.PubMed/NCBI
|
|
16
|
Mataraci EA, Ozgüven BY and Kabukçuoglu F:
Expression of cytokeratin 19, HBME-1 and galectin-3 in neoplastic
and nonneoplastic thyroid lesions. Pol J Pathol. 63:58–64.
2012.PubMed/NCBI
|
|
17
|
Schmitt AC, Cohen C and Siddiqui MT:
Paired box gene 8, HBME-1, and cytokeratin 19 expression in
preoperative fine-needle aspiration of papillary thyroid carcinoma:
Diagnostic utility. Cancer Cytopathol. 118:196–202. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Niedziela M, Maceluch J and Korman E:
Galectin-3 is not an universal marker of malignancy in thyroid
nodular disease in children and adolescents. J Clin Endocrinol
Metab. 87:4411–4415. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mehrotra P, Okpokam A, Bouhaidar R,
Johnson SJ, Wilson JA, Davies BR and Lennard TWJ: Galectin-3 does
not reliably distinguish benign from malignant thyroid neoplasms.
Histopathology. 45:493–500. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mills LJ, Poller DN and Yiangou C:
Galectin-3 is not useful in thyroid FNA. Cytopathology. 16:132–138.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park YJ, Kwak SH, Kim DC, Kim H, Choe G,
Park DJ, Jang HC, Park SH, Cho BY and Park SY: Diagnostic value of
galectin-3, HBME-1, cytokeratin 19, high molecular weight
cytokeratin, cyclin D1 and p27(kip1) in the differential diagnosis
of thyroid nodules. J Korean Med Sci. 22:621–628. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu X, Sun T, Lu H, Zhou X, Lu Y, Cai X
and Zhu X: Diagnostic significance of CK19, RET, galectin-3 and
HBME-1 expression for papillary thyroid carcinoma. J Clin Pathol.
63:786–789. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Barroeta JE, Baloch ZW, Lal P, Pasha TL,
Zhang PJ and LiVolsi VA: Diagnostic value of differential
expression of CK19, Galectin-3, HBME-1, ERK, RET, and p16 in benign
and malignant follicular-derived lesions of the thyroid: An
immunohistochemical tissue microarray analysis. Endocr Pathol.
17:225–234. 2006. View Article : Google Scholar
|
|
24
|
Hirabayashi J: Recent topics on galectins.
Trends Glycosci Glycotechnol. 9:1–180. 1997.
|
|
25
|
Kaltner H and Gabius HJ: A toolbox of
lectins for translating the sugar code: The galectin network in
phylogenesis and tumors. Histol Histopathol. 27:397–416.
2012.PubMed/NCBI
|
|
26
|
Cludts S, Decaestecker C, Mahillon V,
Chevalier D, Kaltner H, André S, Remmelink M, Leroy X, Gabius HJ
and Saussez S: Galectin-8 up-regulation during hypopharyngeal and
laryngeal tumor progression and comparison with galectin-1, -3 and
-7. Anticancer Res. 29:4933–4940. 2009.
|
|
27
|
Remmelink M, de Leval L, Decaestecker C,
Duray A, Crompot E, Sirtaine N, André S, Kaltner H, Leroy X, Gabius
HJ, et al: Quantitative immunohistochemical fingerprinting of
adhesion/growth-regulatory galectins in salivary gland tumours:
Divergent profiles with diagnostic potential. Histopathology.
58:543–556. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dawson H, André S, Karamitopoulou E,
Zlobec I and Gabius HJ: The growing galectin network in colon
cancer and clinical relevance of cytoplasmic galectin-3 reactivity.
Anticancer Res. 33:3053–3059. 2013.PubMed/NCBI
|
|
29
|
Toegel S, Bieder D, André S, Kayser K,
Walzer SM, Hobusch G, Windhager R and Gabius HJ: Human
osteoarthritic knee cartilage: Fingerprinting of
adhesion/growth-regulatory galectins in vitro and in situ indicates
differential upregulation in severe degeneration. Histochem Cell
Biol. 142:373–388. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Saussez S, Decaestecker C, Cludts S,
Ernoux P, Chevalier D, Smetana K Jr, André S, Leroy X and Gabius
HJ: Adhesion/growth-regulatory tissue lectin galectin-1 in relation
to angiogenesis/lymphocyte infiltration and prognostic relevance of
stromal up-regulation in laryngeal carcinomas. Anticancer Res.
29:59–65. 2009.PubMed/NCBI
|
|
31
|
Amano M, Eriksson H, Manning JC, Detjen
KM, André S, Nishimura S, Lehtiö J and Gabius HJ: Tumour suppressor
p16(INK4a) - anoikis-favouring decrease in N/O-glycan/cell surface
sialylation by down-regulation of enzymes in sialic acid
biosynthesis in tandem in a pancreatic carcinoma model. FEBS J.
279:4062–4080. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Smetana K Jr, André S, Kaltner H, Kopitz J
and Gabius HJ: Context-dependent multifunctionality of galectin-1:
A challenge for defining the lectin as therapeutic target. Expert
Opin Ther Targets. 17:379–392. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chiariotti L, Berlingieri MT, Battaglia C,
Benvenuto G, Martelli ML, Salvatore P, Chiappetta G, Bruni CB and
Fusco A: Expression of galectin-1 in normal human thyroid gland and
in differentiated and poorly differentiated thyroid tumors. Int J
Cancer. 64:171–175. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu XC, el-Naggar AK and Lotan R:
Differential expression of galectin-1 and galectin-3 in thyroid
tumors. Potential diagnostic implications. Am J Pathol.
147:815–822. 1995.PubMed/NCBI
|
|
35
|
Salajegheh A, Dolan-Evans E, Sullivan E,
Irani S, Rahman MA, Vosgha H, Gopalan V, Smith RA and Lam AK: The
expression profiles of the galectin gene family in primary and
metastatic papillary thyroid carcinoma with particular emphasis on
galectin-1 and galectin-3 expression. Exp Mol Pathol. 96:212–218.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Saussez S, Glinoer D, Chantrain G, Pattou
F, Carnaille B, André S, Gabius HJ and Laurent G: Serum galectin-1
and galectin-3 levels in benign and malignant nodular thyroid
disease. Thyroid. 18:705–712. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kaltner H, Seyrek K, Heck A, Sinowatz F
and Gabius HJ: Galectin-1 and galectin-3 in fetal development of
bovine respiratory and digestive tracts. Comparison of cell
type-specific expression profiles and subcellular localization.
Cell Tissue Res. 307:35–46. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
39
|
Kopitz J, Vértesy S, André S, Fiedler S,
Schnölzer M and Gabius HJ: Human chimera-type galectin-3: Defining
the critical tail length for high-affinity glycoprotein/cell
surface binding and functional competition with galectin-1 in
neuroblastoma cell growth regulation. Biochimie. 104:90–99. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
de Matos LL, Del Giglio AB, Matsubayashi
CO, de Lima Farah M, Del Giglio A and da Silva Pinhal MA:
Expression of CK-19, galectin-3 and HBME-1 in the differentiation
of thyroid lesions: Systematic review and diagnostic meta-analysis.
Diagn Pathol. 7:972012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dunđerović D, Lipkovski JM, Boričic I,
Soldatović I, Božic V, Cvejić D and Tatić S: Defining the value of
CD56, CK19, Galectin 3 and HBME-1 in diagnosis of follicular cell
derived lesions of thyroid with systematic review of literature.
Diagn Pathol. 10:1962015. View Article : Google Scholar
|
|
42
|
Toegel S, Weinmann D, André S, Walzer SM,
Bilban M, Schmidt S, Chiari C, Windhager R, Krall C, Bennani-Baiti
IM, et al: Galectin-1 couples glycobiology to inflammation in
osteoarthritis through the activation of an NF-κB-regulated gene
network. J Immunol. 196:1910–1921. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhu X, Wang K, Zhang K, Xu F, Yin Y, Zhu L
and Zhou F: Galectin-1 knockdown in carcinoma-associated
fibroblasts inhibits migration and invasion of human MDA-MB-231
breast cancer cells by modulating MMP-9 expression. Acta Biochim
Biophys Sin (Shanghai). 48:462–467. 2016. View Article : Google Scholar
|
|
44
|
Saussez S, Cludts S, Capouillez A,
Mortuaire G, Smetana K Jr, Kaltner H, André S, Leroy X, Gabius HJ
and Decaestecker C: Identification of matrix metalloproteinase-9 as
an independent prognostic marker in laryngeal and hypopharyngeal
cancer with opposite correlations to adhesion/growth-regulatory
galectins-1 and -7. Int J Oncol. 34:433–439. 2009.PubMed/NCBI
|
|
45
|
Nucera C, Nehs MA, Mekel M, Zhang X, Hodin
R, Lawler J, Nose V and Parangi S: A novel orthotopic mouse model
of human anaplastic thyroid carcinoma. Thyroid. 19:1077–1084. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sewell W, Reeb A and Lin RY: An orthotopic
mouse model of anaplastic thyroid carcinoma. J Vis Exp. 74:2013.
View Article : Google Scholar
|
|
47
|
Antonello ZA and Nucera C: Orthotopic
mouse models for the preclinical and translational study of
targeted therapies against metastatic human thyroid carcinoma with
BRAF(V600E) or wild-type BRAF. Oncogene. 33:5397–5404. 2014.
View Article : Google Scholar
|
|
48
|
Camby I, Belot N, Lefranc F, Sadeghi N, de
Launoit Y, Kaltner H, Musette S, Darro F, Danguy A, Salmon I, et
al: Galectin-1 modulates human glioblastoma cell migration into the
brain through modifications to the actin cytoskeleton and levels of
expression of small GTPases. J Neuropathol Exp Neurol. 61:585–596.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Martínez-Bosch N, Fernández-Barrena MG,
Moreno M, Ortiz-Zapater E, Munné-Collado J, Iglesias M, André S,
Gabius HJ, Hwang RF, Poirier F, et al: Galectin-1 drives pancreatic
carcinogenesis through stroma remodeling and Hedgehog signaling
activation. Cancer Res. 74:3512–3524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Solís D, Bovin NV, Davis AP,
Jiménez-Barbero J, Romero A, Roy R, Smetana K Jr and Gabius HJ: A
guide into glycosciences: How chemistry, biochemistry and biology
cooperate to crack the sugar code. Biochim Biophys Acta.
1850:186–235. 2015. View Article : Google Scholar
|