Introduction
Colorectal cancer (CRC), one of the most commonly
registered cancers worldwide, is associated with high mortality,
especially for advanced and metastatic patients (1). Although enormous efforts have been
made in both basic and clinical research in the fight against CRC,
there are more than one million new cases per year in the world. It
is estimated that approximately 95,520 (colon cancer only) new
cases and 50,260 (colon and rectal cancers combined) deaths from
CRC will occur in the United States in 2017 (2,3).
Although surgery can cure approximately half of the patients, it is
reported that disease recurrence is nearly 50% among patients with
CRC who received resection (4).
Aberrant angiogenesis is an essential step in the
progression of CRC, which provides nutrients and oxygen for the
survival, growth and metastasis of the tumor cells (5). Anti-angiogenic therapy has been
proved to be one of the most crucial and promising approaches to
control tumor growth (6). The
combination of anti-angiogenic therapy and chemotherapy has been
well accepted as a first-line regimen for metastatic CRC (7).
Ginsenoside, Rg3, one of the major active components
of ginseng, displays anti-angiogenesis ability (5,8) and
has been widely used in traditional medicine. Rg3 has been reported
to exhibit antitumor effects in several types of cancer, including
ovarian, breast and lung cancer (9,10).
Rg3 also repressed migration and induced apoptosis in CRC cells
in vitro (9,10).
Therefore, in the present study, we attempted to
verify whether Rg3 could be applied to the treatment of CRC in
orthotopic xenograft models; the mechanisms involved were also
investigated.
Materials and methods
Cell line and cultures
The human CRC cell lines, LoVo, SW620 and HCT116,
were purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA). Cells were maintained in Dulbecco's modified
Eagle's medium (DMEM; Gibco, Grand Island, NY, USA) supplemented
with 10% fetal calf serum (FCS; Hyclone Laboratories, Inc., Logan,
UT, USA), 100 U/m penicillin and 100 mg/ml streptomycin. The
cultures were incubated at 37°C in a humidified atmosphere with 5%
CO2. Cells were passaged every 2–3 days to obtain
exponential growth.
Reagents
Rg3 was purchased from Shanghai Jinsui
Bio-Technology Co., Ltd., (Shanghai, China), fluorouracil (5-FU)
was purchased from Shanghai Xudong Haipu Pharmaceutical Co., Ltd.,
(Shanghai, China) and oxaliplatin was purchased from Jiangsu
Hengrui Medicine Co., Ltd. (Lianyungang, China).
MTT assay
Cellular growth was evaluated by an MTT (methyl
thiazolyl-tetrazolium) assay (11). Cells were seeded into 24-well
tissue culture plates at 5×104 cells/well. After
treatment, MTT (Sigma-Aldrich, St. Louis, MO, USA) was added to
each well to a final concentration of 0.5 mg/ml, followed by
incubation at 37°C for 4 h. The medium was then removed, and 800
μl of dimethyl sulfoxide (DMSO) was added per well. The
absorbance in each well was measured at 490 nm using a microplate
enzyme-linked immunosorbent assay (ELISA) reader (Bio-Rad
Laboratories, Hercules, CA, USA). The relative cell viability was
calculated as follows: Relative cell viability = (mean experimental
absorbance/mean control absorbance) × 100%.
Wound healing assay
Cells (1×104/well) were seeded in 96-well
plates and grown to confluence. The monolayer culture was
artificially scraped/wounded with a sterile micropipette tip to
create a denuded zone of constant width. Each well was washed with
phosphate-buffered saline (PBS) twice to remove the detached cells.
Cell migration to the wounded region was observed using an XDS-1B
inverted microscope (MIC Optical and Electrical Instrument,
Chongqing, China) and photographed (×40 magnification). Images were
captured at 0, 3, 6, 9 and 12 h to monitor the wound healing
process. The wound areas were measured using ImageJ (NIH, Bethesda,
MA, USA).
Evaluation of protein levels
The protein levels of CD24, CD44 and EpCAM in colon
cancer cells were measured by flow cytometry. Following treatment,
the cells were harvested, fixed with 4% paraformaldehyde and were
permeabilized using 0.1% Triton X-100. After washing with PBS three
times, cells were incubated with anti-CD24 (FITC-conjugated; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA), anti-CD44
(PE-conjugated; Santa Cruz Biotechnology), and anti-EpCAM
(PerCP-cy5.5-conjugated; Santa Cruz Biotechnology) antibodies,
respectively, for 30 min at 4°C. Subsequently, the cells were
analyzed using a Beckman Coulter FC500 flow cytometer (Beckman
Coulter, Indianapolis, IN, USA).
Stemness evaluation of cancer cells
Cells were harvested, washed twice with 2% fetal
bovine serum (FBS)/PBS solution, and resuspended in 100 μl
of 2% FBS/PBS, before incubating with anti-CD24 (FITC-conjugated),
anti-CD44 (PE-conjugated), and anti-EpCAM (PerCP-cy5.5-conjugated)
antibodies for 30 min at 4°C. Cells were then washed twice with 2%
FBS/PBS solution, and resuspended in 500 μl of 2% FBS/PBS
before assessment on a Beckman Coulter FC500 flow cytometer
(Beckman Coulter).
Plate clone formation assay
LoVo and HCT116 cells were used in this assay. LoVo
cells were seeded at a density of 250 cells/well and HCT116 cells
were seeded at a density of 500 cells/well in 24-well plates and
treated with different concentrations of Rg3 3 days later. After
treatment for 12 days, the cells were stained with 1%
methylrosanilinium chloride and the numbers of visible colonies
were counted. The relative clone formation ability was calculated
as relative clone formation ability = (mean experimental clone
number/mean control clone number) × 100%.
Real-time PCR
Total RNA was extracted using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA), according to the manufacturer's
protocol. After spectrophotometric quantification, 1 μg of
total RNA in a final volume of 20 μl was used for reverse
transcription with a PrimeScript RT Reagent kit (Takara, Bio,
Shiga, Japan), according to the manufacturer's protocol. Aliquots
of cDNA corresponding to equal amounts of RNA were used to quantify
the mRNA by real-time PCR using the LightCycler 96 Real-time
Quantitative PCR Detection system (Roche Diagnostics, Indianapolis,
IN, USA). The reaction system (25 μl) contained the
corresponding cDNA, forward and reverse primers, and the SYBR-Green
PCR Master Mix (Roche Diagnostics). All data were analyzed using
the expression of the B2M gene as an internal standard. The
specific primers are presented in Table I.
 | Table IPrimers used in the present
study. |
Table I
Primers used in the present
study.
| Genes | Sense (5′–3′) | Antisense
(5′–3′) | Product size
(bp) |
|---|
| Cancer stem cell
markers |
| CD24 |
CAGGGCAATGATGAATGAGAAT |
CCTGGGCGACAAAGTGAGA | 233 |
| CD44 |
GTGATGGCACCCGCTATGTC |
AACCTCCTGAAGTGCTGCTCC | 129 |
| EpCAM |
TAATCGTCAATGCCAGTGTACTTC |
GCCATTCATTTCTGCCTTCAT | 100 |
|
Angiogenesis-related genes |
| ANG |
CAAGAATGGAAACCCTCACAGA |
AAATGGAAGGCAAGGACAGC | 246 |
| ANGPT1 |
AGAGGTCAGAAGAAAGGAGCAAG |
GTGAGTCAGAATGGCAGCGAG | 109 |
| ANGPT2 |
AGAGGAACAAAGGACCGTGAAAG |
CTGTCAGATTGCAGTGGGAAG | 91 |
| CCL1 |
TGGATGGGTTCAGAGGCAC |
GCAGGGCAGAAGGAATGGT | 147 |
| CCL13 |
AGGAGAAGTGGGTCCAGAATTAT |
CTCAAATAAACTCCAAACCAGCAAC | 265 |
| CCL5 |
GAGAAGAAATGGGTTCGGGAGT |
AGGACAAGAGCAAGCAGAAACAGGC | 109 |
| CCL7 |
GCTCAGCCAGTTGGGATTAAT |
TCATGGCTTGTTTTCAGTTCAGTC | 164 |
| COL18A |
TCAGACCACGGCTCGATTTC |
CTCAGCTCCCATTGCCTCA | 154 |
| CSF3 |
CCTTCGCCTCTGCTTTCCA |
CGTTCTGCTCTTCCCTGTCTTT | 199 |
| CXCL1 |
CACCCCAAGAACATCCAAAGT |
CCTTCAGGAACAGCCACCA | 210 |
| CXCL2 |
GCTTATTGGTGGCTGTTCCTG |
ACACATTAGGCGCAATCCAG | 101 |
| CXCL3 |
GCCCAAACCGAAGTCATAGC |
GAACCCTCGTAAGAAATAGTCAAAC | 271 |
| CXCL5 |
ACAGTGCCCTACGGTGGAAGT |
CTCATCAAAGCAGGGAGTTCATA | 266 |
| EGF |
GGGTGACCGTTTGGGAGTT |
ATCCACCACGTCGTCCATG | 335 |
| PLG |
GACTATCTGGTTTGTGGATGCGT |
TTCTTCGTCCTCCTCACATTTT | 201 |
| FGF-2 |
CTGTCTGGTTTGCTGCTGTATCT |
GGTTTCTGGGATTTGCTTTATTC | 95 |
| FIGF |
CATCCCATCGGTCCACTAGGT |
CAGCCACCACATCGGAACA | 190 |
| FLT4 |
GCTGTGCCTGCGACTGTG |
CGTGTCCTCGCTGTCCTTGT | 138 |
| GM-CSF |
ACACTGCTGCTGAGATGAATGA |
AAAGGTGATAATCTGGGTTGCA | 218 |
| IFNG |
TCCAACGCAAAGCAATACATG |
TTGCAGGCAGGACAACCAT | 137 |
| IGF1 |
GGTGGATGCTCTTCAGTTCGT |
GCAATACATCTCCAGCCTCCTTAG | 182 |
| IL10 |
TGGTGAAGGAGGATCGCTAGA |
CCTTGATGTCTGGGTCTTGGTT | 204 |
| IL1A |
TGACGACGCACTTGTAGCCAC |
GCCAATGAAATGACTCCCTCT | 111 |
| IL1B |
ATTTGAGTCTGCCCAGTTCCC |
AACCTTTCTGTTCCCTTTCTGC | 207 |
| IL2 |
CAGTAACCTCAACTCCTGCCAC |
CTGGTGAGTTTGGGATTCTTGTA | 227 |
| IL4 |
CCCCTCTGTTCTTCCTGCTAG |
TGTCCTTCTCATGGTGGCTGT | 181 |
| IL8 |
CTGGGTGCAGAGGGTTGTG |
ACTGGCATCTTCACTGATTCTTG | 98 |
| KDR |
CCCAATAATCAGAGTGGCAGTG |
CATAGACATAAATGACCGAGGCC | 163 |
| MMP1 |
GCTGAAAGTGACTGGGAAACC |
TCTTGGCAAATCTGGCGTGT | 166 |
| PDFGB |
GCTGTTGAGGTGGCTGTAGATG |
GTCGTGGCTGGGTTGGAAT | 281 |
| PECAM1 |
AGGTCAGCAGCATCGTGGT |
GTGAAGTTGGCTGGAGGTG | 136 |
| PGF |
AAGGGAGCTGCTGTCTGCG |
CTTGCGGAGTCAGGAGCCCGTAGGT | 192 |
| PIGF |
ACTGTGCCTTGCTTATGTTTGTT |
CCAAGCCATGCTCCTACAAAG | 137 |
| PLAUR |
GCCGGGCTGTCACCTATT |
CCACATCCAGGCACTGTTCTTC | 132 |
| TEK |
TAACTATGACTGTGGACAAGGGAG |
GGCCGAGGTGAAGAGGTTT | 221 |
| TGFB1 |
CTGGCGATACCTCAGCAACC |
CTAAGGCGAAAGCCCTCAAT | 126 |
| THPO |
TCTCAGACACTGCCGACATCA |
GGGCTTTGGGTTTCAGGAGA | 112 |
| TIMP1 |
GGTTGTGGGACCTGTGGAAGTA |
CCAAGATGTATAAAGGGTTCCAAG | 108 |
| TIMP2 |
CCCCTGTTCGCTTCCTGTATG |
GCGTTCCACTCTGGGTCAAAT | 207 |
| TPO |
AAGCAAGCGCCTGGTGGA |
CAGGAAGTTTGGAAAAAGACAGAAG | 156 |
| VEGFA |
CACCCACCCACATACATACATTT |
CCTCCCAACTCAAGTCCACAG | 170 |
| Internal
control |
| B2M |
TCAAGAAGGTGGTGAAGCAG |
AAGGTGGAGGAGTGGGTGTC | 112 |
Nude mouse orthotopic tumor xenograft
model and treatments
Four-week-old female BALB/c athymic nude mice were
purchased from Shanghai SLAC Laboratory Animal Co., Ltd.,
(Shanghai, China) and received humane care according to the Soochow
University Institutional Animal Care and Treatment Committee.
The orthotopic xenograft model was established as
follows: Cells were injected into the left flanks of the mice in a
total volume of 100 μl (0.5×107 cells),
separately. Ten days later, when the tumors were palpable, the
subcutaneous xenograft tumors were harvested and used to establish
the orthotopic xenograft model, according to previous reports
(12). In brief, the tumors were
harvested and cut into pieces (1.5 mm in diameter). A small midline
incision was made and the sigmoideum part of the intestine was
exteriorized. Serosa of the site where the tumor pieces were to be
implanted was removed using an amyxis. Single pieces of the tumor
were then implanted into the wall of the sigmoideum and fixed with
surgical sutures. The intestine was returned to the abdominal
cavity, and the abdominal wall was closed with surgical sutures.
The animals were kept in a sterile environment. To validate the
success of the orthotopic xenograft, exploratory laparotomy was
performed on 5 randomly chosen mice 7 days after tumor
inoculation.
The mice were observed and weighed once per day
during the study period. After 12 days, all mice were euthanized
and the tumors were carefully resected and weighed. The tumor
tissue was then formalin-fixed, paraffin-embedded, cut into
4-μm sections and immunohistochemically stained.
Immunohistochemistry
The resection specimens were fixed in 10% buffered
formalin and paraffin-embedded by routine processing. Sections were
cut at a thickness of 4 μm, heated at 60°C for 30 min, and
then deparaffinized and hydrated through a series of xylene and
alcohol baths. The slides were microwaved in antigen retrieval
solution (citrate buffer, pH 6.0, containing 0.3% trisodium citrate
and 0.04% citric acid) for 5 min. After replenishment of this
solution, the slides were microwaved again for 5 min, and then
allowed to cool for 20 min. The sections were then rinsed in PBS
and immersed in 3% H2O2 for 15 min to block
the endogenous peroxidase activity. Thereafter, the sections were
incubated with 10% bovine serum albumin (BSA) at room temperature
for 1 h to block non-specific antibodies. Immunohistochemical
staining was performed using mouse anti-Ki-67 antibody (GM724029;
GeneTech, Shanghai, China), rabbit anti-CD24 antibody (ab202963;
Abcam,Cambridge, UK), rabbit anti-CD44 antibody (ab157107; Abcam),
rabbit anti-EpCAM antibody (ab71916; Abcam), rabbit anti-B7-H1
antibody (ab205921; Abcam), mouse anti-B7-H3 antibody (ab105922;
Abcam and rabbit anti-CD34 antibody (ab81289; Abcam) at room
temperature for 2 h. After incubation with the corresponding
secondary antibodies for 20 min, the bound complexes were
visualized using a SuperPicture Polymer Detection kit (no. 87-8963;
Invitrogen).
Angiogenesis vascularity evaluation
Angiogenesis vascularity was defined as the number
of vessels per field counted in the area of the highest vascular
density, termed as microvessel density (MVD). Endothelial cells
were marked with an anti-CD34 antibody. The CD34 antigen was
localized in the cytoplasm and cellular membrane of vascular
endothelial cells. Single endothelial cells, endothelial cell
clusters, and microvessels in the tumors, clearly separated from
adjacent microvessels, were counted. Peritumoral vascularity and
vascularity in areas of necrosis were not scored. A vascular lumen
was not a requirement for a structure to be counted as a
microvessel. Branching structures were counted as one, unless there
was a break in the continuity of the vessel, in which case it was
counted as two distinct vessels. Areas with a higher density of
CD34+ cells and cell clusters relative to adjacent areas
were classified as 'hot spots'. The slides were initially screened
at low power to identify the areas with the highest number of
microvessels or vascularity hot spots. Microvessels were counted in
×400 magnification fields. MVD was defined as the number of
manually counted vessel profiles per mm2, taken as the
average from three hot-spot counts.
Collection of human CRC tissues
The study material was obtained from 129 patients
with metastatic CRC whose tissue samples were available (mean age
62 years, range 26–82 years) and who were treated from January 2007
to July 2016 at the First Affiliated Hospital of Soochow
University. Patient characteristics are detailed in Table II. All human tissue samples were
obtained and handled in accordance with an approved Institutional
Review Board application (the Committee on Medical Ethics, the
First Affiliated Hospital of Soochow University). Prognostic
analyses were performed regarding overall survival (OS).
| Table IIClinicopathological features of 129
patients with metastatic CRC. |
Table II
Clinicopathological features of 129
patients with metastatic CRC.
| Clinicopathological
features | n | B7H3
| B7H1
|
|---|
| Low (n) | High (n) | χ2 | P-value | Low (n) | High (n) | χ2 | P-value |
|---|
| Sex |
| Male | 53 | 28 | 25 | 0.574 | 0.449 | 23 | 30 | 1.066 | 0.302 |
| Female | 76 | 35 | 41 | | | 40 | 36 | | |
| Age (years) |
| >56 | 62 | 35 | 27 | 2.770 | 0.096 | 32 | 30 | 0.368 | 0.544 |
| ≤56 | 67 | 28 | 39 | | | 31 | 36 | | |
| BMI |
| >25 | 65 | 32 | 33 | 0.008 | 0.928 | 28 | 37 | 1.740 | 0.187 |
| ≤25 | 64 | 31 | 33 | | | 35 | 29 | | |
| Liver
metastasis |
| No | 51 | 31 | 20 | 4.818 | 0.028 | 34 | 17 | 10.731 | 0.001 |
| Yes | 78 | 32 | 46 | | | 29 | 49 | | |
Statistical analysis
Each experiment was performed at least in
triplicate. The results are expressed as the mean ± standard
deviation. Kaplan-Meier curves were constructed, and the
statistical analysis was carried out using the log-rank test. OS
was defined as the time from the diagnosed date to the time of
death from any cause. Statistical analysis was performed using an
unpaired Student's t-test. P<0.05 was considered
significant.
Results
Rg3 suppresses the growth and migration
of CRC cells in vitro
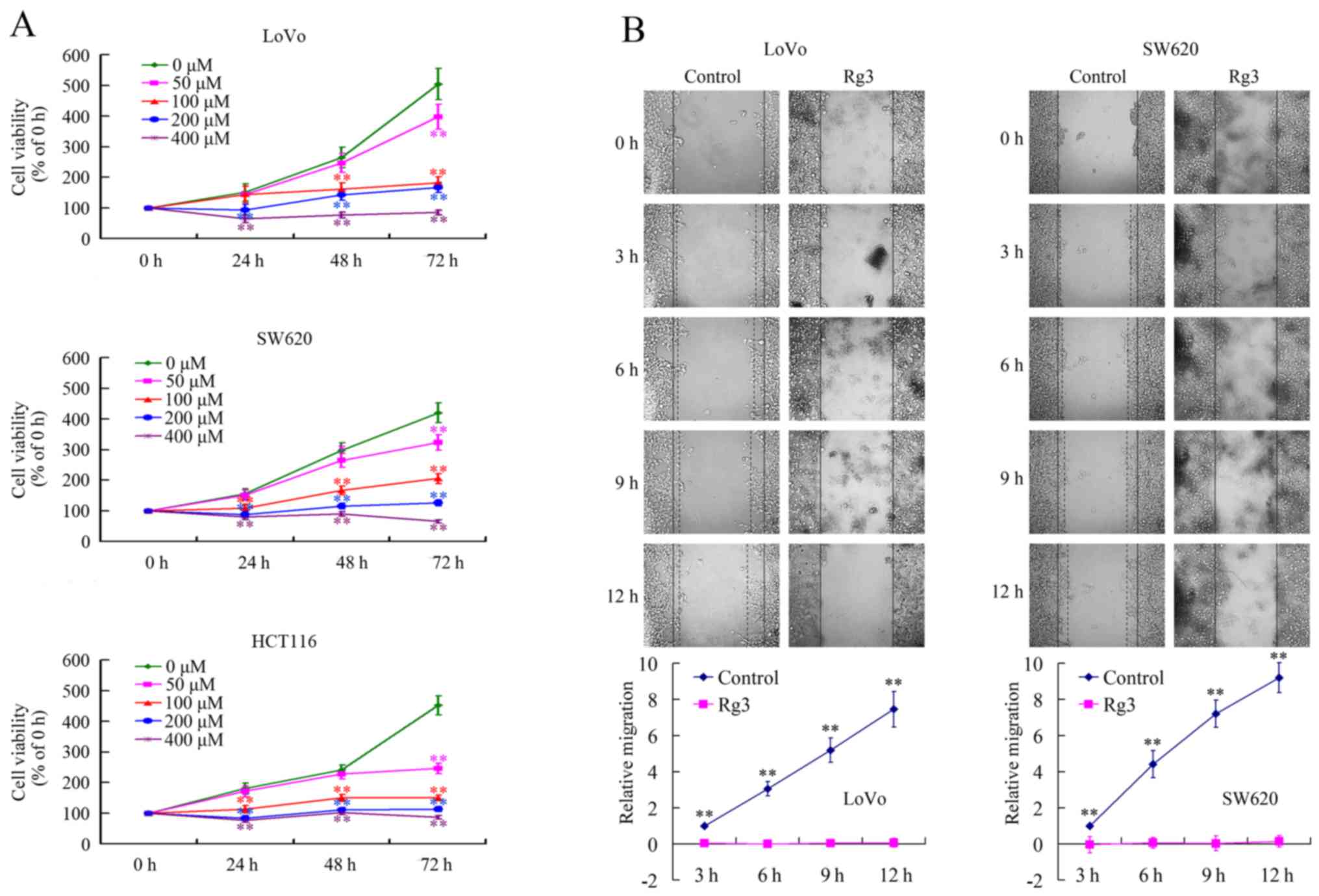
To investigate the effects of Rg3 on the biological
behavior of CRC cells, MTT and wound healing assays were performed
to evaluate cell growth and migration in vitro. The
proliferation of LoVo, SW620, and HCT116 cells was all
significantly inhibited by Rg3 in time- and dose-dependent manner
(Fig. 1A). The wound healing assay
verified that Rg3 inhibited migration of both LoVo and SW620 cells
remarkably (Fig. 1B). These in
vitro data confirmed that Rg3 had antitumor effects against CRC
cells.
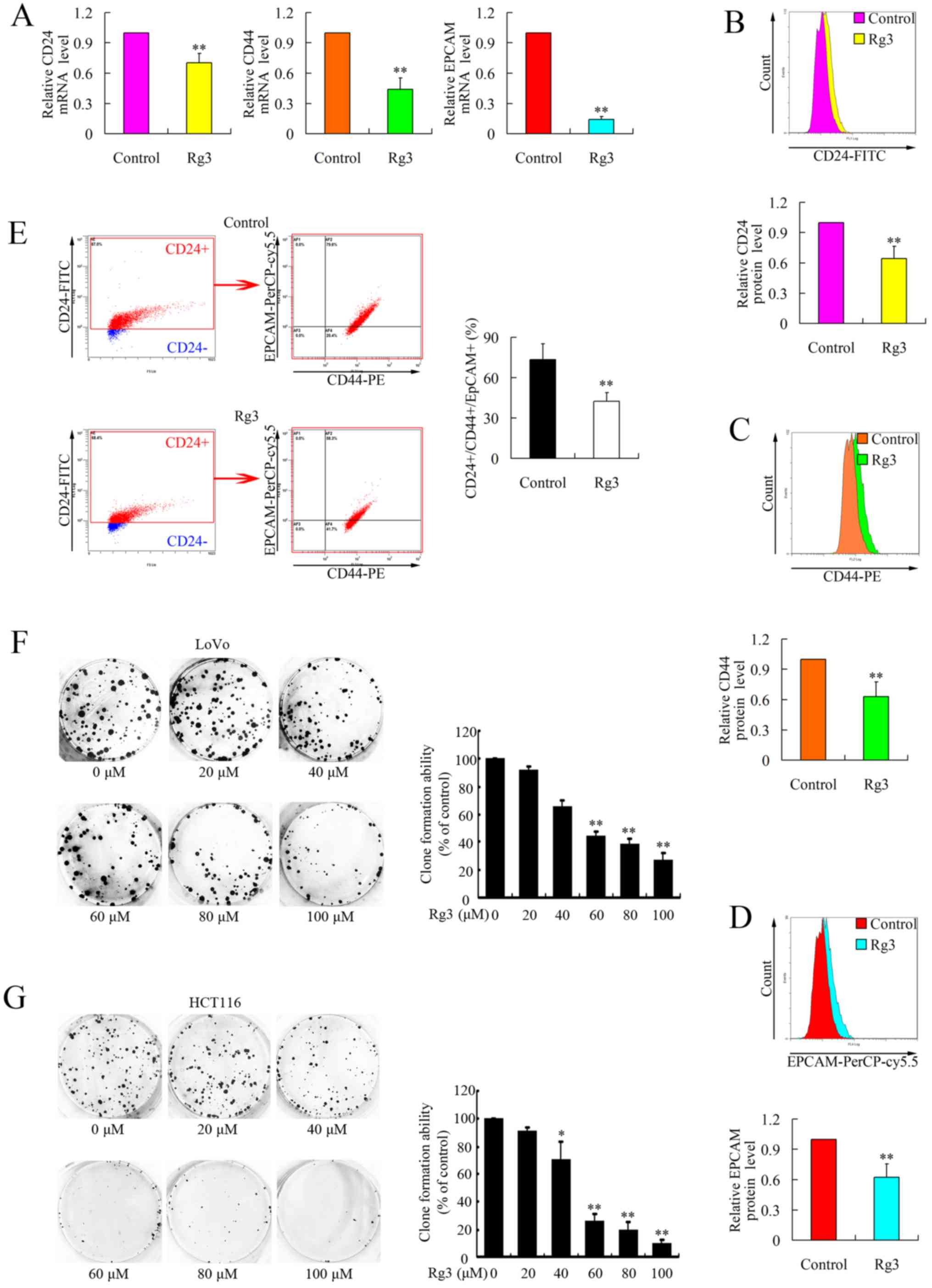
Rg3 impairs the stemness of CRC cells in
vitro
It has been well accepted that there is a special
subgroup of cancer cells in tumors, termed cancer stem cells
(CSCs), which have been proven to preserve the abilities of
extensive proliferation, self-renewal, multi-lineage
differentiation, drug-resistance, high metastasis and high
tumorigenic potential (13–16).
CD24, CD44 and EpCAM are the well accepted
colorectal CSC markers (13–16).
To investigate whether Rg3 could affect the stemness of CRC cells,
we examined the expressions of CD24, CD44 and EpCAM at both the
mRNA and protein levels using real-time PCR and flow cytometry. As
shown in Fig. 2A–D, the expression
of these three markers was downregulated after treatment with Rg3.
We then evaluated the positivity of CD24, CD44 and EpCAM on the
surface of CRCs. The proportion of
CD24+/CD44+/EpCAM+ cells decreased
significantly (Fig. 2E),
suggesting the repressed stemness of CRCs after Rg3 treatment.
In addition, we further confirmed the stemness of
the cells using a plate clone formation assay. LoVo and HCT116 were
treated with Rg3 at different low doses and clone formation ability
was then evaluated by calculating the visible clones. As shown in
Fig. 2F and G, Rg3 treatment
caused significant dose-dependent inhibition of the clone formation
ability of CRC cells, which was consistent with the downregulation
of CSC markers.
Rg3 represses growth and stemness of CRC
in vivo
We used an established orthotopic xenograft model to
evaluate the antitumor effect of Rg3 in vivo. Treatment with
Rg3 (25 mg/kg) for 12 consecutive days significantly repressed the
growth of the tumors (Fig. 3A and
B). As shown in Fig. 3C and D,
Rg3 downregulated the Ki-67 level, a major prognostic factor of
tumors in general (17).
Immunohistochemical assays further confirmed that Rg3 treatment
downregulated the levels of stemness markers, CD24, CD44 and EpCAM,
in the orthotopic xenografts (Fig.
3E–J), which was consistent with the in vitro data.
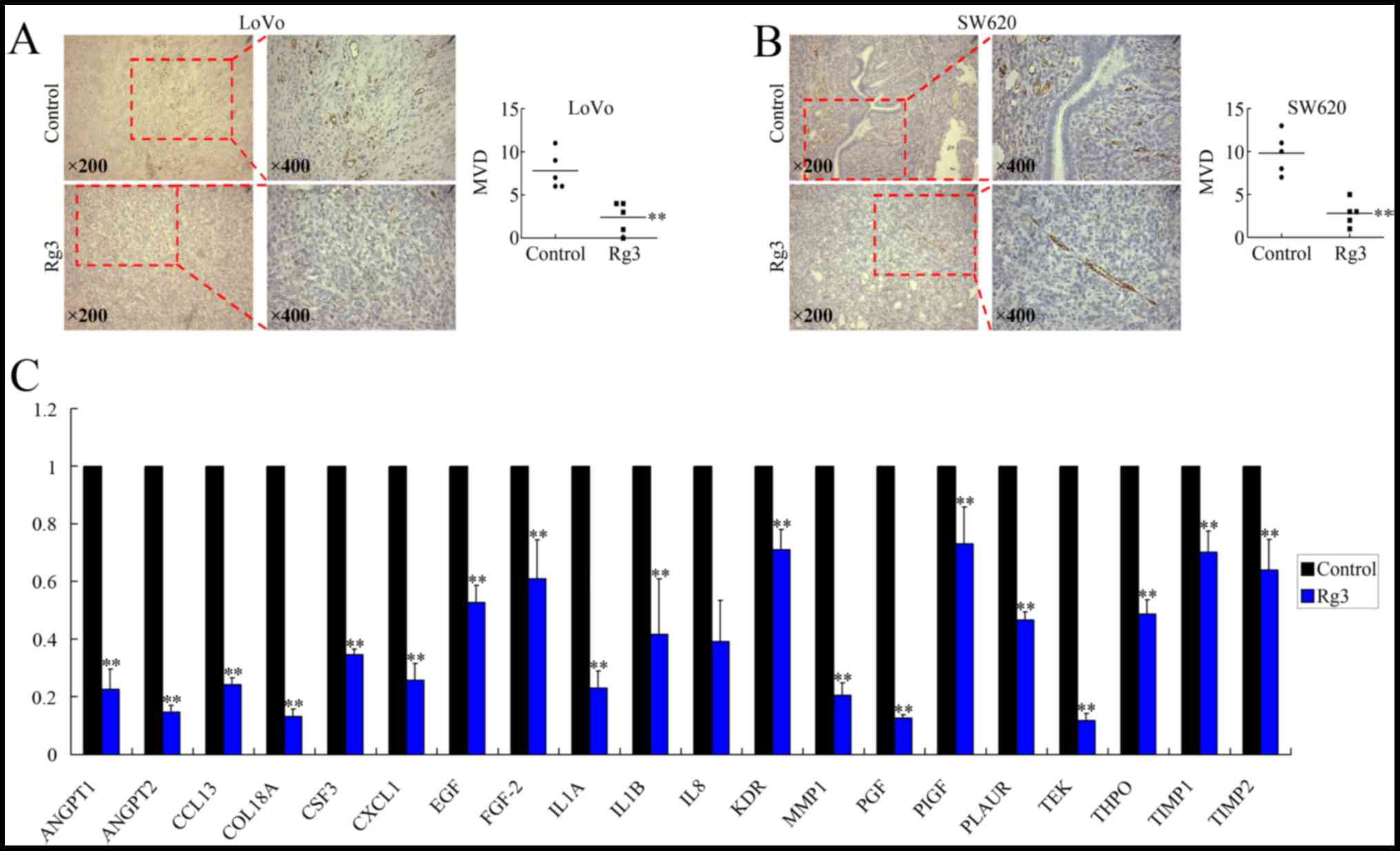
Rg3 represses angiogenesis of CRC
Rg3 is believed to inhibit angiogenesis in tumors;
therefore, we investigated whether Rg3 could also affect
vascularization of the CRC orthotopic xenografts. Endothelial cells
in the tissue were positively stained using an anti-CD34 antibody
(Fig. 4A). The results showed that
treatment with Rg3 repressed angiogenesis, according to the
microvessel density (MVD) levels (Fig.
4B).
To investigate the mechanisms involved in
Rg3-repressed angiogenesis, we analyzed the expression of 41
angiogenesis-related genes (Table
I) using real-time PCR. Among these genes, although ANG,
CCL1 and CXCL5 were found to be upregulated (data not
shown), 22 pro-angiogenic genes, including ANGPT1,
ANGPT2, CCL13, COL18A, CSF3,
CXCL1, EGF, FGF-2, IL1A, IL1B,
IL8, KDR, MMP1, PGF, PIGF,
PLAUR, TEK, THPO, TIMP1 and
TIMP2 were downregulated at the mRNA level after treatment
with Rg3 (Fig. 4C). The
expressions of the other 18 genes were unchanged after treatment
with Rg3 (data not shown). Therefore, it is possible that the
anti-angiogenic effect of Rg3 is executed by downregulating the
expression of certain angiogenesis-related genes.
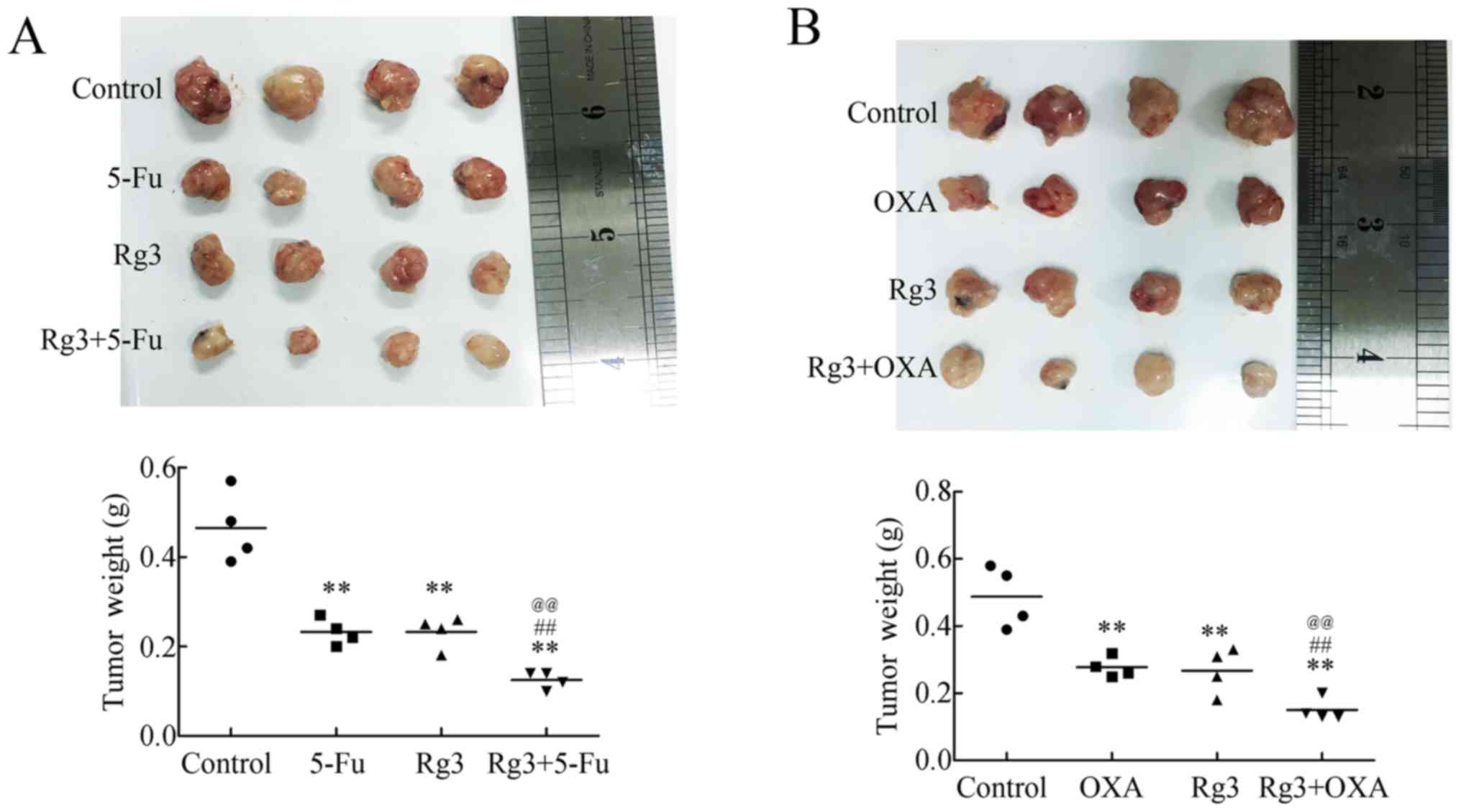
Rg3 strengthens the cytotoxicity of 5-Fu
and oxaliplatin in vivo
Rg3 impaired the stemness of CRC cells and repressed
angiogenesis; therefore, we next investigated whether Rg3 could
improve the cytotoxicity of 5-Fu and oxaliplatin, two widely used
first line pharmacotherapeutics in clinical treatments.
Separately, Rg3, 5-Fu and oxaliplatin could repress
growth of the xenografts (Fig. 5).
However, the combinations of Rg3 and 5-Fu, or Rg3 and oxaliplatin,
decreased the volumes of the tumors to much more significant
levels. Furthermore, the weights of the tumors were also consistent
with the images (Fig. 5),
suggesting that Rg3 could act as a supplement for chemotherapy
regimens containing 5-Fu or oxaliplatin.
Rg3 downregulates the levels of B7-H1 and
B7-H3, predictors of adverse clinical outcomes in CRC
B7-H1 and B7-H3 belong to B7 family, and play
important roles in tumor immune responses by integrating T cell
receptor signaling to regulate T cell function. To confirm the
relationship between outcomes of CRC patients and the levels of
B7-H1 and B7-H3, we collected tissue samples from 129 patients with
metastatic CRC, and assessed the levels of B7-H1 and B7-H3 using
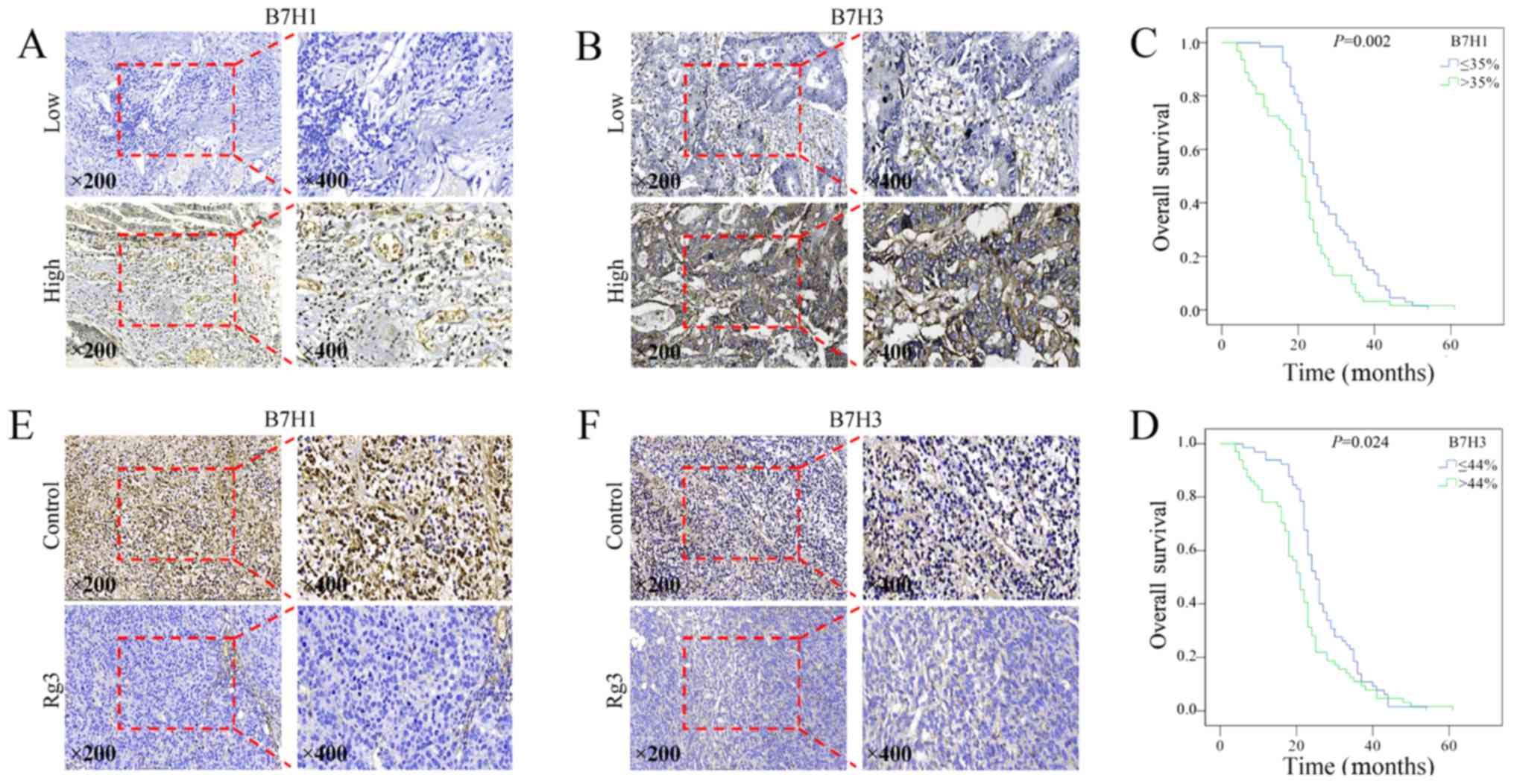
immunohistochemistry. The representative microscope images of
immunohistochemical staining of B7-H1 and B7-H3 are shown in
Fig. 6A and B, respectively. High
levels of B7-H1 and B7-H3 were significantly associated with liver
metastasis (Table II) and reduced
overall survival (Fig. 6C and
D).
The levels of B7-H1 and B7-H3 were then evaluated in
Rg3-treated CRC orthotopic xenografts. As shown in Fig. 6E and F, Rg3 significantly decreased
the levels of both B7-H1 and B7-H3, suggesting that Rg3 might also
promote antitumor immunity.
Discussion
Ginsenoside Rg3 exhibits antitumor activity in
various tumors (9,10). In the present study, Rg3 exerted an
inhibitory effect on the proliferation and migration of CRC cells
in a dose- and time-dependent manner. In addition, Rg3 inhibited
the growth of CRC orthotopic xenografts in vivo.
Immunohistochemistry showed that Ki-67, a well-known proliferation
index, was dramatically downregulated in Rg3-treated subjects.
CSCs, which have aberrant differentiation programs
that generate progenitor cancer cells, play a crucial role in the
formation of many solid tumors, including CRC. Deregulation of the
pathways of self-renewal and differentiation in CSCs result in
unlimited self-renewal and a subsequent excess of CSCs, which are
the source of tumor formation (13,18).
Recent studies have demonstrated that CSCs are drug-resistant and
exhibited high propensity of invasiveness and metastasis (13–16).
The presence of CSCs is also related to immune escape (19). These features of CSCs result in
cancer recurrence after eliminating most of the cancer cells by
conventional therapies. Therefore, it is of paramount importance to
develop therapies targeting CSCs. Markers, including CD24, CD44,
EpCAM and CD133, have been utilized to identify CSCs in CRC
(14–16). Combined analysis of these markers
has been used to identify CSCs. In the present study, we used the
panel of CD24, CD44 and EpCAM to analyze the stemness of CRC, and
demonstrated that Rg3 decreased the population of
CD24+/CD44+/EpCAM+ CRCs cells, the
presumed CRCs. In addition, we detected decreased
CD24+/CD44+/EpCAM+ proportions in
orthotopic xenografts tissue samples using immunohistochemistry.
Plate clone formation assays further confirmed the repressed clone
formation ability by Rg3 treatment. Therefore, we speculated that
the anticancer effect of Rg3 might also involve eliminating
colorectal CSCs.
A previous study proved that Rg3 plays a unique role
in impairing angiogenesis in tumors by inhibiting the growth of
vein endothelial cells (5). To
investigate whether anti-angiogenesis is involved as a mechanism of
the antitumor effect of Rg3, we evaluated the MVD, a well
recognized indicator for vascularization. As expected, a lower MVD
was detected in Rg3-treated subjects, suggesting inhibition of
neoangiogenesis might also be involved in the growth-inhibition
effect of Rg3 against CRC.
Using real-time PCR, we further analyzed the
expression of angiogenesis-related genes in Rg3-treated CRC cells,
and found 22 pro-angiogenic genes ANGPT1, ANGPT2,
CCL13, COL18A, CSF3, CXCL1, EGF,
FGF-2, IL1A, IL1B, IL8, KDR,
MMP1, PGF, PIGF, PLAUR, TEK,
THPO, TIMP1 and TIMP2 were significantly
downregulated. ANGPT1 (angiopoietin 1) and ANGPT2 (angiopoietin 2)
belong to the angiopoietin family. Members of this family play
important roles in vascular development, and are correlated with
tumor stage, disease progression, or metastasis (20). CCL13 (C-C motif chemokine ligand
13) and CXCL1 (C-X-C motif chemokine ligand 1) are chemokines that
play important roles in the initial step of inflammation and
angiogenesis (21,22). COL18A (collagen type XVIII alpha 1
chain) is reported to participate in regulating cell growth and
apoptosis, inflammation, angiogenesis and tissue turnover (23). CSF3 (colony stimulating factor 3)
is involved in various types of angiogenesis and skin wound healing
by inducing VEGF release from neutrophils, thus promoting
neovascularization (24). EGF
(epidermal growth factor) is a member of the epidermal growth
factor superfamily, which plays an important role in the growth,
proliferation, and differentiation of numerous cell types, and has
been demonstrated to have a critical role in tumor angiogenesis by
enhancing the expression of VEGF (25). MMP1 (matrix metallopeptidase 1) has
been identified as an important participant in tumor invasion,
metastasis, and angiogenesis (26). IL1A (interleukin 1 alpha) and IL1B
(interleukin 1 beta) are members of the interleukin 1 cytokine
family, which are involved in various immune responses,
inflammatory processes, and hematopoiesis and also play crucial
roles in the tumorigenesis of colorectal cancer (27). IL8 (interleukin 8), PLAUR
(plasminogen activator, urokinase receptor), and THPO
(thrombopoietin) participate in angiogenesis in a WNT/β-catenin
pathway-dependent manner (17).
PGF (placental growth factor) encodes a growth factor found in the
placenta that is homologous to VEGF (vascular endothelial growth
factor). Knockdown of PGF has been reported to exert antitumor
effect through PI3K/AKT and p38 MAPK signal transduction (28). TEK (vascular endothelial cell
specific receptor tyrosine kinase) has been reported to play a
crucial role in embryonic vascular development (29). Therefore, it is possible that Rg3
represses angiogenesis in CRC through mechanisms involving the
downregulation of multiple angiogenesis-related genes.
Much effort has been made towards discovering new
anti-angiogenic agents. Bevacizumab is a partially humanized
monoclonal antibody that binds to VEGF (30). It has been widely accepted that
bevacizumab can reasonably be added to either FOLFIRI or FOLFOX
chemotherapy regimens for patients undergoing first-line treatment
of metastatic CRC (7). Studies
have proven that bevacizumab can improve the median overall
survival (OS) of patients with metastatic CRC. In research
performed in the USA, 820 patients with metastatic CRC were
randomly assigned to chemotherapy with bevacizumab or chemotherapy
without bevacizumab. Patients who received bevacizumab experienced
an improved OS (11.2 months) compared with the patients who did not
receive bevacizumab (9.8 months). Median progression-free survival
was 5.7 months for patients who received bevacizumab plus
chemotherapy and 4.1 months for those who received chemotherapy
without bevacizumab (31). Based
on these data, we concluded that anti-angiogenic therapy is
effective in the treatment of CRC, especially for patients with
relapse and metastasis (2,3).
Rg3 is able to eliminate chemotherapy-resistant CSCs
and preserve its anti-angiogenic ability; therefore, we speculated
that Rg3 could be an effective supplement to chemotherapy regimens.
5-Fu is a pyrimidine class antagonist that interferes with the
growth of cancer cells, and is currently a cornerstone in the
therapeutic regimens of metastatic or advanced-stage CRC.
Oxaliplatin, as a third-generation platinum drug, is commonly used
in the adjuvant and palliative treatments of CRC. Therefore, we
investigated whether Rg3 could be applied to treatments with 5-Fu
and oxaliplatin. Using mouse orthotopic xenografts models, we
proved that the combination of Rg3 and pharmacotherapies of 5-Fu or
oxaliplatin presented stronger cytotoxicity against CRC than either
chemotherapy alone. Therefore, Rg3 shows promise for future
clinical applications.
Immune escape plays an important role in the
development of tumors. B7-H1 is also known as programmed death
ligand-1 (PDL-1) or CD274 and B7-H3 is also known as CD206. Both
belong to the B7 family, and play important roles in tumor immune
responses by integrating T cell receptor signaling to regulate T
cell function. B7-H1 and B7-H3 are recognized as predictive and
prognostic factors in various cancers. The interaction between
B7-H1 and PD-1 inhibits the activation of tumor antigen-specific T
cells, and induces immune tolerance of T cells to tumor cells, by
which the tumor cells evade immune surveillance (32,33).
Higher expression levels of B7-H1 are associated with more advanced
diseases, increased risks of recurrence and/or shorter survival
time (34–36). B7-H3 is induced in activated
dendritic cells, monocytes, and T cells with an immunoglobulin-like
structure, and is highly expressed in numerous types of cancers
(37–41). Aberrant expression of B7-H3 has
been reported to be associated with poor prognosis in patients with
colorectal and breast cancer (42,43).
B7-H3 can promote tumor progression and cancer cell metastasis, as
well as correlating with malignancy grades and the outcomes of
patients with tumors, including CRC (44–46).
In the analysis of CRC tissue samples, we confirmed
that high expression of B7-H1 and B7-H3 was significantly
associated with worse outcomes of patients with CRC, which was
consistent with previous studies. Moreover, by examining CRC
orthotopic xenografts, we found that Rg3 could decrease the level
of B7-H1 and B7-H3, suggesting that Rg3 might be able to promote
antitumor immunity. However, considering that the nude mouse is a
model of deficient T-cell function, we should be wary of making
this conclusion based on the present data. Further investigations
are required to confirm the anti-immune escape effect of Rg3.
Taken together, the results of this study showed
that Rg3 not only inhibited the growth and migration of CRC, but
also strengthened the cytotoxicity of 5-Fu and oxaliplatin in
vivo. This special antitumor effect might partly depend on
attenuating the stemness of CRC cells and remodeling the tumor
microenvironment by repressing angiogenesis. In view of this, Rg3
has the potential for clinical application in CRC treatment in the
future, and may represent a novel approach to treat this aggressive
disease.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81472296, 81602091,
81402176, 81402093, 81272542 and 81200369), the Six Major Talent
Peak Project of Jiangsu Province (grant no. 2015-WSN-022), the
Project of Invigorating Health Care through Science, Technology and
Education, Jiangsu Provincial Medical Youth Talent (grant no.
QNRC2016709), the Project of Jiangsu Provincial Commission of
Health and Family Planning (grant no. H201518), the Science and
Education for Health Foundation of Suzhou for Youth (grant no.
kjxw2015003), and the Science and Technology Project Foundation of
Suzhou (grant nos. SYS201464 and SYS201504).
References
|
1
|
Crotti S, Piccoli M, Rizzolio F, Giordano
A, Nitti D and Agostini M: Extracellular matrix and colorectal
cancer: How surrounding microenvironment affects cancer cell
behavior? J Cell Physiol. 232:967–975. 2017. View Article : Google Scholar
|
|
2
|
Colon Cancer Treatment: (PDQ(R)): Health
Professional Version: PDQ Cancer Information Summaries. Bethesda
(MD): 2002
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pattison AM, Merlino DJ, Blomain ES and
Waldman SA: Guanylyl cyclase C signaling axis and colon cancer
prevention. World J Gastroenterol. 22:8070–8077. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yue PY, Wong DY, Wu PK, Leung PY, Mak NK,
Yeung HW, Liu L, Cai Z, Jiang ZH, Fan TP, et al: The
angiosuppressive effects of 20(R)- ginsenoside Rg3. Biochem
Pharmacol. 72:437–445. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fan F, Schimming A, Jaeger D and Podar K:
Targeting the tumor microenvironment: Focus on angiogenesis. J
Oncol. 2012:2812612012. View Article : Google Scholar
|
|
7
|
Ferrara N, Hillan KJ and Novotny W:
Bevacizumab (Avastin), a humanized anti-VEGF monoclonal antibody
for cancer therapy. Biochem Biophys Res Commun. 333:328–335. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen QJ, Zhang MZ and Wang LX: Gensenoside
Rg3 inhibits hypoxia-induced VEGF expression in human cancer cells.
Cell Physiol Biochem. 26:849–858. 2010. View Article : Google Scholar
|
|
9
|
Junmin S, Hongxiang L, Zhen L, Chao Y and
Chaojie W: Ginsenoside Rg3 inhibits colon cancer cell migration by
suppressing nuclear factor kappa B activity. J Tradit Chin Med.
35:440–444. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yuan HD, Quan HY, Zhang Y, Kim SH and
Chung SH: 20(S)-Ginsenoside Rg3-induced apoptosis in HT-29 colon
cancer cells is associated with AMPK signaling pathway. Mol Med
Rep. 3:825–831. 2010.
|
|
11
|
Li W, Xie L, Chen Z, Zhu Y, Sun Y, Miao Y,
Xu Z and Han X: Cantharidin, a potent and selective PP2A inhibitor,
induces an oxidative stress-independent growth inhibition of
pancreatic cancer cells through G2/M cell-cycle arrest and
apoptosis. Cancer Sci. 101:1226–1233. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang J, Chen C, Wang S, Zhang Y, Yin P,
Gao Z, Xu J, Feng D, Zuo Q, Zhao R, et al: Bufalin inhibits HCT116
colon cancer cells and its orthotopic xenograft tumor in mice model
through genes related to apoptotic and PTEN/AKT pathways.
Gastroenterol Res Pract. 2015:4571932015. View Article : Google Scholar
|
|
13
|
Wahab SMR, Islam F, Gopalan V and Lam AK:
The identifications and clinical implications of cancer stem cells
in colorectal cancer. Clin Colorectal Cancer. 16:93–102. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cherciu I, Bărbălan A, Pirici D,
Mărgăritescu C and Săftoiu A: Stem cells, colorectal cancer and
cancer stem cell markers correlations. Curr Health Sci J.
40:153–161. 2014.
|
|
15
|
Vaiopoulos AG, Kostakis ID, Koutsilieris M
and Papavassiliou AG: Colorectal cancer stem cells. Stem Cells.
30:363–371. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ren F, Sheng WQ and Du X: CD133: A cancer
stem cells marker, is used in colorectal cancers. World J
Gastroenterol. 19:2603–2611. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu L, Zhi Q, Shen M, Gong FR, Zhou BP,
Lian L, Shen B, Chen K, Duan W, Wu MY, et al: FH535, a β-catenin
pathway inhibitor, represses pancreatic cancer xenograft growth and
angiogenesis. Oncotarget. 7:47145–47162. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang WJ, Wu MY, Shen M, Zhi Q, Liu ZY,
Gong FR, Tao M and Li W: Cantharidin and norcantharidin impair
stemness of pancreatic cancer cells by repressing the β-catenin
pathway and strengthen the cytotoxicity of gemcitabine and
erlotinib. Int J Oncol. 47:1912–1922. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Reim F, Dombrowski Y, Ritter C, Buttmann
M, Häusler S, Ossadnik M, Krockenberger M, Beier D, Beier CP, Dietl
J, et al: Immunoselection of breast and ovarian cancer cells with
trastuzumab and natural killer cells: Selective escape of
CD44high/CD24low/HER2low breast cancer stem cells. Cancer Res.
69:8058–8066. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Güveli ME, Duranyildiz D, Karadeniz A,
Bilgin E, Serilmez M, Soydinc HO and Yasasever V: Circulating serum
levels of angiopoietin-1 and angiopoietin-2 in nasopharynx and
larynx carcinoma patients. Tumour Biol. 37:8979–8983. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yamaguchi A, Nozawa K, Fujishiro M,
Kawasaki M, Suzuki F, Takamori K, Ogawa H, Takasaki Y and Sekigawa
I: CC motif chemokine ligand 13 is associated with rheumatoid
arthritis pathogenesis. Mod Rheumatol. 23:856–863. 2013. View Article : Google Scholar
|
|
22
|
Lai TH, Wu PH and Wu WB: Involvement of
NADPH oxidase and NF-κB activation in CXCL1 induction by vascular
endothelial growth factor in human endometrial epithelial cells of
patients with adenomyosis. J Reprod Immunol. 118:61–69. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Luo X, Pan Q, Liu L and Chegini N: Genomic
and proteomic profiling II: Comparative assessment of gene
expression profiles in leiomyomas, keloids, and surgically-induced
scars. Reprod Biol Endocrinol. 5:352007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kameyama H, Udagawa O, Hoshi T, Toukairin
Y, Arai T and Nogami M: The mRNA expressions and
immunohistochemistry of factors involved in angiogenesis and
lymphangiogenesis in the early stage of rat skin incision wounds.
Leg Med (Tokyo). 17:255–260. 2015. View Article : Google Scholar
|
|
25
|
Hung MS, Chen IC, Lin PY, Lung JH, Li YC,
Lin YC, Yang CT and Tsai YH: Epidermal growth factor receptor
mutation enhances expression of vascular endothelial growth factor
in lung cancer. Oncol Lett. 12:4598–4604. 2016.
|
|
26
|
Zhang Z, Wang L, Du J, Li Y, Yang H, Li C,
Li H and Hu H: Lipid raft localization of epidermal growth factor
receptor alters matrix metalloproteinase-1 expression in SiHa cells
via the MAPK/ERK signaling pathway. Oncol Lett. 12:4991–4998.
2016.
|
|
27
|
Yan H, Sun R, Pan X, Li Z, Guo X and Gao
L: Lack of association between an insertion/deletion polymorphism
in IL1A and risk of colorectal cancer. Genet Mol Res. 14:8490–8495.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Akrami H, Mahmoodi F, Havasi S and Sharifi
A: PlGF knockdown inhibited tumor survival and migration in gastric
cancer cell via PI3K/Akt and p38MAPK pathways. Cell Biochem Funct.
34:173–180. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dopheide JF, Geissler P, Rubrech J, Trumpp
A, Zeller GC, Bock K, Dorweiler B, Dünschede F, Münzel T, Radsak
MP, et al: Inflammation is associated with a reduced number of
pro-angiogenic Tie-2 monocytes and endothelial progenitor cells in
patients with critical limb ischemia. Angiogenesis. 19:67–78. 2016.
View Article : Google Scholar
|
|
30
|
Koukourakis GV and Sotiropoulou-Lontou A:
Targeted therapy with bevacizumab (Avastin) for metastatic
colorectal cancer. Clin Transl Oncol. 13:710–714. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bennouna J, Sastre J, Arnold D, Österlund
P, Greil R, Van Cutsem E, von Moos R, Viéitez JM, Bouché O, Borg C,
et al: ML18147 Study Investigators: Continuation of bevacizumab
after first progression in metastatic colorectal cancer (ML18147):
A randomised phase 3 trial. Lancet Oncol. 14:29–37. 2013.
View Article : Google Scholar
|
|
32
|
Chen Z, Pang N, Du R, Zhu Y, Fan L, Cai D,
Ding Y and Ding J: Elevated expression of programmed death-1 and
programmed death Ligand-1 negatively regulates immune response
against cervical cancer cells. Mediators Inflamm. 2016:68914822016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bryan LJ and Gordon LI: Releasing the
brake on the immune system: The PD-1 strategy for hematologic
malignancies. Oncology (Williston Park). 29:431–439. 2015.
|
|
34
|
Baptista MZ, Sarian LO, Derchain SF, Pinto
GA and Vassallo J: Prognostic significance of PD-L1 and PD-L2 in
breast cancer. Hum Pathol. 47:78–84. 2016. View Article : Google Scholar
|
|
35
|
Reiss KA, Forde PM and Brahmer JR:
Harnessing the power of the immune system via blockade of PD-1 and
PD-L1: A promising new anticancer strategy. Immunotherapy.
6:459–475. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ghebeh H, Mohammed S, Al-Omair A, Qattan
A, Lehe C, Al-Qudaihi G, Elkum N, Alshabanah M, Bin Amer S, Tulbah
A, et al: The B7-H1 (PD-L1) T lymphocyte-inhibitory molecule is
expressed in breast cancer patients with infiltrating ductal
carcinoma: Correlation with important high-risk prognostic factors.
Neoplasia. 8:190–198. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sun M, Richards S, Prasad DV, Mai XM,
Rudensky A and Dong C: Characterization of mouse and human B7-H3
genes. J Immunol. 168:6294–6297. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chapoval AI, Ni J, Lau JS, Wilcox RA,
Flies DB, Liu D, Dong H, Sica GL, Zhu G, Tamada K, et al: B7-H3: A
costimulatory molecule for T cell activation and IFN-gamma
production. Nat Immunol. 2:269–274. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Steinberger P, Majdic O, Derdak SV,
Pfistershammer K, Kirchberger S, Klauser C, Zlabinger G, Pickl WF,
Stöckl J and Knapp W: Molecular characterization of human 4Ig-B7-H3
a member of the B7 family with four Ig-like domains. J Immunol.
172:2352–2359. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xu H, Cheung IY, Guo HF and Cheung NK:
MicroRNA miR-29 modulates expression of immunoinhibitory molecule
B7-H3: Potential implications for immune based therapy of human
solid tumors. Cancer Res. 69:6275–6281. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang F, Wang G, Liu T, Yu G, Zhang G and
Luan X: B7-H3 was highly expressed in human primary hepatocellular
carcinoma and promoted tumor progression. Cancer Invest.
32:262–271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ingebrigtsen VA, Boye K, Nesland JM,
Nesbakken A, Flatmark K and Fodstad Ø: B7-H3 expression in
colorectal cancer: Associations with clinicopathological parameters
and patient outcome. BMC Cancer. 14:6022014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Maeda N, Yoshimura K, Yamamoto S, Kuramasu
A, Inoue M, Suzuki N, Watanabe Y, Maeda Y, Kamei R, Tsunedomi R, et
al: Expression of B7-H3 a potential factor of tumor immune evasion
in combination with the number of regulatory T cells, affects
against recurrence-free survival in breast cancer patients. Ann
Surg Oncol. 21(Suppl 4): S546–S554. 2014. View Article : Google Scholar
|
|
44
|
Baral A, Ye HX, Jiang PC, Yao Y and Mao Y:
B7-H3 and B7-H1 expression in cerebral spinal fluid and tumor
tissue correlates with the malignancy grade of glioma patients.
Oncol Lett. 8:1195–1201. 2014.PubMed/NCBI
|
|
45
|
Ingebrigtsen VA, Boye K, Tekle C, Nesland
JM, Flatmark K and Fodstad O: B7-H3 expression in colorectal
cancer: nuclear localization strongly predicts poor outcome in
colon cancer. Int J Cancer. 131:2528–2536. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yamato I, Sho M, Nomi T, Akahori T,
Shimada K, Hotta K, Kanehiro H, Konishi N, Yagita H and Nakajima Y:
Clinical importance of B7-H3 expression in human pancreatic cancer.
Br J Cancer. 101:1709–1716. 2009. View Article : Google Scholar : PubMed/NCBI
|