1. HPV in the general context
Human papilloma viruses (HPVs) are a small group of
non-enveloped viruses belonging to the Papillomaviridae family with
strong similarities to polyoma viruses (1). The viral particles consist of a
genome in the form of a circular double-stranded DNA, encompassing
eight open reading frames, as well as a non-enveloped icosahedral
capsid. Their size is approximately 52–55 nm and they are composed
of 72 pentameric capsomers (2). To
date, >170 types of HPV have been identified (3), the majority of which affect the
genital tract epithelia (approximately half of them), the mucosa of
the upper respiratory tract and the skin developing
epidermodysplasia verruciformis (3,4).
‘High-risk’ mucosal HPV types, predominantly types 16, 18, 31, 33
and 35, have been associated with most cervical, penile, vulvar,
vaginal, anal and oropharyngeal cancers and pre-cancers (5). HPV types 16 and 18 are the most
common high-risk types and are considered to be responsible for
>70% of all cervical cancer cases (5).
HPV are characterized by the presence of three
functional code regions in their genome: The E region that codes
the early viral function, the L region which is responsible for the
late viral function and the long control region (LCR) that is
situated between the previous regions. The density of an intact
virion is approximately 1.34 g/ml in cesium chloride and a
sedimentation coefficient (S20, W) of 300 (6).
The link between HPV and cancer has been extensively
investigated over the past decade and, for this discovery,
Professor Harald zur Hausen, the ‘Father of HPV Virology’, received
the Nobel Prize (7). ‘Low-risk’
HPV types cause benign anogenital warts and recurrent respiratory
papillomatosis and are only rarely found in squamous
intraepithelial lesions (8).
Despite the fact that HPVs are known to be
responsible for the development of cervical cancers, HPV infections
are often asymptomatic, unrecognized and underestimated. The Center
for Disease Control and Prevention (CDC) highlights the fact that
the majority of sexually active individuals become infected with
HPV at least once in their lifetime (9), often without being aware and not
presenting any symptoms. HPV infection is considered the most
common sexually transmitted disease among both males and females.
Indeed, HPV-associated cervical cancer is one of the most prevalent
types of cancer over the past years and the third main cause of
cancer-related mortality among adults (9). Furthermore, HPV types 16 and 18
high-risk genital types are implicated in the majority of
ano-genital tract malignancies, while HPV types 6 and 11 are
considered to be low-risk HPV types, which induce laryngeal
papillomatosis and genital warts. Other identified genital HPV
types are: 31, 33, 35, 39, 45, 51, 52, 56, 58, 66 and 69 (10).
It has recently been demonstrated that in the USA,
women are twice more likely to develop a HPV infection than men,
and that the prevalence of HPV is much higher in women than in men,
the peak time for acquiring the infection being shortly after
becoming sexually active (11). In
the majority of HPV-infected individuals, the virus will clear out
naturally, without the individual being aware they were infected or
exposed to the virus. An immune-deficient woman will develop
cervical cancer within 5 to 10 years, while for a woman with a
normal immune system; this time frame may be extended up to 15 to
20 years or even longer (11).
Oropharyngeal cancer can be induced by members of
the HPV family, among which HPV type 11 is the most prevalent, with
a percentage of 50–84% being the cause of causing laryngeal
papillomatosis. Even so, in the adult population, HPV 16 is the
type most commonly associated with cancer incidence, being
correlated to 86% of all HPV-related cancers. Thus, HPV type 16 has
been identified both in upper respiratory tract papillomatosis, but
also in the normal mucosa adjacent to lesions (12). It has been demonstrated that
tonsilar cancer in particular, has a strong association with HPV
(approximately 60% this type of cancer is ascribed to HPV)
(12).
2. Development of HPV infections
Oncogenic high-risk associated HPV strains have the
strongest contribution to the occurrence of pre-cancerous and
cancerous lesions (11). Age
trends in cervical HPV acquisition and persistence have revealed
that with age, the incidence of new infections decreases, while
persistence increases, emphasizing the utility of HPV screening in
older women (13). This may result
from age-related immune senescence affecting HPV clearance, the
reduced size of squamocolumnar junctions and the replacement of the
cervical mucosa by atrophic stratified squamous epithelium
(13). Of note, HPV DNA is present
in oral and genital samples of asymptomatic neonates soon after
birth (14) and it has been
suggested that HPV infection can be acquired very early in infancy
(15) in utero via the
placenta or cord blood, perinatally or through maternal breast
milk. High carriage rates of HPV DNA have been detected in oral
samples from newborn babies (16)
that decrease gradually during the first 3 years of life (16). Moreover, ‘high-risk’ mucosal types
have been detected in genital samples obtained from asymptomatic
infants, with a decreasing genital HPV DNA carriage rate during the
first year of life (16).
In a recent Romanian study, only 15% of individuals
tested positive for HPV had low-risk strains, while the remainder
had either high-risk or mixed high- and low-risk strain infections.
The distribution of high-risk HPV strains was found to prevail in
women younger than 35 years, indicating that additional attention
may be necessary for this age group regarding the risk of the
development of HPV-related neoplasms (17).
HPVs, including high-risk types 16 and 18, have also
been detected in tonsillar or adenoid samples from children with
tonsillar or adenoid hyperplasia, chronic tonsillitis and normal
mucosa (18), as well as in
genital samples from pre-pubertal girls with no known vulvar
disease (19). A so-called ‘Trojan
horse oncogenic strategy’ for HPV in childhood has been proposed,
stating that children may represent a reservoir of ‘silent’
high-risk HPV types that may be the key to HPV persistence and
related carcinogenesis later on in adulthood (6).
However, apart from persistent HPV infection, other
viral, host and environmental co-factors (20), as well as psychological stressors
(21–23) have been proposed to be involved in
HPV-related carcinogenesis. Oxidative stress is generated in
aerobic cells during infections, inflammation, physical, mechanical
and chemical stresses and it promotes viral infection, viral
persistence and the integration of the viral DNA into the host
genome (20). HPV-infected
epithelial surfaces are a constant target for oxidative
stress-induced genotoxic, anti-apoptotic and pro-angiogenetic
effects and also for oxidative stress/HPV cooperation in initiating
and promoting carcinogenesis (20). Oxidative stress promotes the
activator protein-1-mediated expression of the E6 and
E7 viral oncogenes that further target and promote the
degradation of p53 and some members of the retinoblastoma protein
family, leading to genomic alterations, the inhibition of apoptosis
and neoplasia (20).
3. Tendencies in cancer occurrence in
specific epithelial areas
High-risk HPV are causally associated with 99% of
cervical, 25% of head and neck, 70% of vaginal, 88% of anal, 43% of
vulvar and 50% of penile carcinomas, in descending order by
prevalence (1,24–27).
While the incidence of cervical cancer has declined steadily due to
intensive screening programs, the incidence of HPV-positive
(HPV+) head and neck carcinomas has significantly
increased over the past decades (25), becoming the second most common
localization for HPV-associated tumors (1,25).
By the year 2020, the annual number of HPV+ head and
neck carcinomas has been estimated to be the highest among
HPV-associated cancers in the USA (12). Importantly, HPV+ and
HPV-negative (HPV−) head and neck carcinomas have been
proven to be different clinical entities in terms of risk factors,
anatomical sites of involvement, molecular alterations, pathology
appearance and prognosis (28).
HPV+ head and neck carcinomas primary involve the
oropharynx (tonsils, base of the tongue and other parts of the
oropharynx) (24) and are caused
by HPV type 16 in >90% of cases (25). It has been demonstrated that the
E6 and E7 genes are the main viral oncogenes by which
HPV 16 and other high-risk HPV inactivate p53 and retinoblastoma
protein, respectively, leading to the inhibition of apoptosis, cell
cycle progression (1,24), the accumulation of genetic
alterations, viral integration and ultimately to uncontrolled cell
proliferation (20,29). Experimental data have indicated
that the expression of E6/E7 is mandatory for the initiation
and maintenance of the malignant phenotype in oropharyngeal cancers
(28). By contrast, HPV-related
head and neck carcinomas are associated with exposure to tobacco
and alcohol consumption, with the incidence decreasing in developed
countries due to aggressive smoking cessation campaigns (30).
Furthermore, HPV positivity is associated with a
significantly superior loco regional control (30) and a better prognosis of
recurrent/metastatic head and neck carcinoma (31). In fact, the tumor HPV status is of
paramount importance in predicting the survival for patients with
local regionally advanced oropharyngeal cancer and its
determination is included in the routine evaluation of head and
neck cancers for prognostic assessment (28). The biological explanation for these
different outcomes is currently unknown and, as in other
pathologies (32), the novel
‘-omics’ technologies are promising in the pinpointing of reliable
biomarkers useful for patient stratification, the evaluation of
therapeutic regimens and identification of potentially targetable
pathogenic pathways (30). Of
note, HPV-related head and neck carcinomas share molecular
characteristics with lung squamous carcinoma, whereas
HPV+-related head and neck carcinomas have similar
characteristics with cervical neoplasia (33). This latter observation can be in
part explained by the fact that oral HPV infection and type
concordance in women with cervical HPV infection are frequent, and
increase significantly in human immunodeficiency virus (HIV)
infection, demonstrating a degree of correlation between the two
infection sites (34).
HPV infection is an important risk factor for penile
SCC, the most frequently reported pathology of penile cancer
(>95%). It is a rare malignancy accounting for 0.24% of all
neoplasms among males in the United States with a significantly
higher incidence (up to 20–30-fold greater) in areas of Africa and
South America that usually arises from the epithelium of the inner
prepuce or the glans, and it exists in several histological
subtypes sharing a similar pathology with SCC of other origins
(35–38). Viral DNA has been detected in
70–100% of penile intraepithelial neoplasia and in 30–40% of
invasive cancer tissue samples (35). HPV infection (particularly by the
high-risk HPV 16, 18, 31, 33, 45, 56 and 65) is nowadays recognized
as a major co-factor in penile SCC through interaction with
oncogenes and tumor suppressor genes (p53, retinoblastoma
protein, p16) (37,39). Nevertheless, it is now clear that
apart from the HPV-induced pathway (through which up to 50–80% of
penile SCC cases arise), a non-HPV-induced pathway represents a
divergent molecular pathway accounting for penile carcinogenesis
related to several risk factors, such as chronic inflammation and
specific mediators (37,40).
4. Conditions favoring HPV development
HPV enters epithelial cells of cutaneous or mucosal
surfaces through abrasion (6) and
further requires wounding or micro-wounding in order to allow the
access of the virus to the basal lamina (41). In most cases, HPV infection
resolves spontaneously and is transient and asymptomatic (42). The development of HPV infections
and lesion formation may involve the appearance of the wound
healing response that promotes active cell division and the
proliferation of infected cells (29,43,44).
Persistent infection with high-risk HPV types and
altered viral gene expression are the cornerstone of HPV-induced
carcinogenesis (29). Life-cycles
of low-risk and high-risk HPV types differ in their ability to
drive cell cycle entry and enhance cell proliferation in the basal
cell layers (29). The E6
and E7 viral oncogenes are considered to have a significant
contribution to these differences, as E6 inactivates
p53 and E7 promotes the degradation of several
retinoblastoma protein family members, further leading to cell
cycle deregulation and the inhibition of p53-mediated apoptosis
(1,25,29).
Moreover, while HPV infection requires the appearance of wound
healing and inflammation, in some cases, pro-inflammatory
cytokines/chemokines can trigger malignant transformation.
Tumor-induced inflammation in turn can promote the proliferation
and survival of malignant cells and may alter the response to
cancer-targeted therapeutic agents (45).
However, only a small fraction of infections with
high-risk HPV types will eventually evolve into anogenital or head
and neck carcinomas and various co-factors have been proposed to be
involved in the progression of HPV infection in the epithelial
neoplasia. The most consistently identified factors in HPV-related
carcinogenesis include high parity, the long-term use of oral
contraceptives, smoking, concomitant infection with other sexually
transmitted agents (46), the
immune status, nutrition, endogenous and exogenous hormones, as
well as viral characteristics, such as HPV type, viral load, HPV
variant and viral integration (42,47).
5. HPV occurrence in comparison to other
sexually transmitted diseases
HPV is the most prevalent sexually transmitted
infection worldwide (48–50), associated with a profound social
and economic burden (51). In
fact, the majority of sexually active individuals contract at least
one type of HPV at some point in their lives; however, in the
majority of cases, HPV infection is transient or asymptomatic and
is resolved spontaneously (42).
Therefore, it is considered appropriate that treatment is
vindicable only for clinically visible or microscopically
pre-cancerous lesions caused by HPV and is not recommended for the
virus itself (42). However, new
evidence suggests that the early detection of known high-risk HPV
infection and treatment should be considered even if clinically or
microscopically pre-cancerous lesions are not visible. This early
detection is not currently available on a routine basis; thus, the
development of newer molecular genetic diagnosis methods is
mandatory.
Many types of HPV are sexually transmitted through
the anogenital contact, mainly during vaginal and anal sex, and
also during genital-to-genital contact without penetration and oral
sex (42). Exceptionally, HPV may
be transmitted by non-sexual routes, including casual physical
contact via auto-inoculation or fomites, as well as vertically from
HPV-infected pregnant women to their newborns during delivery
(1,6,42) or
in utero (14). In a 2015
report on global sexually transmitted infection surveillance, WHO
estimated that worldwide, >290 million sexually active women
will at some point become infected with HPV and an estimated 417
million cases are prevalent for herpes simplex viral infection
(52,53). Male prevalence is not yet known,
although it may be even higher due to the high clinically
asymptomatic rates. For a comparison, the global estimate in
incidence for the most common sexually transmitted infections that
are considered curable (chlamydia, gonorrhoea, trichomoniasis, and
syphilis) was approximately 357×106 new infections in
2012 (Table I) (54).
 | Table IIncidence and prevalence of the most
frequent sexually transmitted diseases. |
Table I
Incidence and prevalence of the most
frequent sexually transmitted diseases.
| STD | World
incidence | World
prevalence | US incidence | US prevalence |
|---|
| 1. HPV | N/A | 291,000,000 | 14,100,000 | 79,100,000 |
| 2. Chlamydia | 130,900,000 | N/A | 2,860,000 | 24,100,000 |
| 3.
Trichomoniasis | 142,600,000 | N/A | 1,090,000 | 3,710,000 |
| 4. Gonorrhea | 78,300,000 | N/A | 820,000 | 1,570,000 |
| 5. Syphilis | 5,600,000 | N/A | 55,400 | 422,000 |
| 6. HSV | N/A | 417,000,000 | 776,000 | 908,000 |
| 7. HIV | 1,800,000 | 36,700,000 | 41,400 | 270,000 |
| 8. Hepatitis B | N/A | 248,000,000 | 19,000 | 117,000 |
In 2013, CDC published the latest available data for
the US, demonstrating that roughly >6% of Americans annually
contract a new sexually transmitted infection and approximately 1
in 3 Americans have a sexually transmitted infection (numbers
corresponding to an incidence of 19.7×106 and a
prevalence of 110×106 sexually transmitted infections at
a population of 304,09×106 for the year 2008), with an
estimated economic burden of over 16×109 US $ with
3×109 US $ in direct medical costs (in 2010). Of great
concern is the fact that >20% of these infections
(22.1×106) with an acquisition rate of almost
9.8×106 a year were among men and women aged 15 to 24
years, which in many cases were not clinically evident, accounting
for most of both prevalent and incident infections (48,51).
Although there is no reporting system for HPV in the
USA, more conclusive data is available through CDC statistics. As
stated in a recent brief, among adults aged between 18–69 years in
2013–2014, approximately 45% of men and 40% of women had a genital
HPV infection. Approximately 25% of men and 20% of women had a
high-risk genital HPV infection. The prevalence of any type of oral
HPV infection among adults aged between 18–69 years in 2011–2014
was approximately 7%, and the prevalence of high-risk oral HPV
infection was 4% (55). The vast
majority of HPV infections (approximately 90%) are spontaneously
resolved within 1 or 2 years, being cleared by the immune system
(56,57).
An increasing amount of data suggests that HPV
infection, even asymptomatic, can significantly increase the
morbidity associated with other sexually transmitted diseases. In a
recent study, it was shown that reduced sperm motility/abnormal
morphology significantly decreased fertility in a male heterosexual
population with prostatitis-related symptoms secondary to
Chlamydia trachomatis and HPV co-infection (58). In another study, Chlamydia
trachomatis infection in both HPV+ and
HPV− females was investigated (59). It was concluded that Chlamydia
trachomatis infection was much more prevalent in
HPV+ subjects and when the two sexually transmitted
infections co-existed, there was an increased risk for detecting
single high-risk HPV genotypes and various multiple HPV infections
even in asymptomatic patients. Chlamydia trachomatis
infection was investigated as a potential HPV co-factor in invasive
cervical cancer as early as 2002. Smith et al investigated a
Brazilian and Philippinese invasive cervical cancer population and
found that Chlamydia trachomatis infection increased the
risk of developing SCC among HPV+ patients (60). Although a direct interaction
between Chlamydia trachomatis and HPV was not yet
demonstrated, the authors of that study postulated that
Chlamydia trachomatis can act as a carcinogenic co-factor by
means of chronic inflammation, in which reactive oxygen species may
damage DNA, thus providing a possible link towards malignant
transformation (60).
Another study identified a possible association
between HPV and hepatitis C virus. In liver transplant candidates,
HPV+ testing was prevalent in non-sexually active women,
suggesting a possible reactivation of dormant HPV due to a
dysregulation in T-cell activity by the hepatitis C virus (61). Genital herpes virus (HSV-2)
infection was long thought to be a co-factor in cervical cancer
development (62). On its own,
HSV-2 infection represents a social and economic burden, with
approximately 1.1×106 individuals (15.9%) reporting
asymptomatic genital herpes and 47×106 new cases in 2013
(63). It has been suggested that
in conjunction with HPV, HSV can accelerate the process of
tumorigenesis, in part due to the repeated inflammation caused by
its periodic outbursts. Yet, this theory remains to be validated,
since HSV was not present in all cervical cancer tissue samples as
opposed to HPV. In another study in a population of genital
HPV-infected men, it was found that that Chlamydia
trachomatis and HSV-2 infections were more prevalent (64). However, it remains uncertain as to
whether these infections can influence the acquisition and/or
persistence of HPV through a biological interaction, such as a
decrease in cell-mediated immunity (64).
There is evidence that HIV infection
(>35×106 cases worldwide with approximately
1.8×106 new ones in 2016) can increase the number of
infected HPV particles carried on a single individual (http://www.who.int/mediacentre/factsheets/fs360/en).
This may be the result of the negative impact of HIV on the immune
system functionality. Among couples, HIV infection can impact HPV
carriage. It has been reported that in couples where women are
HIV-positive, their HIV-negative male partners have a significantly
higher rate of HPV penile carriage compared to those of
HIV-negative women (58 vs. 32%) (65). If both partners are HIV positive,
the carriage rate increases to 72%. Moreover, HIV co-infection
increases the risk of type-specific HPV sharing within the couple
(65). Furthermore, two large
meta-analyses, performed in women from the general population and
in HIV-positive women, revealed that HPV prevalence was higher
within the same region in HIV-positive compared to HIV-negative
individuals (66,67).
6. HPV persistence and cancer incidence
CDC outlines that in approximately 90% of cases, HPV
infection is likely to disappear within approximately two years,
causing no harmful effects. However, if there is a persistent
infection, due to high-risk strain HPV strains, coupled with
host-particularities, particular behavior such as smoking and/or
alcohol abuse, environmental co-factors, other associated viral
infection such as HIV, persistence can lead to neoplastic
transformation (9,25,34,68,69).
The link between well-studied genital cancer and oral cancer
triggered by persistent HPV infections is still a subject of
intense investigation (70).
High-grade cervical intraepithelial neoplasia and the association
with HPV have been studied in various cohorts and geographical
regions. In over 8,600 Danish women tested at 2 years, it was
reported that subjects that tested positive for HPV type 16 at the
second examination had an estimated probability of developing CIN
grade 3 within 12 years of 26.7%, while those infected with HPV
type 18 of 19.1% (71). This study
revealed that the persistence of HPV type 16 was particularly
associated with a high risk to develop high-grade cervical lesions
(71). In >2,000 women from
Africa, HPV prevalence was reported as 24.5% for any HPV type and
as 16.1% for the oncogenic ones. Over 21 months of follow-up,
type-specific persistence was 29.8% among all HPV infections giving
good grounds for vaccine development (72).
In a Finish female cohort, the natural HPV type 16
clearance of patients with positive cervical and oral samples was
investigated. Natural clearance correlated with HPV antibody titers
(73). In this context, it was
reported that a duration of HPV persistence of >12 months leads
to an increased risk for disease progression in cervical and oral
neoplasias (73,74). In high-grade squamous
intraepithelial lesions, an extended high-risk HPV persistence of
>6 months was the most potent predictor of progression (74). Later on, when HPV vaccines were
investigated, the results obtained from a study comprising over
15,000 female subjects confirmed that a persistence of HPV
infection of >6 months was the surrogate endpoint of progressive
disease (75). In oral mucosa
infection, HPV type 16 was the main cause of infection (65%), other
low-risk HPV genotypes cleared from the oral mucosa rapidly in
comparison to the high-risk HPV genotypes 12 (75).
When investigating the association between HPV
infection with oropharyngeal and anogenital sites in men engaging
in sexual activities with other men, no congruity between HPV
genotypes discovered at oral sites as compared with the anogenital
sites was found; thus, it is possible that oral HPV infection may
be considered independently (76).
In esophageal SCC diagnosed in a Greek population, it was reported
that there are important positive association factors, such as HPV
positivity, the mutational activation of K-ras and tobacco/alcohol
abuse, that are main risk factors for the development of this type
of cancer (30,77).
HPV infection can trigger cervical cancer,
accounting for approximately 25% of head and neck cancer cases
(78). In female subjects, there
is an increased prevalence of oral HPV infections with concomitant
cervical HPV infection with a moderate type-specific HPV
concordance between genital and oral HPV types. The long-term
persistence of cervical high-risk HPV (over 24 months of
positivity), can induce an increased incidence of oral HPV
infection later in time. Lower titers of HPV type 16 antibodies may
suggest the incapacity to clear chronic cervical HPV infections
(79).
Despite prior findings demonstrating the lack of a
correlation between the oral finding of HPV DNA and HPV serology,
there are data showing the most marked titers of HPV type-specific
antibodies in female subjects clearing cervical HPV type 16
infection and the lowest HPV type 16 antibody levels in females
with acquired cervical HPV type 16 infections (72). There are also studies suggesting
the possibility of a subcategory of women with HPV persistence at a
very young age (72,80,81).
A study conducted by Koskimaa et al in children whose
mothers had cervical intra-epithelial neoplasm lesions,
demonstrated that these children had HPV type 16-specific
cell-mediated immunity. HPV infections persistent earlier in life
can affect the outcome of subsequent genital or oral HPV infections
(82).
In terms of the clinical manifestation of HPV
infection, these may vary from asymptomatic forms, beginning from
benign warts and ending in carcinomas of malignant potential
(17). In an attempt to elucidate
the presence of benign warts as a site of persistence for future
neoplastic transformation, Kofoed et al conducted a study
including 201 men and women with genital wart-like lesions
(83). Their study found HPV in
46.2% of subjects, out of which oral HPV was identified in 10.4% of
subjects. In 21.7% of investigated subjects, concordance was found
between oral and anal HPV types. Patients with genital warts
frequently display extra-genital HPV with differences between men
and women subjects (83).
HPV infection has specific sites for localization.
In order to complete the HPV infection generation cycle, the virus
requires a stratified epithelium. That is why HPV-associated
cancers occur in areas with intermediate epithelia towards cubic
mucosal epithelia, such as the lip, cervix, oral cavity or the
rectum (29). The usual
progression begins with initial virus infestation of the basal cell
nucleus, overcoming the host defense mechanisms. Once established
in the basal cell nucleus, the virus genome multiplies with the
host DNA, producing a reduced number of copies which later, on cell
division, are allocated to daughter cells. In this phase, there is
a low viral gene expression, and thus infected cells are difficult
to detect (84). In light of the
overall body of evidence, the vast majority of HPV infections
(approximately 90%) are cleared naturally by the body’s immune
system within a two-year period; however, when persistence is
established for >6 months, infection induces a high risk of the
malignant transformation of infected cells.
7. HPV prevention
Sexual abstinence is the most reliable method of
protection against HPV, as with other sexually transmitted
infections (8,9). Limiting the number of lifetime sexual
partners, cultivating a monogamous relationship and having safe
sexual habits can reduce the exposure to HPV (85). The consistent and correct use of
condoms, although not offering complete protection, can decrease
the risk of transmitting HPV and developing HPV-associated
conditions (8,9).
Available vaccines (bivalent, quadrivalent and
9-valent vaccines) are effective against high-risk HPV types 16 and
18, and are the optimal prevention strategy for cervical cancer to
date (11). The quadrivalent
vaccine also protects against HPV types 6 and 11, which are
responsible for 90% of ano-genital warts; whereas the 9-valent
vaccine also covers HPV types 31, 33, 45, 52 and 58, that account
for 15% of cervical cancer cases (8,9). All
vaccine types are safe and should be administered as a 3-dose
series of intra-muscular injections over a 6-month period for the
female population aged 11–12 years, but vaccination can begin 9
years of age (86). Boys are
advised to receive the quadrivalent (11) or the 9-valent HPV (8,9)
vaccine at the same age interval as girls. HPV vaccines can be
administered regardless of a history of anogenital warts and
pre-cancerous lesions or abnormal Pap/HPV tests (8,9), in
men or women not older than 26 years (86). Although a ≥1 dose of HPV
vaccination coverage increases notably each year, it is still much
below the expected coverage of 91% by 13 years of age, due to
missed vaccination opportunities/non-adequate efforts in promoting
the advantages of reducing vaccine-preventable infections and
cancers (11). Consequently, the
Advisory Committee on Immunization Practices recommends
administration of HPV vaccine at the same visit with other
age-appropriate vaccines (11).
Geographic variations of different HPV types have been reported by
numerous studies on women with cervical lesions of various degrees
of severity, including cervical cancer (7,87,88)
and should be taken into account when designing future HPV
vaccination programs to prevent HPV-induced carcinogenesis
(7).
8. Proteomics technologies for the
identification of markers
Discoveries of proteomics biomarkers in
HPV-related cancer
Given the expansion of proteomics studies and the
advances brought by this strategy in the diagnosis, prognosis and
therapy monitoring, proteomics have recently focused on HPV-related
neoplasia (73). Oncogenic human
DNA viruses, such as HPV, have co-evolved with their human hosts
and studies of viral proteomics can identify important tumorigenic
pathways, as well as novel therapeutic targets (70,89).
Nevertheless, there are still extensive gaps in proteomic knowledge
related to HPV-related cancer forms, the difficulties due to the
HPV prevalence and type distribution in women with HPV-related
cancers other than cervical ones (27). The complexity of this group of
diseases can be matched only by equally complex proteomic data
(29,30).
Several ‘omic’ fields have studied the associations
with HPV-dependent cancer lesions. Wu et al examined the
molecular pathogenesis of HPV-related oral cavity SCC and reported
that there was a significant difference in the incidence of HPV
type 16 infection-related and HPV-non-related cancer (90). Acknowledging that paxillin can
promote lung tumor progression and that HPV type 16 E6 can
reduce miR-218 expression in cervical cancer, the role of paxillin
in oral cavity SCC was investigated. An increased capacity for
invasion in HPV-infected cancer cells was found to be associated
with paxillin expression. In tumor specimens with HPV type 16/18
infection, a negative association was observed with miR-218
expression. Moreover, a shorter relapse-free survival and overall
survival was associated in patients with tumors with a low miR-218
and high paxillin expression. HPV-infected patients with tumors
with a high paxillin and low miR-218 expression have the most
unfavorable relapse-free and overall survival. There is evidence to
indicate that targeting paxillin in order to suppress tumor
progression/improve the clinical outcome of oral cavity SCC,
particularly in HPV-infected patients, can lead to novel
therapeutic regimes (90).
Proteomics analyses, expanding on the neoplastic
differentiation of HPV-infected cells, have indicated that the
E6-mediated ubiquitination of p53 is present in infections
involving high-risk HPV types. A number of novel E6AP-interacting
proteins have been detected and validated, among which the most
important are NEURL4, HIF1AN and mitogen-activated protein kinase 6
(MAPK6) (91). E6AP comprises
several different protein complexes, including an independent
high-molecular-weight complex containing HECT and RLD domain
containing E3 ubiquitin protein ligase 2 (HERC2), neuralized E3
ubiquitin protein ligase 4 (NEURL4) and MAPK6, as well as a
proteasome (92). These proteomics
studies revealed for first time the E6AP complexity and
involvement in infections caused by high-risk HPV types (91).
Due to the small size of the HPV genome (7–8 kbp,
at most eight genes) the viral lifecycle requires these protein
products to be able to execute multiple functions. Thus, E2
proteins play an important role in the HPV lifecycle. Specifically,
the E2 proteins contain a conserved C-terminal DNA binding
structure, a dimerization domain (90–100 amino acids) and a
conserved N-terminal domain (200–210 amino acids) (92). The specific structural
characteristics of the E2 proteins allow their involvement in viral
transcription and replication; whereby they interact with host’s
proteins in order to fulfill the entire viral life cycle (84). E2 proteins aid viral replication by
establishing links between regions of cellular chromatin under
replication stress and viral replication foci (93).
In HPV+ oropharyngeal cancer, proteomics
markers, such as E2 expression have been recently associated
with CD8+ tumor-infiltrating lymphocyte counts,
age and T-stage to predict progression-free survival and stratify
patients for therapy (94). The
high tumorigenic potential of HPV relies on the cell cycle
deregulation/inhibition of p53-mediated apoptosis; both tumorigenic
processes are induced by E6 and E7, the two main
viral oncogenes. These viral oncogenes inactivate p53 and the
retinoblastoma protein (24).
Recently, the gamma HPV197 interactome investigation has indicated
that the E6 protein interacts with a large number of cellular
proteins, including the TP53 tumor suppressor that is thus
inhibited. Although it seems that the HPV197 E7 structure would not
accommodate the binding to retinoblastoma protein, it was
demonstrated that it does and activates E2F transcription
(95).
Using several proteomic technologies,
matrix-assisted laser desorption ionization time-of-flight mass
spectrometry (MALDI-TOF-MS) and fluorescent two-dimensional
electrophoresis, in cervical cancer, mimecan, actin from aortic
smooth muscle and lumican were found to be overexpressed, while
keratin, type II cytoskeletal 5, peroxiredoxin-1 and 14-3-3 protein
sigma expression levels were found to be decreased (96). Moreover, using one dimensional
electrophoresis, followed by direct mass spectrometry in SCC of the
penis, several proteins were identified. HPV+ penile SCC
and HPV− non-tumorous penile foreskins were analyzed and
63 different proteins were revealed in the HPV+ group.
One of this complex array of proteins proved to be complement C3,
which was useful in the evaluation of patients with penile SCC
(97).
Protein microarray, an emergent proteomic
technology (98), is gaining its
place in proteomics. For the study of the diversity of oncogenic
HPV types, a customized HPV protein microarray with 98 proteins in
the form of C-terminal GST fusion proteins, has been recently
developed. The utilized protein markers are representative of eight
antigens of two low-risk HPV types (6 and 11) and 10 oncogenic
high-risk HPV types (16, 18, 31, 33, 35, 39, 45, 51, 52 and 58)
demonstrating a high reproducibility in human samples.
Statistically significant IgG sero-reactivity for HPV16 E1,
E2 and E7 was obtained in oropharyngeal carcinoma
patients, validating prior ELISA results. That study is one of the
first to emphasize the potential of using protein microarray assays
in HPV-related cancer (99).
Genomic markers in HPV-related
cancer
The majority of studies on HPV-related cancers have
gathered genomic data. The following alterations have recently been
highlighted: Amplifications of regions that comprise mainly
oncogenes, deletions that comprise mainly genes involved in
proliferation and cell cycle regulations, mutations and a rapidly
expanding domain, DNA methylations. Using genomic technologies, in
various human non-melanoma skin cancers, ten different HPVs were
detected in >50% of the tested samples. The reported genomic
sequences comprised the known HPV types 16, 22, 120 and 124, two
previously known putative types and four previously unknown HPV
sequences. The most commonly detected virus was HPV197 in >30%
of all skin cancer samples (100).
In HPV+ tonsillar carcinoma there is a
marked enhancement of 3q24–27 amplifications, comprising
PIK3CA, TP63, SOX2, CCNL1,
PARP1, hTERC and DCUN1D1 oncogenes (101). 3q amplifications have also been
found in the early stages of cervical cancer that progresses to
more advanced stages (102). In
earlier head and neck SCC studies, chromosomal aberrations in 7q,
Xp, 8p, 11q22 (harboring the ATM gene), 13q, 14q32
(harboring the TRAF3 gene), and 15q have also been reported
(103). Recently, a subset of
HPV+ head and neck SCC harboring mutations in
TRAF3 and CYLD have been reported, suggesting their
involvement in HPV-related tumorigenesis (104). Moreover, gene loss
(SMAD4), hinders TGFb signaling and favors uncontrolled cell
proliferation and anti-apoptotic events in head and neck SCC
(24).
The detection of mutations in HPV+
lesions is a comprehensive research domain. TpCp(A/C/T)
substitutions in the APOBEC3B cytosine deaminase gene
altering the enzyme activity have been reported in both
HPV+ head and neck SCC and cervical cancers.
APOBEC is associated with helical domain hot spot mutations
in the PIK3CA gene detected in a number of neoplasms
(105). Certain deregulated
intracellular signaling pathways are caused by the increased
frequency of mutations, such as the PI3K pathway (PIK3CA,
PTEN, PIK3R1 genes mutated) and the MAPK pathway
(KRAS gene mutated). Using human xenografts expressing
mutant PIK3CA harvested from HPV+ and
HPV− head and neck SCC, it was demonstrated that
PIK3CA mutation status can be utilized as a target for
mTOR/PI3K inhibitors. Thus, in animals bearing xenografts with
mTOR/PIK3CA inhibitors, an attenuated tumor growth was observed
(106).
DNA methylation is another genomic domain that is
highlighted in the characterization of HPV+ head and
neck SCC. Thus, the profile identification of global DNA
methylation proves that HPV+ head and neck SCC tissues
display increased hyper methylation compared to HPV−
tumors (107). Global
hypermethylation is possibly induced by E6 ectopic
expression, thus E6 is suggested as the primary methylation
factor. Epigenetic silencing in HPV+ head and neck SCC
can modulate gene families involved in proliferation and cell cycle
regulation. The hypermethylation of the cadherin gene family has
been shown to induce HPV-mediated tumorigenic progression through
epithelial-mesenchymal transition (108). Moreover, it was recently
reported, that in HPV+ head and neck SCC, long
non-coding RNAs (lncRNAs) cluster in five distinct tumor regions
and are associated with DNA methylation and TP53 mutation,
as shown by RNAseq expression data. Importantly, this lncRNA
reported clustering correlates with patient survival and tumor
grade (109).
In terms of therapy, HPV+ tumor
responsiveness to radiotherapy can be increased by the activating
amyloid precursor protein binding protein-1 gene (APP-BP1),
as through this mechanism, the upregulation of p53-mediated
apoptosis can be further induced (110,111). Therefore, gene therapy aiming at
restoring wild-type p53 expression may prove to be a novel
therapeutical route in head and neck SCC (102,112). Another possible therapeutical
approach focuses on the epigenetic field, where the
hypermethylation of the SMG1 promoter appears to increase
HPV+ head and neck SCC cell line sensitivity to
radiation (113). As shown in a
separate study, the promoter methylation signature of five genes,
ALDH1A2low, OSR2low, GATA4high,
GRIA1high and IRX4high, may be utilized as a
prognostic marker. These methylation signatures can identify
specific treatment-responsive patients in oropharyngeal cancer
(114). It is important to
highlight that the HPV− head and neck SCC genetic
pattern differs significantly from the HPV+ one. Thus, a
loss of the 17q/mutation of TP53 and of the
9p/inactivation has been reported (103), along with amplification at locus
11q13 (the site harboring CCND1 oncogene, CTTN and
FADD genes) (25). The loss
of 16q12–24, as well as FGFR1 amplifications
(114), seems to be specific to
HPV− head and neck SCC (110). Of note, EGFR gene
amplifications localized at chromosome 7p, have been found to be
present in >13% of HPV− head and neck SCC (114). In brief, genomic markers for
HPV+ and HPV− head and neck SCC are
summarized in Table II.
| Table IIMarkers in HPV+ and
HPV− head and neck SCCs. |
Table II
Markers in HPV+ and
HPV− head and neck SCCs.
A, HPV+
head and neck SCCs
|
|---|
Marker
| Chromosome | Genetic | Epigenetic |
Trans-criptomic | Proteomic | (Refs.) |
|---|
| Family member | Type |
|---|
| Receptor tyrosine
kinases | FGFR2/3 | |
Mutation/amplification | | | | (112) |
| PI3K pathway | PIK3CA | 3q26 | Mutation | | Upregulated | | (112) |
| PIK3R1 | | Mutation | | | | (112) |
| MAPK pathway | KRAS | | Mutation | | | | (112) |
| TGFb pathway | SMAD4 | 18q21 | Loss | | | | (112) |
| Cell cycle | E2F1 | 20q | Amplification | | | | (112) |
| E2F targetsa | | | | Upregulated | Upregulated | (115) |
| PCNA | | | | Upregulated | Upregulated | (115) |
| APP-BP1 | 16q22 | Loss | | Downregulated | | (116) |
| miR familyb | | | | Upregulated | | (116) |
| miR familyc | | | | Downregulated | | (116) |
| Immune
response | TRAF3 | 14q32 | Loss | | | | (112) |
| IFN-induced
genesd | | | | Downregulated | | (117) |
| IL6 | | | | | Downregulated | (117) |
| IL10 | | | | Downregulated | | (117) |
| IL13 | | | | Downregulated | | (117) |
|
Immunoglobulins | | | | | Downregulated | (117) |
|
Lactotransferrins | | | | | Downregulated | (117) |
| Lymphocyte
activation inducede | | | | Upregulated | | (117) |
| JAK-STAT
pathway | JAK3, STAT5A | | | Promoter
methylated | | | (118) |
| EMT | Cadherin family
membersf | | | Promoter
methylated | | | (118) |
| Tissue development
and regeneration | GATA4 | | | Promoter
methylated | | | (118) |
| GRIA1 | | | Promoter
methylated | | | (118) |
| IRX4 | | | Promoter
methylated | | | (118) |
| DNA repair or
recombination | ATM | 11q22 | Loss | | | | (112) |
| BRCA1/2 | | Mutated | | | | (112) |
| Testis-specific
genesg | | | | Upregulated | | (112) |
| SMG1 | | | Promoter
methylated | | | (112) |
| miR-363 | | | | Upregulated | | (116) |
B, HPV− head and neck SCCs
|
| Receptor tyrosine
kinases | EGFR | 7p12 | Amplification | | | | (103,112) |
| FGFR1 | 8p11 |
Mutation/amplification | | | | |
| MAPK pathway | HRAS | | Mutation | | | | (103,112) |
| TGFb pathway | SMAD4 | 18q21 | Amplification | | | | (103,112) |
| Cell cycle | TP53 | | Mutation | | | | (103,112) |
| CCND1 | 11q13 | Amplification | | Upregulated | | (103,112) |
| CDKN2A | 9p21 | Loss/mutation | Promoter
methylated | Downregulated | Downregulated | (115) |
| RASSF1A | 3p | Loss | | | | (119) |
| FHIT | 3p | Loss | | | | (119) |
| Tissue development
and regeneration | ALDH1A2 | | | Promoter
methylated | | | (107) |
| OSR2 | | | Promoter
methylated | | | (107) |
| FADD | 11q13 | Amplification | | | | (120) |
| CTTN | 11q13 | Amplification | | Upregulated | Upregulated | (120) |
An overview of the ‘omics’ biomarkers related to
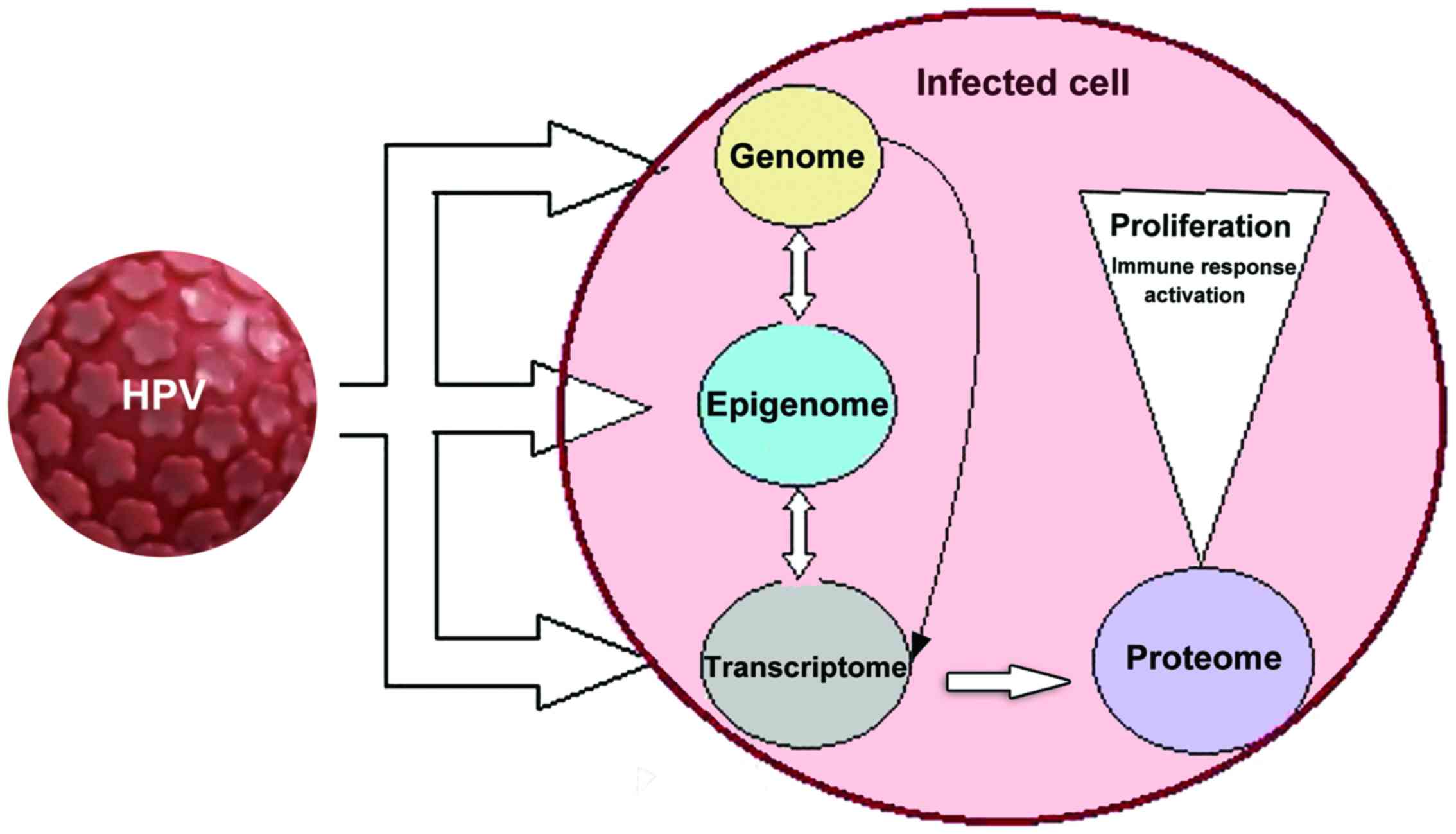
HPV infection is presented in Fig.
1. On all altered levels, beginning from HPV insertion into the
host cell genome, through epigenetic alteration and the
transcription of altered proteins, biomarkers indicative of HPV
infection can be identified.
Use of proteomics technologies for
biomarker identification
To date, major markers able to assist effective
therapy have been identified, However, further steps along this
path, are the study of interactions between the host and the virus
in order to improve disease outcomes and, more concretely, for the
development of biomarkers able to perform the accurate triage of
patients with HPV infection.
Against the increasingly confirmed knowledge that
cancer may begin from proliferative cancer stem cells of high
tumorigenic potential, to which the information is added related to
the capacity of other oncogenic viruses to induce stem
cell-associated gene expression, a study researched the possibility
to detect, on the one hand, the presence of mRNA or proteins from
stem cell associated genes and/or increased cell proliferation, and
on the other hand, in appropriate current cervical-vaginal fluid
samples from high risk HPV+ women. The use of
cervical-vaginal fluid samples has demonstrated benefits as a
biomarker source for lower female reproductive system pathologies,
due to easy collection, with the prospective of utilizing
self-tests. The following detection methods have been used, to
date, for this sample examination: RT-qPCR for mRNA, flow cytometry
and antibody-based proteomic microarray for stem cell-associated
genes and flow cytometry for increased cell proliferation (121).
The study material also included cytological and
histological sample results, indicating moderate-high abnormality
rates. In samples from patients with CIN3, respective analysis
revealed upregulated stem cell-associated proteins (e.g., the
oncogene TP63, human chorionic gonadotropin) and the
SOX2 transcription factor. Moreover, certain samples
collected from a subset of this patient group, the stem
cell-related, surface podocalyxin protein could be detected. In
high-grade disease samples, SOX2, TP63 and human
gonadotrophin mRNAs were upregulated. Immunohistochemistry
experiments performed revealed that SOX2 and TP63 proteins visibly
outlined tumor cells in invasive forms of squamous cervical cancer
(121).
A different study envisaged the identification of a
cervical-vaginal fluid protein biomarker indicative of cervical
cancer or its respective pre-cancer stage. The study involved a
differential proteomics approach applied to cervical-vaginal fluid
samples collected from healthy patients or women at pre-cancer
stages. The quantification of protein expression was performed by
label-free spectral counting and proteome analysis (122). This approach identified 16
candidate biomarkers, the most promising of which were
alpha-actinin-4 and pyruvate kinase isozyme M1/M2. ELISA applied to
alpha-actinin-4 demonstrated the potential of this candidate
biomarker to distinguish between healthy samples and both low- and
high-risk HPV-infected patients. By further analysis of
longitudinal samples, the correlation between levels of
alpha-actinin-4, on one hand, and viral clearing and persistence,
on the other hand, were demonstrated, allowing for 18 pg/ml levels
as discrimination frame (122).
A separate study utilizing a quantitative
proteomics-based approach compared the protein expression profiles
in two HPV+ head and neck cancer cell lines and one
HPV− head and neck cancer cell line, leading to the
identification of 155 differently expressed proteins. In the
HPV+ cell lines, prostate stem cell antigen (PSCA) was
upregulated, whereas the eukaryotic elongation factor 1 alpha was
downregulated, the statuses of which were confirmed by
immunofluorescence and western blot analyses. In addition, in two
clinical series of HPV+ and HPV− oral cavity
carcinoma cases, differences were shown between PSCA and eukaryotic
elongation factor 1 alpha expression levels (123). The study accordingly led to a
first-time demonstration of the association between the
HPV− status and the PSCA and eukaryotic elongation
factor 1 alpha, indicative of the likelihood of the involvement of
these proteins in carcinogenesis associated with HPV−
infection (123).
Expanding the use of monoclonal antibodies in
several domains, was one of the proteomics approaches with intense
development in new treatments for head and neck cancer. Thus,
cetuximab, a monoclonal antibody inhibitor of epidermal growth
factor receptor, was evaluated in comparison with radiotherapy in
an international randomized trial for assessing the best
therapeutic regimen in loco-regionally advanced SCC of the head and
neck (124). This innovative
approach simultaneously used cetuximab and high-dose radiotherapy
for the loco-regional control of the tumors and induced an
important lifetime prolongation (49 vs. 29.3 months for
radiotherapy alone), without increasing the toxic effects related
to radiotherapy (124). Another
recently developed monoclonal antibody against the
thrombin-sensitive site of the osteopontin molecule was used to
test osteopontin as a possible circulating biomarker in cervical
cancers. In both serum and plasma, the osteopontin level was
significantly higher compared to healthy subjects, and was reported
as having the highest values in stage III–IV. However, as
osteopontin can exist in a truncated form and thus, the target
epitope recognized by the monoclonal antibody may be absent,
further studies are warranted to investigate the mechanisms through
which the entire molecule and/or its fragments play a role in
cervical cancer biology (125).
Proteomics may aid in the elucidatation of a
certain status of the HPV-infected milieu. For instance, in
cervical cancer, oxidative stress can be considered as an
insufficiently exploited factor as regards its role in
HPV-initiated carcinogenesis. Along these lines, complex proteomic
and imagistic approaches were used to investigate certain proteins
involved in the overall redox status in HPV type 16-positive
tissues. Thus, tissue samples from patients with invasive squamous
cervical carcinoma, dysplastic HPV lesions and controls, were
assessed for the expression levels of stress markers, such as
endoplasmic reticulum protein 57 (ERp-57), glutathione
S-transferase (GST), inducible nitric oxide synthase (i-NOS), as
well as thioredoxin reductase 2 (TrxR2) levels. In addition,
chemical modifications (carboxylation and oxidation) were evaluated
in some key proteins recognized for their role in cytoskeletal
structure maintenance and cell differentiation. Hence, a HPV tissue
pattern I was registered for each type of lesion. Specifically, in
dysplastic lesions, ERp57 and GST were highly elevated, whereas
TrxR2 and iNOS were gradually decreased; other factors, such as
cytokeratin 6, actin, cornulin, retinal dehydrogenase and GAPDH
were found to be highly carbonylated in comparison with the
controls (126). The results of
the carboxylation degree could account for protein function
failure, providing hints as to the involvement of stress-related
molecular mechanisms in tumor development. For neoplastic lesions,
ERp57 and GST were also found to be elevated, while at the same
time reduced levels of iNOS were determined (126). Indeed, the assessment of chemical
modification indicated that peptidyl-prolyl cis-trans
isomerase A, ERp57, serpin B3, Annexin 2 and GAPDH were found to be
less oxidized in normal as compared with dysplastic tissues.
Furthermore, it was suggested that HPV type 16e neoplastic
progression may be related to a pro-oxidant environment. However,
the oxidative damage seems to be tightly controlled by tumor cells,
as indicated by the selective reduction of carboxylation on key
pro-survival proteins (e.g., ERp57, Serpin B3 and GAPDH), thus
underlining another defense mechanism initiated by the tumor cells
(126). The link between
oxidative stress, UVB radiation and HPV infection were assessed in
a study where a parallel proteomic approach analyzed the expression
pattern and oxidative alterations of proteins in a HPV type
16-transformed keratinocytes (HK-168) cells following UVB exposure.
Protein expression profiles assessed 21 proteins displaying
differential expression levels in cells exposed to UVB radiation in
comparison with unexposed cells. Furthermore, UVB treatment
revealed five proteins highly carbonylated namely: Alpha-enolase,
heat shock protein 75, Annexin 2, elongation factor Tu and
elongation factor gamma. Therefore, proteomics approaches in this
case suggested that UVB-induced oxidative stress triggers a
carbonylated state of specific proteins, which when combined with
the detected protein modifications in HPV-transformed keratinocytes
may contribute to malignant progression (127).
Hypoxia has also been suggested as a factor that
enhances tumor development and metastasis (128). In some pathologies, the
determination of clinically relevant hypoxia may be used to
classify patients who will benefit from the adjustment of the
hypoxia level during the radiotherapy regimen. It was shown that
the osteopontin level in plasma was associated with tumor hypoxia.
Indeed, in patients with head and neck cancer, the higher the
plasma osteopontin concentration, the poorer the outcome after
radiotherapy. This disadvantage may be resolved by a hypoxia
radiosensitiser nimorazole, as in the case of the DAHANCA 5 trial,
which established that nimorazole significantly improved the
outcome of patients subjected to radiotherapy when compared to
placebo (128). Recently, a group
of 63 patients with head and neck SCC was tested in the DAHANCA-5
trial, where the plasma levels of nimorazole were analyzed by high
pressure liquid chromatography (HPLC). The HPLC data revealed a
linear correlation between the peak plasma level and dose, the mean
peak concentration was 32.2±0.9 µg/ml, the median time of
peak concentration was 60 min and the mean half-life was 3.35±0.09
h. Of note, a statistically significantly longer nimorazole
half-life in men compared with women was registered. Overall, this
study that used the HPLC approach may provide a nimorazole scheme
forecast in patients, with nimorazole being generally well
tolerated (129).
Proteomics approaches have also provided insight
related to the cell status upon viral infection. For instance, 2-D
electrophoresis and MALDI-TOF technology were performed in a study
to elucidate the pattern of interactions between some viral
oncogenes expressed in cells infected with HPV type 16 and proteins
related to cell signaling, apoptosis and cell cycle regulation. The
HaCaT keratinocyte cell line was induced to stably express E7,
encoded by one of the viral oncogenes selectively expressed in
carcinoma cells upon HPV type 16 infection. Proteomics data
revealed that >20 important factors are modulated by E7, being
downregulated (e.g., leukocyte elastase inhibitor) or upregulated
(heat shock 60 kDa protein, catalase and peroxiredoxin 2), thus
influencing cell signaling, apoptosis or cell cycle processes
(130).
Studies on HPV type 16 have been continued with
integrative proteomics approaches linked to E6 oncoproteins and
their targets; E6 from high-risk mucosotropic HPV types can trigger
the degradation of the p53 tumor suppressor and the
apoptosis-inducing Bak protein, through the interaction with the
E6AP ubiquitin-protein ligase. On the contrary, E6 from low-risk
mucosotropic HPV types interacts in a lower affinity manner with
E6AP, but the target proteins are still unknown (131). Additionally, this E6 type,
unlike cutaneous E6 proteins, can induce the degradation of
proteins containing PDZ-binding domain. Importantly, that study
could define the differences and similarities of how HPV-related
mucosal and cutaneous E6 proteins interact with intracellular
targets, such as cellular ubiquitin-protein ligases (131).
Comprehensive tumor profiling was used for the
study of a possible link between HPV infection and relapse risk in
vulvar SCC (VSCC), which befall in two distinct subtypes, one
related to high-risk HPV and one negative for HPV, both
acknowledged to develop through distinct intracellular signaling
pathways. A number of 14 VSCC tumor proteome specimens were
analyzed by liquid-chromatography tandem mass spectrometry followed
by complex protein quantification techniques, the fractionation of
labeled peptides by high-resolution isoelectric focusing and
liquid-chromatography tandem mass spectrometry. As a result of such
complex proteomic inquires, 1,579 proteins were quantified and
further analyzed, in a novel approach, for generating individual
tumor pathway alteration data, HPV multivariate statistics and
relapse status comparisons. Thus, 4 proteins identified as main
classifiers of relapse grade were validated by
immunohistochemistry, two of which were found to be
interferon-regulated and HPV-repressed at the mRNA level. Further,
liquid-chromatography tandem mass spectrometry and
immunohistochemistry identified a subgroup of HPV
negative/relapse-associated tumors. The ubiquitin-proteasome
pathway was further identified to be altered in the high risk
subgroup (132).
A correlation between tumor biology and clinical
consequences dictated by HPV subtype infection is likewise a
challenge in the case of oropharyngeal carcinoma. Through a
standardized global proteomic analysis platform, 2,653 proteins
were identified, out of which 31 were selected on the basis of
expression differences between HPV+, HPV− and
normal epithelium. The emerging protein pattern registered an
involvement of proteins associated with epithelial cell
development, keratinization and extracellular matrix organization
in HPV−, while for HPV+ tumors the results
indicated a profile of proteins involved in DNA initiation,
replication and the cell cycle. Tissue microarray also identified
differences in terms of transcription factors and essential amino
acid dependency based on the presence of HPV, contributing to
deciphering molecular pathways for HPV+/HPV−
oropharyngeal carcinoma suitable for therapy and disease monitoring
(117).
9. HPV markers
Understanding the structure of HPV and the biology
of its infection was difficult until the 1990s, when the first
virus propagation in organotypic cultures was produced, leading to
the discovery of the genomic sequences. Even if, the latest
molecular studies have provided a coherent mechanism of the HPV
gene expression and replications, there are still unanswered
questions regarding the HPV biology (133).
The eight reading frames of each DNA strand are
divided into three parts: The L (late) region responsible
for the codification of structural proteins L1–L2, which are
implicated in the virion assembly; the E (early) region that
encodes proteins E1–E7, which are involved in viral replications;
and the LCR necessary for the DNA transcription and
replication (77,134,135). Each papillomaviruses protein has
specific characteristics, and is designed to support a specific
genetic role. Thus, E1 is a viral protein with a size of 72 kDa and
is involved in viral replication. This viral protein binds to a
specific DNA sequence and through the E2 viral protein, it
assembles into hexameric complexes. This complex formation is
responsible for the initiation of the DNA unwinding that produces
subsequent synthesis of progeny DNA (77). E2 is responsible for viral gene
transcription; therefore, low levels of this protein will activate
transcription from the viral LCR, while high levels of E2 act as a
transcriptional repressor. E2 is also involved in DNA replication.
E4 is associated with viral assembly and release, but also with
inducing the G2 arrest and distinguishing the nuclear domain 10.
Importantly, this protein is active in the late part of the viral
life cycle. E5 is implicated in inducing unscheduled cell
proliferation, it activates the protein kinases and growth factor
signaling, but it also inhibits cell apoptosis and the circulation
of major histocompatibility complexes to the surface of cells
(77). The E6 viral protein plays
a role in inducing DNA synthesis, telomerase activity, cell
polarity and motility, as well as in the regulation of
transcriptional co-activators and tumor suppressors together with
E7. The last papillomavirus protein is implicated in unscheduled
cell proliferation and interacts with both negative regulators of
the cell cycle and with histone acetyl transferases (77). As regards the late structural
protein, L1 (major viral structural protein) and L2 (minor viral
structural protein), they are involved in encoding neutralizing
epitopes, but also in assembling the capsomers and the capsid.
Indeed, these proteins interact interact with other cell receptors)
and facilitate virion assembly, interacting with both DNA and
nuclear domain 10 (L2).
Importantly, HPV proteins are useful as markers.
The past or present HPV infections can be confirmed after a serious
epidemiological research, the antibody response to the HPV capsid
being a predictable marker for the cumulative exposure to HPV. The
cervical or oropharyngeal HPV-induced cancers are characterized by
the presence of E6 (the main viral effector E6 gene) and E7
antibodies (108). The
development of SCC is produced by genetic alterations, such as
mutations and copy number alterations at the level of DNA, but also
by epigenetic changes characterized by the aberrant methylation and
histone genetic modifications (90).
However, even if the E6 and E7 proteins antibodies
are markers for HPV type 16- and 18-induced cancer, not all
patients with malignant oropharyngeal disease express these
antibodies, and thus they cannot be used as diagnostic markers
(137). There are epidemiological
studies in progress, in order to further examine the correlation
between HPV-induced cancer and the presence of E6 and E7 proteins
antibodies by either peptide ELISA or western blot analysis
(130). Other researches have
indicated that there is a correlation between cervical cancer and
the presence of E2 and E4 antibodies or some specific linear
sequences of these proteins (138).
HPV type 16 is considered to be the most prevalent
viral genotype implicated in head and neck SCCs. Studies have
suggested that SCC associated with HPV DNA positive/RNA positive
has a better survival expectation in comparison to carcinomas
induced by HPV DNA-positive/RNA-negative or HPV-negative malignant
tumor (137,138). Head and neck cancers induced by
HPV have an HPV-mediated DNA methylation in tumor cells (107). DNA methylation is an epigenetic
mechanism that is characterized by the addition of methyl (CH3) to
cytosine. This process is an important regulator of gene
transcription being controlled by an enzyme family known as DNA
methyltransferases (DNMTs). Furthermore, hypermethylation is most
predominant at the level of TSS 200 promoter regions, but also at
the gene bodies, both of these being good markers of gene
silencing. It is well known that an important role in cellular
transformation is played by the hypermethylation of the promoter
region of tumor suppressor genes (107).
There are data available that have correlated
serine/threonine-protein kinase SMG-1 (involved in both mRNA
surveillance and genotoxic stress response pathways) with
oropharyngeal SCC. SMG-1 is underexpressed in HPV-positive head and
neck cancers in comparison with HPV-negative ones. It has been
demonstrated that HPV type 16 E6 and E7 induce
SMG-1 promoter hypermethylation (113). Other tumor suppressor genes
implicated in carcinogenesis and hypermethylation are various
cadherin subtypes, such as E-cadherin (CDH1), T-cadherin
(CDH13) and proto-cadherin 10 (PCDH10) (108). Recent studies had indicated the
involvement of CDH8 and CDH13, both determined to be
hypermethylated in cervical cancer, in HPV-induced SCCs together
with CDH18, CDH19, CDH23, PCDH10,
PCDH15, PCDHB1, PCDHB4 and PCDHB15
(108). Specifically, the
expression and methylation status of CDH8, PCDH10 and
PCDHB11 can potentially be used to discriminate between
HPV+ and HPV− induced SCC, as well as in
establishing a precise treatment response and in evaluating the
evolution of the disease (108).
Moreover, a decreased E-cadherin expression and subsequent impaired
immune response is also related to HPV infections (108).
CDKN2A or cyclin-dependent kinase inhibitor
2A is a gene located at chromosome 9, band p21.3. It codes for two
proteins, among which INK 4 family member p16 (p16 INK4a) and
p14arf have been annotated the roles of tumor suppressors. This
kinase was found to be overexpressed in carcinomas induced by HPV
and to be reduced in HPV-negative tumors. There is evidence of a
strong correlation between increased levels of HPV E6/E7
transcripts with significant viral loads and high expression levels
of CDKN2A (p16INK4A). SCCs HPV+ can be the result
of a CDKN2A promoter hypermethylation and/or loss of 9p21.
Of note, CDKN2A is the second most commonly activated gene
in carcinogenesis after p53 (110). Furthermore, the expression of
genes CDKN2A, CDKN2B and CDKN2C was also
associated with HPV+ malignant tumors together with
genes involved in the cell-based immune response (CxCL,
CXCL11 and IL-19) (110,112).
Another potential biomarker in HPV
infection-related carcinomas may be the mitochondrial DNA of the
patient. Thus, it has been shown that a large number of patients
suffering from head and neck SCC had mitochondrial DNA mutations
(139). The association between
HPV infection and the mutagenesis of mitochondrial DNA is currently
under investigation. Of note, the quantitative determination of
mitochondrial DNA has proven to be a valuable indicator of local
recurrence and distant metastases in SCC (135,140).
Another line of research has demonstrated that
there are connections between HPV-induced cancer and HPV
chromosomal alterations, such as gains at 20p13–q13.33 and losses
at 13q21.1–21.33. On the other side, lost regions at 3p and 5q
together with amplifications at 11q13.3 were common in HPV-negative
carcinomas and absent in HPV-induced tumors. Indeed, these low
level gains of chromosome 20q can be produced by E7 expression and
the consequent inactivation of the retinoblastoma protein pathway
in epithelial cells (120).
Furthermore, head and neck tumors, but also cervical and anal
carcinomas induced by HPV may be associated with the deletion of
part of the long arm of chromosome 13 (120).
Some researchers have demonstrated the presence of
specific genes at 20q and 13q chromosomes, underlying the
importance of these related genes in HPV-mediated carcinogenesis.
Indeed, they seem to be associated with continuous E7-regulated
E2F1 activation, determining changes in cellular maintenance
systems (nucleic acid metabolism). Furthermore, at the level of 20q
region, cancer-related genes, such as E2F1, PIGU
(phosphatidylinositol glycan anchor biosynthesis, class U-involved
in cell cycle control and associated with bladder cancer) and
DNMT3B (DNA cytosine 5-methyltrans-ferase 3 beta) which is a
de novo DNA methyl transferase (involved in cervical cancer)
have been localized (120). There
are also gains at 3q and losses at 11q associated with epithelial
SCCs (120). PPFIA1,
DHCR7, CTTN, TMEM16A and GAL genes are
located in the 11q13 region which is affected in HPV-related
tumors. For this reason, their expression in HPV+
carcinomas is decreased together with MAGEA4, MAGEA12
and HMGA2 (120). As
regards the detection of HPV capsid antibody, it was demonstrated
that with the exception of HPV type 6, which contains shared
epitopes and type-specific epitopes on intact capsids,
conformationally dependent epitopes on intact capsids are HPV-type
specific (136,153).
10. HPV treatment and cancer
The treatment of head and neck SCC caused by HPV
infection depends on the presence or absence of the HPV load in the
tumors. It is considered that in the case of HPV-negative tumors,
the prognosis is better as compared to that in HPV-positive ones,
where the treatment is focused on improving the quality of life and
the survival rate of patients (141). The treatment is specific
according to the type and stage of the tumor. In the case of SCC
and depending on tumor spread surgical resection of the tumor,
chemotherapy, as well as radiotherapy may be necessary. For stage
one and two tumors, external or internal radiotherapy is used as an
elective treatment. Aggressive carcinomas can sometimes mimic
benign lesions, but follow-up during evolution, as well as a poor
response to treatment eventually characterize their biological
behavior (142). As regards stage
three and four tumors, the standard treatment is platinum salt
chemotherapy combined with radiotherapy (143). In advanced forms of cancers,
former platinum-based and fluorouracil regimens have been replaced
with combinations of cetuximab and platinum salts that have
demonstrated their superior efficacy (144).
Cervical cancer remains the most widespread form of
cancer induced by the HPV virus and the fourth most common type of
malignancy affecting females. There is no proper treatment of
genital HPV infection. In the majority of women, the infection
heals alone due to immune system activity. The treatments available
for HPV genital infection are targeted at changes in the skin and
mucosa caused by HPV infection such as condylomas, pre-cancerous
lesions in the cervix or cervical cancer. Pre-cancerous lesions
that bleed may over-infect with pathogenic germs (145) or fungus (Aspergillus or
Fusarium genus) (146)
which are difficult to treat. As alternative adjuvant therapy,
various chemical compounds with anti-bacterial and
anti-proliferative effects (147,148) or plant extracts with cytotoxic
and apoptotic effects may be administered (149). All of these therapeutic methods
can remove the lesions, but do not remove the virus from the body.
It can be eliminated exclusively by the action of the immune
system. As the virus persists in the body, condylomas often relapse
following treatment. Since the discovery of HPV, a need to
formulate a vaccine to prevent infection and, subsequently, the
development of cancer has existed. The vaccine mimics the disease
and creates resistance. It is not a live virus or inactivated virus
vaccine and thus it does not represent any risk for the individuals
that use it (150).
The antipapillomavirus vaccines
In natural infection, HPV stimulates both humoral
immunity (mediated by monoclonal antibodies), as well as
cell-mediated immunity (mediated by T lymphocytes) in the human
body. The body synthesizes antibodies to surface proteins L1 and
L2; however, the intensity of the immune response varies from
individual to individual and depends on the HPV type that causes
the infection and on its persistence. Thus, the development of
anti-HPV vaccines began after careful observations as to how the
human body reacts as a result of infection with HPV (150). Studies on the vaccines began in
the 1980s, and the results materialized only in 2006, when the two
vaccines containing L1 viral proteins produced by genetic
recombination techniques appeared. The proteins are assembled into
morphologically similar particles to the viral, but do not contain
the genome (DNA-viral double-stranded DNA molecule). Thus, these
vaccines do not present infectious or oncogenic risk, as they only
stimulate specific antibodies (151,152). The objectives of the vaccination
are (151): i) Providing
prophylaxis of HPV infection by stimulating mediated humoral
immunity and preventing the virus from reaching the target
(keratinocytes); and ii) stimulation of cell-mediated immunity (T
helper lymphocytes) as the body must destroy the abnormal
keratinocytes and eliminate the lesions induced by infection.
Due to ethical reasons, the rate of cervical cancer
development or the mortality rate cannot be analyzed through
long-term studies; thus, the final stages of CIN investigated were
CIN2 and CIN3. It is ideal to administer the vaccine prior to the
commencement of sexual activities. The vaccination does not protect
patients already infected with any of the strains studied (types 6,
11, 16, 18 and 16, 18), but the tetravalent vaccine can prevent
(44%) cervical lesions in patients who have been previously
infected and received only one or two of the three required doses
of vaccine. The mechanisms of action of the HPV vaccines are
presented in Fig. 2.
The tetravalent vaccine (Gardasil or Silgard; Merck
& Co., Whitehouse Station, NJ, USA) was approved in June 2006
for women aged between 19–26 years to prevent cervical, vulvar and
vaginal cancer caused by HPV types 16 and 18, and genital lesions
caused by HPV types 6, 11, 16 and 18. The bivalent vaccine
(Cervarix; GlaxoSmithKline Biologicals, Rockville, MD, USA) was
approved in the USA only in October 2009, two years later than in
the rest of the countries. Gardasil is a prophylactic vaccine that
prevents intraepithelial cervical neoplasia, intraepithelial vulvar
neoplasia and intraepithelial vaginal neoplasia, recommended to be
administered prior to sexual intercourse. The double-blind clinical
trial required for the approval of the vaccine was performed on
12,000 women aged between 16 and 26 years, in thirteen countries,
and studies are currently being conducted to approve the vaccine
for women aged between 27 and 45 years (153). An important aspect is that this
vaccine is also effective for men, protecting them from
pre-cancerous lesions or anal cancer caused by some HPV strains.
Its use for this purpose has been permitted since October 2009 (the
age category for men is 9–15 years). The vaccine dose is 0.5 ml
administered intramuscularly (never intravenously) at 2 and 4
months, respectively, from the previous dose (3 in total) and
contains the following: 20 µg of L1 type 6 protein, 40
µg of L1 type 11 protein, 40 µg L1 type 16 proteins
and 20 µg of L1 type 18 protein. The L1 protein is in the
form of virus-like particles produced on yeast cells,
Saccharomyces cerevisiae strain 1895, by recombinant DNA
technology (152). The vaccine’s
effectiveness against HPV infection is almost 100% for strains 6,
11, 16 and 18 over a period of >4 years. In addition, it has
been shown to have an affect (38%) on another 10 strains
responsible for carcinogenesis. The reported side-effects range
from swelling of the area of administration, local pain (stronger
than with other vaccines), syncope, Guillain-Barre syndrome (acute
demyelinating inflammatory polyneuropathy triggered by acute
infectious disease) to pulmonary embolism and death 1:1,000,000,
with approximately 20 cases out of total doses administered
worldwide. The vaccine does not contain mercury, thiomersal (high
potency toxic organomercury compound), antibiotics or viral genome
traces, but contains 50 mcg of polysorbate 80 that can cause
infertility, being one of the main reasons why its use is not
universally accepted. Unfortunately, as a new vaccine, long-term
effects (including the effects on pregnancy/breastfeeding) are not
yet known; however, from the current perspective, the benefits
(both pathological and financial) are greater than the risks of
using it (154).
Cervarix (Merck & Co.) is a bivalent vaccine
designed to prevent HPV types 18 and 16 infection (the latter is
also associated with oropharyngeal cancers), clinical tests showing
that it also offers some protection against strains 45 and 31
(155). The vaccine contains 20
µg of L1 type 16 protein, 20 µg of L1 type 18
protein, 50 µg of AS04 3-O-desacil-4′-monophosphoryl lipid A
(MPL) adjuvant which generates and maintains longer immune response
to the body, absorbed on aluminum hydroxide hydrate (0.5 mg of
aluminum). It is not recommended for patients with severe fever or
immunodeficiency (these contraindications also apply to Gardasil).
Compared to Gardasil, Cervarix generates twice as many anti-HPV 16
antibodies and 6-fold more anti-HPV 18 antibodies at 7 months
post-dosing, but was not active on strains 6 and 11 responsible for
genital warts or in the case of male patients (156). In spite of these findings and
recent studies, anti-HPV vaccination remains an open subject
because of multiple unclear aspects: Long-term side effects,
duration of immunogenicity, effective efficacy of male patients
and, last but not least, the effects on the Papanicolau test or
colposcopy (the intervals between the tests could increase.
11. Conclusions
HPV infections are strongly implicated in the
pathogenesis of different types of cancer. The screening for HPV is
necessary in carcinoma therapy determination and prognosis
evaluation.
Notes
[1] Competing
interests
Demetrios A. Spandidos is the Editor-in-Chief for
the journal, but had no personal involvement in the reviewing
process, or any influence in terms of adjudicating on the final
decision, for this article.
References
|
1
|
zur Hausen H: Papillomaviruses and cancer:
From basic studies to clinical application. Nat Rev Cancer.
2:342–350. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ramqvist T, Grün N and Dalianis T: Human
papillomavirus and tonsillar and base of tongue cancer. Viruses.
7:1332–1343. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lacey CJ, Lowndes CM and Shah KV: Chapter
4: Burden and management of non-cancerous HPV-related conditions:
HPV-6/11 disease. Vaccine. 24(Suppl 3): S3/35–41. 2006. View Article : Google Scholar
|
|
4
|
Cogliano V, Baan R, Straif K, Grosse Y,
Secretan B and El Ghissassi F; WHO International Agency for
Research on Cancer: Carcinogenicity of human papillomaviruses.
Lancet Oncol. 6:2042005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Clifford GM, Smith JS, Plummer M, Muñoz N
and Franceschi S: Human papillomavirus types in invasive cervical
cancer worldwide: A meta-analysis. Br J Cancer. 88:63–73. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mammas IN, Sourvinos G and Spandidos DA:
The paediatric story of human papillomavirus (Review). Oncol Lett.
8:502–506. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mammas IN, Vageli D and Spandidos DA:
Geographic variations of human papilloma virus infection and their
possible impact on the effectiveness of the vaccination programme.
Oncol Rep. 20:141–145. 2008.PubMed/NCBI
|
|
8
|
Workowski KA and Bolan GA; Centers for
Disease Control and Prevention: Sexually transmitted diseases
treatment guidelines, 2015. MMWR Recomm Rep. 64(No. RR-3): pp.
1–137. 2015
|
|
9
|
Workowski KA and Berman S; Centers for
Disease Control and Prevention (CDC): Sexually transmitted diseases
treatment guidelines, 2010. MMWR Recomm Rep. 59:1–110.
2010.PubMed/NCBI
|
|
10
|
de Villiers EM, Fauquet C, Broker TR,
Bernard HU and zur Hausen H: Classification of papillomaviruses.
Virology. 324:17–27. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stokley S, Jeyarajah J, Yankey D, Cano M,
Gee J, Roark J, Curtis RC and Markowitz L; Immunization Services
Division, National Center for Immunization and Respiratory
Diseases, CDC: Human papillomavirus vaccination coverage among
adolescents, 2007–2013, and postlicensure vaccine safety
monitoring, 2006–2014-United States. MMWR Morb Mortal Wkly Rep.
63:620–624. 2014.PubMed/NCBI
|
|
12
|
Chaturvedi AK, Engels EA, Pfeiffer RM,
Hernandez BY, Xiao W, Kim E, Jiang B, Goodman MT, Sibug-Saber M,
Cozen W, et al: Human papillomavirus and rising oropharyngeal
cancer incidence in the United States. J Clin Oncol. 29:4294–4301.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Castle PE, Schiffman M, Herrero R,
Hildesheim A, Rodriguez AC, Bratti MC, Sherman ME, Wacholder S,
Tarone R and Burk RD: A prospective study of age trends in cervical
human papillomavirus acquisition and persistence in Guanacaste,
Costa Rica. J Infect Dis. 191:1808–1816. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sedlacek TV, Lindheim S, Eder C, Hasty L,
Woodland M, Ludomirsky A and Rando RF: Mechanism for human
papillomavirus transmission at birth. Am J Obstet Gynecol.
161:55–59. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Roman A and Fife K: Human papillomavirus
DNA associated with foreskins of normal newborns. J Infect Dis.
153:855–861. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rintala MA, Grénman SE, Järvenkylä ME,
Syrjänen KJ and Syrjänen SM: High-risk types of human
papillomavirus (HPV) DNA in oral and genital mucosa of infants
during their first 3 years of life: Experience from the Finnish HPV
Family Study. Clin Infect Dis. 41:1728–1733. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Boda D, Neagu M, Constantin C, Voinescu
RN, Caruntu C, Zurac S, Spandidos DA, Drakoulis N, Tsoukalas D and
Tsatsakis AM: HPV strain distribution in patients with genital
warts in a female population sample. Oncol Lett. 12:1779–1782.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen R, Sehr P, Waterboer T, Leivo I,
Pawlita M, Vaheri A and Aaltonen LM: Presence of DNA of human
papillomavirus 16 but no other types in tumor-free tonsillar
tissue. J Clin Microbiol. 43:1408–1410. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Powell J, Strauss S, Gray J and
Wojnarowska F: Genital carriage of human papilloma virus (HPV) DNA
in prepubertal girls with and without vulval disease. Pediatr
Dermatol. 20:191–194. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
De Marco F: Oxidative stress and HPV
carcinogenesis. Viruses. 5:708–731. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Căruntu C, Boda D, Musat S, Căruntu A and
Mandache E: Stress-induced mast cell activation in glabrous and
hairy skin. Mediators Inflamm. 2014:1059502014. View Article : Google Scholar :
|
|
22
|
Căruntu C, Grigore C, Căruntu A,
Diaconeasa A and Boda D: The role of stress in skin disease. Intern
Med. 8:73–84. 2003.
|
|
23
|
Fang CY, Miller SM, Bovbjerg DH, Bergman
C, Edelson MI, Rosenblum NG, Bove BA, Godwin AK, Campbell DE and
Douglas SD: Perceived stress is associated with impaired T-cell
response to HPV16 in women with cervical dysplasia. Ann Behav Med.
35:87–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Leemans CR, Braakhuis BJ and Brakenhoff
RH: The molecular biology of head and neck cancer. Nat Rev Cancer.
11:9–22. 2011. View Article : Google Scholar
|
|
25
|
Gillison ML, Castellsagué X, Chaturvedi A,
Goodman MT, Snijders P, Tommasino M, Arbyn M and Franceschi S:
Eurogin Roadmap: Comparative epidemiology of HPV infection and
associated cancers of the head and neck and cervix. Int J Cancer.
134:497–507. 2014. View Article : Google Scholar
|
|
26
|
Giuliano AR, Nyitray AG, Kreimer AR,
Pierce Campbell CM, Goodman MT, Sudenga SL, Monsonego J and
Franceschi S: EUROGIN 2014 roadmap: Differences in human
papillomavirus infection natural history, transmission and human
papillomavirus-related cancer incidence by gender and anatomic site
of infection. Int J Cancer. 136:2752–2760. 2015. View Article : Google Scholar
|
|
27
|
Ang KK, Harris J, Wheeler R, Weber R,
Rosenthal DI, Nguyen-Tân PF, Westra WH, Chung CH, Jordan RC, Lu C,
et al: Human papillomavirus and survival of patients with
oropharyngeal cancer. N Engl J Med. 363:24–35. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gillison ML, Alemany L, Snijders PJ,
Chaturvedi A, Steinberg BM, Schwartz S and Castellsagué X: Human
papillomavirus and diseases of the upper airway: Head and neck
cancer and respiratory papillomatosis. Vaccine. 30(Suppl 5):
F34–F54. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Doorbar J, Quint W, Banks L, Bravo IG,
Stoler M, Broker TR and Stanley MA: The biology and life-cycle of
human papillomaviruses. Vaccine. 30(Suppl 5): F55–F70. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sepiashvili L, Bruce JP, Huang SH,
O’Sullivan B, Liu FF and Kislinger T: Novel insights into head and
neck cancer using next-generation ‘omic’ technologies. Cancer Res.
75:480–486. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Psyrri A, Rampias T and Vermorken JB: The
current and future impact of human papillomavirus on treatment of
squamous cell carcinoma of the head and neck. Ann Oncol.
25:2101–2115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ghita MA, Caruntu C, Lixandru D, Pitea A,
Batani A and Boda D: The quest for novel biomarkers in early
diagnosis of diabetic neuropathy. Curr Proteomics. 14:86–99. 2017.
View Article : Google Scholar
|
|
33
|
Hoadley KA, Yau C, Wolf DM, Cherniack AD,
Tamborero D, Ng S, Leiserson MDM, Niu B, McLellan MD, Uzunangelov
V, et al Cancer Genome Atlas Research Network: Multiplatform
analysis of 12 cancer types reveals molecular classification within
and across tissues of origin. Cell. 158:929–944. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Termine N, Giovannelli L, Matranga D,
Caleca MP, Bellavia C, Perino A and Campisi G: Oral human
papillomavirus infection in women with cervical HPV infection: New
data from an Italian cohort and a metanalysis of the literature.
Oral Oncol. 47:244–250. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hakenberg OW, Compérat EM, Minhas S,
Necchi A, Protzel C and Watkin N; European Association of Urology:
EAU guidelines on penile cancer: 2014 update. Eur Urol. 67:142–150.
2015. View Article : Google Scholar
|
|
36
|
Lupu M, Caruntu A, Caruntu C, Papagheorghe
LML, Ilie MA, Voiculescu V, Boda D, Constantin C, Tanase C, Sifaki
M, et al: Neuroendocrine factors: The missing link in non melanoma
skin cancer (Review). Oncol Rep. 38:1327–1340. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Spiess PE, Dhillon J, Baumgarten AS,
Johnstone PA and Giuliano AR: Pathophysiological basis of human
papillomavirus in penile cancer: Key to prevention and delivery of
more effective therapies. CA Cancer J Clin. 66:481–495. 2016.
View Article : Google Scholar
|
|
38
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kayes O, Ahmed HU, Arya M and Minhas S:
Molecular and genetic pathways in penile cancer. Lancet Oncol.
8:420–429. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Protzel C and Spiess PE: Molecular
research in penile cancer-lessons learned from the past and bright
horizons of the future? Int J Mol Sci. 14:19494–19505. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Schiller JT, Day PM and Kines RC: Current
understanding of the mechanism of HPV infection. Gynecol Oncol.
118(Suppl): S12–S17. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Steben M and Duarte-Franco E: Human
papillomavirus infection: Epidemiology and pathophysiology. Gynecol
Oncol. 107(Suppl 1): S2–S5. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Fuchs E and Nowak JA: Building epithelial
tissues from skin stem cells. Cold Spring Harb Symp Quant Biol.
73:333–350. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ledwaba T, Dlamini Z, Naicker S and Bhoola
K: Molecular genetics of human cervical cancer: Role of
papillomavirus and the apoptotic cascade. Biol Chem. 385:671–682.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Neagu M, Căruntu C, Constantin C, Boda D,
Zurac S, Spandidos DA and Tsatsakis AM: Chemically induced skin
carcinogenesis: Updates in experimental models (Review). Oncol Rep.
35:2516–2528. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Castellsagué X, Bosch FX and Muñoz N:
Environmental co-factors in HPV carcinogenesis. Virus Res.
89:191–199. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bosch FX and Muñoz N: The viral etiology
of cervical cancer. Virus Res. 89:183–190. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Satterwhite CL, Torrone E, Meites E, Dunne
EF, Mahajan R, Ocfemia MC, Su J, Xu F and Weinstock H: Sexually
transmitted infections among US women and men: Prevalence and
incidence estimates, 2008. Sex Transm Dis. 40:187–193. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nielsen A, Kjaer SK, Munk C and Iftner T:
Type-specific HPV infection and multiple HPV types: Prevalence and
risk factor profile in nearly 12,000 younger and older Danish
women. Sex Transm Dis. 35:276–282. 2008. View Article : Google Scholar
|
|
50
|
Skaaby S and Kofoed K: Anogenital warts in
Danish men who have sex with men. Int J STD AIDS. 22:214–217. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Owusu-Edusei K Jr, Chesson HW, Gift TL,
Tao G, Mahajan R, Ocfemia MC and Kent CK: The estimated direct
medical cost of selected sexually transmitted infections in the
United States, 2008. Sex Transm Dis. 40:197–201. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
World Health Organization: Implementation
of the global strategy for prevention and control of sexually
transmitted infections: 2006–2015. http://apps.who.int/iris/bitstream/10665/183117/1/9789241508841_eng.pdf.
Accessed Dec 21, 2015.
|
|
53
|
de Sanjosé S, Diaz M, Castellsagué X,
Clifford G, Bruni L, Muñoz N and Bosch FX: Worldwide prevalence and
genotype distribution of cervical human papillomavirus DNA in women
with normal cytology: A meta-analysis. Lancet Infect Dis.
7:453–459. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Newman L, Rowley J, Vander Hoorn S,
Wijesooriya NS, Unemo M, Low N, Stevens G, Gottlieb S, Kiarie J and
Temmerman M: Global estimates of the prevalence and incidence of
four curable sexually transmitted infections in 2012 based on
systematic review and global reporting. PLoS One. 10:e01433042015.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
McQuillan G, Kruszon-Moran D, Markowitz
LE, Unger ER and Paulose-Ram R: Prevalence of HPV in adults aged
18–69: United States, 2011–2014. NCHS Data Brief. (280): 1–8.
2017.PubMed/NCBI
|
|
56
|
Braaten KP and Laufer MR: Human
papillomavirus (HPV), HPV-related disease, and the HPV vaccine. Rev
Obstet Gynecol. 1:2–10. 2008.PubMed/NCBI
|
|
57
|
Dunne EF and Markowitz LE: Genital human
papillomavirus infection. Clin Infect Dis. 43:624–629. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Cai T, Wagenlehner FM, Mondaini N, D’Elia
C, Meacci F, Migno S, Malossini G, Mazzoli S and Bartoletti R:
Effect of human papillomavirus and Chlamydia trachomatis
co-infection on sperm quality in young heterosexual men with
chronic prostatitis-related symptoms. BJU Int. 113:281–287. 2014.
View Article : Google Scholar
|
|
59
|
Panatto D, Amicizia D, Bianchi S, Frati
ER, Zotti CM, Lai PL, Domnich A, Colzani D, Gasparini R and Tanzi
E: Chlamydia trachomatis prevalence and chlamydial/HPV co-infection
among HPV-unvaccinated young Italian females with normal cytology.
Hum Vaccin Immunother. 11:270–276. 2015. View Article : Google Scholar :
|
|
60
|
Smith JS, Muñoz N, Herrero R, Eluf-Neto J,
Ngelangel C, Franceschi S, Bosch FX, Walboomers JMM and Peeling RW:
Evidence for Chlamydia trachomatis as a human papillomavirus
cofactor in the etiology of invasive cervical cancer in Brazil and
the Philippines. J Infect Dis. 185:324–331. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Tarallo PA, Smolowitz J, Carriero D,
Tarallo J, Siegel A, Jia H and Emond JC: Prevalence of high-risk
human papilloma virus among women with hepatitis C virus before
liver transplantation. Transpl Infect Dis. 15:400–404. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Smith JS, Herrero R, Bosetti C, Muñoz N,
Bosch FX, Eluf-Neto J, Castellsagué X, Meijer CJ, Van den Brule AJ,
Franceschi S and Ashley R; International Agency for Research on
Cancer (IARC) Multicentric Cervical Cancer Study Group: Herpes
simplex virus-2 as a human papillomavirus cofactor in the etiology
of invasive cervical cancer. J Natl Cancer Inst. 94:1604–1613.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Global Burden of Disease Study 2013
Collaborators: Global, regional, and national incidence,
prevalence, and years lived with disability for 301 acute and
chronic diseases and injuries in 188 countries, 1990–2013: A
systematic analysis for the Global Burden of Disease Study 2013.
Lancet. 386:743–800. 2013.
|
|
64
|
Alberts CJ, Schim van der Loeff MF,
Papenfuss MR, da Silva RJC, Villa LL, Lazcano-Ponce E, Nyitray AG
and Giuliano AR: Association of Chlamydia trachomatis infection and
herpes simplex virus type 2 serostatus with genital human
papillomavirus infection in men: The HPV in men study. Sex Transm
Dis. 40:508–515. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Mbulawa ZZ, Coetzee D, Marais DJ, Kamupira
M, Zwane E, Allan B, Constant D, Moodley JR, Hoffman M and
Williamson AL: Genital human papillomavirus prevalence and human
papillomavirus concordance in heterosexual couples are positively
associated with human immunodeficiency virus coinfection. J Infect
Dis. 199:1514–1524. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
66
|
Bruni L, Diaz M, Castellsagué X, Ferrer E,
Bosch FX and de Sanjosé S: Cervical human papillomavirus prevalence
in 5 continents: Meta-analysis of 1 million women with normal
cytological findings. J Infect Dis. 12:1789–1799. 2010. View Article : Google Scholar
|
|
67
|
Clifford GM, Gonçalves MA and Franceschi
S; HPV and HIV Study Group: Human papillomavirus types among women
infected with HIV: A meta-analysis. AIDS. 20:2337–2344. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Koshiol J, Lindsay L, Pimenta JM, Poole C,
Jenkins D and Smith JS: Persistent human papillomavirus infection
and cervical neoplasia: A systematic review and meta-analysis. Am J
Epidemiol. 168:123–137. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
van Rijn VM, Mooij SH, Mollers M, Snijders
PJF, Speksnijder AGCL, King AJ, de Vries HJ, van Eeden A, van der
Klis FR, de Melker HE, et al: Anal, penile, and oral high-risk HPV
infections and HPV seropositivity in HIV-positive and HIV-negative
men who have sex with men. PLoS One. 9:e922082014. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kjær SK, Frederiksen K, Munk C and Iftner
T: Long-term absolute risk of cervical intraepithelial neoplasia
grade 3 or worse following human papillomavirus infection: Role of
persistence. J Natl Cancer Inst. 102:1478–1488. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Nielsen A, Kjaer SK, Munk C, Osler M and
Iftner T: Persistence of high-risk human papillomavirus infection
in a population-based cohort of Danish women. J Med Virol.
82:616–23. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Fukuchi E, Sawaya GF, Chirenje M, Magure
T, Tuveson J, Ma Y, Shiboski S, Da Costa M, Palefsky J, Moscicki
AB, et al: Cervical human papillomavirus incidence and persistence
in a cohort of HIV-negative women in Zimbabwe. Sex Transm Dis.
36:305–311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Paaso AE, Louvanto K, Syrjänen KJ,
Waterboer T, Grénman SE, Pawlita M and Syrjänen SM: Lack of
type-specific concordance between human papillomavirus (HPV)
serology and HPV DNA detection in the uterine cervix and oral
mucosa. J Gen Virol. 92:2034–2046. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Syrjänen K, Shabalova I, Naud P,
Kozachenko V, Derchain S, Zakharchenko S, Roteli-Martins C,
Nerovjna R, Longatto-Filho A, Kljukina L, et al New Independent
States of the Former Soviet Union and the Latin American Screening
Study Research Groups: Persistent high-risk human papillomavirus
infections and other end-point markers of progressive cervical
disease among women prospectively followed up in the New
Independent States of the Former Soviet Union and the Latin
American Screening study cohorts. Int J Gynecol Cancer. 19:934–942.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Syrjänen K, Shabalova I, Naud P,
Kozachenko V, Derchain S, Zakharchenko S, Roteli-Martins C,
Nerovjna R, Longatto-Filho A, Kljukina L, et al; NIS and LAMS Study
Research Groups. Risk estimates for persistent high-risk human
papillomavirus infections as surrogate endpoints of progressive
cervical disease critically depend on reference category: Analysis
of the combined prospective cohort of the New Independent States of
the Former Soviet Union and Latin American Screening studies. Int J
STD AIDS. 22:315–323. 2011. View Article : Google Scholar
|
|
76
|
King EM, Gilson R, Beddows S, Soldan K,
Panwar K, Young C, Jit M, Edmunds WJ and Sonnenberg P: Oral human
papillomavirus (HPV) infection in men who have sex with men:
Prevalence and lack of anogenital concordance. Sex Transm Infect.
91:284–286. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Lyronis ID, Baritaki S, Bizakis I,
Krambovitis E and Spandidos DA: K-ras mutation, HPV infection and
smoking or alcohol abuse positively correlate with esophageal
squamous carcinoma. Pathol Oncol Res. 14:267–273. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zaravinos A, Bizakis J and Spandidos DA:
Prevalence of human papilloma virus and human herpes virus types
1–7 in human nasal polyposis. J Med Virol. 81:1613–1619. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Du J, Nordfors C, Ahrlund-Richter A,
Sobkowiak M, Romanitan M, Näsman A, Andersson S, Ramqvist T and
Dalianis T: Prevalence of oral human papillomavirus infection among
youth, Sweden. Emerg Infect Dis. 18:1468–1471. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Nielsen A, Kjaer SK, Munk C, Osler M and
Iftner T: Persistence of high-risk human papillomavirus infection
in a population-based cohort of Danish women. J Med Virol.
82:616–623. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Rosa MI, Fachel JM, Rosa DD, Medeiros LR,
Igansi CN and Bozzetti MC: Persistence and clearance of human
papillomavirus infection: A prospective cohort study. Am J Obstet
Gynecol. 199:617.e1–617.e7. 2008. View Article : Google Scholar
|
|
82
|
Koskimaa HM, Waterboer T, Pawlita M,
Grénman S, Syrjänen K and Syrjänen S: Human papillomavirus
genotypes present in the oral mucosa of newborns and their
concordance with maternal cervical human papillomavirus genotypes.
J Pediatr. 160:837–843. 2012. View Article : Google Scholar
|
|
83
|
Kofoed K, Sand C, Forslund O and Madsen K:
Prevalence of human papillomavirus in anal and oral sites among
patients with genital warts. Acta Derm Venereol. 94:207–211. 2014.
View Article : Google Scholar
|
|
84
|
Jang MK, Anderson DE, van Doorslaer K and
McBride AA: A proteomic approach to discover and compare
interacting partners of papillomavirus E2 proteins from diverse
phylogenetic groups. Proteomics. 15:2038–2050. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Yarbro CH, Wujcik D and Gobel BH: Cancer
Nursing: Principles and Practice. 7th. Jones & Bartlett;
Learning, Burlington, MA: 2010
|
|
86
|
Markowitz LE, Dunne EF, Saraiya M, Chesson
HW, Curtis CR, Gee J, Bocchini JA Jr and Unger ER; Centers for
Disease Control and Prevention (CDC): Human papillomavirus
vaccination: Recommendations of the Advisory Committee on
Immunization Practices (ACIP). MMWR Recomm Rep. 63:1–30.
2014.PubMed/NCBI
|
|
87
|
Clifford GM, Rana RK, Franceschi S, Smith
JS, Gough G and Pimenta JM: Human papillomavirus genotype
distribution in low-grade cervical lesions: Comparison by
geographic region and with cervical cancer. Cancer Epidemiol
Biomarkers Prev. 14:1157–1164. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Clifford GM, Smith JS, Aguado T and
Franceschi S: Comparison of HPV type distribution in high-grade
cervical lesions and cervical cancer: A meta-analysis. Br J Cancer.
89:101–105. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Louvanto K, Rautava J, Willberg J, Wideman
L, Syrjänen K, Grénman S and Syrjänen S: Genotype-specific
incidence and clearance of human papillomavirus in oral mucosa of
women: A six-year follow-up study. PLoS One. 8:e534132013.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Wu DW, Chuang CY, Lin WL, Sung WW, Cheng
YW and Lee H: Paxillin promotes tumor progression and predicts
survival and relapse in oral cavity squamous cell carcinoma by
microRNA-218 targeting. Carcinogenesis. 35:1823–1829. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Martínez-Noël G, Galligan JT, Sowa ME,
Arndt V, Overton TM, Harper JW and Howley PM: Identification and
proteomic analysis of distinct UBE3A/E6AP protein complexes. Mol
Cell Biol. 32:3095–3106. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
McBride AA: The papillomavirus E2
proteins. Virology. 445:57–79. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Jang MK, Shen K and McBride AA:
Papillomavirus genomes associate with BRD4 to replicate at fragile
sites in the host genome. PLoS Pathog. 10:e10041172014. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Bersani C, Mints M, Tertipis N, Haeggblom
L, Sivars L, Ährlund-Richter A, Vlastos A, Smedberg C, Grün N,
Munck-Wikland E, et al: A model using concomitant markers for
predicting outcome in human papillomavirus positive oropharyngeal
cancer. Oral Oncol. 68:53–59. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Grace M and Munger K: Proteomic analysis
of the gamma human papillomavirus type 197 E6 and E7 associated
cellular proteins. Virology. 500:71–81. 2017. View Article : Google Scholar
|
|
96
|
Serafín-Higuera I, Garibay-Cerdenares OL,
Illades-Aguiar B, Flores-Alfaro E, Jiménez-López MA,
Sierra-Martínez P and Alarcón-Romero LC: Differential proteins
among normal cervix cells and cervical cancer cells with HPV-16
infection, through mass spectrometry-based proteomics (2D-DIGE) in
women from Southern México. Proteome Sci. 14:102016. View Article : Google Scholar
|
|
97
|
Koifman L, Ornellas P, Ornellas AA,
Pereira DA, Zingali BR, Cavalcanti SM, Afonso LA, Sandim V and
Alves G: Proteomics analysis of tissue samples from patients with
squamous cell carcinoma of the penis and positive to human
papillomavirus. Int Braz J Urol. 41:642–654. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Tanase CP, Albulescu R and Neagu M:
Application of 3D hydrogel microarrays in molecular diagnostics:
Advantages and limitations. Expert Rev Mol Diagn. 11:461–464. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ewaisha R, Meshay I, Resnik J, Katchman BA
and Anderson KS: Programmable protein arrays for immunoprofiling
HPV-associated cancers. Proteomics. 16:1215–1224. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Arroyo Mühr LS, Hultin E, Bzhalava D,
Eklund C, Lagheden C, Ekström J, Johansson H, Forslund O and
Dillner J: Human papillomavirus type 197 is commonly present in
skin tumors. Int J Cancer. 136:2546–2555. 2015. View Article : Google Scholar
|
|
101
|
Dahlgren L, Mellin H, Wangsa D,
Heselmeyer-Haddad K, Björnestål L, Lindholm J, Munck-Wikland E,
Auer G, Ried T and Dalianis T: Comparative genomic hybridization
analysis of tonsillar cancer reveals a different pattern of genomic
imbalances in human papillomavirus-positive and −negative tumors.
Int J Cancer. 107:244–249. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Mountzios G, Rampias T and Psyrri A: The
mutational spectrum of squamous-cell carcinoma of the head and
neck: Targetable genetic events and clinical impact. Ann Oncol.
25:1889–1900. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Smeets SJ, Braakhuis BJ, Abbas S, Snijders
PJ, Ylstra B, van de Wiel MA, Meijer GA, Leemans CR and Brakenhoff
RH: Genome-wide DNA copy number alterations in head and neck
squamous cell carcinomas with or without oncogene-expressing human
papillomavirus. Oncogene. 25:2558–2564. 2006. View Article : Google Scholar
|
|
104
|
Hajek M, Sewell A, Kaech S, Burtness B,
Yarbrough WG and Issaeva N: TRAF3/CYLD mutations identify a
distinct subset of human papillomavirus-associated head and neck
squamous cell carcinoma. Cancer. 123:1778–1790. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Henderson S, Chakravarthy A, Su X, Boshoff
C and Fenton TR: APOBEC-mediated cytosine deamination links PIK3CA
helical domain mutations to human papillomavirus-driven tumor
development. Cell Reports. 7:1833–1841. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Lui VW, Hedberg ML, Li H, Vangara BS,
Pendleton K, Zeng Y, Lu Y, Zhang Q, Du Y, Gilbert BR, et al:
Frequent mutation of the PI3K pathway in head and neck cancer
defines predictive biomarkers. Cancer Discov. 3:761–769. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Kostareli E, Holzinger D, Bogatyrova O,
Hielscher T, Wichmann G, Keck M, Lahrmann B, Grabe N,
Flechtenmacher C, Schmidt CR, et al: HPV-related methylation
signature predicts survival in oropharyngeal squamous cell
carcinomas. J Clin Invest. 123:2488–2501. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Lechner M, Fenton T, West J, Wilson G,
Feber A, Henderson S, Thirlwell C, Dibra HK, Jay A, Butcher L, et
al: Identification and functional validation of HPV-mediated
hypermethylation in head and neck squamous cell carcinoma. Genome
Med. 5:152013. View
Article : Google Scholar : PubMed/NCBI
|
|
109
|
de Lena PG, Paz-Gallardo A, Paramio JM and
García-Escudero R: Clusterization in head and neck squamous
carcinomas based on lncRNA expression: Molecular and clinical
correlates. Clin Epigenetics. 9:362017. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Jung AC, Briolat J, Millon R, de Reyniès
A, Rickman D, Thomas E, Abecassis J, Clavel C and Wasylyk B:
Biological and clinical relevance of transcriptionally active human
papillomavirus (HPV) infection in oropharynx squamous cell
carcinoma. Int J Cancer. 126:1882–1894. 2010.
|
|
111
|
Kimple RJ, Smith MA, Blitzer GC, Torres
AD, Martin JA, Yang RZ, Peet CR, Lorenz LD, Nickel KP, Klingelhutz
AJ, et al: Enhanced radiation sensitivity in HPV-positive head and
neck cancer. Cancer Res. 73:4791–4800. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Seiwert TY, Zuo Z, Keck MK, Khattri A,
Pedamallu CS, Stricker T, Brown C, Pugh TJ, Stojanov P, Cho J, et
al: Integrative and comparative genomic analysis of HPV-positive
and HPV-negative head and neck squamous cell carcinomas. Clin
Cancer Res. 21:632–641. 2015. View Article : Google Scholar :
|
|
113
|
Gubanova E, Brown B, Ivanov SV, Helleday
T, Mills GB, Yarbrough WG and Issaeva N: Downregulation of SMG-1 in
HPV-positive head and neck squamous cell carcinoma due to promoter
hypermethylation correlates with improved survival. Clin Cancer
Res. 18:1257–1267. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Lawrence MS, Sougnez C, Lichtenstein L,
Cibulskis K, Lander E, Gabriel SB, Getz G, Ally A, Balasundaram M,
Birol I, et al Cancer Genome Atlas Network: Comprehensive genomic
characterization of head and neck squamous cell carcinomas. Nature.
517:576–582. 2015. View Article : Google Scholar
|
|
115
|
Pyeon D, Newton MA, Lambert PF, den Boon
JA, Sengupta S, Marsit CJ, Woodworth CD, Connor JP, Haugen TH,
Smith EM, et al: Fundamental differences in cell cycle deregulation
in human papillomavirus-positive and human papillomavirus-negative
head/neck and cervical cancers. Cancer Res. 67:4605–4619. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Lajer CB, Garnæs E, Friis-Hansen L,
Norrild B, Therkildsen MH, Glud M, Rossing M, Lajer H, Svane D,
Skotte L, et al: The role of miRNAs in human papilloma virus
(HPV)-associated cancers: Bridging between HPV-related head and
neck cancer and cervical cancer. Br J Cancer. 106:1526–1534. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Slebos RJC, Jehmlich N, Brown B, Yin Z,
Chung CH, Yarbrough WG and Liebler DC: Proteomic analysis of
oropharyngeal carcinomas reveals novel HPV-associated biological
pathways. Int J Cancer. 132:568–579. 2013. View Article : Google Scholar
|
|
118
|
Colacino JA, Dolinoy DC, Duffy SA, Sartor
MA, Chepeha DB, Bradford CR, McHugh JB, Patel DA, Virani S, Walline
HM, et al: Comprehensive analysis of DNA methylation in head and
neck squamous cell carcinoma indicates differences by survival and
clinicopathologic characteristics. PLoS One. 8:e547422013.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Klussmann JP, Mooren JJ, Lehnen M,
Claessen SM, Stenner M, Huebbers CU, Weissenborn SJ, Wedemeyer I,
Preuss SF, Straetmans JM, et al: Genetic signatures of HPV-related
and unrelated oropharyngeal carcinoma and their prognostic
implications. Clin Cancer Res. 15:1779–1786. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Wilting SM, Smeets SJ, Snijders PJF, van
Wieringen WN, van de Wiel MA, Meijer GA, Ylstra B, Leemans CR,
Meijer CJLM, Brakenhoff RH, et al: Genomic profiling identifies
common HPV-associated chromosomal alterations in squamous cell
carcinomas of cervix and head and neck. BMC Med Genomics. 2:322009.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Canham M, Charsou C, Stewart J, Moncur S,
Hoodless L, Bhatia R, Cong D, Cubie H, Busby-Earle C, Williams A,
et al: Increased cycling cell numbers and stem cell associated
proteins as potential biomarkers for high grade human
papillomavirus+ve pre-neoplastic cervical disease. PLoS One.
9:e1153792014. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Van Raemdonck GAA, Tjalma WAA, Coen EP,
Depuydt CE and Van Ostade XWM: Identification of protein biomarkers
for cervical cancer using human cervicovaginal fluid. PLoS One.
9:e1064882014. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Descamps G, Wattiez R and Saussez S:
Proteomic study of HPV-positive head and neck cancers: Preliminary
results. BioMed Res Int. 2014:4309062014. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Bonner JA, Harari PM, Giralt J, Azarnia N,
Shin DM, Cohen RB, Jones CU, Sur R, Raben D, Jassem J, et al:
Radiotherapy plus cetuximab for squamous-cell carcinoma of the head
and neck. N Engl J Med. 354:567–578. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Leung DT, Lim PL, Cheung TH, Wong RR, Yim
SF, Ng MH, Tam FC, Chung TK and Wong YF: Osteopontin fragments with
intact thrombin-sensitive site circulate in cervical cancer
patients. PLoS One. 11:e01604122016. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
De Marco F, Bucaj E, Foppoli C, Fiorini A,
Blarzino C, Filipi K, Giorgi A, Schininà ME, Di Domenico F, Coccia
R, et al: Oxidative stress in HPV-driven viral carcinogenesis:
Redox proteomics analysis of HPV-16 dysplastic and neoplastic
tissues. PLoS One. 7:e343662012. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Perluigi M, Giorgi A, Blarzino C, De Marco
F, Foppoli C, Di Domenico F, Butterfield DA, Schininà ME, Cini C
and Coccia R: Proteomics analysis of protein expression and
specific protein oxidation in human papillomavirus transformed
keratinocytes upon UVB irradiation. J Cell Mol Med. 13(8B):
1809–1822. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Overgaard J, Eriksen JG, Nordsmark M and
Alsner J; Danish Head and Neck Cancer Study Group:: Plasma
osteopontin, hypoxia, and response to the hypoxia sensitiser
nimorazole in radiotherapy of head and neck cancer: Results from
the DAHANCA 5 randomised double-blind placebo-controlled trial.
Lancet Oncol. 6:757–764. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Hassan Metwally MA, Jansen JA and
Overgaard J: Study of the population pharmacokinetic
characteristics of nimorazole in head and neck cancer patients
treated in the DAHANCA-5 trial. Clin Oncol (R Coll Radiol).
27:168–175. 2015. View Article : Google Scholar
|
|
130
|
Lee KA, Kang JW, Shim JH, Kho CW, Park SG,
Lee HG, Paik SG, Lim JS and Yoon DY: Protein profiling and
identification of modulators regulated by human papillomavirus 16
E7 oncogene in HaCaT keratinocytes by proteomics. Gynecol Oncol.
99:142–152. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Thomas M, Tomaić V, Pim D, Myers MP,
Tommasino M and Banks L: Interactions between E6AP and E6 proteins
from alpha and beta HPV types. Virology. 435:357–362. 2013.
View Article : Google Scholar
|
|
132
|
Sandberg A, Lindell G, Källström BN,
Branca RM, Danielsson KG, Dahlberg M, Larson B, Forshed J and
Lehtiö J: Tumor proteomics by multivariate analysis on individual
pathway data for characterization of vulvar cancer phenotypes. Mol
Cell Proteomics. 11:M112.0169982012. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Jamison JH, Kaplan DW, Hamman R, Eagar R,
Beach R and Douglas JM Jr: Spectrum of genital human papillomavirus
infection in a female adolescent population. Sex Transm Dis.
22:236–243. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Blas MM, Brown B, Menacho L, Alva IE,
Silva-Santisteban A and Carcamo C: HPV Prevalence in multiple
anatomical sites among men who have sex with men in Peru. PLoS One.
10:e01395242015. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Uzawa K, Baba T, Uchida F, Yamatoji M,
Kasamatsu A, Sakamoto Y, Ogawara K, Shiiba M, Bukawa H and Tanzawa
H: Circulating tumor-derived mutant mitochondrial DNA: A predictive
biomarker of clinical prognosis in human squamous cell carcinoma.
Oncotarget. 3:670–677. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
No. 90-IARC Working Group on the
evaluation of carcinogenic risk to humans: IARC Monographs on the
evaluation of carcinogenic risks to humans. IARC; Lyon: 2007
|
|
137
|
Elrefaey S, Massaro MA, Chiocca S, Chiesa
F and Ansarin M: HPV in oropharyngeal cancer: The basics to know in
clinical practice. Acta Otorhinolaryngol Ital. 34:299–309.
2014.
|
|
138
|
Faridi R, Zahra A, Khan K and Idrees M:
Oncogenic potential of human papillomavirus (HPV) and its relation
with cervical cancer. Virol J. 8:2692011. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Hamana K, Uzawa K, Ogawara K, Shiiba M,
Bukawa H, Yokoe H and Tanzawa H: Monitoring of circulating
tumour-associated DNA as a prognostic tool for oral squamous cell
carcinoma. Br J Cancer. 92:2181–2184. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Voiculescu V, Calenic B, Ghita M, Lupu M,
Caruntu A, Moraru L, Voiculescu S, Ion A, Greabu M, Ishkitiev N and
Caruntu C: From normal skin to squamous cell carcinoma: A quest for
novel biomarkers. Dis Markers. 2016:45174922016. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Ang KK, Harris J, Wheeler R, Weber R,
Rosenthal DI, Nguyen-Tân PF, Westra WH, Chung CH, Jordan RC, Lu C,
et al: Human papillomavirus and survival of patients with
oropharyngeal cancer. N Engl J Med. 363:24–35. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Stanescu B, Miron R and Badiu C:
Anaplastic carcinoma of the thyroid mimicking acute thyroiditis.
Acta Endocrinol (Bucur). 3:493–502. 2007. View Article : Google Scholar
|
|
143
|
Forastiere AA, Goepfert H, Maor M, Pajak
TF, Weber R, Morrison W, Glisson B, Trotti A, Ridge JA, Chao C, et
al: Concurrent chemotherapy and radiotherapy for organ preservation
in advanced laryngeal cancer. N Engl J Med. 349:2091–2098. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Pignon JP, le Maître A, Maillard E and
Bourhis J; MACH-NC Collaborative Group: Meta-analysis of
chemotherapy in head and neck cancer (MACH-NC): An update on 93
randomised trials and 17,346 patients. Radiother Oncol. 92:4–14.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Călina D, Docea AO, Roşu L, Zlatian O,
Roşu AF, Anghelina F, Rogoveanu O, Arsene AL, Nicolae AC, Drăgoi
CM, et al: Antimicrobial resistance development following surgical
site infections. Mol Med Rep. 15:681–688. 2017. View Article : Google Scholar :
|
|
146
|
Tănase A, Coliță A, Ianoşi G, Neagoe D,
Brănişteanu DE, Călina D, Docea AO, Tsatsakis A and Ianoşi SL: Rare
case of disseminated fusariosis in a young patient with graft vs.
host disease following an allogeneic transplant. Exp Ther Med.
12:2078–2082. 2016. View Article : Google Scholar :
|
|
147
|
Mioc M, Soica C, Bercean V, Avram S,
Balan-Porcarasu M, Coricovac D, Ghiulai R, Muntean D, Andrica F,
Dehelean C, et al: Design, synthesis and pharmaco-toxicological
assessment of 5-mercapto-1,2,4-triazole derivatives with
antibacterial and antiproliferative activity. Int J Oncol.
50:1175–1183. 2017. View Article : Google Scholar :
|
|
148
|
Oprean C, Borcan F, Pavel I, Dema A,
Danciu C, Soica C, Dehelean C, Nicu A, Ardelean A, Cristea M, et
al: In vivo biological evaluation of polyurethane nanostructures
with ursolic and oleanolic acids on chemically-induced skin
carcinogenesis. In Vivo. 30:633–638. 2016.PubMed/NCBI
|
|
149
|
Sani TA, Mohammadpour E, Mohammadi A,
Memariani T, Yazdi MV, Rezaee R, Calina D, Docea AO, Goumenou M,
Etemad L and Shahsavand S: Cytotoxic and apoptogenic properties of
Dracocephalum kotschyi aerial part different fractions on calu-6
and mehr-80 lung cancer cell lines. Farmacia. 65:189–199. 2017.
|
|
150
|
Suzich JA, Ghim SJ, Palmer-Hill FJ, White
WI, Tamura JK, Bell JA, Newsome JA, Jenson AB and Schlegel R:
Systemic immunization with papillomavirus L1 protein completely
prevents the development of viral mucosal papillomas. Proc Natl
Acad Sci USA. 92:11553–11557. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Roden RB, Monie A and Wu TC: Opportunities
to improve the prevention and treatment of cervical cancer. Curr
Mol Med. 7:490–503. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Joura EA, Leodolter S, Hernandez-Avila M,
Wheeler CM, Perez G, Koutsky LA, Garland SM, Harper DM, Tang GW,
Ferris DG, et al: Efficacy of a quadrivalent prophylactic human
papillomavirus (types 6, 11, 16, and 18) L1 virus-like-particle
vaccine against high-grade vulval and vaginal lesions: A combined
analysis of three randomised clinical trials. Lancet.
369:1693–1702. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Varsani A, Williamson AL, de Villiers D,
Becker I, Christensen ND and Rybicki EP: Chimeric human
papillomavirus type 16 (HPV-16) L1 particles presenting the common
neutralizing epitope for the L2 minor capsid protein of HPV-6 and
HPV-16. J Virol. 77:8386–8393. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Paavonen J, Jenkins D, Bosch FX, Naud P,
Salmerón J, Wheeler CM, Chow SN, Apter DL, Kitchener HC,
Castellsague X, et al HPV PATRICIA study group: Efficacy of a
prophylactic adjuvanted bivalent L1 virus-like-particle vaccine
against infection with human papillomavirus types 16 and 18 in
young women: An interim analysis of a phase III double-blind,
randomised controlled trial. Lancet. 369:2161–2170. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Panatto D, Amicizia D, Bragazzi NL,
Rizzitelli E, Tramalloni D, Valle I and Gasparini R: Human
papillomavirus vaccine: State of the art and future perspectives.
Adv Protein Chem Struct Biol. 101:231–322. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Toft L, Storgaard M, Müller M, Sehr P,
Bonde J, Tolstrup M, Østergaard L and Søgaard OS: Comparison of the
immunogenicity and reactogenicity of Cervarix and Gardasil human
papillomavirus vaccines in HIV-infected adults: A randomized,
double-blind clinical trial. J Infect Dis. 209:1165–1173. 2014.
View Article : Google Scholar
|