Introduction
Colon cancer is the second leading cause of
cancer-associated mortality in the United States (1). The current standard treatment for
patients with colon cancer is surgical resection followed by
chemotherapy (2). However,
patients that experience recurrence following surgery or are
diagnosed when they are at an advanced stage colon cancer are
difficult to treat, despite the development of novel
chemotherapeutic regimens and molecular targeted therapy (3). In addition, continuous chemotherapy
and molecular targeted therapy induce toxicity in normal tissues.
Therefore, novel treatments are required that do not induce these
problems in patients.
Apoptosis is a form of regulated cell death that
results in the removal of damaged or potentially harmful cells
(4). The Bcl-2 family of proteins
consists of pro- (Bax and Bak) and anti-apoptotic proteins (Bcl-xL
and Mcl-1) and they regulate apoptosis (5). Apoptosis is stimulated by the
downregulation of anti-apoptotic proteins and/or the upregulation
of pro-apoptotic proteins. Changes in the balance of these proteins
promote the permeability of the outer membrane of mitochondria,
resulting in the release of cytochrome c and the induction of
apoptosis via the caspase cascade (6). Therefore, targeting the Bcl-2 family
proteins, particularly the anti-apoptotic proteins, may be a novel
method of treating cancer (7–10).
Apigenin is a common flavonoid and is present in
many plants, fruits and vegetables, including oranges, onions,
parsley, tea, chamomile, wheat sprouts and certain seasonings
(11). One of the most common
sources of consumed apigenin is chamomile tea, which is prepared
from the dried flowers of Matricaria chamomilla (12). Apigenin possesses remarkable
anti-inflammatory, -oxidant and -carcinogenic properties, and
inhibits the proliferation, arrests the cell cycle and induces
apoptosis in various types of cancer, including breast, lung, liver
and ovarian cancer, without affecting healthy cells. It has been
demonstrated that apigenin reduces the expression of Mcl-1 in colon
cancer cells (13). However, to
the best of our knowledge, the mechanism by which apigenin affects
the Bcl-2 family of proteins in colon cancer remains unknown.
The signal transducer and activator of transcription
(STAT) family consists of cytoplasmic transcription factors that
form dimers upon phosphorylation, interact with other
transcriptional modulators that migrate to the nucleus and bind to
specific promoter sequences to induce gene expression (14). Several studies have demonstrated
that STAT family proteins, including STAT3 and STAT5, recognize
constitutive activity in different types of human cancer cells and
stimulate the progression and development of human cancer (15,16).
Inhibition of STAT3 or STAT5 activation is associated with the
suppression of cancer growth and the induction of cell death
(17,18). Previous studies have demonstrated
that apigenin exhibits an antitumor effect in different types of
cancer by inhibiting the STAT3 signaling pathway (19,20).
However, the effect of apigenin on the interaction between STAT
signaling and the expression of Bcl-2 family proteins in colon
cancer cells has not yet been investigated.
The present study demonstrated that apigenin induces
the apoptosis of colon cancer cells by downregulating Bcl-xL and
Mcl-1 expression via inhibition of the STAT signaling pathway.
These results demonstrate that apigenin may be used as a novel
therapeutic method of treating patients with colon cancer.
Materials and methods
Reagents
Antibodies against Bcl-xL (1:1,000, cat. no. 54H6),
Mcl-1 (1:1,000, cat. no. D5V5L), STAT3 (1:1,000, cat. no. D3Z2G),
phosphorylated (p)-STAT3 (1:2,000, cat. no. Tyr705), STAT5
(1:1,000, cat. no. D206Y), p-STAT5 (1:1,000, cat. no. Tyr694),
poly-(ADP-ribose) polymerase (PARP; 1:1,000, cat. no. 46D11) and
GAPDH (1:1,000, cat. no. 14C10) were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). The secondary antibodies
polyclonal goat anti-rabbit immunoglobulins (Igs) conjugated to
horseradish peroxidase (HRP; 1:2,000, cat. no. P0448) and
polyclonal goat anti-mouse Igs/HRP (1:2,000, cat. no. P0447) were
purchased from Dako; Agilent Technologies, Inc. (Santa Clara, CA,
USA). Apigenin (4′,5,7-trihydroxyflavone) was purchased from
R&D Systems, Inc. (Minneapolis, MN, USA). Human recombinant
interleukin-6 (IL-6) and human recombinant soluble IL-6 receptor
were purchased from PeproTech, Inc. (Rocky Hill, NJ, USA). IL-6 was
used supplemented with soluble IL-6 receptor (40 ng/ml).
Cell culture
The human colon cancer cell lines HT29, DLD-1,
COLO320 and HCT116 were obtained from the American Type Culture
Collection (Manassas, VA, USA). HT29 (KRAS wild-type) cells were
cultured in Dulbecco’s modified Eagle’s medium (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) supplemented with 10% fetal bovine
serum (FBS; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
1% antibiotic-antimycotic (Sigma-Aldrich; Merck KGaA). DLD-1 (KRAS
mutant-type) and COLO320 (KRAS wild-type) cells were cultured in
RPMI-1640 medium (Sigma-Aldrich; Merck KGaA) supplemented with 10%
FBS and 1% antibiotic-antimycotic. HCT116 (KRAS mutant-type) cells
were cultured in McCoy’s 5A medium (Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS and 1% antibiotic-antimycotic. All cell
lines were maintained at 37°C in a humidified incubator containing
5% CO2.
Patients and tumor samples
Colon cancer tissue samples were obtained from 40
patients (25 males and 15 females) with Stage Ⅳ colon cancer who
had undergo surgery at the Nagoya City University Hospital (Nagoya,
Japan) between January 2012 and December 2014. Resected specimens
were staged by pathological evaluation according to the Japanese
Classification of Colorectal Carcinoma (21). The mean age of the patients was
66.2±10 years (range, 37–88 years). All patients gave their written
informed consent for the use of their tissue samples in the present
study.
Immunohistochemistry
Resected specimens were fixed with formalin at room
temperature for 2 days. Formalin-fixed, paraffin-embedded sections
4-µm-thick were deparaffinized in xylene and hydrated in
graded alcohols. Following washing with running water, samples were
soaked in soak in 10 mM citric acid buffer (9 ml 0.1 M citric acid
+ 41 ml 0.1 mM triso-dium citrate dihydrate + 450 ml distilled
water) and boiled for 10 min. Following 3 washes with PBS (5
min/time) the slide was immersed in a mixture of 1.5 ml 30%
hydrogen peroxide solution and 150 ml 100% methanol for 30 min to
block endogenous peroxidase. Following washing with PBS, slides
were blocked with 400 µl 4% Block Ace® Powder
(cat. no. UK-B80; DS Pharma Biomedical Co., Ltd., Osaka, Japan) for
10 min in a humidity box at room temperature. Subsequently, slides
were incubated with primary antibodies against Bcl-xL (1:300) and
Mcl-1 (1:100) onto at 4°C overnight. Following washing with PBS,
slides were incubated with 100 µl secondary antibody
(EnVision + Single Reagents HRP; cat. no. K4003; Dako; Agilent
Technologies, Inc.) for 45 min at room temperature. The detection
step was performed using Liquid DAB + Substrate Chromogen System
(cat. no. K3467; Dako; Agilent Technologies, Inc.) for 10 min at
room temperature and the sections were counterstained with
hematoxylin for 30 sec at room tenmperature. Slides were analyzed
using a light microscope (magnification, ×400). Ethical approval
for the use of human tissue was granted by the Graduate School of
Medicine, Nagoya City University (Nagoya, Japan) and all patients
provided their informed consent for the use of their tissues in the
current study.
Preparation of apigenin
Apigenin was dissolved in dimethyl sulfoxide (DMSO;
Wako Chemicals GmbH, Neuss, Germany) to produce 50 mM stock
solutions that were aliquoted and stored at −28°C prior to use.
Stock solutions were diluted with culture medium to the indicated
concentrations (final DMSO concentration, 0.1%).
Transfection of small interfering
(si)RNA
Bcl-xL siRNA (5 nM, ID#:s1922; forward,
5′-GGAACUCUAUGGGAACAAUTT-3′ and reverse,
5′-AUUGUUCCCAUAGAGUUCCAC-3′), Mcl-1 siRNA (5 nM, ID#:s8583;
forward, 5′-CCAGUAUACUUCUUAGAAATT-3′ and reverse,
5′-UUUCUAAGAAGUAUACUGGGA-3′), negative control siRNA (5 nM, cat.
no. 4390843, Silencer Select Negative Control #1) and STAT3 siRNA
(5 nM, ID#:s744, sequence: forward, 5′-GGCUGGACAAUAUCAUUGATT-3′ and
reverse, 5′-UCAAUGAUAUUGUCCAGCCAG-3′) were purchased from Thermo
Fisher Scientific, Inc. On the day before siRNA transfection, DLD-1
and HCT116 cells (1.0×104 cells) were seeded to 60–80%
confluence in their respective growth mediums without antibiotics.
siRNAs and Lipofectamine RNAiMAX were diluted in Opti-MEM medium
(both from Thermo Fisher Scientific, Inc.), mixed gently and
incubated for 10–20 min at room temperature. Cells were transfected
by adding the siRNA-Lipofectamine RNAiMAX complex dropwise to the
medium to achieve a siRNA final concentration of 100 nM. Cells were
incubated at 37°C in a CO2 incubator and knockdown
efficiency was evaluated 24 h post-transfection.
DNA fragmentation assay
Cells (5×105/well) were seeded into
6-well culture plates and incubated at 37°C overnight. Medium was
replenished with serum-free medium and incubated for a further 4 h.
Cells were then treated with apigenin at various doses 0, 5, 15 and
50 µM or transfected to target Bcl-xL, Mcl-1 or Bcl-xL and
Mcl-1 together. DNA fragmentation was analyzed using a Cell Death
Detection ELISAPlus kit (Sigma-Aldrich; Merck KGaA)
following the manufacturer’s protocol. The degree of apoptosis was
evaluated using an enrichment factor (mU of treated cells/mU of
non-treated cells).
Cell proliferation assay
The effect of apigenin or siRNA on cell
proliferation was measured using WST-1 and BrdU assays. Each assay
was performed at least three times. The WST-1 assay was performed
using a Premix WST-1 cell proliferation assay system (Takara Bio,
Inc., Tokyo, Japan). Cells (5×103–5×104/well)
were seeded into 96-well tissue culture plates and incubated at
37°C overnight. Cells were treated with 0, 5, 15 and 50 µM
apigenin or transfected to target Bcl-xL, Mcl-1 or Bcl-xL and Mcl-1
together in serum-free medium. After 48 h, the medium was removed
and replaced with fresh medium (90 µl/well) containing
Premix WST-1 (10 µl/well). Cells were then incubated at 37°C
for 2 h. Absorbance was measured using a plate reader at 450 nm.
The BrdU assay was performed using BrdU Cell Proliferation ELISA
kit (colorimetric; cat. no. ab126556; Abcam, Cambridge, MA, USA),
following the manufacturer’s protocol. Cells
(5×103–5×104/well) were seeded into 96-well
tissue culture plates and incubated at 37°C overnight. Cells were
treated with 0, 5, 15 and 50 µM apigenin or transfected to
target Bcl-xL, Mcl-1, or Bcl-xL and Mcl-1 together in serum-free
medium for 32 h at 37°C. A total of 20 µl 1/500 diluted BrdU
was added and incubated for 16 h at room temperature. Subsequently,
fixing solution was added for 30 min at room temperature to fix and
permeabilize cells, and denature the DNA. Subsequently, anti-BrdU
antibody was added to each well for 1 h and peroxidase-conjugated
goat anti-mouse immunoglobulin G antibody was added for 30 min.
Incubation with primary and secondary antibodies was performed at
room temperature. Finally, 3,3′,5,5′-tetra-methylbenzidine
peroxidase substrate was added to each well and following
incubation in the dark for 30 min, stop solution was added to each
well. Absorbance was measured at 450 nm using a microplate
reader.
Identifying the mechanism of action of
apigenin
Western blotting was performed to identify the
mechanism of action of apigenin. Cells (DLD-1, HCT116:
1×106/dish) were seeded in 4 dishes and incubated at
37°C overnight. The following day, cells were treated with 0, 5, 15
and 50 µM IL-6 in serum-free medium for 48 h. Proteins were
the extracted and expression of p-STAT3 was evaluated by western
blotting.
Cells (DLD-1, HCT116: 1×106/dish) were
seeded in 4 dishes and incubated at 37°C overnight. The following
day, cells were treated with or without 50 µM (DLD-1), 15
µM (HCT 116) apigenin for 2 h and then with or without 50
µM IL-6 for a further 48 h. Proteins were extracted and
expression of p-STAT3, Bcl-xL and Mcl-1 was evaluated by western
blotting.
Western blot analysis
Protein samples were prepared in
radioimmunoprecipitation lysis buffer with Protease Inhibitor
Single Use Cocktail and Phosphatase Inhibitor Cocktail (all from
Thermo Fisher Scientific, Inc.). Protein concentrations were
measured using a BCA protein assay kit (Thermo Fisher Scientific,
Inc.). Equal amounts of protein extract were denatured by boiling
at 90°C for 5 min. Proteins (20 µg) were fractionated on
4–15% Mini-PROTEAN TGX gels and transferred to nitrocellulose
membranes (both from Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The primary and secondary antibody reactions were performed
using an iBind Flex Western System (Thermo Fisher Scientific,
Inc.). Membranes were incubated with iBind Flex Solution (iBind
Flex Buffer, iBind Flex Additive and distilled water) for 10 min at
room temperature to block nonspecific binding. Primary (Bcl-xL,
Mcl-1, STAT3, p-STAT3, STAT5, p-STAT5, PARP and GAPDH) and
secondary (polyclonal goat anti-rabbit IGs conjugated to HRP)
antibody reactions were performed at room temperature for 2.5 h,
following the manufacturer’s protocol. Protein-antibody complexes
were visualized using SuperSignal West Pico Chemiluminescent
Substrate or SuperSignal West Femto Chemiluminescent Substrate
(Thermo Fisher Scientific, Inc.). The immunoreactive protein band
was detected and band density was quantified by densitometry using
an Amersham Imager 600 (GE Healthcare Life Sciences, Little
Chalfont, UK).
Statistical analysis
All statistical analyses were performed using EZR
software (Easy R) version 1.27 (Saitama Medical Center, Jichi
Medical University, Saitama, Japan). All data are presented as the
mean ± standard deviation. Statistical comparisons with paired
observations were determined using the Student’s t-test.
Comparisons of more than two groups were made by a one-way analysis
of variance followed by Dunnett’s test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Apigenin reduces the proliferation of
colon cancer cells
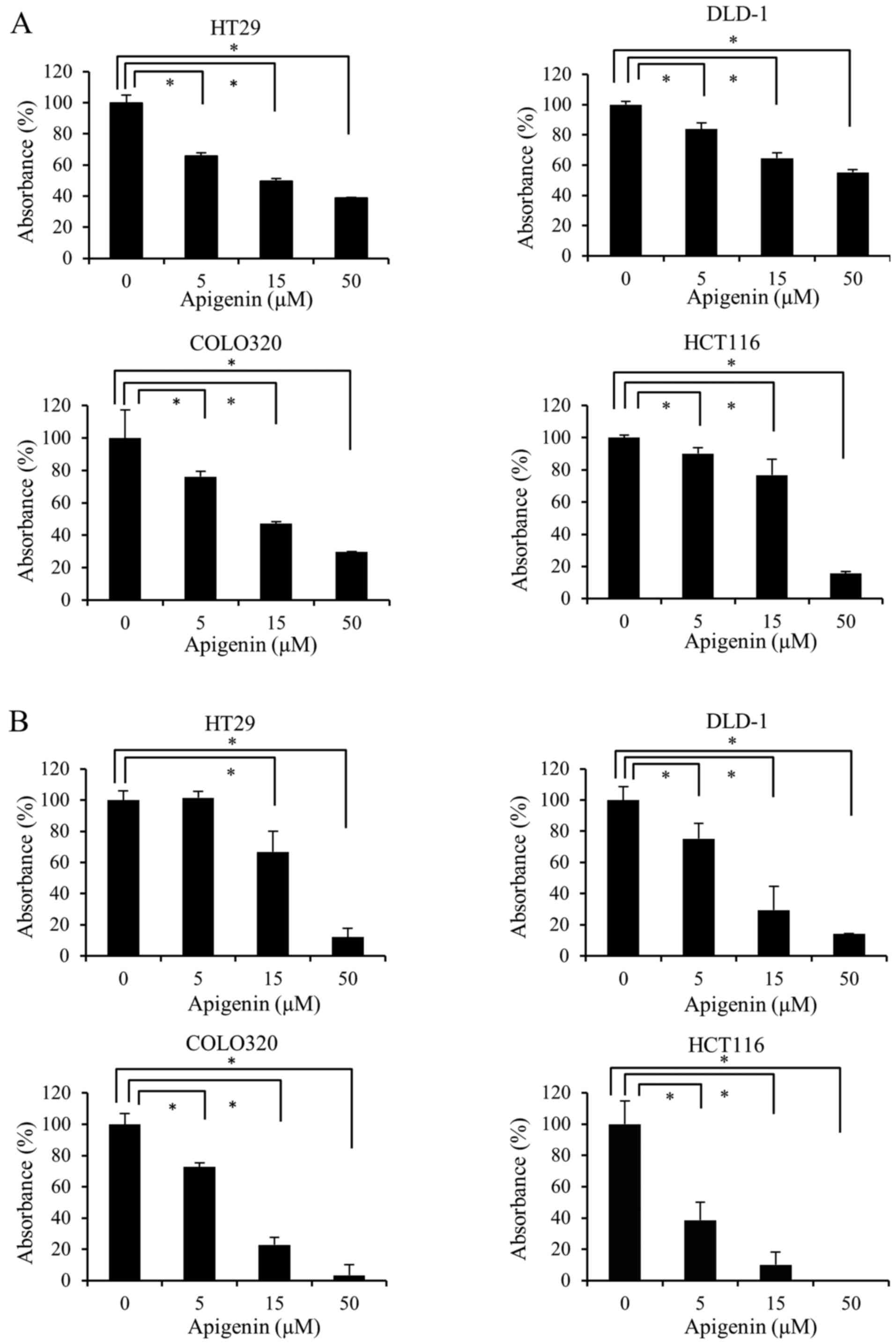
To investigate whether apigenin inhibits the
proliferation of colon cancer cells, four colon cancer cell lines
(HT29, DLD-1, COLO320 and HCT116) were treated with 0, 5, 15 and 50
µM apigenin for 48 h. Cell proliferation was measured using
WST-1 (Fig. 1A) and BrdU (Fig. 1B) assays. The results demonstrated
that apigenin significantly reduced the proliferation of all four
colon cancer cell lines in a dose-dependent manner.
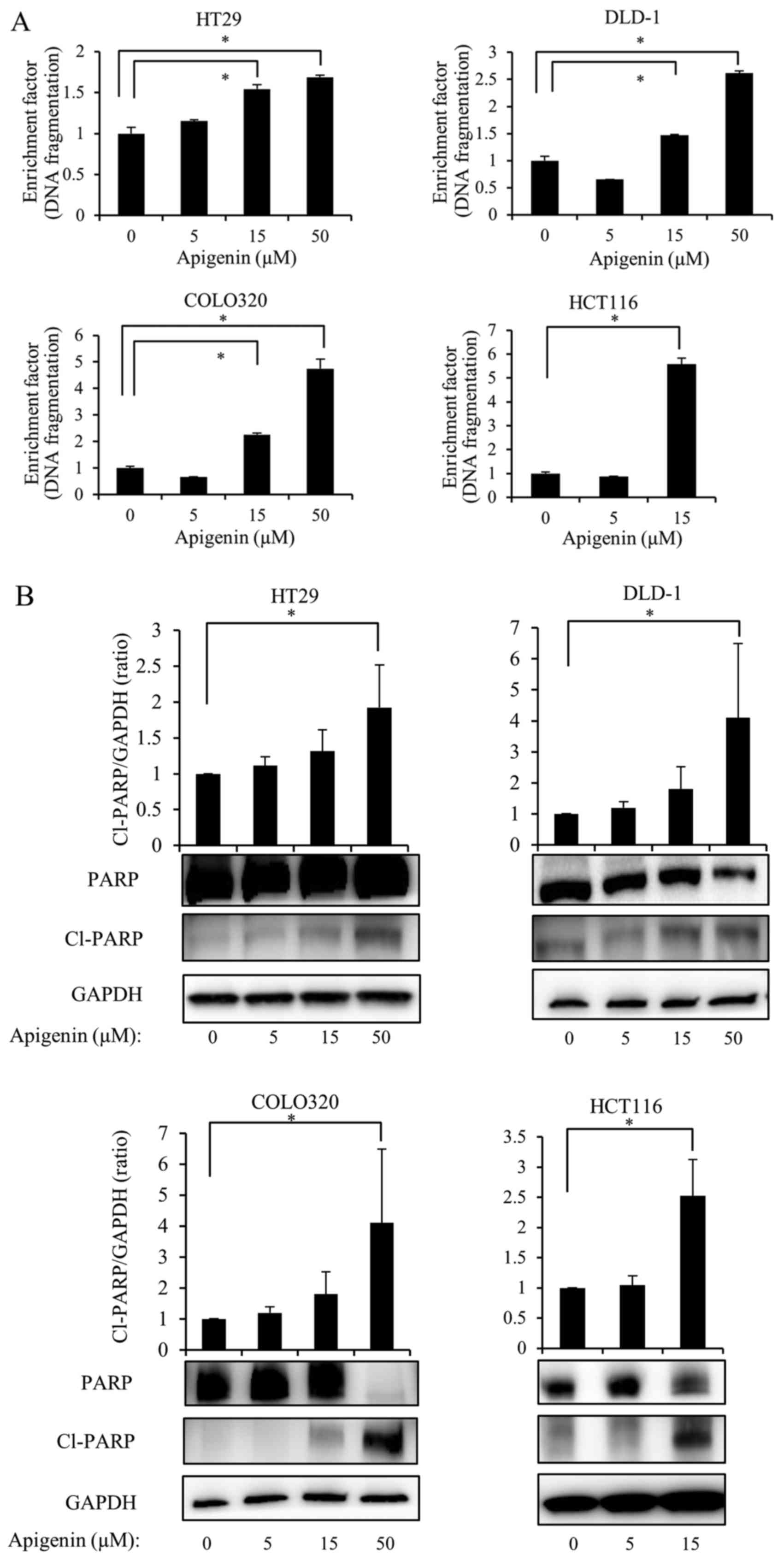
Apigenin induces apoptosis in colon
cancer cells via a caspase-dependent pathway
To investigate whether the inhibition of cell
proliferation by apigenin was due to the induction of apoptosis,
four colon cancer cell lines were exposed to various doses of
apigenin for 24 h and the degree of DNA fragmentation in the cells
was then measured. The results demonstrated that apigenin induced
DNA fragmentation in a dose-dependent manner in all four colon
cancer cell lines (Fig. 2A).
Subsequently, it was determined whether the effect
of apigenin on apoptosis was mediated by caspase activation. The
four colon cancer cell lines were treated with various doses of
apigenin for 24 h and the cleavage of PARP was evaluated by western
blotting. The results demonstrated that apigenin induced the
cleavage of PARP in a dose-dependent manner (Fig. 2B). In HCT116 cells, the
apoptosis-inducing effect of 50 µM apigenin was weaker than
that of 15 µM apigenin. However, as presented in Fig. 1, the suppression of cell
proliferation in HCT116 occurred in a dose-dependent manner, with
50 µM apigenin inducing the most marked inhibition. Thus, it
was considered that this phenomenon may involve other processes, as
well as apoptosis, such as necrosis. Therefore, only 5 and 15
µM apigenin were used to treat HCT116 cells in subsequent
experiments investigating the effects of apigenin on the apoptosis
of colon cancer cells.
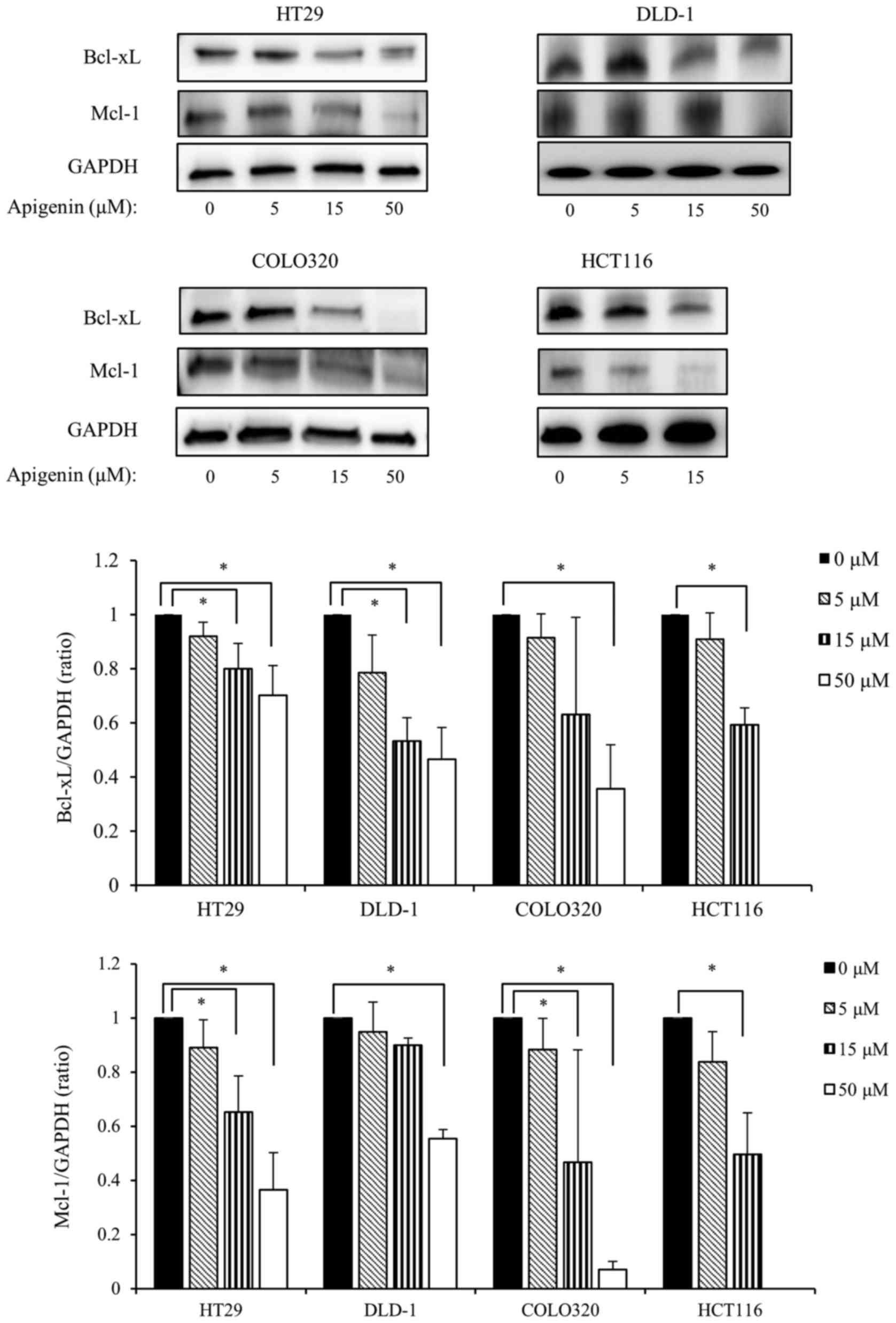
Apigenin downregulates the expression of
the anti-apoptotic Bcl-2 family members Bcl-xL and Mcl-1
The anti-apoptotic Bcl-2 family of proteins serves
an important role in protecting against DNA damage-induced
apoptosis. Therefore, the present study investigated the effect of
apigenin on the expression of anti-apoptotic Bcl-2 family proteins.
The four colon cancer cell lines were exposed to various doses of
apigenin for 24 h and the expression of Bcl-xL and Mcl-1 were
measured by western blotting. The results demonstrated that
apigenin significantly downregulated the expression of Bcl-xL and
Mcl-1 in all colon cancer cells lines in a dose-dependent manner
(Fig. 3).
The simultaneous downregulation of Bcl-xL
and Mcl-1 strongly induces apoptosis via a caspase-dependent
pathway in colon cancer cells
Apigenin downregulated the expression of Bcl-xL and
Mcl-1; therefore, it was hypothesized that the anti-apoptotic Bcl-2
family proteins Bcl-xL and Mcl-1 are involved in apigenin-induced
apoptosis. To determine whether this was the case, the DLD-1 and
HCT116 cell lines were used. As a preliminary experiment,
immunostaining of clinical specimens of colon cancer was performed.
The results demonstrated that expression of Bcl-xL was very strong
in colon cancer and that the expression of Mcl-1 was relatively
weak (data not presented). Since these data were in accordance with
the results of western blotting regarding Bcl-xL and Mcl-1
expression in the DLD-1 and HCT116 cell lines, the following
experiments were performed on these two cell lines.
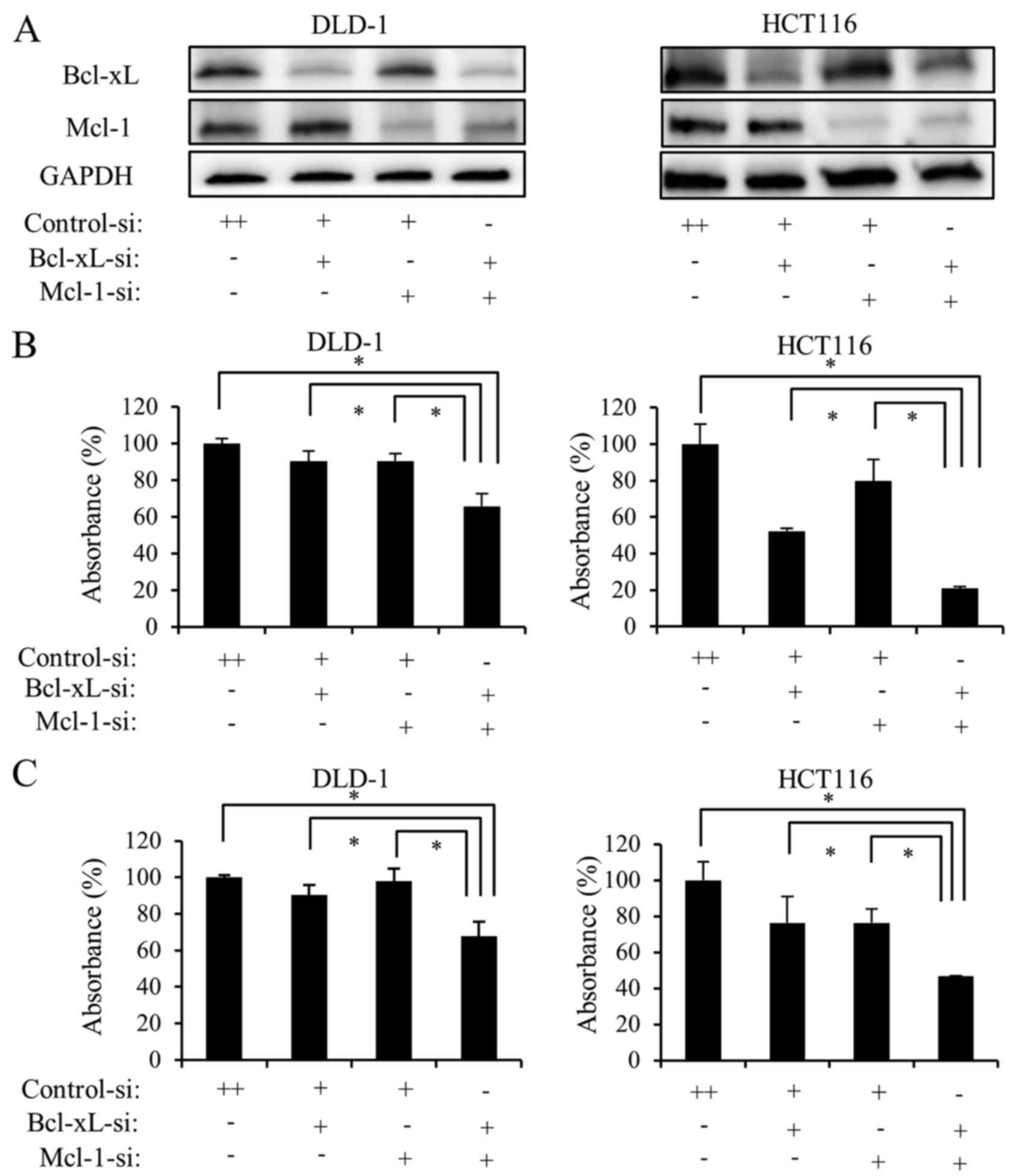
Cells were transfected with siRNA targeting Bcl-xL,
Mcl-1, or Bcl-xL and Mcl-1 together. Transfection with the siRNAs
downregulated Bcl-xL and MCl-1 expression in DLD-1 and HCT116 cells
(Fig. 4A). To investigate whether
transfection with siRNA inhibits colon cancer cell proliferation,
DLD-1 and HCT116 cells were transfected with siRNA targeting
Bcl-xL, Mcl-1, or Bcl-xL and Mcl-1 together for 48 h. Cell
proliferation was measured using WST-1 (Fig. 4B) and BrdU assays (Fig. 4C). Downregulation of Bcl-xL alone
or Mcl-1 alone reduced cell proliferation. However, the
simultaneous downregulation of Bcl-xL and Mcl-1 significantly
reduced the proliferation of the two cell lines.
Subsequently, it was determined whether transfection
with siRNA induces apoptosis in colon cancer cells. DLD-1 and
HCT116 cells were transfected with siRNA targeting Bcl-xL, Mcl-1,
or Bcl-xL and Mcl-1 together for 24 h. Subsequently, the degree of
DNA fragmentation in the cells was measured. Downregulation of
Bcl-xL or Mcl-1 alone induced DNA fragmentation in DLD-1 and HCT116
cells. However, the simultaneous downregulation of Bcl-xL and Mcl-1
significantly induced DNA fragmentation (Fig. 4D).
Subsequently, it was determined whether the effect
of the simultaneous downregulation of Bcl-xL and Mcl-1 on apoptosis
was mediated by the activation of caspases. DLD-1 and HCT116 cells
were transfected with siRNA targeting Bcl-xL, Mcl-1 or Bcl-xL and
Mcl-1 together for 24 h and the degree of PARP cleavage was
evaluated using western blotting. Downregulation of Bcl-xL or Mcl-1
alone slightly increased the cleavage of PARP in DLD-1 and HCT116
cells. However, the simultaneous downregulation of Bcl-xL and Mcl-1
significantly increased the cleavage of PARP in the two cell lines
(Fig. 4E), which was consistent
with the results of the cell proliferation and DNA fragmentation
studies. These results suggest that the simultaneous downregulation
of Bcl-xL and Mcl-1 strongly induces the apoptosis of colon cancer
cells.
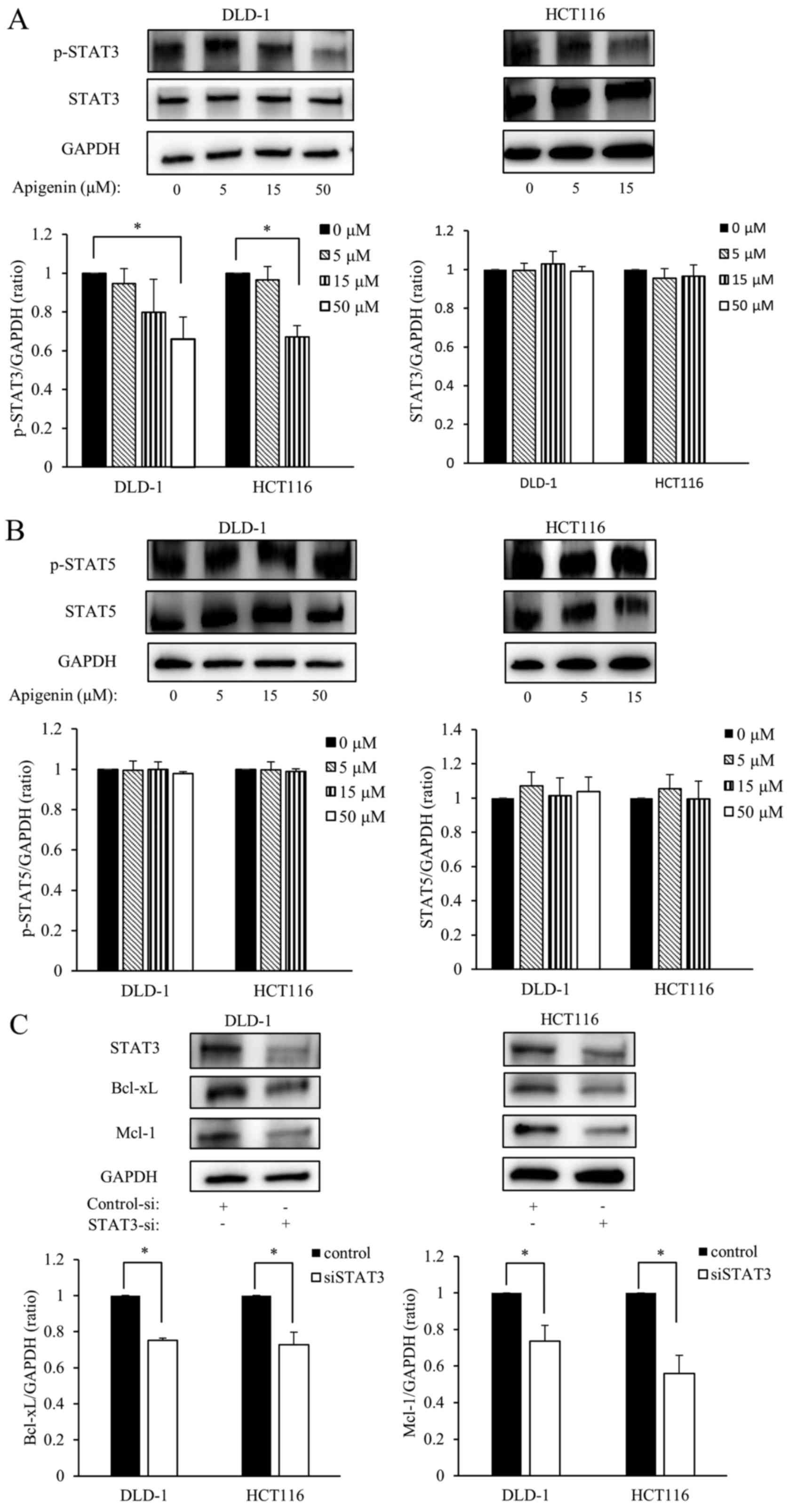
Apigenin reduces constitutive STAT3
phosphorylation and siRNA targeting STAT3 downregulates the
expression of Bcl-xL and Mcl-1
To further investigate the mechanism by which
apigenin induces apoptosis, the effect of apigenin on the
expression of p-STAT3 and STAT5 in DLD-1 and HCT116 cells was
investigated. DLD-1 and HCT116 cells were exposed to various doses
of apigenin for 24 h, then the expression of p-STAT3, p-STAT5 and
total STAT3, STAT5 were measured by western blotting. Levels of
p-STAT3 significantly decreased in a dose-dependent manner;
however, apigenin treatment did not affect the total amount of
STAT3 expression (Fig. 5A).
Furthermore, apigenin did not affect the expression of p- and total
STAT5 (Fig. 5B). Subsequently, it
was determined whether Bcl-xL and Mcl-1 were down-regulated by
STAT3 knockdown. DLD-1 and HCT116 cells were transfected with siRNA
targeting STAT3 for 24 h and subsequently the expression of Bcl-xL
and Mcl-1 were measured using western blotting. The results
demonstrated that levels of Bcl-xL and Mcl-1 were significantly
downregulated in DLD-1 and HCT116 cells following transfection with
siSTAT3 (Fig. 5C).
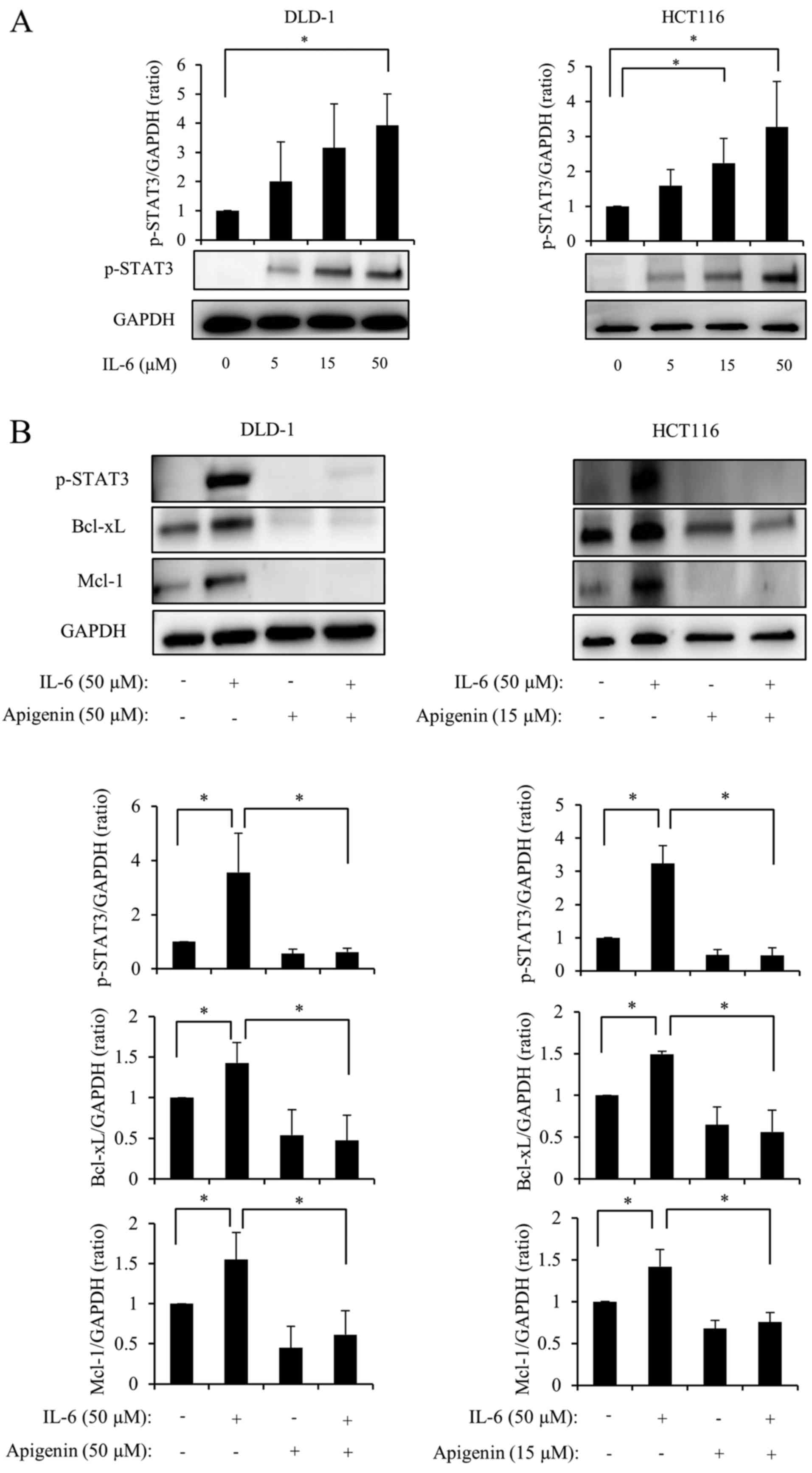
Apigenin suppresses Bcl-xL/Mcl-1 via
STAT3 signaling
It has been reported that IL-6 leads to STAT3
phosphorylation by activating janus kinase (22). DLD-1 and HCT116 cell lines were
administered various doses of IL-6 for 48 h and the expression of
p-STAT3 was measured using western blotting. The results indicated
that p-STAT3 expression increased in a dose-dependent manner
following treatment with IL-6 (Fig.
6A). Subsequently, 50 µM IL-6 was administered to cells
2 h following treatment with apigenin. After 48 h, levels of
p-STAT3, Bcl-xL and Mcl-1 were measured and it was demonstrated
that the expression of p-STAT3, Bcl-xL and Mcl-1 significantly
increased following stimulation with IL-6; however, this increase
was almost completely reversed following administration of apigenin
(Fig. 6B). Therefore, it was
suggested that apigenin suppresses the expression of Bcl-xL and
Mcl-1 via STAT3 signaling (Fig.
6C).
Discussion
The results of the current study demonstrated that
the natural dietary flavonoid, apigenin, reduces proliferation and
induces apoptosis in colon cancer cells. It was also demonstrated
that apigenin induces apoptosis by suppressing Bcl-xL and Mcl-1 and
that apigenin downregulated Bcl-xL and Mcl-1 by inhibiting the
phosphorylation of STAT3.
Apigenin is found in many fruits and vegetables and
is one of the most bioactive flavonoids (12). Apigenin exhibits low cytotoxicity
and has a marked effect on cancer cells compared with normal cells
(23). It has been reported that
apigenin reduces cell proliferation and induces apoptosis in colon
cancer cells (24–26). However, the detailed mechanism of
apigenin-induced apoptosis of colon cancer cells remains unknown.
Therefore, the molecular mechanism by which apigenin induces
apoptosis in colon cancer cells was assessed in the current
study.
The results demonstrated that the apoptosis inducing
effect of apigenin is mediated via the caspase cascade; therefore,
it was postulated that the anti-apoptotic Bcl-2 family proteins are
involved in the apoptosis-inducing effect of apigenin. Apigenin
suppressed the expression of Bcl-xL and Mcl-1 in all four colon
cancer cell lines. Previous studies demonstrated that apigenin
suppresses the expression of various anti-apoptotic Bcl-2 family
proteins, including Bcl-xL, Mcl-1 and Bcl-2 in several cancer cell
lines (27,28). However, it has only been
demonstrated that apigenin suppresses Bcl-xL and Mcl-1 in bladder
cancer and multiple myeloma cell lines (29,30).
To the best of our knowledge, the current study is the first to
report that apigenin suppresses the expression of Bcl-xL and Mcl-1
in colon cancer cells. Shao et al (13) reported that apigenin reduces Mcl-1
expression and sensitizes the Bcl-2 homology domain 3 mimetic
ABT-263. These results are similar to those of the current study,
although they did not measure Bcl-xL expression. The results of
present study demonstrated that apigenin simultaneously reduces
Bcl-xL and Mcl-1 expression and strongly induces apoptosis in colon
cancer cells, suggesting that apigenin itself exhibits strong
antitumor activity in colon cancer cells.
The anti-apoptotic Bcl-2 family of proteins serve a
protective role against various apoptotic stimuli (31). The over-expression of
anti-apoptotic Bcl-2 family proteins, including Bcl-xL and Mcl-1,
is observed in different types of human cancer and this
overexpression is involved in the development of resistance to
treatments such as chemotherapy, as well as in cell survival
(32,33). In the present study, the
overexpression of Bcl-xL and Mcl-1 was observed in four colon
cancer cell lines. Therefore, these proteins may be effective
therapeutic targets in colon cancer. The results demonstrated that
the simultaneous suppression of Bcl-xL and Mcl-1 strongly reduced
cell proliferation and induced apoptosis compared with the
suppression of each protein alone. This effect has been reported in
ovarian carcinoma, mesothelioma and pancreatic cancer (34–36),
but has not yet been reported in colon cancer. The results of the
present study and those of previous studies (34–36)
demonstrated that the simultaneous downregulation of Bcl-xL and
Mcl-1 induces cancer cell apoptosis.
STAT3 and STAT5 are upstream of Bcl-xL and Mcl-1
signaling (37). Therefore, it was
speculated that apigenin suppresses the expression of Bcl-xL and
Mcl-1 by inhibiting the signaling of STAT3 and STAT5. The present
study demonstrated that apigenin inhibits the phosphorylation of
STAT3 but does not inhibit the phosphorylation of STAT5. The marked
suppression of Bcl-xL and Mcl-1 expression was observed in the
DLD-1 and HCT116 cell lines following STAT3 knockdown. However,
STAT5 knockdown did not affect Bcl-xL and Mcl-1 expression. These
results indicate that apigenin does not induce apoptosis via STAT5
signaling. In addition, the expression of p-STAT3, Bcl-xL and Mcl-1
increased following IL-6 stimulation, but decreased following
administration of apigenin. This suggests that apigenin suppresses
Bcl-xL and Mcl-1 by inhibiting the phosphorylation of STAT3.
Four colon cancer cell lines were used in the
current study. HT-29 and COLO320 are KRAS wild-type whereas DLD-1
and HCT116 are KRAS mutant-type cells. Furthermore, the BRAF
mutation status (V600E) of DLD-1 and HCT116 is wild-type and that
of HT-29 is mutant (38).
Anti-EGFR therapy, which is effective at treating patients with
wild-type colon cancer, however, it generally ineffective against
KRAS or BRAF mutant colon cancer. However, the results of the
present study demonstrated that apigenin exhibited a similar effect
in all colon cell lines regardless of KRAS or BRAF status.
Therefore, apigenin may be beneficial at treating patients with
wild-type and mutant KRAS or BRAF colon cancer.
Apigenin may be an effective method of treating
patients with colon cancer; however, the mechanism by which
apigenin induces an antitumor effect in colon cancer has not been
reported. Shan et al (39)
reported that apigenin induces an antitumor effect by inhibiting
pyruvate kinase M2 (PKM2) activity via blocking of the
β-catenin/c-Myc/polypyrimidine tract-binding protein 1 signaling
pathway. McVean et al (40)
reported that apigenin induces G2/M arrest by inhibiting p34
cyclin-dependent kinase protein expression and activity in a
p21-independent manner. However, the results of the current study
indicated that apigenin induces an antitumor effect by inhibiting
the anti-apoptotic proteins Bcl-xL and Mcl-1 via the STAT3
signaling pathway. This suggests that apigenin acts on various
pathways and may therefore be developed as a novel treatment for
colon cancer. Furthermore, it was confirmed that Bcl-xL and Mcl-1
are strongly expressed in more advanced colon cancer by
immunostaining. This suggests that apigenin may be effective at
treating patients with advanced colon cancer.
In conclusion, the results of the current study
demonstrated that apigenin induces apoptosis by two anti-apoptotic
proteins, Bcl-xL and Mcl-1, via the STAT3 signaling pathway in
colon cancer cells. It was also demonstrated that the simultaneous
suppression of Bcl-xL and Mcl-1 induces strong apoptosis in colon
cancer cells. These data suggest that Bcl-xL and Mcl-1 are
important therapeutic targets in colon cancer and that apigenin may
be developed as an effective therapeutic agent for colon cancer by
simultaneously suppressing the expression of these two
proteins.
Acknowledgments
The authors wish to thank Nagoya City University for
providing valuable proposals and assistance to members of the
Department of Gastroenterological Surgery. The authors also wish to
thank Seiko Inumaru and Ryoko Hara for providing excellent
technical support.
Abbreviations:
|
DMSO
|
dimethyl sulfoxide
|
|
PARP
|
poly-(ADP-ribose) polymerase
|
|
STAT
|
signal transducer and activator of
transcription
|
Notes
[1]
Funding
No funding was received.
[2] Availability
of data and materials
All data generated or analyzed during this study are
included in the current published article.
[3] Authors’
contributions
HT, HI, YoM and ST conceived and designed the study.
NN and TY performed the cell proliferation assay, and analyzed and
interpreted the data on cell proliferation. NA and TO performed
immunohistochemistry, and analyzed and interpreted data on the
protein expression of resected specimens. KeS and TH performed
western blotting, and analyzed and interpreted data on protein
expression and change in cell lines. KaS and MH performed apoptosis
assay, and analyzed and interpreted data on apoptosis. YuM planned,
analyzed and interpreted all the experiments and drafted the
manuscript.
[4] Ethics
approval and consent to participate
Ethical approval for the use of human tissue was
granted by the Graduate School of Medicine, Nagoya City University
(Nagoya, Japan). All patients provided their written informed
consent for the use of their tissues.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee DH, Sung KS, Bartlett DL, Kwon YT and
Lee YJ: HSP90 inhibitor NVP-AUY922 enhances TRAIL-induced apoptosis
by suppressing the JAK2-STAT3-Mcl-1 signal transduction pathway in
colorectal cancer cells. Cell Signal. 27:293–305. 2015. View Article : Google Scholar
|
|
3
|
Vo MC, Nguyen-Pham TN, Lee HJ, Jaya
Lakshmi T, Yang S, Jung SH, Kim HJ and Lee JJ: Combination therapy
with dendritic cells and lenalidomide is an effective approach to
enhance antitumor immunity in a mouse colon cancer model.
Oncotarget. 8:27252–27262. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hail N Jr, Carter BZ, Konopleva M and
Andreeff M: Apoptosis effector mechanisms: a requiem performed in
different keys. Apoptosis. 11:889–904. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kirkin V, Joos S and Zörnig M: The role of
Bcl-2 family members in tumorigenesis. Biochim Biophys Acta.
1644:229–249. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Danial NN and Korsmeyer SJ: Cell death:
Critical control points. Cell. 116:205–219. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McDonnell TJ, Troncoso P, Brisbay SM,
Logothetis C, Chung LW, Hsieh JT, Tu SM and Campbell ML: Expression
of the protooncogene bcl-2 in the prostate and its association with
emergence of androgen-independent prostate cancer. Cancer Res.
52:6940–6944. 1992.PubMed/NCBI
|
|
8
|
Teixeira C, Reed JC and Pratt MA: Estrogen
promotes chemotherapeutic drug resistance by a mechanism involving
Bcl-2 proto-oncogene expression in human breast cancer cells.
Cancer Res. 55:3902–3907. 1995.PubMed/NCBI
|
|
9
|
Peddaboina C, Jupiter D, Fletcher S, Yap
JL, Rai A, Tobin RP, Jiang W, Rascoe P, Rogers MK, Smythe WR, et
al: The down-regulation of Mcl-1 via USP9X inhibition sensitizes
solid tumors to Bcl-xl inhibition. BMC Cancer. 12:5412012.
View Article : Google Scholar
|
|
10
|
Takahashi H, Chen MC, Pham H, Angst E,
King JC, Park J, Brovman EY, Ishiguro H, Harris DM, Reber HA, et
al: Baicalein, a component of Scutellaria baicalensis, induces
apoptosis by Mcl-1 down-regulation in human pancreatic cancer
cells. Biochim Biophys Acta. 1813:1465–1474. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Patel D, Shukla S and Gupta S: Apigenin
and cancer chemoprevention: Progress, potential and promise
(Review). Int J Oncol. 30:233–245. 2007.
|
|
12
|
Shukla S and Gupta S: Apigenin: A
promising molecule for cancer prevention. Pharm Res. 27:962–978.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shao H, Jing K, Mahmoud E, Huang H, Fang X
and Yu C: Apigenin sensitizes colon cancer cells to antitumor
activity of ABT-263. Mol Cancer Ther. 12:2640–2650. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Turkson J and Jove R: STAT proteins: Novel
molecular targets for cancer drug discovery. Oncogene.
19:6613–6626. 2000. View Article : Google Scholar
|
|
15
|
Bowman T, Garcia R, Turkson J and Jove R:
STATs in oncogenesis. Oncogene. 19:2474–2488. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xiong H, Su WY, Liang QC, Zhang ZG, Chen
HM, Du W, Chen YX and Fang JY: Inhibition of STAT5 induces G1 cell
cycle arrest and reduces tumor cell invasion in human colorectal
cancer cells. Lab Invest. 89:717–725. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Donato NJ, Wu JY, Zhang L, Kantarjian H
and Talpaz M: Down-regulation of
interleukin-3/granulocyte-macrophage colony-stimulating factor
receptor beta-chain in BCR-ABL(+) human leukemic cells: Association
with loss of cytokine-mediated Stat-5 activation and protection
from apoptosis after BCR-ABL inhibition. Blood. 97:2846–2853. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Garcia R, Bowman TL, Niu G, Yu H, Minton
S, Muro-Cacho CA, Cox CE, Falcone R, Fairclough R, Parsons S, et
al: Constitutive activation of Stat3 by the Src and JAK tyrosine
kinases participates in growth regulation of human breast carcinoma
cells. Oncogene. 20:2499–2513. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Seo HS, Ku JM, Choi HS, Woo JK, Jang BH,
Go H, Shin YC and Ko SG: Apigenin induces caspase-dependent
apoptosis by inhibiting signal transducer and activator of
transcription 3 signaling in HER2-overexpressing SKBR3 breast
cancer cells. Mol Med Rep. 12:2977–2984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cao HH, Chu JH, Kwan HY, Su T, Yu H, Cheng
CY, Fu XQ, Guo H, Li T, Tse AK, et al: Inhibition of the STAT3
signaling pathway contributes to apigenin-mediated anti-metastatic
effect in melanoma. Sci Rep. 6:217312016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tomita S, Yamauchi M, Ichikawa K, Mitomi H
and Fujimori T: The brand new trend of colorectal carcinoma
pathology. Nihon Rinsho. 72:63–70. 2014.In Japanese. PubMed/NCBI
|
|
22
|
Wang SW and Sun YM: The IL-6/JAK/STAT3
pathway: Potential therapeutic strategies in treating colorectal
cancer (Review). Int J Oncol. 44:1032–1040. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gupta S, Afaq F and Mukhtar H: Selective
growth-inhibitory, cell-cycle deregulatory and apoptotic response
of apigenin in normal versus human prostate carcinoma cells.
Biochem Biophys Res Commun. 287:914–920. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Turktekin M, Konac E, Onen HI, Alp E,
Yilmaz A and Menevse S: Evaluation of the effects of the flavonoid
apigenin on apoptotic pathway gene expression on the colon cancer
cell line (HT29). J Med Food. 14:1107–1117. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Horinaka M, Yoshida T, Shiraishi T, Nakata
S, Wakada M and Sakai T: The dietary flavonoid apigenin sensitizes
malignant tumor cells to tumor necrosis factor-related
apoptosis-inducing ligand. Mol Cancer Ther. 5:945–951. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee Y, Sung B, Kang YJ, Kim DH, Jang JY,
Hwang SY, Kim M, Lim HS, Yoon JH, Chung HY, et al: Apigenin-induced
apoptosis is enhanced by inhibition of autophagy formation in
HCT116 human colon cancer cells. Int J Oncol. 44:1599–1606. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Budhraja A, Gao N, Zhang Z, Son YO, Cheng
S, Wang X, Ding S, Hitron A, Chen G, Luo J, et al: Apigenin induces
apoptosis in human leukemia cells and exhibits anti-leukemic
activity in vivo. Mol Cancer Ther. 11:132–142. 2012. View Article : Google Scholar
|
|
28
|
Lu HF, Chie YJ, Yang MS, Lee CS, Fu JJ,
Yang JS, Tan TW, Wu SH, Ma YS, Ip SW, et al: Apigenin induces
caspase-dependent apoptosis in human lung cancer A549 cells through
Bax- and Bcl-2-triggered mitochondrial pathway. Int J Oncol.
36:1477–1484. 2010.PubMed/NCBI
|
|
29
|
Shi MD, Shiao CK, Lee YC and Shih YW:
Apigenin, a dietary flavonoid, inhibits proliferation of human
bladder cancer T-24 cells via blocking cell cycle progression and
inducing apoptosis. Cancer Cell Int. 15:332015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao M, Ma J, Zhu HY, Zhang XH, Du ZY, Xu
YJ and Yu XD: Apigenin inhibits proliferation and induces apoptosis
in human multiple myeloma cells through targeting the trinity of
CK2, Cdc37 and Hsp90. Mol Cancer. 10:1042011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Brunelle JK and Letai A: Control of
mitochondrial apoptosis by the Bcl-2 family. J Cell Sci.
122:437–441. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kelly PN and Strasser A: The role of Bcl-2
and its pro-survival relatives in tumourigenesis and cancer
therapy. Cell Death Differ. 18:1414–1424. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Czabotar PE, Lessene G, Strasser A and
Adams JM: Control of apoptosis by the BCL-2 protein family:
Implications for physiology and therapy. Nat Rev Mol Cell Biol.
15:49–63. 2014. View Article : Google Scholar
|
|
34
|
Brotin E, Meryet-Figuière M, Simonin K,
Duval RE, Villedieu M, Leroy-Dudal J, Saison-Behmoaras E, Gauduchon
P, Denoyelle C and Poulain L: Bcl-XL and MCL-1 constitute pertinent
targets in ovarian carcinoma and their concomitant inhibition is
sufficient to induce apoptosis. Int J Cancer. 126:885–895.
2010.
|
|
35
|
Varin E, Denoyelle C, Brotin E,
Meryet-Figuière M, Giffard F, Abeilard E, Goux D, Gauduchon P,
Icard P and Poulain L: Downregulation of Bcl-xL and Mcl-1 is
sufficient to induce cell death in mesothelioma cells highly
refractory to conventional chemotherapy. Carcinogenesis.
31:984–993. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Takahashi H, Chen MC, Pham H, Matsuo Y,
Ishiguro H, Reber HA, Takeyama H, Hines OJ and Eibl G: Simultaneous
knock-down of Bcl-xL and Mcl-1 induces apoptosis through Bax
activation in pancreatic cancer cells. Biochim Biophys Acta.
1833:2980–2987. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Buettner R, Mora LB and Jove R: Activated
STAT signaling in human tumors provides novel molecular targets for
therapeutic intervention. Clin Cancer Res. 8:945–954.
2002.PubMed/NCBI
|
|
38
|
Sueda T, Sakai D, Kawamoto K, Konno M,
Nishida N, Koseki J, Colvin H, Takahashi H, Haraguchi N, Nishimura
J, et al: BRAF V600E inhibition stimulates AMP-activated protein
kinase-mediated autophagy in colorectal cancer cells. Sci Rep.
6:189492016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shan S, Shi J, Yang P, Jia B, Wu H, Zhang
X and Li Z: Apigenin restrains colon cancer cell proliferation via
targeted blocking of pyruvate kinase M2-dependent glycolysis. J
Agric Food Chem. 65:8136–8144. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
McVean M, Weinberg WC and Pelling JCA: A
p21(waf1)-independent pathway for inhibitory phosphorylation of
cyclin-dependent kinase p34(cdc2) and concomitant G(2)/M arrest by
the chemopreventive flavonoid apigenin. Mol Carcinog. 33:36–43.
2002. View Article : Google Scholar : PubMed/NCBI
|