Introduction
Increased neutrophil counts can sometimes predict a
poor prognosis among patients with advanced cancers (1-3).
Pan-cancer analysis performed by Templeton et al aided in
the identification of polymorphonuclear leukocyte signaling as the
most potent predictor of a poor prognosis (4). Increased neutrophil/lymphocyte (N/L)
ratios have recently been attracting interest as a suitable
prognostic marker for cancer patients (4,5).
Several reports have indicated that increased N/L ratios are a
marker of a poor prognosis and a weak response to chemotherapy
among patients with ovarian cancer (6,7).
Although a number of studies have demonstrated the significance of
increased neutrophil counts as a prognostic marker, an in-depth
analysis has not yet been performed regarding its role in cancer
progression, at least to the best of our knowledge.
Oncogenes are known to promote cell proliferation
and prevent apoptosis in malignant cells. Although the association
between oncogenes and tumor microenvironment (TME) modifications
has not yet been sufficiently elucidated, recent studies have
revealed that several oncogenes are associated with TME modulation
(8,9). KRAS is a frequently mutated oncogene
in cancer. In a previous study, we established a KRAS-transduced
mouse ovarian cancer cell line, ID8-KRAS, and demonstrated that the
oncogene KRAS promoted tumorigenesis and aggravated cancer-induced
inflammation, accompanied by an increased number of neutrophils in
ascites (8). RAS is considered to
promote cancer progression by sustaining proliferation, metabolic
reprogramming, anti-apoptosis, and remodeling of the TME (10,11).
RAS followed by the activation of RAS-GTP and PI3K/AKT signaling
may upregulate nuclear factor (NF)-κB activity and lead to an
inflammatory microenvironment (12). Similarly, several oncogenes may be
associated with severe inflammation accompanied by an increased
number of neutrophils.
Although neutrophils have been regarded as a
protumor marker, the role of tumor-associated neutrophils (TANs) in
cancer progression has only recently become a focus area (13). There are two types of TANs: An
interferon (IFN)-β-induced antitumor type (14) and a transforming growth factor
(TGF)-β-induced protumor type (15). The majority of studies have focused
on the protumor properties of neutrophils and have associated
neutrophils with a poor prognosis (1,16,17).
Recent findings have revealed that TANs exert antitumor effects in
several cancer types through the stimulation of T cell activity
(17,18). However, the role of neutrophils in
the oncogene-induced TME has not yet been elucidated.
In the KRAS-transduced ovarian cancer model, a
marked increase in the number of neutrophils was observed; however,
it is still unclear whether the increased number of neutrophils
exerts a pro-tumor or antitumor effect in this model. Therefore, in
this study, we investigated the role of recruited neutrophils in
KRAS-induced ovarian cancer progression, as well as their influence
on the intraperitoneal microenvironment.
Materials and methods
Cell lines and establishment of
oncogene-transduced ID8 cells
We established oncogene-transduced mouse ovarian
cancer cell lines by transducing KRAS into the mouse ovarian
epithelial immortalized cell line, ID8, which was established from
C57BL/6 mice (8). The ID8 cells
were a kindly gift from Dr Kathy Roby, Department of Anatomy and
Cell Biology, University of Kansas Medical Center (Kansas City, KS,
USA). An oncogenic mutant form of human KRAS (KRASG12V)
was recombined into pDEST-CLXSN to generate
pCLXSN-KRASG12V. Retrovirus packaging was performed as
previously described (19).
ID8-KRAS cells were established by infection of the
LXSN-KRASG12V virus at a multiplicity of infection of 1,
followed by G418 selection at a concentration of 800 µg/ml
for 1 week.
Mouse model
The ID8 and ID8-KRAS cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; Wako, Tokyo, Japan,
043-30085) containing 10% FBS, 100 U/ml penicillin, 0.1 mg/ml
streptomycin, and 0.25 g/ml amphotericin B. The ID8 and ID8-KRAS
cells (2×106) suspended in 1,000 µl DMEM were
injected into the peritoneal cavities of 8-week-old female C57/BL6
mice as previously described (8).
The mice were obtained from Japan SLC and weighed 18-20 g. The
animals were maintained at room temperature in a
humidity-controlled room with a 12 h-light/12 h-dark cycle and were
provided sterilized solid food and water ad libidum during the
entire experimental period. Mice were sacrificed to minimize
suffering when moribund behaviors were observed. In all cases,
tissue collection procedures were initiated after animals had been
euthanized by isoflurane overdose; mice were placed into a chamber
filled with the vapor of the anesthetic isoflurane until
respiration ceased (within 2 min) (20). For the mouse survival analysis,
mice were sacrificed when their body weight (BW) exceeded 23 g
after inoculation as in our previous study (8), we confirmed that that approximately 5
ml of ascites were accumulated with the formation of dissemination
when the BW reached 23 g. BW and ascites weight were assessed at
the time of sacrifice. The total number of mice used in this study
was 115 as follows: Experiment of the in vitro effects of
neutrophils on CD8 T cell activation, 32 mice (no cancer mice,
n=16; ID8 mice, n=8; ID8-KRAS mice, n=8); experiment of T cell
costimulatory molecules on neutrophils, 83 mice (CD80: no cancer,
n=8; ID8, n=5; ID8-KRAS, n=7; CD86: no cancer, n=6; ID8, n=6;
ID8-KRAS, n=5; 4-1BBL: no cancer, n=11; ID8, n=5; ID8-KRAS, n=7;
OX40L: no cancer, n=10; ID8, n=5; ID8-KRAS, n=8).
Neutrophil depletion
Neutrophils were depleted using anti-Ly6G mAb
(mouse) (Nimp-R14, AdipoGen Life Sciences, San Diego, CA, USA,
AG-20B-0043-C100). This mAb has been reported to selectively
deplete neutrophils in vivo (21,22).
The mice were injected intraperitoneally with 250 µg
anti-Ly6G mAb or matched isotype (rat IgG2b isotype control, Bio X
Cell, West Lebanon, NH, USA, BE0090) in 0.5 ml of PBS or with PBS
alone every 3 days by the following two medication methods: Method
A, from day 7 after inoculation of ID8-KRAS cells. Mice were
sacrificed when their body weight exceeded 23 g (isotype, n=8; and
anti-Ly6G, n=9). Method B, from day 10 after inoculation of
ID8-KRAS cells. Mice were sacrificed on day 16 (isotype, n=15; and
anti-Ly6G, n=16). As a control, we used mice with no cancer which
were not injected with any substance (method A, n=6; method B,
n=6).
Isolation of myelocyte peritoneal
ascites
Mice were peritoneally inoculated with ID8-KRAS
cells by the described medication described above in method B.
Following the injection of 3 ml of PBS, peritoneal myelocytes were
recovered from peritoneal cavities of no cancer mice (n=6) and
ID8-KRAS mice treated with anti-Ly6G or isotype mAb. Peritoneal
myelocytes were centrifuged at 300 × g at room temperature for 5
min. Red blood cells were removed by RBC lysed.
Isolation of neutrophils from blood and
peritoneal ascites
After euthanized by isoflurane overdose, peritoneal
ascites and approximately 1.5 ml of blood by cardiac puncture were
obtained from ID8 and ID8-KRAS mice. Neutrophils were isolated from
the collected peritoneal ascites and blood using a magnetic cell
sorting kit (Neutrophil Isolation kit, MACS KK; Miltenyi Biotec,
Cologne, Germany) according to the manufacturer's instructions.
Isolation of lymphocytes from ascites and
splenocytes
Mice were peritoneally inoculated with ID8-KRAS
cells by the described medication method B. Peritoneal cells were
obtained from peritoneal cavities of no cancer mice and ID8-KRAS
mice treated with anti-Ly6G or isotype mAb after injection of 3 ml
of PBS. Splenocytes were prepared as follows: spleens obtained from
no cancer mice and ID8-KRAS mice treated with anti-Ly6G or isotype
mAb were homogenized between glass slides, RBC lysed, washed twice,
and filtered with 100 µm filter. Peritoneal cells and
splenocytes were centrifuged (300 × g, 15 min) to pellet them. The
pellet was resuspended in 3 ml of PBS and then layered onto a
discontinuous Percoll gradient (44%/70%; Percoll, GE Healthcare,
Chicago, IL, USA) followed by centrifugation at 400 × g for 18 min.
The band between the 44 and 70% layers was pipetted into another
round-bottom centrifuge tube, diluted with 10 ml of PBS, and
centrifuged at 300 × g for 5 min to remove the Percoll or small
particulates. The collected pellet was then used for flow
cytometric analysis.
Isolation of naïve CD8+ T
cells from splenocytes
Splenocytes were obtained from the spleens of no
cancer mice. Naïve CD8+ T cells from splenocytes were
isolated using a magnetic cell sorting kit (Naïve CD8a+
T Cell Isolation Kit, mouse, MACS KK; Miltenyi Biotec) according to
the manufacturer's instructions.
Flow cytometry
Samples (2×106 cells/ml) were suspended
in 500 µl of PBS/1% BSA and analyzed by flow cytometry
(FACSCalibur flow cytometer; Becton-Dickinson, Mountain View, CA,
USA). Matched isotype antibodies were used as controls. T cells and
neutrophils were stained with the antibodies listed in Table I.
 | Table IAntibodies used in flow
cytometry. |
Table I
Antibodies used in flow
cytometry.
| Antibody (clone,
company, cat. no.) | Isotype (clone,
company, cat. no.) |
|---|
| FITC-anti-mouse
CD45 (B3821F4A/N901/UCHT1, Beckman Coulter, Tokyo, Japan,
CO6607071) | FITC-rat IgG, 2b, k
isotype (eBRG1, eBioscience, 11-4031-82) |
| FITC-anti-mouse
c-mesenchymal-epithelial transition (eBioclone7, eBioscience,
Tokyo, Japan, 11-8854-80) | FITC-rat IgG1, k
isotype (eBRG1, eBioscience, 11-4301-82) |
| FITC-anti-mouse
CD80 (16-10A1, BioLegend, Tokyo, Japan, 104705) | FITC-Arm Hamster,
IgG isotype (eBio299Arm, eBioscience, 11-4888-81) |
| FITC-anti-mouse
CD86 (GL1, BioLegend, 105005) | FITC-rat IgG, 2b, k
isotype (eB149/10H5, eBioscience, 11-4031-82) |
| PE-anti-mouse 4-1BB
ligand (TKS-1, BioLegend, 107105) | PE-mouse IgG, 2a, k
isotype (MOPC-173, BioLegend, 400211) |
| FITC-anti-mouse
CD11b (M1/70, eBioscience, 11-0122-85) | FITC-rat IgG, 2b, k
isotype (eBRG1, eBioscience, 11-4031-82) |
| APC-anti-mouse Ly6G
(RB6-8C5, eBioscience, 17-5931-82) | APC-Rat, IgG, 2a, k
isotype (eBR2a, eBioscience, 17-4321-81) |
| PE-anti-mouse Ly6G
(RB6-8C5, eBioscience, 12-5931-82) | PE-mouse IgG, 2a, k
isotype (MOPC-173, BioLegend, 40021) |
| APC-anti-mouse Ly6C
(HK1.4, BioLegend, 128015) | APC-Rat, IgG, 2c, k
isotype (RTK4174, BioLegend, 400713) |
|
PE-Cyanine5-anti-mouse F4/80 (BM8,
BioLegend, 123111) | PE-Cyanine5-Rat,
IgG, 2a, k isotype (eBR2a, eBioscience, 35-4321-80) |
| FITC-anti-mouse
CD8a (53-6.7, eBioscience, 11-0081-81) | FITC-Rat, IgG, 2a,
k isotype (eBR2a, eBioscience, 11-4321-82) |
|
PE-Cyanine5-anti-mouse CD3e (145-2C11,
eBioscience, 15-0031-82) | PE-Cyanine5-Arm
Hamster, IgG isotype (eBio299Arm, eBioscience, 15-4888-82) |
| APC-anti-mouse CD4
(RM4-5, eBioscience, 17-0042-81) | APC-Rat, IgG, 2a, k
isotype (eBR2a, BioLegend, 400512) |
| FITC-anti-mouse
CD25 (PC61.5, BD, 558689) | FITC-mouse IgG1, λ
isotype (G0114F7, BioLegend, 401913) |
| PE-anti-mouse FoxP3
(NRRF-30, eBioscience, 12-4771-80) | PE-mouse IgG, 2a, k
isotype (MOPC-173, BioLegend, 400211) |
ELISA
Cytokine levels [interleukin (IL)-6 and IFN-γ] were
measured in ascites or culture media with a specific ELISA kit
(DuoSet ELISA; R&D Systems, Minneapolis, MN, USA) according to
the manufacturer's instructions.
Coculture of neutrophils and naïve
CD8+ T cells
T cell proliferation induced by plate-bound
anti-mouse CD3ε (1 µg/ml; BioLegend, San Diego, CA, USA;
clone: 145-2C11) was assessed using standard carboxyfluorescein
diacetate succinimidyl ester (CFSE; Cayman Chemical, Ann Arbor, MI,
USA) dilution methods. Purified naïve CD8+ T cells
(2.0×106 cells/ml) were labeled with CFSE and cocultured
with neutrophils (2.0×106 cells/ml) obtained from the
blood or ascites of each mouse (no-cancer control, ID8 mice, or
ID8-KRAS mice) in CD3-coated plates for 3 days in complete cell
culture medium. The complete cell culture medium was composed of
RPMI-1640 medium (Wako) supplemented with 10% FBS, 100 U/ml
penicillin, 0.1 mg/ml streptomycin and 0.25 g/ml amphotericin B. To
establish whether T cell proliferation was induced by intercellular
communication, we assessed cocultures using chambers. Five hundred
microliters of neutrophils obtained from the ascites of ID8-KRAS
mice were added to the upper compartments of the chambers at a
concentration of 2.0×106 cells/ml. Five hundred
microliters of purified naïve CD8+ T cells were seeded
onto the bottom chamber at a concentration of 2.0×106
cells/ml. Following a 3-day incubation, cells on the upper chamber
were completely removed. The CFSE signal was analyzed by flow
cytometry.
Statistical analysis
Data are presented as the means ± SEM. The log-rank
test was used to detect differences in animal survival
(Kaplan-Meier survival curves). All other comparisons were
performed with the two-tailed Student's t-test. For performing
multiple comparisons, all the P-values were adjusted by the Holm's
method using the Microsoft Office Excel 2015 (BellCurve, Tokyo,
Japan). Other statistical analyses were conducted using JMP11 (SAS
Institute Japan, Tokyo, Japan). Other statistical analyses were
conducted using JMP11 (SAS Institute Japan, Tokyo, Japan). A value
of P<0.05 was considered to indicate a statistically significant
difference.
Study approval
Animal studies were approved by the University of
Tokyo Animal Committee. Our IACUC permitted this study and provided
this study the approval numbers P-14-027 and P15-060.
Results
Neutrophil depletion modulates the
peritoneal microenvironment
We previously reported that the neutrophil count is
markedly increased in ID8-KRAS-induced ascites (8), and demonstrated that neutrophil
recruitment into the peritoneal cavity begins at around 10 days
after the intraperitoneal injection. Therefore, in this study, we
focused in particularly on neutrophil functions in the late phase
and began the depletion of neutrophils 10 days after the injection.
To investigate the role of increased neutrophils in the peritoneal
microenvironment, neutrophils were depleted with a specific
antibody, anti-Ly6G mAb, and its isotype antibody was used as the
control. Profiles of intraperitoneal myelocytes were assessed by
flow cytometry. We first confirmed that the isotype antibody had no
effect on the myelocyte population in ascites (data not shown).
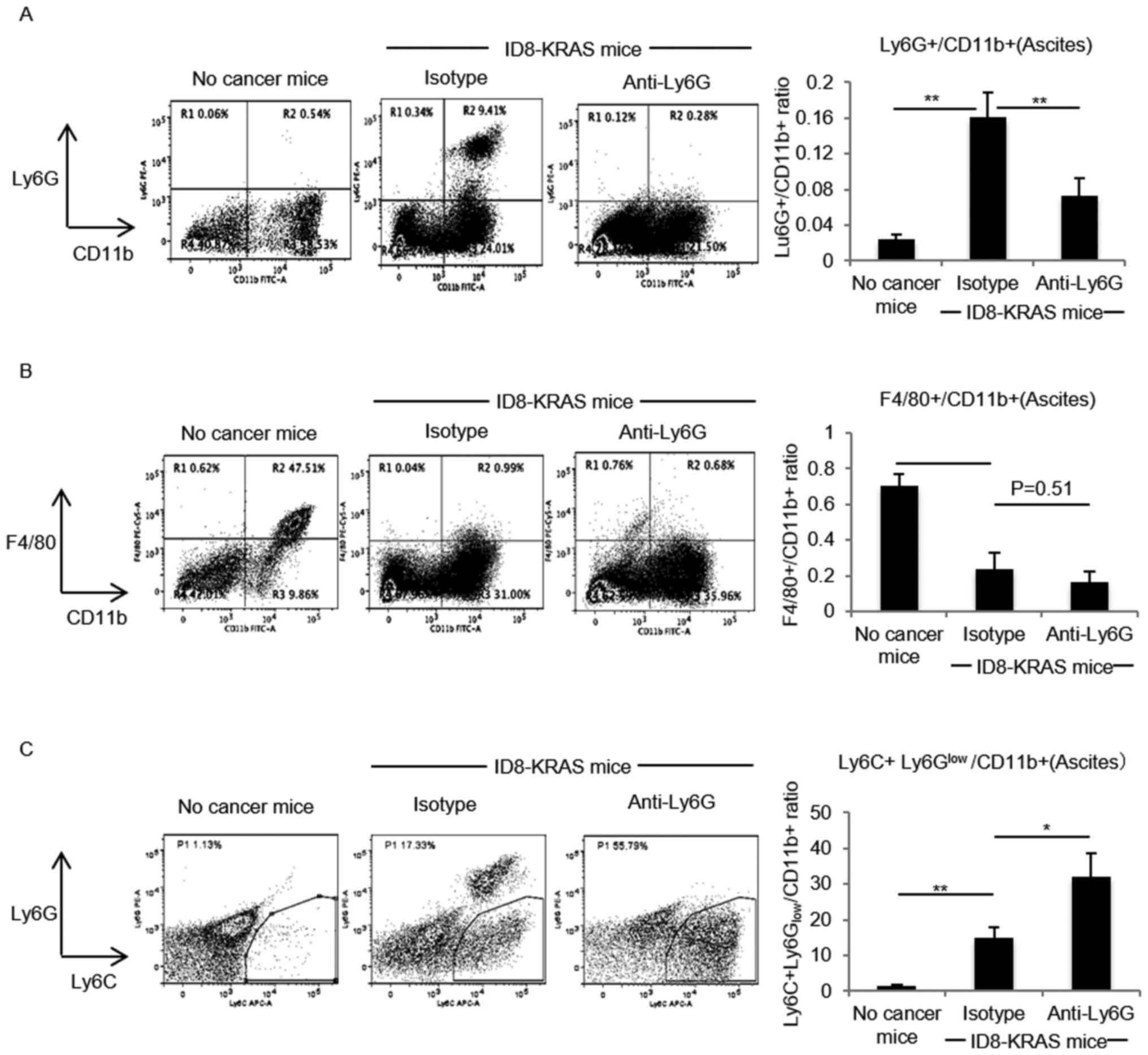
When compared with the isotype antibody, the anti-Ly6G antibody
successfully decreased the Ly6G+/CD11b+
neutrophil numbers (P<0.01, Fig.
1A); however, no significant difference was observed in the
number of F4/80+/CD11b+ macrophages between
the isotype and neutrophil-depleted groups (P=0.51, Fig. 1B). Focusing on the profiles of
myeloid-derived suppressor cells (MDSCs), the amount of
Ly6C+Ly6Glow/CD11b+monocytic-MDSCs
(M-MDSCs) was significantly increased between the isotype and
neutrophil-depleted groups (P<0.05, Fig. 1C).
Neutrophil depletion accelerates cancer
progression and aggravates inflammation
We then investigated the effects of neutrophils on
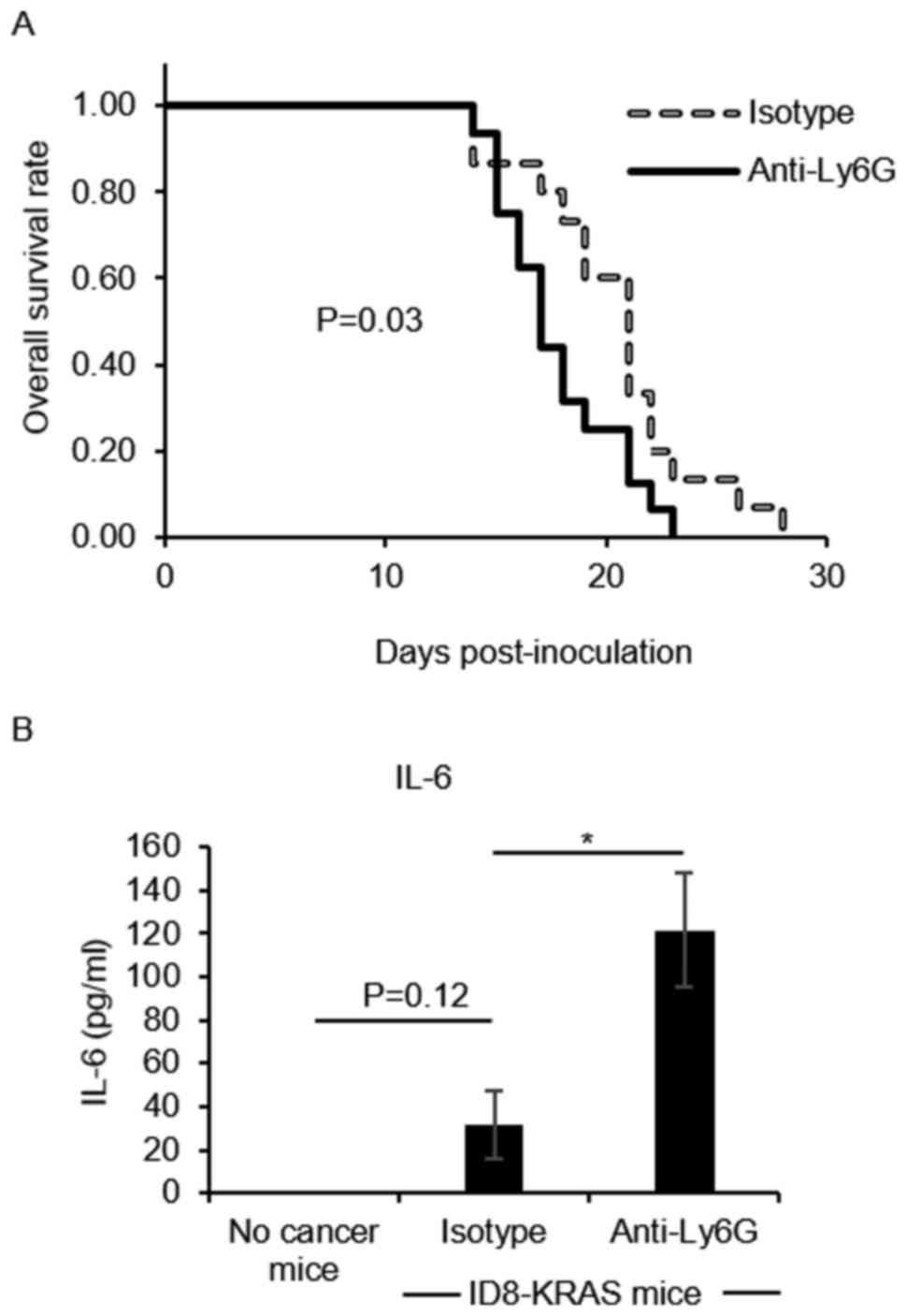
cancer progression. We first confirmed that the isotype antibody
had no effect on the accumulation of ascites (P=0.18, data not
shown). When compared with the isotype group, the
neutrophil-depleted group exhibited a more rapid accumulation of
ascites (P=0.03, Fig. 2A).
A higher level of IL-6 is a hallmark of ovarian
cancer progression (23,24). In addition, increases in IL-6
production have been observed in KRAS-induced ovarian cancer
(8); therefore, in this study, we
measured the IL-6 concentration as a surrogate marker of cancer
progression. Neutrophil depletion significantly increased the IL-6
concentration in ascites compared with that in the isotype group
(P<0.05, Fig. 2B).
Neutrophil depletion modulates the T cell
profile in ascites and spleen
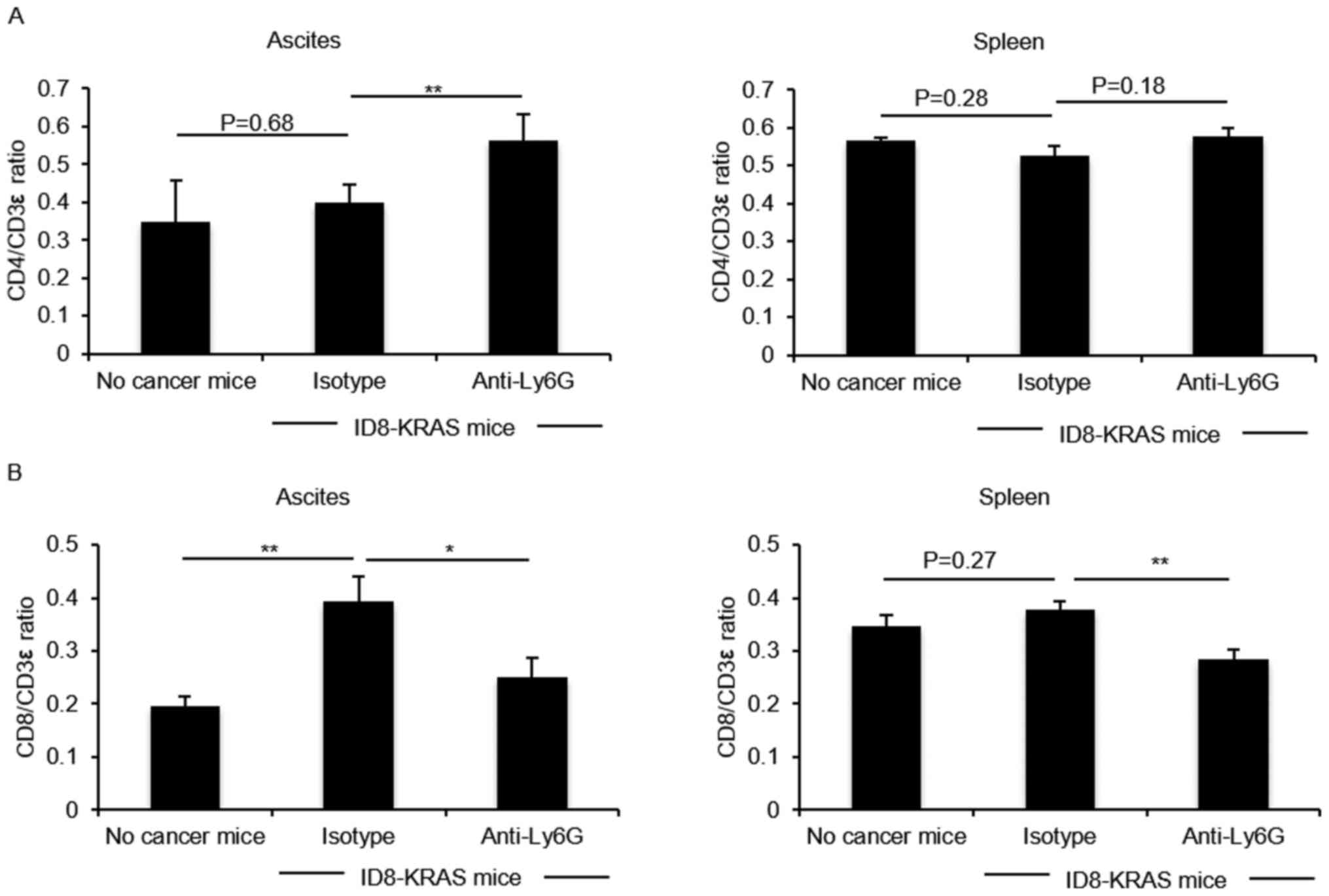
We examined the effects of neutrophils on the T
lymphocyte population. Differences in the CD4/CD3ε ratio and
CD8/CD3ε ratio in the spleen and ascites were compared between the
isotype and neutrophil-depleted groups. The CD4/CD3ε ratio in the
KRAS-induced ascites was significantly increased by neutrophil
depletion (P<0.05, Fig. 3A). On
the contrary, the CD8/CD3e ratio in the KRAS-induced ascites and
spleen was reduced by neutrophil depletion (P<0.05, Fig. 3B).
Neutrophil depletion increases the number
of regulatory T cells in ascites
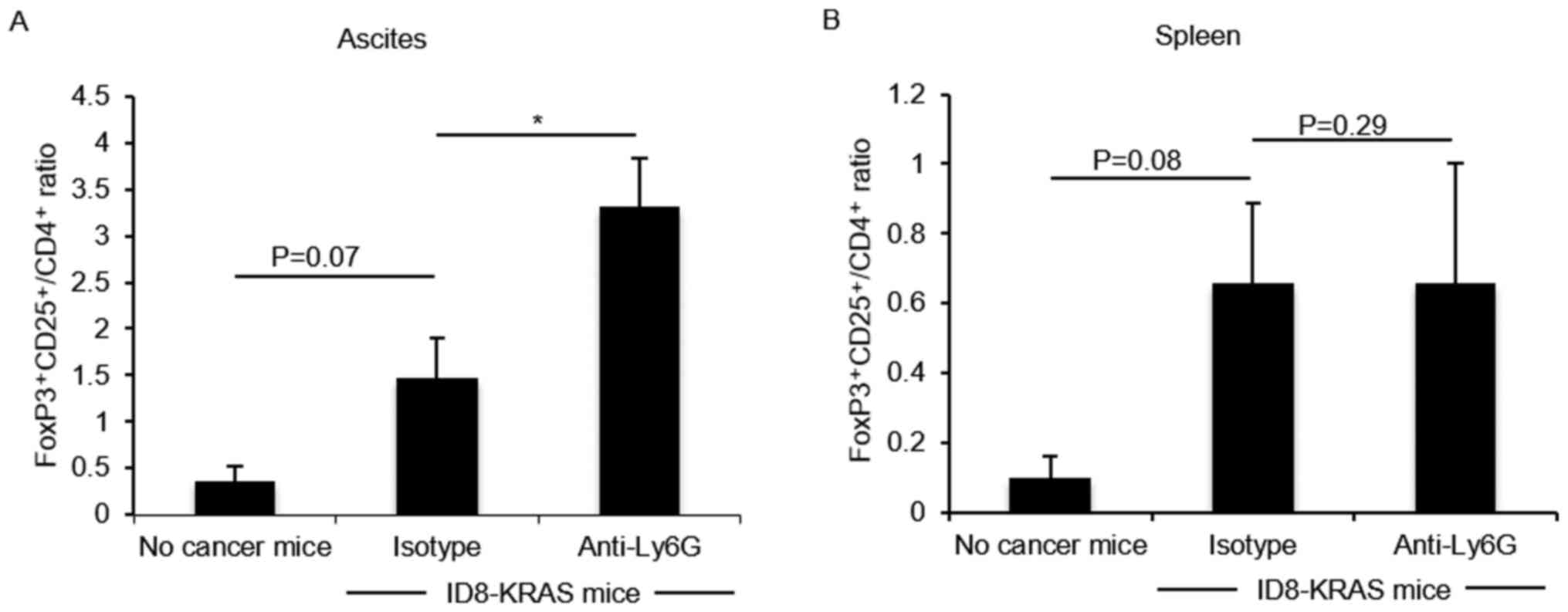
Previous studies have demonstrated that M-MDSCs
induce regulatory T cell (Treg) differentiation (25-27).
The data of this study indicated that an increased number of
Ly6C+Ly6Glow/CD11b+ M-MDSCs was accompanied
by an increased number of CD4+ T cells. Therefore, we
hypothesized that in the neutrophil-depleted group, the number of
Tregs would be increased. The number of Tregs was assessed by
FoxP3-CD25-CD4 flow cytometry. The number of
FoxP3+CD25+/CD4+ Tregs tended to
be increased in the KRAS-induced ascites, and their number was
markedly increased by neutrophil depletion (P<0.05, Fig. 4A); however, their number did not
significantly differ in the spleen (Fig. 4B).
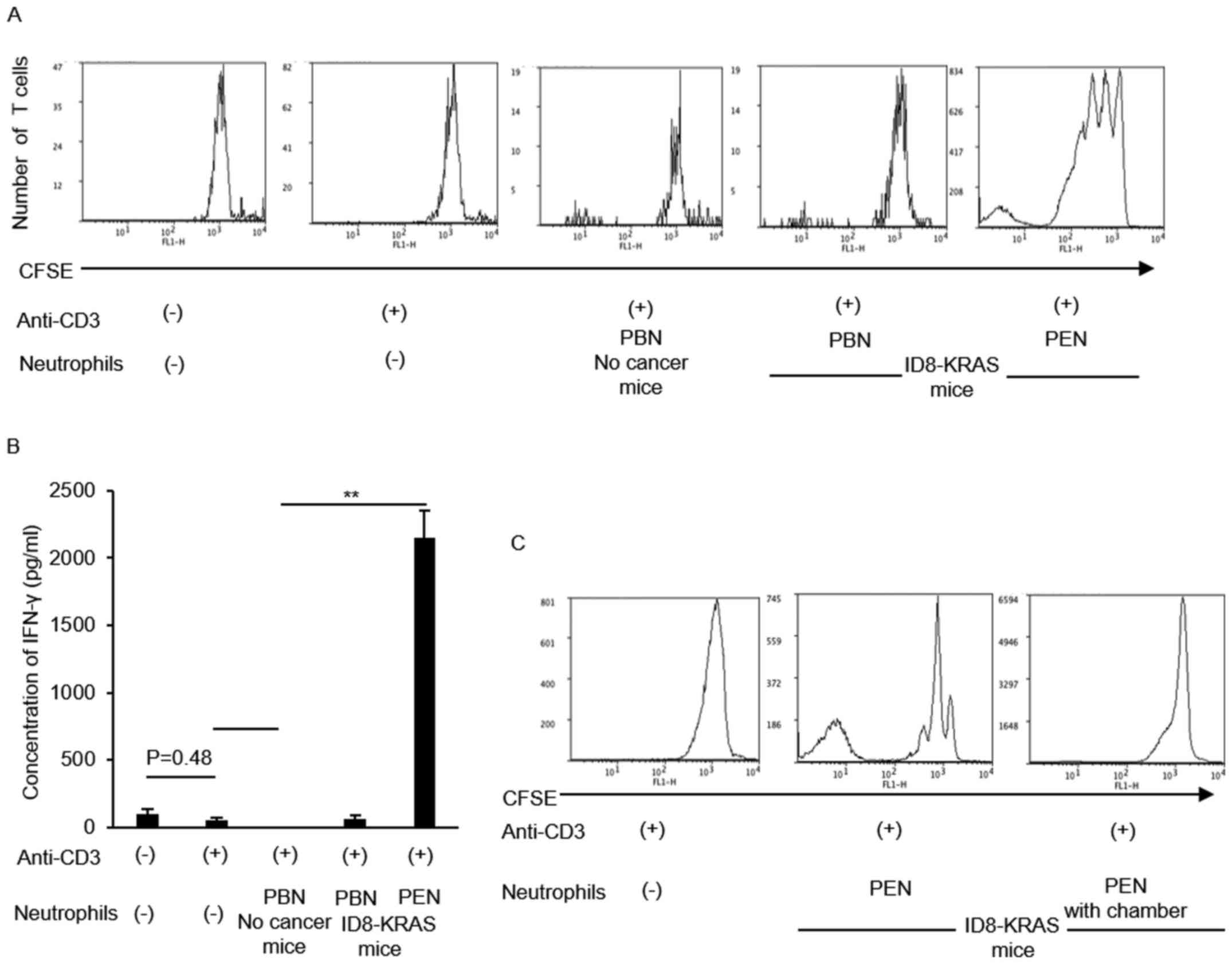
Neutrophils from ID8-KRAS ascites
stimulated CD8 T cell activity
The results of the in vivo assessment
suggested that neutrophils also modulate CD8+ T cell
activity. Therefore, we assessed the effects of neutrophils on
CD8+ T cell activity in vitro. Naïve
CD8+ T cells were cultured with anti-CD3 antibody alone
or with peripheral blood neutrophils (PBNs) or peritoneal
neutrophils (PENs) from each mouse (no cancer mice and ID8-KRAS
mice). The proliferation of naïve CD8+ T cells was
assessed using CFSE staining. CD8+ T cell proliferation
markedly increased in coculture with PENs from ID8-KRAS mice
(Fig. 5A). IFN-γ production by
CD8+ T cells was also assessed in each group using
specific ELISA kits. When compared with the PBNs from no cancer
mice or ID8-KRAS mice, coculture with PENs from ID8-KRAS mice
markedly increased IFN-γ production by naïve CD8+ T
cells (Fig. 5B). The T cell
stimulatory effects of PENs from ID8-KRAS mice were reversed when
PENs from ID8-KRAS mice were added to the coculture chamber,
suggesting that cell-to-cell contact was indispensable for the T
cell stimulatory effects of KRAS-related neutrophils (Fig. 5C).
T cell costimulatory molecules are
expressed at high levels in neutrophils from ID8-KRAS ascites
To elucidate the mechanisms through which PENs from
ID8-KRAS mice strongly promote IFN-γ production and the
proliferation of CD8+ T cells, T cell costimulatory
molecules CD80, CD86, OX40 ligand (OX40L) and 4-1BB ligand (4-1BBL)
on the surface of neutrophils from no cancer mice, ID8 mice and
ID8-KRAS mice were assessed by flow cytometry. Almost no difference
in CD80 and CD86 expression in the PBNs and PENs was observed among
these 3 groups (Fig. 6A and B).
Furthermore, no significant differences in CD80 and CD86 expression
between the PBNs and PENs was noted in each mouse group (Fig. 6A and B). The ID8-KRAS mice
expressed higher levels of 4-1BBL in both the PBNs and PENs
(Fig. 6C) (PBNs: no cancer vs.
ID8-KRAS, P<0.01; ID8 vs. ID8-KRAS, P=0.05; PENs: ID8 vs.
ID8-KRAS, P<0.05). No significant differences were observed
between the PBNs and PENs in each mouse group (ID8 mice, P=0.20;
ID8-KRAS mice, P=0.10, respectively). Marked differences were
observed in the expression levels of OX40L. In both the PBNs and
PENs, the OX40L expression levels were significantly higher in the
ID8-KRAS mice than in the no cancer mice and ID8 mice (Fig. 6D) (PBNs: no cancer vs. ID8-KRAS,
P<0.01; ID8 vs. ID8-KRAS, P=0.03; PENs: ID8 vs. ID8-KRAS,
P<0.05). When the PBNs and PENs from each mouse were compared,
the PENs from the ID8-KRAS mice exhibited a higher expression of
OX40L than the PBNs (Fig. 6D) (ID8
mice, P=0.30; ID8-KRAS mice, P<0.01, respectively).
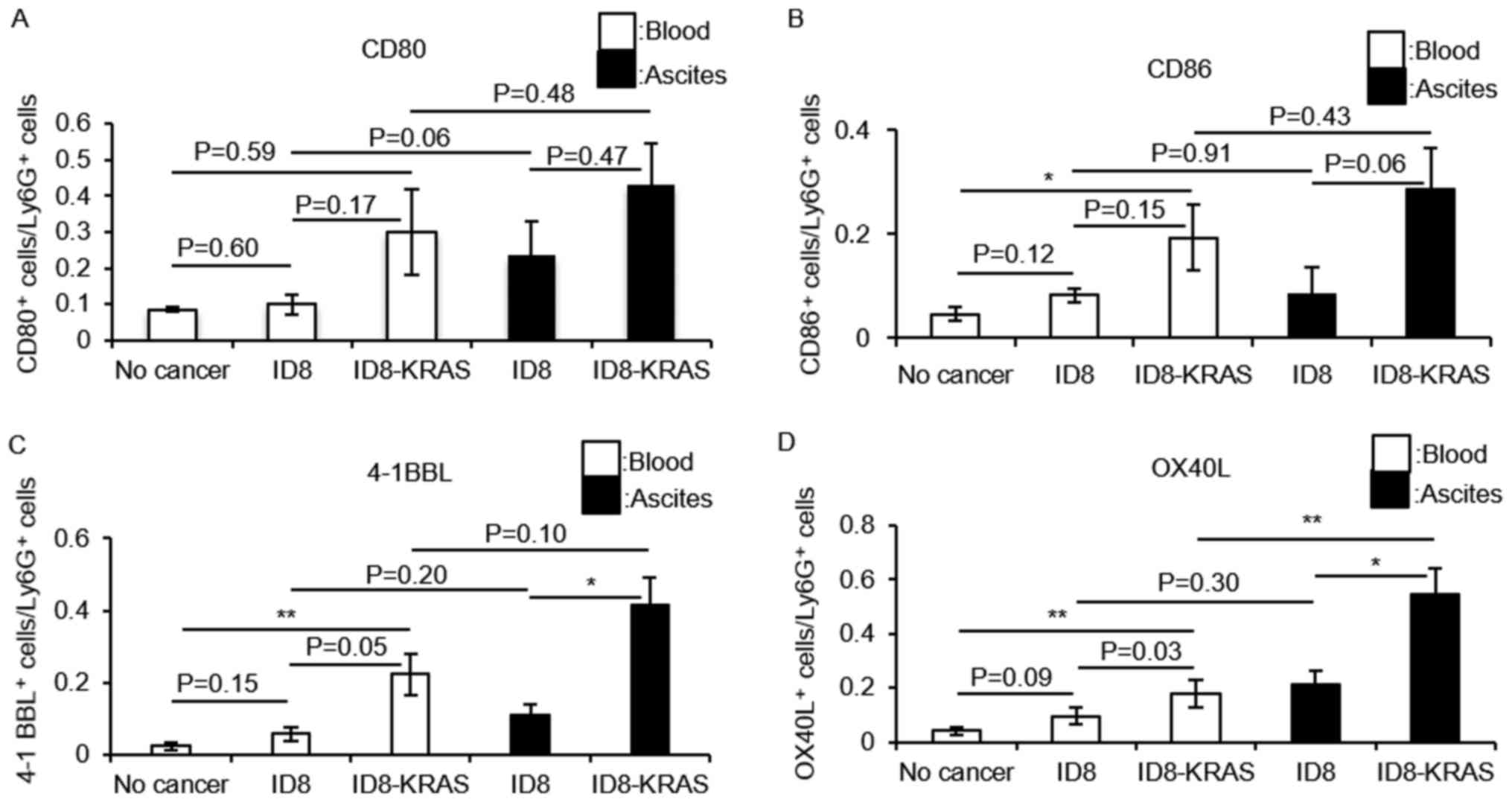 | Figure 6Expression of T cell costimulatory
molecules on neutrophils. Murine ID8 cells (2×106) and
ID8-KRAS cells (2×106) were injected into mice, and
neutrophils were obtained from ascites or blood when the body
weight exceeded 23 g. Neutrophils from no cancer mice were used as
a control. (A-D) The expression of costimulatory molecules on the
gated Ly6g+ neutrophils from ascites or blood was
analyzed by flow cytometry. The number of mice used in each
experiment was as follows: (A) CD80: no cancer, n=8; ID8, n=5;
ID8-KRAS, n=7; (B) CD86: no cancer, n=6; ID8, n=6; ID8-KRAS, n=5;
(C) 4-1BBL: no cancer, n=11; ID8, n=5; ID8-KRAS, n=7; (D) OX40L: no
cancer, n=10; ID8, n=5; ID8-KRAS, n=8. Error bars represent the
means ± SEM. Statistical analysis was performed using the Student's
t-test. The P-value was adjusted using the Holm's method
(*P<0.05, **P<0.01). PBNs and PENs
indicate peripheral blood neutrophils and peritoneal neutrophils,
respectively. |
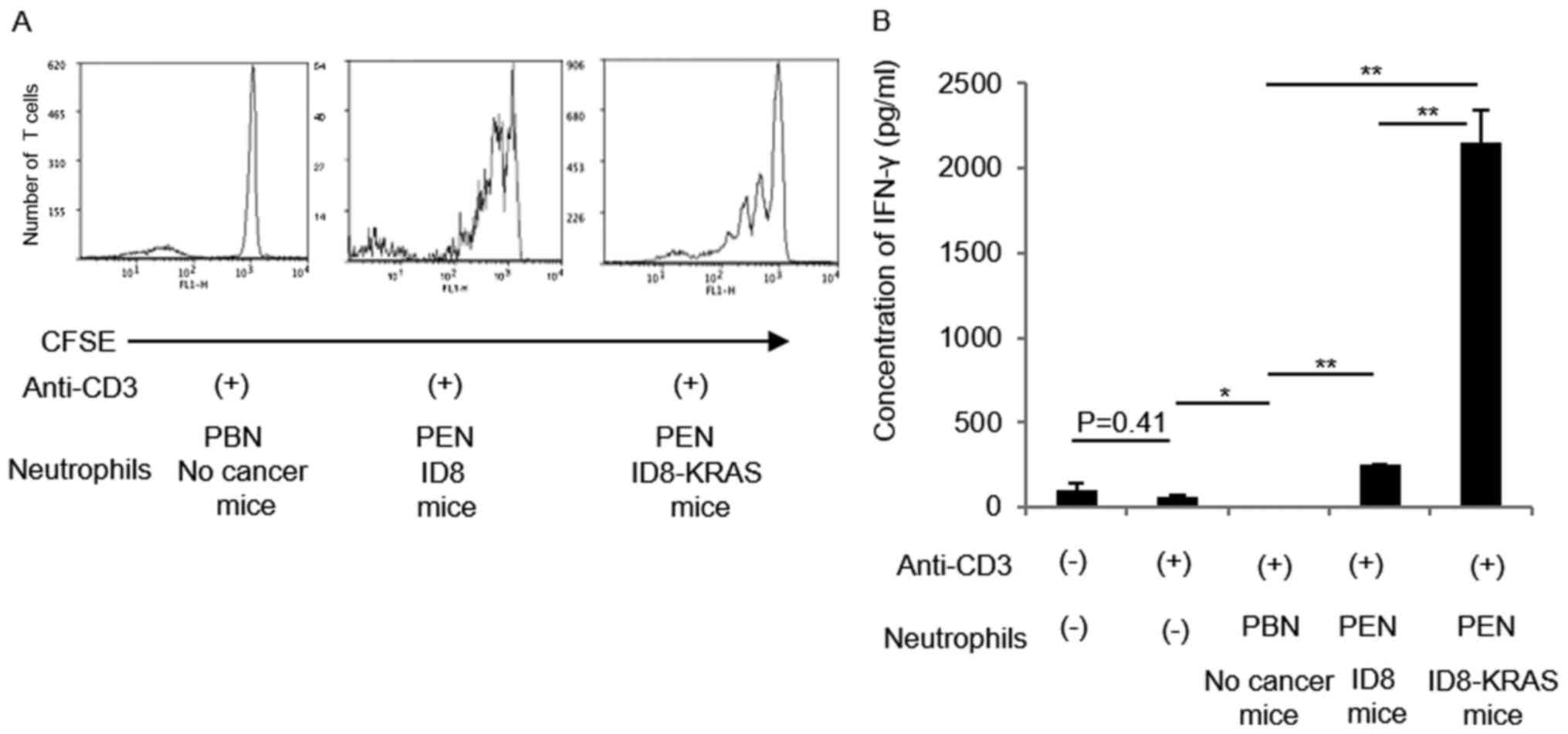
CD8+ T cell stimulatory
function on neutrophils is much stronger in ID8-KRAS mice than in
ID8 mice
ID8-KRAS-associated neutrophils possessed T cell
stimulatory function with higher expression of OX40L and 4-1BBL. We
next evaluated whether T cell stimulatory properties of neutrophils
were present in any cancer-related microenvironment. Therefore, we
compared T cell stimulatory effects between neutrophils from
ID8-induced ascites and those from ID8-KRAS-induced ascites. PENs
from ID8-induced ascites slightly activated CD8+ T cells
by inducing IFN-γ production, as well as stimulating
CD8+ T cell proliferation, which was much weaker than
that promoted by PENs from ID8-KRAS-induced ascites (Fig. 7).
A comparison of T cell costimulatory molecules
between ID8 and ID8-KRAS revealed that the OX40L and 4-1BBL
expression levels in PENs were significantly lower in ID8 mice than
in ID8-KRAS mice (Fig. 8).
Discussion
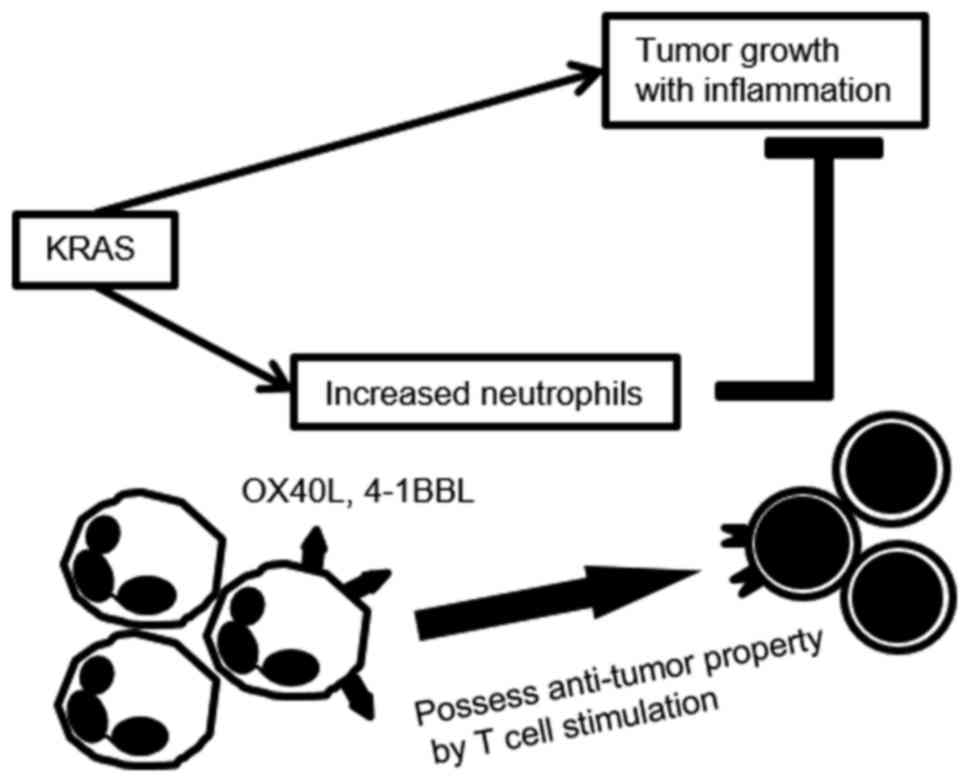
In the current study, we found that neutrophil
depletion in the ID8-KRAS model accelerated tumor formation and
aggravated IL-6-associated inflammation. Neutrophil depletion
increased the number of
Ly6C+Ly6Glow/CD11b+ M-MDSCs
accompanied with an increased number of Tregs in ascites.
Furthermore, the neutrophil depletion decreased the CD8+
T cell population. PENs from ID8-KRAS mice exhibited strong
potential to enhance T cell proliferation with a higher expression
of T cell costimulatory molecules compared with the PENs from ID8
mice.
A number of studies have reported that an elevated
neutrophil count or an increased N/L ratio in blood is a poor
prognostic marker in several cancer types (28-31).
The results of our previous study (8) also support the above finding that an
increased neutrophil count, which may induce an increased N/L
ratio, was observed in ID8-KRAS-induced rapid cancer progression.
However, in our model, neutrophil depletion appeared to accelerate
tumor formation. These findings suggest that the increase in TANs
in our model had antitumor effects.
The results of this study revealed that neutrophil
depletion increased the number of CD4+ T cells in
ascites, particularly the Tregs fraction. Furthermore, we revealed
that neutrophil depletion increased the M-MDSC population. MDSCs
are subdivided into two major groups: Polymorphonuclear MDSCs
(PMN-MDSCs) and M-MDSCs. PMN-MDSCs have a morphology similar to
that of granulocytes, and M-MDSCs are morphologically similar to
monocytes. In mice, PMN-MDSCs have a phenotype of
CD11b+Ly6C−Ly6Ghigh, whereas
M-MDSCs have a phenotype of
CD11b+Ly6G−Ly6Chigh (32,33).
In our model, the MDSCs increased by neutrophil depletion were
classified as M-MDSCs with the phenotype of
CD11b+Ly6G-Ly6Chigh. There is substantial
variability in the M-MDSC proportion depending on the type of
cancer. Patients with melanoma and prostate cancer have a
substantially higher proportion of M-MDSCs in the peripheral blood
than PMN-MDSCs (34). In ovarian
cancer, M-MDSCs have been reported to be present in both the
periphery and ascites. Ascite-derived IL-6 and IL-10 and their
downstream signal transducer and activator of transcription 3
(STAT3) signal are critically responsible for the accumulation and
suppressive activity of M-MDSCs (35). M-MDSCs potently suppress
nonspecific T cell responses, and on a per-cell basis, M-MDSCs have
a high suppressive activity with a higher production of nitric
oxide (NO), Arg1, TGF-β and immunosuppressive cytokines (36-40).
Furthermore, it has been previously demonstrated that TGF-β induced
Foxp3 gene expression in T cell receptor-challenged
CD4+CD25- peripheral naïve T cells, which were then
transformed toward a Treg phenotype with potent immunosuppressive
potential (41). In this model,
M-MDSCs may contribute to cancer progression by inducing Treg
differentiation and suppressing CD8 T cell activity.
The results of this study revealed that neutrophil
depletion decreased the CD8+ T cell population in
KRAS-induced ascites, suggesting that neutrophils promote
CD8+ T cell proliferation or survival in the
KRAS-induced environment. The finding that TANs in KRAS-induced
ascites had a strong potential for promoting the proliferation of
naïve CD8+ T cells in vitro supports the results
obtained in these in vivo assessments. In lung cancer, T
cell costimulatory molecules are responsible for T cell activation
by TANs (18). We also confirmed
that TANs in KRAS-induced ascites expressed OX40L and 4-1BBL at
higher levels than neutrophils from blood or TANs in ID8-induced
ascites. The T cell-stimulatory effects of TANs from KRAS-induced
ascites were reversed by chamber separation, suggesting that
cell-to-cell interactions are indispensable for T cell activation.
T cell costimulatory molecules on neutrophils may be responsible
for the involvement of TANs in KRAS-induced ascites in promoting T
cell proliferation.
Although the function of neutrophils in promoting T
cell activation has already been reported (39), the types of tumors that induce
neutrophils with antitumor properties have not yet been identified.
In this study, PENs from ID8-KRAS mice possessed markedly stronger
potential for T cell activation than PENs of ID8 mice. A difference
was also observed between the OX40L and 4-1BBL expression levels on
the neutrophil surfaces. Our results suggest that the KRAS-induced
environment generated neutrophils with portent antitumor
properties. To the best of our knowledge, no study to date has
demonstrated an association between oncogenes and the properties of
neutrophils. The cancer-specific modulation of neutrophils has not
yet been reported; however, the antitumor or protumor function of
neutrophils may be related to the activated oncogenes and their
specific TME. Furthermore, given that KRAS-related neutrophils
possessed antitumor properties with increased expression of OX40L
and 4-1BBL, OX40L- or 4-1BBL-based immunotherapy may be a
therapeutic strategy for KRAS mutated cancer.
In the current study, we used anti-Ly6G mAb to
deplete neutrophils. It is possible that
CD11b+Ly6C−Ly6Ghigh PMN-MDSCs were
also depleted in our model as it is difficult to distinguish
neutrophils from PMN-MDSCs. However, in our model, the depletion of
Ly6Ghigh populations resulted in an enhanced cancer formation
accompanied by the immunosuppressive microenvironment. Considering
that MDSCs have an immunosuppressive function, it appeared that, in
our model, a decreased number of neutrophils with immunostimulatory
effects potently affected the TME compared with a decreased number
of PMN-MDSCs with immunosuppressive effects. In addition, we only
used a mouse ovarian cancer model to elucidate the association
between the oncogene KRAS and the properties of neutrophils.
Further studies utilizing clinical samples are warranted for the
confirmation of the association identified in this study.
We herein propose that the increased number of
neutrophils in KRAS-induced ascites possessed antitumor properties
by modulating the TME, and that this effect was more potent in
KRAS-associated neutrophils. Knowledge regarding these
characteristics of neutrophil modification may provide a better
understanding of the cancer-induced microenvironment.
Acknowledgments
The authors would like to thank Ms. Chiho Kohno
(Division of Virology, National Cancer Center Research Institute)
for the construction of the ID8-KRAS cells.
Funding
This study was supported by Japan Society for the
Promotion of Science KAKENHI Grant number 16K11131 and
15H06172.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MY, AT, KKa, KK, TN and KO were involved in the
conception and design of the study. MY, AT, KKa, KK, MM, OWH and TK
were involved in the development of methodology. TK and OWH were
involved in the establishment of cell lines and technical support.
MY, AT, KKa and KA were involved in the acquisition of data. MY,
AT, KKa, JO, AK, HN, MS, AF, TI, KT, MM, TN, TA, KKo, OWH, KO, TK,
YO and TF were involved in the analysis and interpretation of data.
MY, AT, KKa and KA were involved in the writing, reviewing, and/or
revision of the manuscript. MY, AT, KKa, JO, KA and AK were
involved in administrative, technical, or material support. MY, AT,
KKa, MM, TN, TA, KKo, OWH, KO, TK, YO and TF supervised the
study.
Ethics approval and consent to
participate
The animal studies were approved by the University
of Tokyo Animal Committee. Our IACUC permitted this study and gave
this study the approval numbers P-14-027 and P15- 060.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Trellakis S, Bruderek K, Dumitru CA,
Gholaman H, Gu X, Bankfalvi A, Scherag A, Hütte J, Dominas N,
Lehnerdt GF, et al: Polymorphonuclear granulocytes in human head
and neck cancer: Enhanced inflammatory activity, modulation by
cancer cells and expansion in advanced disease. Int J Cancer.
129:2183–2193. 2011. View Article : Google Scholar
|
|
2
|
Jensen HK, Donskov F, Marcussen N,
Nordsmark M, Lundbeck F and von der Maase H: Presence of
intratumoral neutrophils is an independent prognostic factor in
localized renal cell carcinoma. J Clin Oncol. 27:4709–4717. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jensen TO, Schmidt H, Møller HJ, Donskov
F, Høyer M, Sjoegren P, Christensen IJ and Steiniche T:
Intratumoral neutrophils and plasmacytoid dendritic cells indicate
poor prognosis and are associated with pSTAT3 expression in AJCC
stage I/II melanoma. Cancer. 118:2476–2485. 2012. View Article : Google Scholar
|
|
4
|
Templeton AJ, McNamara MG, Šeruga B,
Vera-Badillo FE, Aneja P, Ocaña A, Leibowitz-Amit R, Sonpavde G,
Knox JJ, Tran B, et al: Prognostic role of neutrophil-to-lymphocyte
ratio in solid tumors: A systematic review and meta-analysis. J
Natl Cancer Inst. 106:dju1242014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Taguchi S, Nakagawa T, Matsumoto A, Nagase
Y, Kawai T, Tanaka Y, Yoshida K, Yamamoto S, Enomoto Y, Nose Y, et
al: Pretreatment neutrophil-to-lymphocyte ratio as an independent
predictor of survival in patients with metastatic urothelial
carcinoma: A multi-institutional study. Int J Urol. 22:638–643.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nakamura K, Nagasaka T, Nishida T, Haruma
T, Ogawa C, Kusumoto T, Seki N and Hiramatsu Y: Neutrophil to
lymphocyte ratio in the pre-treatment phase of final-line
chemotherapy predicts the outcome of patients with recurrent
ovarian cancer. Oncol Lett. 11:3975–3981. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Feng Z, Wen H, Bi R, Ju X, Chen X, Yang W
and Wu X: Preoperative neutrophil-to-lymphocyte ratio as a
predictive and prognostic factor for high-grade serous ovarian
cancer. PLoS One. 11:e01561012016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yoshida M, Taguchi A, Kawana K, Adachi K,
Kawata A, Ogishima J, Nakamura H, Fujimoto A, Sato M, Inoue T, et
al: Modification of the tumor microenvironment in KRAS or
c-MYC-induced ovarian cancer-associated peritonitis. PLoS One.
11:e01603302016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chandler RL, Damrauer JS, Raab JR,
Schisler JC, Wilkerson MD, Didion JP, Starmer J, Serber D, Yee D,
Xiong J, et al: Coexistent ARID1A-PIK3CA mutations promote ovarian
clear-cell tumorigenesis through pro-tumorigenic inflammatory
cytokine signalling. Nat Commun. 6:61182015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Eser S, Schnieke A, Schneider G and Saur
D: Oncogenic KRAS signalling in pancreatic cancer. Br J Cancer.
111:817–822. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pylayeva-Gupta Y, Grabocka E and Bar-Sagi
D: RAS oncogenes: Weaving a tumorigenic web. Nat Rev Cancer.
11:761–774. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Daniluk J, Liu Y, Deng D, Chu J, Huang H,
Gaiser S, Cruz-Monserrate Z, Wang H, Ji B and Logsdon CD: An NF-κB
pathway-mediated positive feedback loop amplifies Ras activity to
pathological levels in mice. J Clin Invest. 122:1519–1528. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Caruso RA, Bellocco R, Pagano M, Bertoli
G, Rigoli L and Inferrera C: Prognostic value of intratumoral
neutrophils in advanced gastric carcinoma in a high-risk area in
northern Italy. Mod Pathol. 15:831–837. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jablonska J, Leschner S, Westphal K,
Lienenklaus S and Weiss S: Neutrophils responsive to endogenous
IFN-beta regulate tumor angiogenesis and growth in a mouse tumor
model. J Clin Invest. 120:1151–1164. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fridlender ZG, Sun J, Kim S, Kapoor V,
Cheng G, Ling L, Worthen GS and Albelda SM: Polarization of
tumor-associated neutrophil phenotype by TGF-beta: 'N1' versus "N2"
TAN. Cancer Cell. 16:183–194. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li YW, Qiu SJ, Fan J, Zhou J, Gao Q, Xiao
YS and Xu YF: Intratumoral neutrophils: A poor prognostic factor
for hepatocellular carcinoma following resection. J Hepatol.
54:497–505. 2011. View Article : Google Scholar
|
|
17
|
Houghton AM: The paradox of
tumor-associated neutrophils: Fueling tumor growth with cytotoxic
substances. Cell Cycle. 9:1732–1737. 2010. View Article : Google Scholar
|
|
18
|
Eruslanov EB, Bhojnagarwala PS, Quatromoni
JG, Stephen TL, Ranganathan A, Deshpande C, Akimova T, Vachani A,
Litzky L, Hancock WW, et al: Tumor-associated neutrophils stimulate
T cell responses in early-stage human lung cancer. J Clin Invest.
124:5466–5480. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sasaki R, Narisawa-Saito M, Yugawa T,
Fujita M, Tashiro H, Katabuchi H and Kiyono T: Oncogenic
transformation of human ovarian surface epithelial cells with
defined cellular oncogenes. Carcinogenesis. 30:423–431. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Overmyer KA, Thonusin C, Qi NR, Burant CF
and Evans CR: Impact of anesthesia and euthanasia on metabolomics
of mammalian tissues: Studies in a C57BL/6J mouse model. PLoS One.
10:e01172322015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Arnold L, Tyagi RK, Mejia P, Van Rooijen
N, Pérignon JL and Druilhe P: Analysis of innate defences against
Plasmodium falciparum in immunodeficient mice. Malar J. 9:1972010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xiao H, Heeringa P, Liu Z, Huugen D, Hu P,
Maeda N, Falk RJ and Jennette JC: The role of neutrophils in the
induction of glomerulonephritis by anti-myeloperoxidase antibodies.
Am J Pathol. 167:39–45. 2005. View Article : Google Scholar
|
|
23
|
Tempfer C, Zeisler H, Sliutz G, Haeusler
G, Hanzal E and Kainz C: Serum evaluation of interleukin 6 in
ovarian cancer patients. Gynecol Oncol. 66:27–30. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Scambia G, Testa U, Benedetti Panici P,
Foti E, Martucci R, Gadducci A, Perillo A, Facchini V, Peschle C
and Mancuso S: Prognostic significance of interleukin 6 serum
levels in patients with ovarian cancer. Br J Cancer. 71:354–356.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang J, Yang L, Yu L, Wang YY, Chen R,
Qian J, Hong ZP and Su XS: Surgery-induced monocytic
myeloid-derived suppressor cells expand regulatory T cells in lung
cancer. Oncotarget. 8:17050–17058. 2017.PubMed/NCBI
|
|
26
|
O'Connor MA, Vella JL and Green WR:
Reciprocal relationship of T regulatory cells and monocytic
myeloid-derived suppressor cells in LP-BM5 murine
retrovirus-induced immunodeficiency. J Gen Virol. 97:509–522. 2016.
View Article : Google Scholar
|
|
27
|
Idorn M, Køllgaard T, Kongsted P, Sengeløv
L and Thor Straten P: Correlation between frequencies of blood
monocytic myeloid-derived suppressor cells, regulatory T cells and
negative prognostic markers in patients with castration-resistant
metastatic prostate cancer. Cancer Immunol Immunother.
63:1177–1187. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rao HL, Chen JW, Li M, Xiao YB, Fu J, Zeng
YX, Cai MY and Xie D: Increased intratumoral neutrophil in
colorectal carcinomas correlates closely with malignant phenotype
and predicts patients' adverse prognosis. PLoS One. 7:e308062012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamanaka T, Matsumoto S, Teramukai S,
Ishiwata R, Nagai Y and Fukushima M: The baseline ratio of
neutrophils to lymphocytes is associated with patient prognosis in
advanced gastric cancer. Oncology. 73:215–220. 2007. View Article : Google Scholar
|
|
30
|
Nuhn P, Vaghasia AM, Goyal J, Zhou XC,
Carducci MA, Eisenberger MA and Antonarakis ES: Association of
pretreatment neutrophil-to-lymphocyte ratio (NLR) and overall
survival (OS) in patients with metastatic castration-resistant
prostate cancer (mCRPC) treated with first-line docetaxel. BJU Int.
114(6b): E11–E17. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Azab B, Bhatt VR, Phookan J, Murukutla S,
Kohn N, Terjanian T and Widmann WD: Usefulness of the
neutrophil-to-lymphocyte ratio in predicting short- and long-term
mortality in breast cancer patients. Ann Surg Oncol. 19:217–224.
2012. View Article : Google Scholar
|
|
32
|
Movahedi K, Guilliams M, Van den Bossche
J, Van den Bergh R, Gysemans C, Beschin A, De Baetselier P and Van
Ginderachter JA: Identification of discrete tumor-induced
myeloid-derived suppressor cell subpopulations with distinct T
cell-suppressive activity. Blood. 111:4233–4244. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Youn JI, Nagaraj S, Collazo M and
Gabrilovich DI: Subsets of myeloid-derived suppressor cells in
tumor-bearing mice. J Immunol. 181:5791–5802. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Solito S, Marigo I, Pinton L, Damuzzo V,
Mandruzzato S and Bronte V: Myeloid-derived suppressor cell
heterogeneity in human cancers. Ann N Y Acad Sci. 1319:47–65. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen MF, Kuan FC, Yen TC, Lu MS, Lin PY,
Chung YH, Chen WC and Lee KD: IL-6-stimulated CD11b+
CD14+ HLA-DR− myeloid-derived suppressor
cells, are associated with progression and poor prognosis in
squamous cell carcinoma of the esophagus. Oncotarget. 5:8716–8728.
2014.PubMed/NCBI
|
|
36
|
Cuenca AG, Delano MJ, Kelly-Scumpia KM,
Moreno C, Scumpia PO, Laface DM, Heyworth PG, Efron PA and Moldawer
LL: A paradoxical role for myeloid-derived suppressor cells in
sepsis and trauma. Mol Med. 17:281–292. 2011. View Article : Google Scholar :
|
|
37
|
Dolcetti L, Peranzoni E, Ugel S, Marigo I,
Fernandez Gomez A, Mesa C, Geilich M, Winkels G, Traggiai E, Casati
A, et al: Hierarchy of immunosuppressive strength among
myeloid-derived suppressor cell subsets is determined by GM-CSF.
Eur J Immunol. 40:22–35. 2010. View Article : Google Scholar
|
|
38
|
Haverkamp JM, Smith AM, Weinlich R, Dillon
CP, Qualls JE, Neale G, Koss B, Kim Y, Bronte V, Herold MJ, et al:
Myeloid-derived suppressor activity is mediated by monocytic
lineages maintained by continuous inhibition of extrinsic and
intrinsic death pathways. Immunity. 41:947–959. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Umemura N, Saio M, Suwa T, Kitoh Y, Bai J,
Nonaka K, Ouyang GF, Okada M, Balazs M, Adany R, et al:
Tumor-infiltrating myeloid-derived suppressor cells are
pleiotropic-inflamed monocytes/macrophages that bear M1- and
M2-type characteristics. J Leukoc Biol. 83:1136–1144. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sumida K, Wakita D, Narita Y, Masuko K,
Terada S, Watanabe K, Satoh T, Kitamura H and Nishimura T:
Anti-IL-6 receptor mAb eliminates myeloid-derived suppressor cells
and inhibits tumor growth by enhancing T-cell responses. Eur J
Immunol. 42:2060–2072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen W, Jin W, Hardegen N, Lei KJ, Li L,
Marinos N, McGrady G and Wahl SM: Conversion of peripheral
CD4+CD25− naive T cells to
CD4+CD25+ regulatory T cells by TGF-beta
induction of transcription factor Foxp3. J Exp Med. 198:1875–1886.
2003. View Article : Google Scholar : PubMed/NCBI
|