Introduction
In South Korea, cancer has been a leading health
issue since the 1980s. Currently, lung cancer is the most commonly
diagnosed type of cancer in Korean males followed by gastric,
colorectal, liver and prostate cancer (1). In females, however, the 5 most
commonly diagnosed types of cancer are breast, thyroid, colorectal,
gastric and lung cancer. In 2015, cancer was the leading cause of
death in the Republic of Korea (2).
The World Health Organization (WHO) report cites
that approximately 80% of the population of developing countries in
Asia and Africa still use traditional herbs and plants as medicine
(3,4). Pharmaceutical companies now rely more
on experiments conducted on various traditionally known medicinal
plants to discover novel molecules and bioactive substances that
are more efficient against disease targets (5,6).
There has been an increase in the concern associated with the fatal
side-effects of some of the medicine available on the market and
this has led researchers to focus more on studying traditional
medicinal plants. Considering the fact that only 1% of >500,000
known plant species has been investigated, the need for novel
medicinal bioactive compounds is substantial (7,8).
Gastric cancer is the fourth most commonly diagnosed
type of cancer globally and the second most fatal in cancer
patients (9-11). The incidence rate of gastric cancer
is markedly high in Eastern Asia, Eastern Europe and South America
(12,13). At present, the treatment of gastric
cancer involves surgery, chemotherapy, radiotherapy, chemoradiation
and targeted therapy; however, the prognosis remains poor (14-17).
In addition, the effects of current chemotherapeutic drugs are not
particularly satisfactory and these drugs have various
side-effects. Therefore, it is important to search for novel agents
from natural sources, which have the ability to treat gastric
cancer (7,18-21).
Cancer is characterized by the loss of cell cycle
regulation followed by uncontrolled cell growth (22,23).
Cancer cells grow and constitute tumors in certain types of cancer.
After a period of time, cancer cells are able to leave the primary
site of establishment and invade other organs of the body. This
process is termed metastasis and is one of the reasons cancer is
difficult to cure. The control of the cell cycle by the induction
of cell death through the activation of cell cycle arrest or the
activation of apoptosis is the major goal of cancer experiments.
Various proteins regulate the cell cycle. The interaction between
cyclins and specific cyclin-dependent kinases (CDKs) allow the
cells to progress from one phase to another (24). The complexes which play a
significant role in G1/S cell phase transition are cyclin
D1,3/CDK4,6 and cyclin E/CDK2. These cyclin/CDK complexes are
activated and regulated by phosphorylation.
Apoptosis is an extraordinarily controlled cell
death mechanism through which cells are eliminated without inducing
inflammation around the dying cell (25,26).
Apoptotic cell death is triggered by the interaction between
ligands and membrane extrinsic death receptors or by the intrinsic
mitochondrial-mediated pathway (27,28).
While the extrinsic pathway is triggered by ligand-receptor
interaction, the intrinsic pathway is initiated by intracellular
processes, such as cell stress and leads to the release of
cytochrome c from the mitochondria. The cytochrome in turn
activates apoptosis through a number of cascade reactions in the
cell. The activation of downstream caspases results in the
formation of apoptotic bodies, DNA fragmentation and chromatin
condensation. Dying cells ultimately shrink and are removed by
phagocytic cells (29,30).
Aster incisus, also known as Kalimeris
incisa, is a flowering plant of the Asteraceae family. It is an
aromatic plant and mostly grows in many regions of the Northern
Hemisphere. Other plants belonging to the Asteraceae family have
been used for traditional medicinal purposes and most recently,
plants from the Aster genus were studied and reported for
their biological activities. Aster tataricus can reduce the
damage of neurons in epileptic rats (31), inhibit oxidative stress (32), regulate the expression of
inflammatory cytokines in THP-1 cells (33) and inhibit the proliferation of
SCC-9 human oral squamous carcinoma (34). Aster yomena can inhibit
adipogenesis in 3T3-L1 preadipocytes (35) and inflammation in RAW 264.7
macrophages (36). Aster
tripolium and Aster spathulifolius both have
anti-obesity effects (37,38).
In the current study, we evaluated the anticancer
effects, as well as the possible signaling pathways involved in the
cell death of human gastric adenocarcinoma AGS cells treated with
methanolic extract of Aster incisus referred to as AIE,
Aster incisus extract.
Materials and methods
Cell culture and cell viability
assay
HaCaT cells (normal human keratinocytes) were
obtained from AddexBio (San Diego, CA, USA) and A549 (lung
adenocarcinoma), Hep3B (hepatocellular carcinoma), MDA-MB-231
(breast adenocarcinoma) and AGS (gastric adenocarcinoma) cells were
purchased from ATCC (Manassas, VA, USA). The cells were maintained
in Dulbecco's modified Eagle's medium (DMEM; HyClone, Logan, UT,
USA) and Roswell Park Memorial Institute 1640 Medium (RPMI-1640;
HyClone) Both media were supplemented with 10% heat-inactivated
fetal bovine serum (FBS; HyClone) and 1% antibiotics (100 U/ml
penicillin and 10 µg/ml streptomycin; PAA Laboratories GmbH,
Pasching, Austria) and incubated at 37°C in 5% CO2 in a
humidified air atmosphere. For the cell viability assay, all the
cells were separately plated in 96-well plates at a final
concentration of 104 cells per well and were incubated
for 24 h. All the cells were later treated with various
concentrations (50, 100 or 150 µg/ml) of AIE and incubated
at 37°C for a further 24 h. AIE (voucher no. 016-001) was purchased
from Korean Plant Extract Bank (KPEB, Cheongju, Korea) with 99.9%
HPLC purity. Following treatment for 24 h, the medium was replaced
with fresh medium and 10 µl of WST-1® solution
were added to each well followed by 3 h of additional incubation at
37°C. Cell viability was determined by reading the absorbance using
an ELISA microplate reader (Molecular Devices, Silicon Valley, CA,
USA) at 460 nm and the percentages of inhibition were calculated.
Additionally, all cells were separately plated in a 96-well plate
and incubated at 37°C. Following 24 h of incubation, the cells were
incubated with 40 µM of the caspase inhibitor, z-VAD-fmk
(Sigma-Aldrich, St. Louis, MO, USA), for 1 h at 37°C. Following
incubation with z-VAD-fmk, the medium was removed and the cells
were treated with 150 µg/ml of AIE for 24 h. Following
treatment for 24 h, the medium was replaced with fresh medium and
10 µl of WST-1® solution were added to each well
followed by 3 h of additional incubation at 37°C. Cell viability
was determined by reading the absorbance using an ELISA microplate
reader (Molecular Devices) at 460 nm and the percentages of
inhibition were calculated.
4,6-Diamidino-2-phenylindole (DAPI)
staining
The HaCaT, A549, Hep3B, MDA-MB-231 and AGS cells
were plated on coverglass bottom dishes and incubated at 37°C for
24 h. The following day, the cells were challenged with 150
µg/ml of AIE and were incubated at 37°C again for an
additional 24 h. After the additional incubation, the cells were
washed once with phosphate-buffered saline (PBS) buffer (135 mM
sodium chloride, 2.7 mM potassium chloride, 4.3 mM sodium
phosphate, and 1.4 mM potassium dihydrogen phosphate) and stained
with 1 µg/ml of DAPI solution (Thermo Fisher Scientific
Inc., Waltham, MA, USA) diluted in methanol. Following incubation
in the dark at 37°C for 20 min, the dishes were rinsed with PBS and
fixed with 4% formaldehyde for 15 min at room temperature. The
fixed cells were mounted on the slide with Prolong Gold Antifade
Reagent and observed under a ZEISS LSM 710 confocal laser scanning
microscope (Carl Zeiss, Jena, Germany).
Western blot analysis
The AGS cells seeded in 100-mm dishes and treated
with the various concentrations (80, 100 or 140 µg/ml) were
harvested and lysed in ice-cold lysis buffer. Proteins were
quantified using CBB solution and were separated on a 12% SDS-PAGE
gel by electrophoresis. Following electrophoresis, the proteins
were transferred onto nitrocellulose membranes which were blocked
using PBST buffer (135 mM sodium chloride, 2.7 mM potassium
chloride, 4.3 mM sodium phosphate, 1.4 mM potassium dihydrogen
phosphate and 0.5% Tween-20) containing 5% skim milk powder. The
blots were probed overnight at 4°C with the following primary
antibodies: Cleaved caspase-8 (Cat. no. 9496), cleaved caspase-9
(Cat. no. 20750), caspase-3 (Cat. no. 9662), cleaved caspase-3
(Cat. no. 9661), FLIP (Cat. no. 8510), cleaved PARP (Cat. no.
5625), cytochrome c (Cat. no. 4272), Bcl-xL (Cat. no. 2762),
Bcl-2 (Cat. no. 2872), Bad (Cat. no. 9292), Bak (Cat. no. 3814),
Bid (Cat. no. 2002), AIF (Cat. no. 5318), cyclin D1 (Cat. no.
9932), cyclin D3 (Cat. no. 9932), cyclin E2 (Cat. no. 9870), CDK2
(Cat. no. 9932), CDK4 (Cat. no. 9932), CDK6 (Cat. no. 9932), p-p53
(Cat. no. 9919), p-Chk2 (Cat. no. 9931), p16 (Cat. no. 80772), p18
(Cat. no. 9932), p21 (Cat. no. 2947), p27 (Cat. no. 3686), E2F-1
(Cat. no. 3742), pRb (Cat. no. 9969), cdc25A (Cat. no. 3652) and
β-actin (Cat. no. 4970) obtained from Cell Signaling Technology
(CST; Danvers, MA, USA) and diluted according to the manufacturer's
instructions (1:1,000 in 1X PBS, 5% BSA and 0.1% Tween-20). The
blots were then washed 3 times in PBST, followed by incubation at
room temperature for 1 h with HRP conjugated anti-rabbit (Cat. no.
7074) or anti-mouse IgG secondary antibody (Cat. no. 7076) obtained
from CST and diluted according to the manufacturer's instructions
(1:1,000 in 1X PBS, 5% BSA and 0.1% Tween-20). The immunoblots were
then washed in PBST and visualized using ECL detection
solutions.
Immunofluorescence staining
The AGS cells were plated in coverglass bottom
dishes, incubated at 37°C and treated with 140 µg/ml of AIE
for 24 h. The cells were first stained with 1 µg/ml of a
solution of DAPI for 20 min at room temperature. After staining
with DAPI, the cells were then fixed with 4% formaldehyde for 15
min at room temperature and blocked for 1 h in a blocking solution,
including 5% rabbit and mouse normal sera (Santa Cruz Biotechnolgy
Inc., Santa Cruz, CA, USA) with 0.3% Triton X-100. The fixed and
blocked cells were incubated with the primary antibodies: β-actin
(CST, Cat. no. 4970) and cleaved caspase-3 (CST, Cat. no. 9661) for
3 h and washed 3 times with PBS buffer. After washing, the cells
were treated with 0.1 µg/ml of anti-mouse IgG (H+L), F(ab')2
fragment (Alexa Fluor® 555 Conjugate) (CST, Cat. no.
4409) and anti-rabbit IgG (H+L), F(ab')2 fragment (Alexa
Fluor® 488 Conjugate) (CST, Cat. no. 4412) for 1 h. The
stained cells were mounted on the slide with Prolong Gold Antifade
Reagent and observed in a ZEISS LSM 710 confocal laser scanning
microscope (Carl Zeiss).
Flow cytometric analysis
Following treatment with 80, 100 or 140 µg/ml
of AIE for 24 h, the cells were harvested by trypsinization and
fixed with 70% ethanol at 4°C overnight. Subsequently, the cells
were resuspended in PBS buffer containing 0.2 mg/ml RNase A and
incubated for 1 h at 37°C. The cells were stained with 40
µg/ml propidium iodide for 30 min at room temperature in the
dark. The distribution of sub-G1 DNA was analyzed using a BD
FACSVerse™ flow cytometer (BD Biosciences, San Jose, CA, USA).
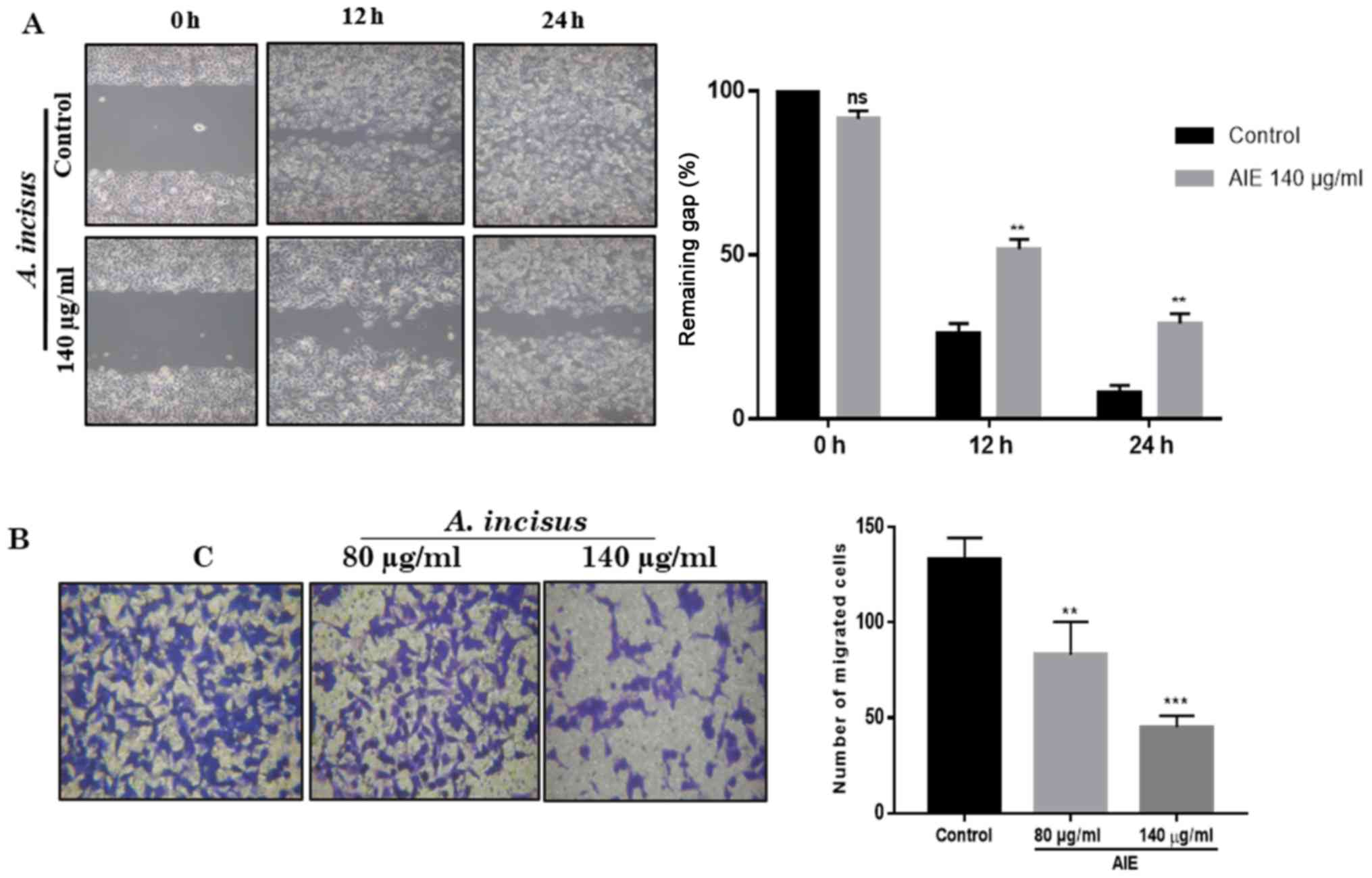
Wound healing assay
For the wound healing assay, the AGS cells were
seeded in a culture-Insert (ibidi, Planegg/Matrinsied, Germany) and
the cells were cultured until they reached 80-90% confluency. The
culture insert was removed and the cells were then washed with PBS
to remove the non-adherent cells. Fresh medium containing 140
µg/ml of AIE was provided to the cells prior to
photographing the plates with an Olympus CKX41 inverted microscope
(Olympus Corp., Tokyo, Japan) at 0, 12 and 24 h to capture two
different fields at each time point for each plate. The average
wound width was measured between the two lines representative of
cell migration determined by the mean of the furthest and the
nearest cells at the leading edge.
Cell invasion assay
The invasion of the AGS cells was investigated using
24-well plate Transwell inserts (8 µm pore size; Corning
Inc., New York, NY, USA). The inserts were first coated with 100
µl of Matrigel (BD Biosciences). RPMI-1640 medium containing
FBS in the lower compartment was used as a chemoattractant. During
and after treatment with AIE, the media in the inserts were
replaced with FBS-free media and the plate was incubated at 37°C in
5% CO2 for 24 h. Following incubation, the non-invading
cells were removed from the inner side of the inserts using a
cotton swab and the invading cells in the lower surface were
stained with 1% of crystal violet (Junsei Chemical Co., Tokyo,
Japan) for 20 min at 37°C. Images were taken using an Olympus CKX41
inverted microscope (Olympus Corp.).
Statistical analysis
Data are presented as the means ± standard deviation
(SD) for the indicated number of separate experiments. The mean of
the control was compared with the mean of each individual treatment
group using Tukey's multiple comparisons analysis following two-way
ANOVA by GraphPad Prism 7 software. (Graphpad Software Inc., La
Jolla, CA, USA), and statistical significance was set at
P<0.05.
Results
AIE suppresses the proliferation of human
cancer cell lines
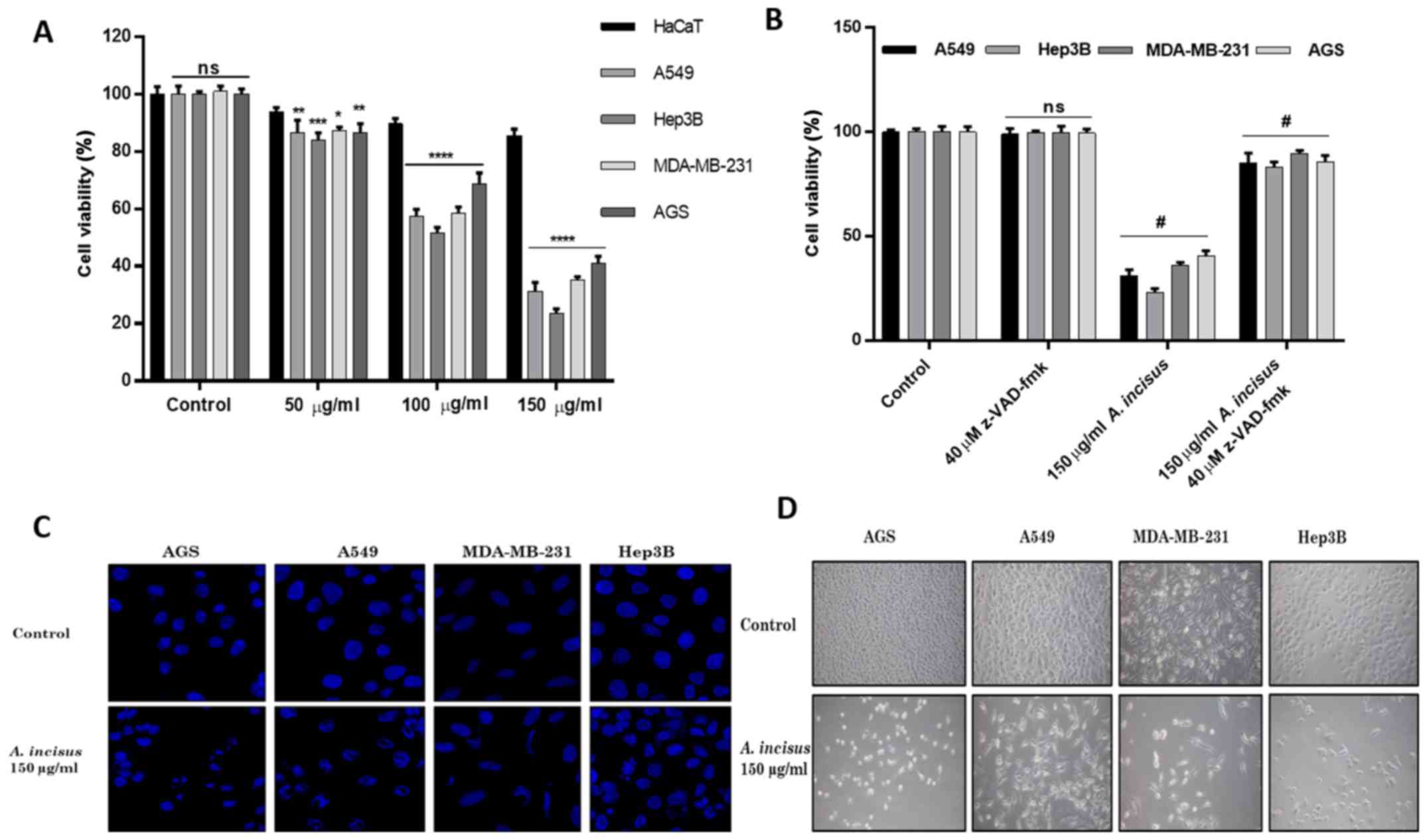
We examined the viability of the HaCaT, A549, Hep3B,
MDA-MB-231 and AGS cells treated with various concentrations of AIE
and found that AIE effectively suppressed the growth of both the
treated cancer cell lines, but did not affect the proliferation of
the non-cancerous keratinocytes, HaCaT cells, up to a concentration
of 150 µg/ml. These results determined that AIE was safe at
concentrations from 200 µg/ml and below. The treatment of
the human cancer cell lines with AIE demonstrated that 150
µg/ml was the most active concentration for the suppression
of cancer cell growth without affecting human non-cancerous cells.
In order to verify whether the effects of AIE were dose-dependent,
we therefore selected 3 different concentrations ≤150 µg/ml
for use in further experiments. The results revealed that AIE
gradually suppressed the viability of the cancer cells in a
dose-dependent manner (Fig. 1A).
Furthermore, the shrinkage of cells in all the treated cancer cells
was observed under an Olympus CKX41 inverted microscope and the
obtained images revealed that AIE significantly reduced the growth
of the AGS cells and affected their morphology (magnification,
×100) (Fig. 1D).
AIE induces apoptotic changes in treated
cancer cell lines
The A549, Hep3B, MDA-MB-231 and AGS cells were
incubated with the caspase inhibitor (z-VAD-fmk) followed by
treatment with 150 µg/ml of AIE. The results of cell
viability assay with WST1 revealed that treatment with the caspase
inhibitor significantly increased the proliferation of both the
cancerous and non-cancerous treated cells (Fig. 1B), demonstrating that the
caspase-related cell death pathway may play a major role in the
anti-proliferative effects of AIE. Additionally, changes in nuclear
morphology indicative of apoptosis were observed after the cancer
cells were treated with 150 µg/ml of AIE and stained with
DAPI solution. Images were obtained and analyzed under a ZEISS LSM
710 confocal laser scanning microscope (magnification, ×1,000). The
results revealed that the AIE-treated cells were characterized by
apoptotic morphological changes, such as the nuclear condensation
and the formation of apoptotic bodies, whereas the untreated cells
exhibited normal round-shaped nuclei (Fig. 1C).
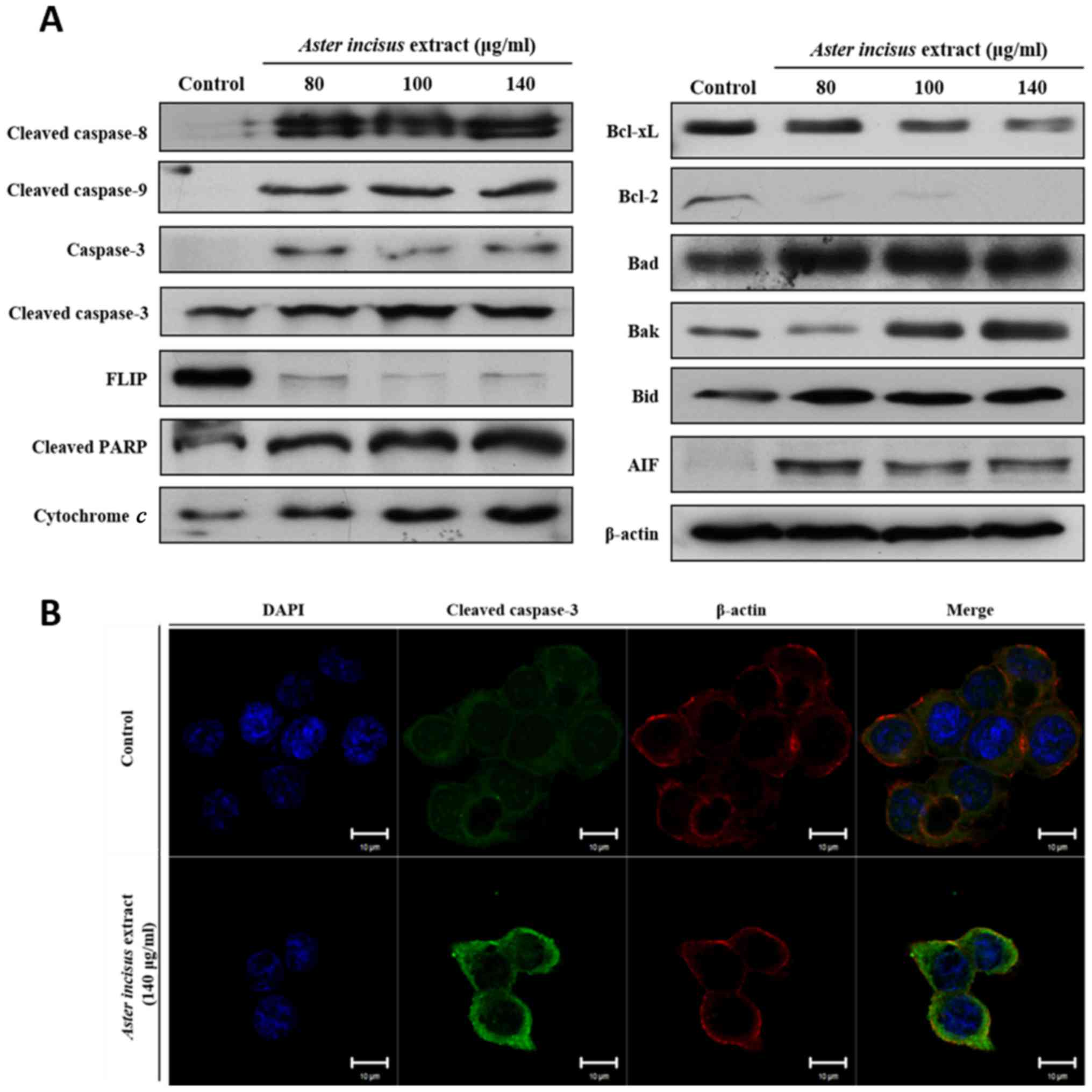
AIE induces the apoptosis of AGS gastric
cell lines
After the initial investigation of the 4 different
cancer cell lines, we then focused on AGS cell lines specifically.
Following the results mentioned above, we examined the expression
of apoptosis-related proteins by western blot analysis. The results
revealed that the expression of proteins from both the intrinsic
and extrinsic pathways varied in a dose-dependent manner. The
expression levels of pro-apoptotic proteins, such as Bak, Bid, Bad,
AIF, cytochrome c, cleaved caspase-8,-9 and -3, and cleaved
PARP were significantly upregulated, while the expression levels of
anti-apoptotic proteins, such as FLIP, Bcl-2 and Bcl-xL were lower
in the AIE-treated AGS cells when compared with the untreated
cells. These results confirmed that AIE induced the apoptosis of
the AGS cells (Fig. 2A). To
further confirm these results, we conducted an immunofluorescence
staining assay of the AGS cells using caspase-3 antibody. The
results revealed that caspase-3 protein was highly expressed in the
AGS cells treated with AIE when compared with the controls
(Fig. 2B). Taking all these
results into consideration, it can be concluded that AIE suppresses
the proliferation of the AGS gastric cell line by inducing
apoptosis.
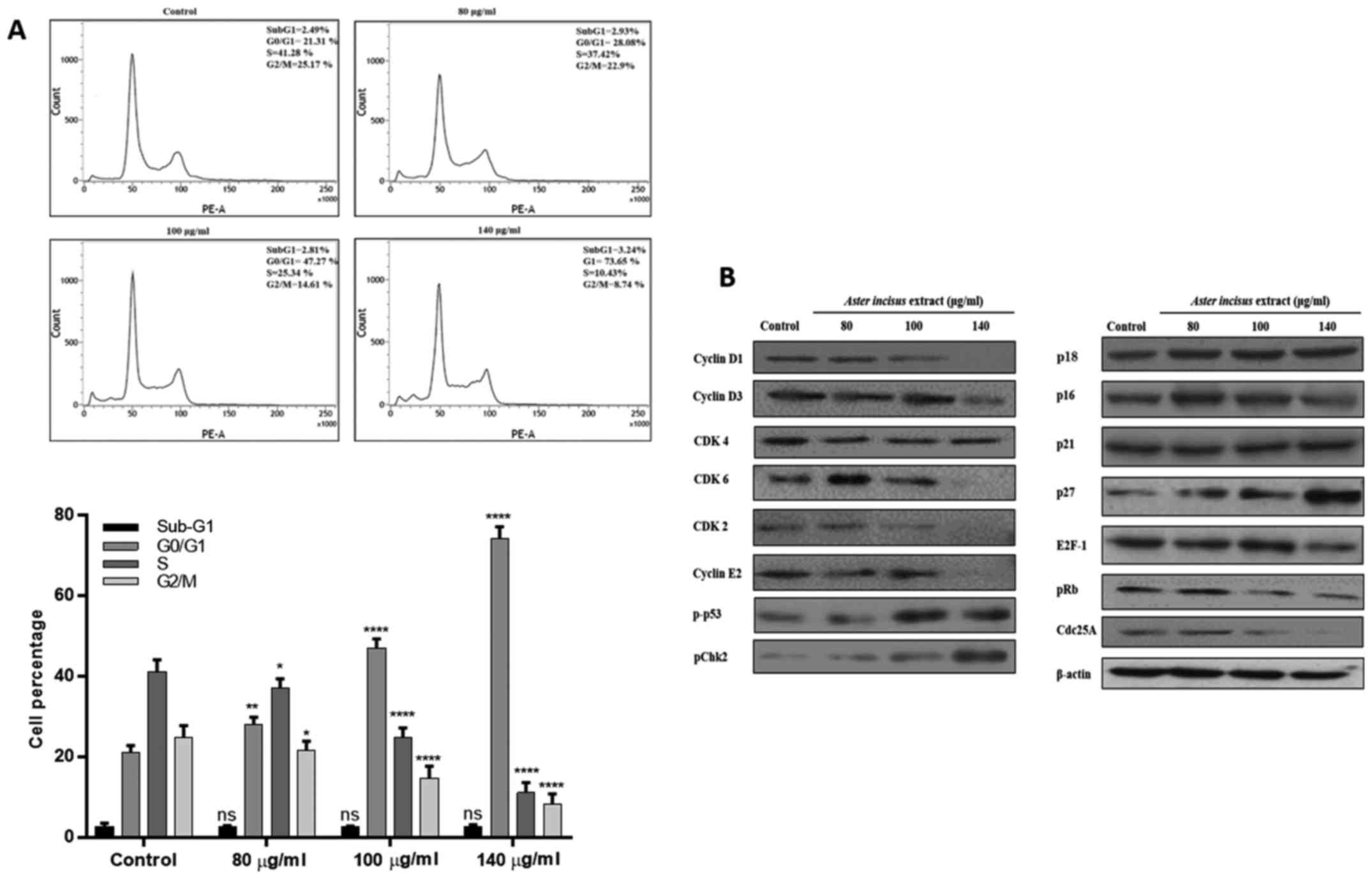
AIE induces G1/S phase cell cycle arrest
in the AGS gastric cancer cell line
We investigated the effects of AIE on cell cycle
phases by using PI staining in FACS analysis. This assay allowed us
to quantify the percentage of cells in the different phases of the
cell cycle. For this analysis, the AGS cells were treated with 80,
100, 140 µg/ml of AIE for 24 h and later stained with PI.
The results of flow cytometric analysis revealed the accumulation
of cells in the G0/G1 phase in a dose-dependent manner. The number
of cells in the G0/G1 phase were 21.3% in the untreated cells, and
28.08, 47.2 and 73.6% in the cells treated with AIE at 80, 100 and
140 µg/ml, respectively (Fig.
3A). To validate these results, we conducted a western blot
analysis of proteins that are involved in the G1/S phase,
particularly cyclins, CDKs and CDK inhibitors. The results revealed
that AIE downregulated the expression levels of proteins involved
in the G1/S phase progression, including cyclin D1, cyclin D3,
cyclin E2, CDK2, CDK4 and CDK6, while it increased the expression
levels of CDK-related inhibitors, such as p16, p18, p21 and p27
(Fig. 3B). Taking these results
into consideration, it can be concluded that AIE successfully
induced cell cycle arrest in the treated AGS cells, leading to the
accumulation of cells in the G1/S phase (please also see Fig. 5).
AIE suppresses the migration and invasion
of AGS cells
To explore the effects of AIE on cell migration, a
wound healing assay was conducted. For the wound healing assay,
images were acquired after 24 h of treatment using an inverted
microscope to record the progression of cells from the beginning of
the treatment up to 24 h. The results shown in Fig. 4A indicatd that AIE was suppressed
the migration of the treated AGS cells. The treated AGS cells were
not able to fill the gap between the cells, while the untreated
cells filled the surface within 24 h. These results confirm that
AIE suppresses the migration of AGS gastric cancer cells.
From the invasion assay, we observed that AIE was
able to successfully prevent the invasion of AGS cells through the
Matrigel layer. These results suggest that AIE has the ability to
inhibit the degradation of extracellular Matrigel by proteases from
the cancer cells, and thus, prevent the spread of cancer cells to
other organs (Fig. 4B).
Discussion
AIE belongs to the Asteraceae family and has been
used for medicinal purposes in South Korea. However, to date, and
at least to the best of our knowledge, there is no report on the
use of AIE for the treatment of cancer cells. In this study, we
investigated the potential anticancer effects of AIE in 4 different
cancer cell lines, A549, Hep3B, MDA-MB-231 and AGS cells. The
results suggested that AIE reduced cell viability in a
dose-dependent manner (Fig.
1A).
Apoptosis involves well-defined morphological
changes, resulting from the activation of specific signaling
pathways. Apoptotic cells exhibit DNA fragmentation, nuclear
condensation and the blebbing of the plasma membrane, leading to
the release of small membrane-enclosed particles, known as
apoptotic bodies (39,40). In this study, we found that AIE was
able to initiate nuclear condensation in both the cancer cell lines
when treated with 150 µg/ml (Fig. 1D). The results of WST-1 assay for
the cells challenged with the caspase inhibitor before treatment
clearly demonstrated that the inhibitor significantly increased the
survival of AGS gastric cancer cells that were treated with AIE
(Fig. 1B)
The two major most common pathways involved in
apoptosis are the extrinsic and the intrinsic pathways. The first
pathway (extrinsic) is initiated through the interaction between
cellular death receptors and their respective ligands. Following
the interaction, intracellular caspase-8 is recruited by the
intracellular part of the death receptor and is activated.
Activated caspase-8 then initiates apoptosis through the cleavage
of effector caspases or cleavage of Bid into tBid (41). The results of this study revealed
that AIE increased the expression of cleaved caspase-8 and Bid in
the AGS cells (Fig. 2A).
The second pathway (intrinsic pathway) is initiated
by different stimuli that do not require a transmembrane receptor,
such as cellular stresses, in which Bcl-2 family proteins play a
crucial role. Bcl-2 family proteins are a large group of proteins
that include anti-apoptotic proteins (Bcl-2, Bcl-xL, Mcl-1 and
Bcl-w), pro-apoptotic proteins (Bax and Bak), and finally the
BH3-only proteins (Bad and Bid). BH3-only proteins monitor cellular
processes and transduce both extrinsic and intrinsic death signals
to the Bcl-2 family of proteins at the outer mitochondrial membrane
(OMM) to modulate mitochondrial outer membrane permeability (MOMP).
The increase in MOMP leads to the release of cytochrome c,
AIF and EndoG in the cytosol. Cytochrome c and the Apaf-1
complex cleave procaspase-9 and cleaved caspase-9 cleaves
procaspase-3 to activate caspase-3 (42). AIF and EndoG translocate to the
nucleus where they participate in DNA fragmentation (43). A number of in vitro studies
have confirmed that the induction of apoptosis leads to the death
of cancer cells (44-46).
In this study, we observed the morphological changes
and nuclear condensation in the cells. The results revealed that
AIE induced mitochondrial dysfunction via the increased expression
of Bak and pro-apoptotic proteins, and by the decreased expression
of Bcl-2 and Bcl-xL, anti-apoptotic proteins, in the treated AGS
cells specifically in a dose-dependent manner (Fig. 2A). The results of western blot
analysis and immunofluorescence staining of caspase-3 as shown in
these figures, clearly indicate a significant increase in the
levels of pro-apoptotic proteins and the inhibition of
anti-apoptotic proteins in the treated cells when compared with the
untreated cells. The results were corroborated with the results of
immunofluorescence staining for caspase-3 expression in the treated
AGS cells (Fig. 2B).
The cell cycle is the most biologically conserved
process throughout the years. The cell cycle is regulated by
numerous proteins and variations in the cell regulation usually
result in cancer. Cancer development can be due to various causes,
including a dysregulation in cellular functions. The human cell
cycle is divided into 4 phases: The G1 phase, S phase, G2 phase and
M phase. Cyclins and CDKs are crucial for cell division; therefore,
the expression and regulation of these proteins play a crucial role
in the initiation of cancer or in cancer treatment. Each cell cycle
phase is regulated by specific cyclins and CDKs. The interaction
between cyclins and CDKs allow the cells to progress with their
division. Genetic alterations in cells, such as a mutation can lead
to the loss of cell regulation. Components of the cell cycle
machinery are frequently altered in human cancer. A group of
inhibitory proteins, cyclin-dependent kinases inhibitors or CDK
inhibitors, control the cyclin-CDK activity thereby restraining
cell cycle progression in response to extracellular and
intracellular signals (29). The
crucial role of CDKs has prompted great interest in the development
of specific kinase inhibitors that could be expected to block cell
cycle progression and induce growth arrest. CDK inhibitors are
divided into two major groups: The CDK4 inhibitors named due to
their ability to inhibit CDK4 and CDK6 by competing with cyclin D,
include p16 and p18. The second family is Cip/Kip family (for CDK
interacting protein/kinase inhibitory protein) which include p21
and p27 (47). As other studies
have reported previously, the activation of cyclin-dependent
kinases inhibitors is a very effective method with which to inhibit
the proliferation of cancer cells (48,49).
The G1 phase is a very important part of the cell cycle as it
provides the signal to the cell to allow it to enter the cell
division stage.
The cyclin D-CDK4/6 complex initiates the
phosphorylation of Rb during G1 to S phase transition, which leads
to the release of E2F-1 by Rb allowing the initiation of
transcription and translation. Later in the G1 phase, the complex
cyclin A-CDK2 and the cyclin E-CDK2 complex in the early S phase
regulate the transition from the G1 phase to S phase.
Cyclin-dependent kinases inhibitors, by interfering in the
interaction of cyclins with CDKs, trigger the arrest of cell cycle
in G1 phase. Many plants used as traditional medicine have been
reported to arrest the cell cycle of various cancer cells lines
during the G1/S phase transition (50-53).
One of the other most important tumor suppressors is
tumor protein p53, which plays a key role in the cellular response
to DNA damage. Similar to cyclin-CDK complexes, p53 is very
important in the transition of cells during division from the G1 to
S phase. p53 also arrests the cells in the G1 phase after DNA
damage by activating the cyclin-dependent kinases inhibitors
(54-56).
Chk2 protein is also phosphorylated following DNA
damage, which in turn inactivates the cell division cycle 25A
phosphatase (Cdc25A). The deactivation of Cdc25A results in the
elevation of the phosphorylated (inactive) form of Cdk2; therefore,
cells cannot progress into the S phase and replicate their DNA
(57,58).
The results from the flow cytometric analysis in the
current study demonstrated the accumulation of treated AGS cells in
the G1 phase. Additional results from western blot analysis
revealed that AIE reduced the expression of cyclins and CDKs
involved in the G1 phase (more specifically, cyclin D1, cyclin D3,
cyclin E, CDK2 CDK4 and CDK6), but increased the expression of the
cyclin-dependent kinases inhibitors, p16, p18, p21 and p27
(Fig. 3B).
To deepen our understanding, we analyzed the effects
of AIE on cell migration and cell invasion. Migration and invasion
are very important steps in metastasis. In cancer patients, some
cancer cells can spread from the initial site to secondary sites.
This process is known as metastasis and is made possible by the
ability to migrate, degrade the extracellular matrix and spread to
other parts of the body through the bloodstream. Once cancer cells
start to diffuse, the prognosis for the patient worsens and reduces
the chances of survival considerably. The results of this study
demonstrated that AIE successfully suppressed the migration of the
AIE-treated AGS gastric cancer cell lines in a wound healing assay.
AGS cells plated in inserts coated with Matrigel when treated with
AIE were not able to migrate. These results revealed that AIE
considerably reduced the ability of AGS gastric cancer cells to
invade tissues by degrading the extracellular matrix components. We
thus concluded that AIE has the ability to inhibit the migration of
AGS cells and their invasion (Fig.
4).
In conclusion, the findings of this study
demonstrate that AIE induces the apoptosis of AGS gastric cancer
cells via both the intrinsic mitochondrial-mediated cell death
pathway and the caspase-dependent pathway via the activation of
caspases. AIE also significantly induced cell cycle arrest in G1/S
phase and furthermore, AIE suppressed the migration and invasion of
AGS cells. The results of this study, at least to the best of our
knowledge, are the first report of any biological activity of
Aster incisus. Further studies are warranted for the
isolation and the identification of the active single compounds
responsible for the anti-proliferative activities of AIE in gastric
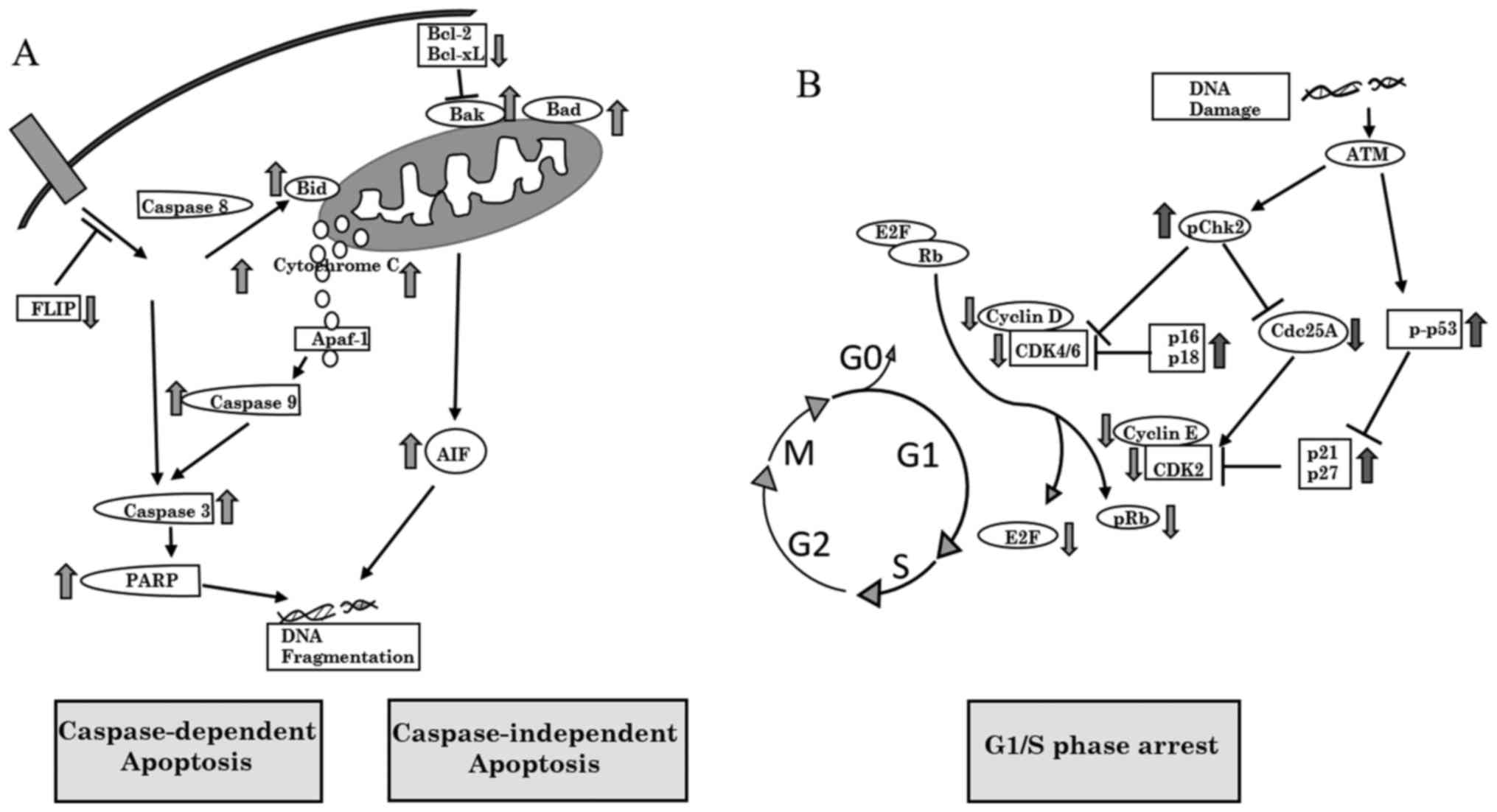
cancer cells. A schematic representation of the pathways through
which AIE induces apoptosis and cell cycle arrest in AGS cells is
shown in Fig. 5.
Funding
This study was supported by the National Research
Foundation of Korea (NRF) grant funded by the Korea government
(MSIT)(NRF-2016R1A2B4014909).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DN conducted the experiments and wrote the
manuscript; YAS provided the plant extract and advice in the
conception and design of the study; YBS and GDK conducted parts of
the experiments, MPP and IN contributed to the analysis of the
data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Authors would like to express their deep gratitude
to Courtney T. Green for her support during the editing of the
manuscript.
References
|
1
|
Oh CM, Won YJ, Jung KW, Kong HJ, Cho H,
Lee JK, Lee DH and Lee KH; Community of Population-Based Regional
Cancer Registries: Cancer statistics in Korea: Incidence,
mortality, survival, and prevalence in 2013. Cancer Res Treat.
48:436–450. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jung KW, Won YJ, Kong HJ and Lee ES;
Community of Population-Based Regional Cancer Registries: Cancer
Statistics in Korea: Incidence, mortality, survival, and prevalence
in 2015. Cancer Res Treat. 50:303–316. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
World Health Organization: 2013, WHO
traditional medicine strategy. pp. 2014–2023, http://www.who.int/medicines/publications/traditional/trm_strategy14_23/en/.
Accessed April 28, 2018.
|
|
4
|
Ma JK-C, Chikwamba R, Sparrow P, Fischer
R, Mahoney R and Twyman RM: Plant-derived pharmaceuticals - the
road forward. Trends Plant Sci. 10:580–585. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Padmaja R, Arun PC, Prashanth D, Deepak M,
Amit A and Anjana M: Brine shrimp lethality bioassay of selected
Indian medicinal plants. Fitoterapia. 73:508–510. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bais S: A phytopharmacological review on
an important medicinal plant: Leea indica. Cancer. 15:162013.
|
|
7
|
Cragg GM and Newman DJ: Plants as a source
of anti-cancer agents. J Ethnopharmacol. 100:72–79. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mann J: Natural products in cancer
chemotherapy: Past, present and future. Nat Rev Cancer. 2:143–148.
2002. View
Article : Google Scholar
|
|
9
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar
|
|
12
|
Fock KM: Review article: The epidemiology
and prevention of gastric cancer. Aliment Pharmacol Ther.
40:250–260. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Satolli MA, Buffoni L, Spadi R and Roato
I: Gastric cancer: The times they are a-changin. World J
Gastrointest Oncol. 7:303–316. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yoong J, Michael M and Leong T: Targeted
therapies for gastric cancer: Current status. Drugs. 71:1367–1384.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cravo M, Fidalgo C, Garrido R, Rodrigues
T, Luz G, Palmela C, Santos M, Lopes F and Maio R: Towards curative
therapy in gastric cancer: Faraway, so close! . World J
Gastroenterol. 21:11609–11620. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Davidson M, Okines AF and Starling N:
Current and future therapies for advanced gastric cancer. Clin
Colorectal Cancer. 14:239–250. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee JH, Kim KM, Cheong JH and Noh SH:
Current management and future strategies of gastric cancer. Yonsei
Med J. 53:248–257. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cragg GM, Grothaus PG and Newman DJ:
Impact of natural products on developing new anti-cancer agents.
Chem Rev. 109:3012–3043. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yin SY, Wei WC, Jian FY and Yang NS:
Therapeutic applications of herbal medicines for cancer patients.
Evid Based Complement Alternat Med. 2013:3024262013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bishayee A: Editorial: Current advances in
cancer prevention and treatment by natural products. Curr Pharm
Biotechnol. 13:115–116. 2012. View Article : Google Scholar
|
|
21
|
Amin AR, Kucuk O, Khuri FR and Shin DM:
Perspectives for cancer prevention with natural compounds. J Clin
Oncol. 27:2712–2725. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nakanishi M, Shimada M and Niida H:
Genetic instability in cancer cells by impaired cell cycle
checkpoints. Cancer Sci. 97:984–989. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jin Z and El-Deiry WS: Overview of cell
death signaling pathways. Cancer Biol Ther. 4:139–163. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Morgan DO: Principles of CDK regulation.
Nature. 374:131–134. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hassan M, Watari H, AbuAlmaaty A, Ohba Y
and Sakuragi N: Apoptosis and molecular targeting therapy in
cancer. BioMed Res Int. 2014:1508452014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cory S and Adams JM: The Bcl2 family:
Regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Langerak P and Russell P: Regulatory
networks integrating cell cycle control with DNA damage checkpoints
and double-strand break repair. Philos Trans R Soc Lond B Biol Sci.
366:3562–3571. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Al-Ejeh F, Kumar R, Wiegmans A, Lakhani
SR, Brown MP and Khanna KK: Harnessing the complexity of DNA-damage
response pathways to improve cancer treatment outcomes. Oncogene.
29:6085–6098. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Branzei D and Foiani M: Regulation of DNA
repair throughout the cell cycle. Nat Rev Mol Cell Biol. 9:297–308.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jackson SP and Bartek J: The DNA-damage
response in human biology and disease. Nature. 461:1071–1078. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu B, Wei-chen Ho, Li X, Ma L, Bai X, Liu
Q and Liu B: Aster tataricus attenuates the neuronal cell damage
and restores the cognitive functions in epileptic rat. Biomed Res
(Aligarh). 28:1189–1194. 2017.
|
|
32
|
Yao X, Dong X, Zhang H-S, Wang Y and
Lan-Yuzhu X-SL: Preventive effect of Aster tataricus on oxidative
stress and biomarker of renal function in rat fed with high fat
diet and sucrose diet. Biomed Res (Aligarh). 28:1647–1653.
2017.
|
|
33
|
Zhang HT, Tian M, He QW, Chi N, Xiu CM and
Wang YB: Effect of Aster tataricus on production of inflammatory
mediators in LPS stimulated rat astrocytoma cell line (C6) and
THP-1 cells. Saudi Pharm J. 25:370–375. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang R, Xiao S and Niu Z: Anti-cancer
activity of Aster tataricus on SCC-9 human oral squamous carcinoma.
Afr J Tradit Complement Altern Med. 14:142–147. 2017. View Article : Google Scholar :
|
|
35
|
Han MH, Jeong JS, Jeong JW, Choi SH, Kim
SO, Hong SH, Park C, Kim BW and Choi YH: Ethanol extracts of Aster
yomena (Kitam) Honda inhibit adipogenesis through the activation of
the AMPK signaling pathway in 3T3-L1 preadipocytes. Drug Discov
Ther. 11:281–287. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Choi JH, Kim DW, Park SE, Choi BS, Sapkota
K, Kim S and Kim SJ: Novel thrombolytic protease from edible and
medicinal plant Aster yomena (Kitam) Honda with anticoagulant
activity: Purification and partial characterization. J Biosci
Bioeng. 118:372–377. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Duarte B, Cabrita MT, Gameiro C, Matos AR,
Godinho R, Marques JC and Caçador I: Disentangling the
photochemical salinity tolerance in Aster tripolium L.: Connecting
biophysical traits with changes in fatty acid composition. Plant
Biol Stuttg. 19:239–248. 2017. View Article : Google Scholar
|
|
38
|
Kim SJ, Bang CY, Guo YR and Choung SY:
Anti-obesity effects of Aster spathulifolius extract in high-fat
diet-induced obese rats. J Med Food. 19:353–364. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kantari C and Walczak H: Caspase-8 and
bid: Caught in the act between death receptors and mitochondria.
Biochim Biophys Acta. 1813:558–563. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Brunelle JK and Letai A: Control of
mitochondrial apoptosis by the Bcl-2 family. J Cell Sci.
122:437–441. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Parrish AB, Freel CD and Kornbluth S:
Cellular mechanisms controlling caspase activation and function.
Cold Spring Harb Perspect Biol. 5:a0086722013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Redza-Dutordoir M and Averill-Bates DA:
Activation of apoptosis signalling pathways by reactive oxygen
species. Biochim Biophys Acta. 1863:2977–2992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kitazumi I and Tsukahara M: Regulation of
DNA fragmentation: The role of caspases and phosphorylation. FEBS
J. 278:427–441. 2011. View Article : Google Scholar
|
|
44
|
Cazal CM, Choosang K, Severino VG, Soares
MS, Sarria AL, Fernandes JB, Silva MF, Vieira PC, Pakkong P,
Almeida GM, et al: Evaluation of effect of triterpenes and
limonoids on cell growth, cell cycle and apoptosis in human tumor
cell line. Anticancer Agents Med Chem. 10:769–776. 2010. View Article : Google Scholar
|
|
45
|
Kastan MB and Bartek J: Cell-cycle
checkpoints and cancer. Nature. 432:316–323. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: A changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hoshino R, Tanimura S, Watanabe K, Kataoka
T and Kohno M: Blockade of the extracellular signal-regulated
kinase pathway induces marked G1 cell cycle arrest and apoptosis in
tumor cells in which the pathway is constitutively activated:
Up-regulation of p27(Kip1). J Biol Chem. 276:2686–2692. 2001.
View Article : Google Scholar
|
|
48
|
Chen ZY and Istfan NW: Docosahexaenoic
acid, a major constituent of fish oil diets, prevents activation of
cyclin-dependent kinases and S-phase entry by serum stimulation in
HT-29 cells. Prostaglandins Leukot Essent Fatty Acids. 64:67–73.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Harbour JW and Dean DC: Rb function in
cell-cycle regulation and apoptosis. Nat Cell Biol. 2:E65–E67.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Agami R and Bernards R: Distinct
initiation and maintenance mechanisms cooperate to induce G1 cell
cycle arrest in response to DNA damage. Cell. 102:55–66. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Petronelli A, Pannitteri G and Testa U:
Triterpenoids as new promising anticancer drugs. Anticancer Drugs.
20:880–892. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang Y, Zhou Y, Zhou H, Jia G, Liu J, Han
B, Cheng Z, Jiang H, Pan S and Sun B: Pristimerin causes G1 arrest,
induces apoptosis, and enhances the chemosensitivity to gemcitabine
in pancreatic cancer cells. PLoS One. 7:e438262012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wu G, Qian Z, Guo J, Hu D, Bao J, Xie J,
Xu W, Lu J, Chen X and Wang Y: Ganoderma lucidum extract induces G1
cell cycle arrest, and apoptosis in human breast cancer cells. Am J
Chin Med. 40:631–642. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Il Jung H, Jo MJ, Kim HR, Choi YH and Kim
GD: Extract of Saccharina japonica induces apoptosis companied by
cell cycle arrest and endoplasmic reticulum stress in SK-Hep1 human
hepatocellular carcinoma cells. Asian Pac J Cancer Prev.
15:2993–2999. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Banin S, Moyal L, Shieh S, Taya Y,
Anderson CW, Chessa L, Smorodinsky NI, Prives C, Reiss Y, Shiloh Y,
et al: Enhanced phosphorylation of p53 by ATM in response to DNA
damage. Science. 281:1674–1677. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Reinhardt HC and Schumacher B: The p53
network: Cellular and systemic DNA damage responses in aging and
cancer. Trends Genet. 28:128–136. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Qiu P, Guan H, Dong P, Li S, Ho CT, Pan
MH, McClements DJ and Xiao H: The p53-, Bax- and p21-dependent
inhibition of colon cancer cell growth by 5-hydroxy
polymethoxyflavones. Mol Nutr Food Res. 55:613–622. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Bartek J and Lukas J: Chk1 and Chk2
kinases in checkpoint control and cancer. Cancer Cell. 3:421–429.
2003. View Article : Google Scholar : PubMed/NCBI
|