Introduction
Lymphatic malformations are non-malignant masses
that result from the abnormal development of the lymphatic vessels
(1,2). Cystic lymphatic malformations (cLM)
comprise a significant portion of congenital lymphatic malformation
and can be classified into three histological subtypes based on the
diameter of the cyst, including macrocystic (>1 cm), microcystic
(<1 cm) and mixed-type (3).
While the exact etiology is unknown, cLM is believed to result from
the loss of connection between local lymphatic tissues and the
lymphatic or vascular systems during critical development stages of
the lymphatic system (4,5). While the genetics and molecular
mechanisms underlying hemangiomas and vascular malformations have
been elucidated (6-8), those of cLM remain unknown at this
time.
Hypoxia-inducible factor-1 (HIF-1) is a protein
complex that consists of two similar subunits, HIF-1α and HIF-1β.
Biological roles of HIF-1 include mediation of hypoxia on cells and
regulation of the metabolism (9,10).
The HIF-1 cascade occurs when HIF-1α binds to HIF-1β in the cell
nucleus under hypoxic conditions. Several researchers are currently
investigating the role of HIF-1α in lymphangiogenesis and
angiogenesis during tumor development (11,12).
Unlike blood vessels, lymphatic vessels are exposed to an
oxygen-deprived environment due to the lack of oxygen-carrying red
blood cells. In hypoxic milieus, tumor cells enhance the growth of
lymphatic vessels by increasing the production of vascular
endothelial growth factor (VEGF)-C, which provides a route for
tumor cells metastasis (13).
VEGF receptor 3 (VEGFR-3), which is primarily
localized to lymphatic endothelial cells, is the tyrosine kinase
receptor of VEGF-C and its signaling pathway has been demonstrated
to associated with lymphatic vascular development during multiple
stages (14).
Angiopoietin-1-induced upregulation of VEGFR-3 activates sprouting
and hyperplasia of lymphatic vessels (15). Previous studies have suggested that
hypoxia upregulates the expression of VEGFR-3 and VEGF-C in several
cancer cell lines, which yields a significant and vital association
between HIF-1α and VEGFR-3 (16-18).
Although the existence of the HIF-1α/VEGFR-3/VEGF-C axis has been
demonstrated in tumor lymphangiogenesis, it is unknown if the axis
serves a crucial role in the malformation of lymphatic vessels.
The current study aimed to assess the biological
function of HIF-1α in the malformation of human lymphatic
endothelial cells (HLECs) via regulation of VEGFR-3. The results
have demonstrated that HIF-1α expression was upregulated in human
cLM specimens compared with normal tissue, which in turn
upregulated VEGFR-3 expression and in vitro led to the
promotion of colony formation, migration and tube malformation in
HLECs.
Materials and methods
Clinical data and sample collection
cLM and matched cLM-adjacent normal tissues were
obtained from 20 patients (median age, 12.5 months; age range, 16
days to 5 years; male, 11; female, 9), who underwent surgical
resection or a combined operation (endoscopic cautery and
postoperative intratumoral negative pressure) (19) in the Department of Burns and
Plastic Surgery at the Children’s Hospital of Nanjing Medical
University (Nanjing, China) between August 2015 and May 2016.
Clinical characteristics of the patients are summarized in Table I. All tissue samples were
immediately frozen in liquid nitrogen following resection and
stored at −80°C for subsequent experiments. The study protocol was
approved by the Ethics Committee of Children’s Hospital of Nanjing
Medical University (Nanjing, China). Written consent was obtained
prior to the initiation of the study.
 | Table ICharacteristic of 20 patients with
cystic lymphatic malformations. |
Table I
Characteristic of 20 patients with
cystic lymphatic malformations.
| Sex | Age | Location | Histological
type | Lymph property | Treatment |
|---|
| Male | 6 months | Neck | Microcystic | Transparent | CO |
| Female | 2 years | Torso | Mixed cystic | Transparent | CO |
| Female | 3 years | Neck | Mixed cystic | Bloody | CO |
| Male | 2 years | Extremity | Macrocystic | Transparent | SR |
| Male | 1 years | Neck | Macrocystic | Transparent | CO |
| Female | 1 years | Neck | Microcystic | Transparent | CO |
| Male | 2 years | Neck | Mixed cystic | Bloody | CO |
| Male | 5 years | Extremity | Macrocystic | Transparent | SR |
| Female | 5 months | Neck | Microcystic | Transparent | CO |
| Male | 16 days | Neck | Macrocystic | Transparent | CO |
| Male | 1 months | Neck | Mixed cystic | Transparent | CO |
| Female | 4 years | Torso | Macrocystic | Bloody | SR |
| Male | 11 months | Extremity | Mixed cystic | Transparent | SR |
| Female | 2 months | Neck | Macrocystic | Bloody | CO |
| Female | 10 months | Extremity | Microcystic | Transparent | SR |
| Female | 4 years | Neck | Macrocystic | Transparent | CO |
| Male | 20 days | Neck | Microcystic | Transparent | CO |
| Male | 6 years | Torso | Mixed cystic | Bloody | SR |
| Female | 3 months | Neck | Mixed cystic | Transparent | CO |
| Male | 2 months | Neck | Macrocystic | Transparent | CO |
Cell culture
HLECs (cat. no. 2500) were obtained from the Type
Culture Collection of the Chinese Academy of Sciences (Shanghai,
China). Cells were cultured in endothelial cell medium (ECM;
ScienCell Research Laboratories, Inc., San Diego, CA, USA) in a
humidified atmosphere containing 5% CO2 at 37°C.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated from the tissues and cells
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) according to the manufacturer’s instructions. RNA
quantity was measured using a NanoDrop 2000 spectrophotometer
(Thermo Fisher Scientific, Inc.) and RNA quality was assessed via
gel electrophoresis. cDNA was synthesized from total RNA (1
µg) using the PrimeScript® RT kit (Takara
Biotechnology Co., Ltd., Dalian, China) according to the
manufacturer’s instructions by incubation at 37°C for 15 min,
followed by 85°C for 5 sec and storage at 4°C. qPCR was performed
using the SYBR Green PCR master mix (Takara Biotechnology Co.,
Ltd.) in 20 µl reactions according to the manufacturer’s
instructions. Expression of the target genes was relative to that
of β-actin. The reaction protocol included incubations at 95°C for
30 sec, followed by 40 cycles of 95°C for 5 sec and 60°C for 30
sec. All reactions were performed in triplicate and quantification
was performed using the 2−ΔΔCq method (20). Primer sequences were as follows:
HIF1-α, forward, 5′-GAAGACATCGCGGGGAC-3′ and reverse,
5′-TGGCTGCATCTCGAGACTTT-3′; VEGFR-3, forward,
5′-CTCTGCCTGGGACTCCTG-3′ and reverse, 5′-GGTGTCGATGACGTGTGACT-3′;
VEGF-A, forward, 5′-CCCTGGCTTTACTGCTGTAC-3′ and reverse, 5′-TCT
GAACAAGGCTCACAGTG-3′; VEGF-C, forward, 5′-AGTGTCAGGCAGCGAACAAGA-3′
and reverse, 5′-CTTCCTGAGCCAGGCATCTG-3′; sonic hedgehog (SHH),
forward, 5′-GAAAGCAGAGAACTCGGTGG-3′ and reverse,
5′-GGAAAGTGAGGAAGTCGCTG-3′; transforming growth factor (TGF)-β,
forward, 5′-AGCAACAATTCCTGGGGTTACCT-3′ and reverse.
5′-CGAAGCCCTGATTCCGTCTCC-3′; prospero homeobox protein 1 (Prox1),
forward, 5′-TGATCTGAGCAACTTCCAGG-3′ and reverse,
5′-CAACGATGGGGTCACCAGTA-3′; lymphatic vessel endothelial hyaluronan
receptor 1 (LYVE-1), forward, 5′-AGCCTACAGGCCTCCTTAGC-3′ and
reverse, 5′-CTCCTGGTCCAAGGCTCTTT-3′; and β-actin, forward,
5′-AGCGAGCATCCCCCAAAGTT-3′ and reverse,
5′-GGGCACGAAGGCTCATCATT-3′.
Western blot analysis
Frozen tissues or cells were washed twice with
ice-cold PBS and incubated for 30 min on ice with
radioimmunoprecipitation assay buffer (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). The mixture was centrifuged (12,700 × g;
20 min; 4°C), the supernatant was collected and the protein
concentration was determined using a Bradford assay (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Next, 20 µg protein
were separated on 8% SDS-PAGE gels and transferred to 0.22
µm polyvinylidene difluoride membranes. Following blocking
with 5% non-fat milk in TBS containing 0.1% Tween-20 (TBST) at 25°C
for 1 h, primary antibodies for VEGFR-3 (cat. no. ab27278; 1:1,000;
Abcam, Cambridge, UK), HIF-1α (cat. no. 3716S; 1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA) and β-actin (cat. no.
4967L; 1:2,000; Cell Signaling Technology, Inc.) were incubated
with the membranes overnight at 4°C. Following washing with TBST
(3x), membranes were incubated for 2 h at 37°C with horseradish
peroxidase-conjugated secondary antibody to rabbit IgG (cat. no.
ab6727; 1:2,000; Abcam). Proteins were detected using the enhanced
chemiluminescence detection method (Pierce; Thermo Fisher
Scientific, Inc.) and the specific protein bands were detected with
the UVP Bio Imaging system (Tanon Science and Technology Co., Ltd.,
Shanghai, China). The expression of target proteins was
quantitatively analyzed using ImageJ (version 1.43; National
Institutes of Health, Bethesda, MD, USA). β-actin was used as the
loading control.
Cell transfection
cDNA of HIF-1α (2.5 µg; cat. no. ab185916;
Abcam) was cloned into a pCDH-CMV-MCS-EF1-GFP lentivirus vector
(GeneChem, Inc., Daejeon, Korea). Prior to viral infection,
Lentivirus complex containing 4 µg lentiviral plasmid, 4
µg each viral packaging vectors (pRRE, pMD2.G and pRSV;
GeneChem, Inc.) and 800 µl serum-free ECM were added to
HLECs (5×106 cells/well) cultured in 10 cm plates.
Culture medium was collected and filtered through 0.45 µm
polyethersulfone filter. Next, 50 µl polybrene (1
µg/µl) and 1 ml of the viral supernatant containing
the plasmids were added, along with fresh media containing 5 ml ECM
with 10% FBS. At 24 h, the medium was changed and at 72 h, GFP
expression was observed under a fluorescent microscope
(magnification, ×100). HIF-1α-overexpressing cells were selected by
puromycin (1 µg/ml) over 6 days. Lentivirus overexpressing
HIF-1α (GeneChem, Inc.), VEGFR-3 (GeneChem, Inc.) or containing
empty vector (GeneChem, Inc.) were cultured using the enhanced
infection reagent (cat. no. REVG0002; GeneChem, Inc.) according to
the manufacturer’s instructions.
HIF-1α- and VEGFR-3-overexpressing HLECs were seeded
at 1×106 cells/well into 6-well culture plates in ECM
with 10% FBS. Short interfering (si)RNA against HIF-1α (si-HIF-1α),
VEGFR-3 (si-VEGFR-3) and corresponding scramble negative controls
(si-NC) were designed and synthesized by Shanghai GenePharma Co.,
Ltd. (Shanghai, China). Transfections with 50 nM siRNA were
performed in using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer’s instructions.
Subsequent assays were performed at 48 h post transfection. RT-qPCR
and western blot assays were used to analyze HIF-1α and VEGFR-3
expression to determine transfection efficiency. siRNA sequences
were as follows: si-HIF-1α, 5′-CAAGUAGCCUCU UUCACAA-3′; si-VEGFR-3,
5′-CGCCCGAGUUCCAGUGGUA-3′; and si-NC,
5′-UUCUCCGAACGUGUCACGU-3′.
The following experimental groups were established:
HIF-1α, HLECs overexpressing HIF-1α; VEGFR-3, HLECs overexpressing
VEGFR-3; empty vector, HLECs transfected with empty expression
vector; HIF-1α overexpression + VEGFR-3 knockdown, si-VEGFR-3
transfected into HLECs overexpressing HIF-1α; VEGFR-3
overexpression + HIF-1α knockdown, si-HIF-1α transfected into HLECs
overexpressing VEGFR-3; HIF-1α knockdown, HLECs transfected with
si-HIF-1α; VEGFR-3 knockdown, HLECs transfected with si-VEGFR-3;
NC, HLECs transfected with si-NC.
Colony formation and migration
assays
To evaluate colony formation, transfected HLECs were
seeded at 1×103 cells/well into 6-well plates. Plates
were kept at 37°C in an incubator with 5% CO2 for 12
days. Next, cells were washed with PBS, fixed with methanol at 37°C
for 15 min and stained with 2% Giemsa stain (Sigma-Aldrich; Merck
KGaA) at 37°C for 30 min. Positive colonies were photographed and
scored under an inverted light microscope (magnification,
×100).
To evaluate the migration of cells in vitro,
5×105 HLECs from each group were seeded into 35-mm
dishes. When cells had spread to 80-90% of the well surface and
formed a monolayer, the well was scratched with a 200-µl
pipette tip perpendicular to the surface. Cells were washed with
PBS twice followed by the addition of fresh media. Pictures were
taken at 0, 24, 48 and 72 h using an inverted light microscope
(magnification, ×100).
Lymphatic tube formation assay
Mixed fibrinogen gel was prepared by combining 500
µl human fibrinogen (3 mg/ml; Beijing Bio-Ekon Biotechnology
Co., Ltd., Beijing, China) and 10 µl human thrombin (50 U/ml
in PBS; Sigma-Aldrich; Merck KGaA) with 100 µl ECM. For
gelling, the mixture was incubated at 37°C in 5% CO2 for
30 min. Fibrinogen gel (0.05 ml/well) was used to coat the walls of
96-well plates and incubated at 37°C for 1 h. Transfected HLECs
(1×105/ml; 100 µl/well) from the HIF-1α, empty
vector, HIF-1α overexpression + VEGFR-3 knockdown, HIF-1α knockdown
or NC groups were seeded on the gel and cultured in an incubator
with 5% CO2 at 37°C. Media was changed every 4 days.
Following 10 days, cells were observed by inverted phase-contrast
light microscopy (magnification, ×100). Lymphatic tube formation
was determined using morphological characteristics and the number
of lymphatic tubes observed in three random microscopic fields per
well. Western blot assays were performed to analyze HIF-1α and
VEGFR-3 expression at days 4, 6 and 10 during HLECs
lymphangiogenesis.
Determination of adenosine triphosphate
(ATP) content
ATP levels in transfected HLECs from different
groups were determined at 48 h post transfection using the Enhanced
ATP Assay kit (cat. no. S0027; Beyotime Institute of Biotechnology,
Haimen, China) according to the manufacturer’s instructions. Total
ATP levels (range, 0.1-10 µM) were calculated from the
luminescence signals and results are presented in arbitrary
units.
Statistical analysis
Experiments were repeated ≥3 times and data are
presented as mean ± standard deviation. SPSS 19.0 (IBM Corp.,
Armonk, NY, USA) and GraphPad Prism 6.0 (GraphPad Software, Inc.,
La Jolla, CA, USA) were used for statistical analyses. Statistical
differences were evaluated using the Student’s t-test or one-way
analysis of variance followed by Student-Newman-Keuls post-hoc
tests. Correlation between HIF-1α and VEGFR-3 protein expression in
cLM tissues was determined using Pearson analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
HIF-1α and VEGFR-3 are overexpressed in
cLM specimen
mRNA expression of HIF-1α, VEGFR-3, VEGF-A, VEGF-C,
SHH and TGF-β in cLM and matched cLM-adjacent healthy tissues were
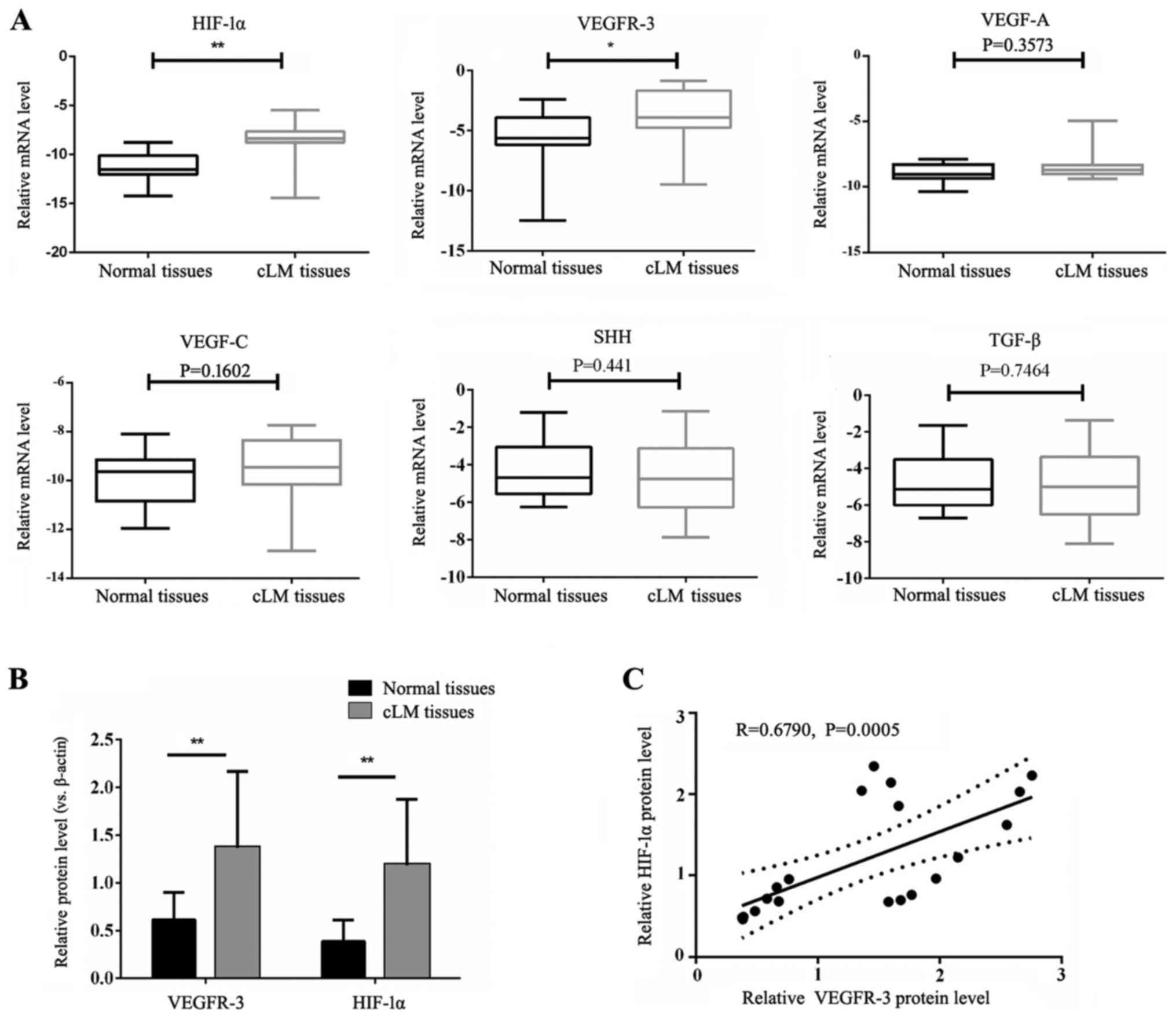
determined by RT-qPCR. As presented in Fig. 1A, HIF-1α and VEGFR-3 mRNA levels
were significantly upregulated in cLM tissues compared with the
adjacent tissues (P<0.05). There were no significant differences
in VEGF-A, VEGF-C, SHH and TGF-β mRNA expression between cLM and
matched healthy tissues. Significantly increased HIF-1α and VEGFR-3
protein levels were observed in cLM tissues compared with matched
healthy tissues (P<0.01; Fig.
1B). Furthermore, analysis revealed a strong positive
correlation between HIF-1α and VEGFR-3 protein expression in cLM
tissues, with a Pearson’s correlation coefficient of 0.6790
(P=0.0005; Fig. 1C).
Characterization of HIF-1α-overexpressing
HLECs
To investigate the biological role of HIF-1α in the
development and progression of lymphatic malformation in HLECs,
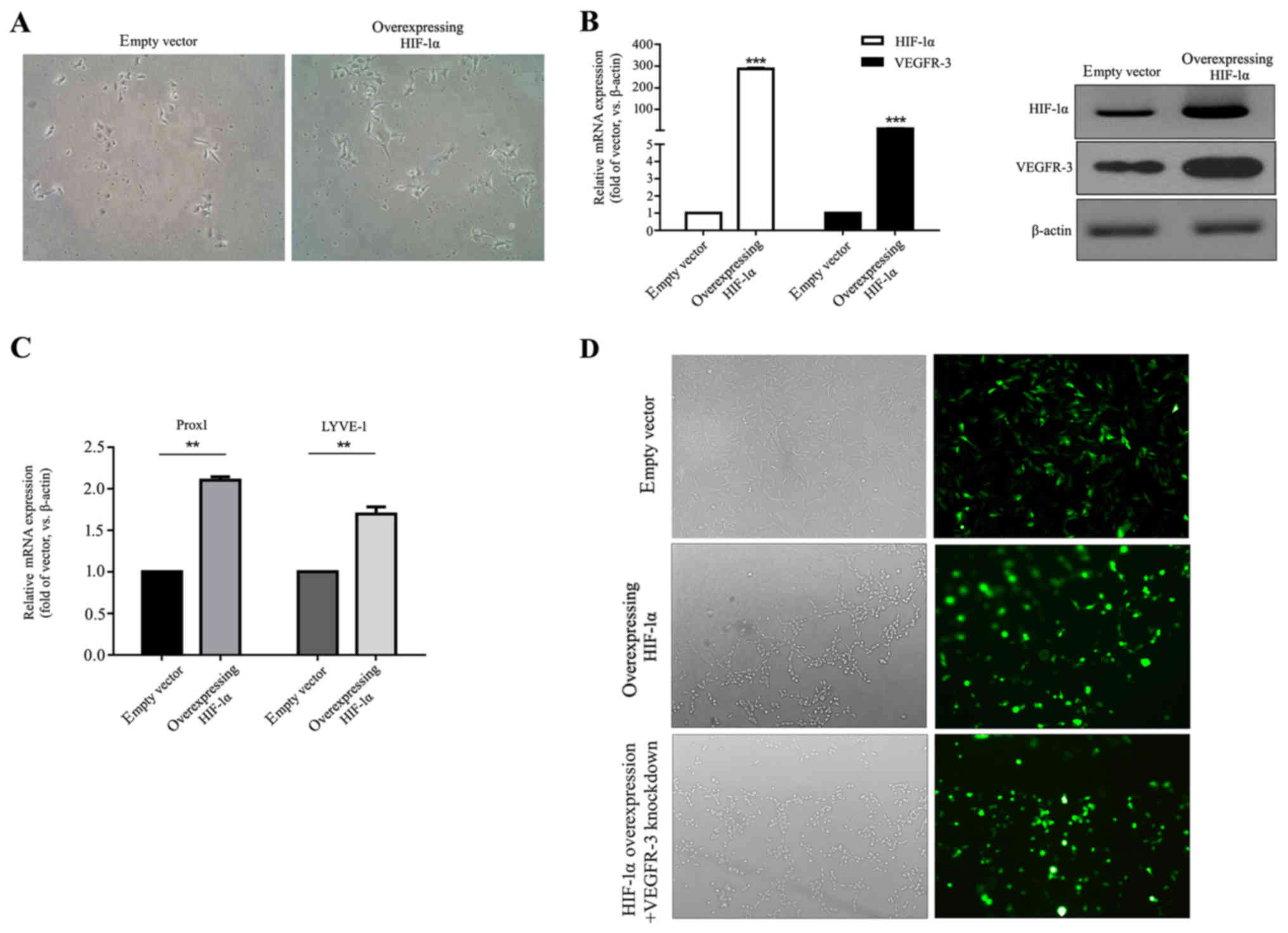
cells overexpressing HIF-1α were created by lentiviral infection.
HIF-1α-overexpressing HLECs maintained the morphology of normal
endothelial cells; however, they appeared multi-angular and
irregular compared with empty vector-transfected HLECs
(magnification, ×100; Fig. 2A).
RT-qPCR and western blot were used to determine HIF-1α and VEGFR-3
expression in HIF-1α-overexpressing HLECs at 72 h post transfection
(Fig. 2B). It was observed that in
HIF-1α-overexpressing HLECs HIF-1α and VEGFR-3 mRNA levels were
significantly increased compared with the empty vector control
(P<0.001) and protein expression was markedly increased in the
HIF-1α-overexpressing HLECs. HIF-1α overexpression significantly
increased expression of lymphatic endothelial markers Prox1 and
LYVE-1 in HLECs compared with the empty vector control (P<0.01;
Fig. 2C). Fluorescence microscopy
to evaluate the GFP content revealed that >80% of HLECs in the
empty vector, the HIF-1α and the HIF-1α overexpression + VEGFR-3
knockdown groups expressed GFP following transfection, indicating a
successfully transfection (Fig.
2D).
Overexpression of HIF-1α promotes colony
formation and migration of HLECs
To determine the role of HIF-1α in cLM progression,
the impact of HIF-1α on colony formation and migration was examined
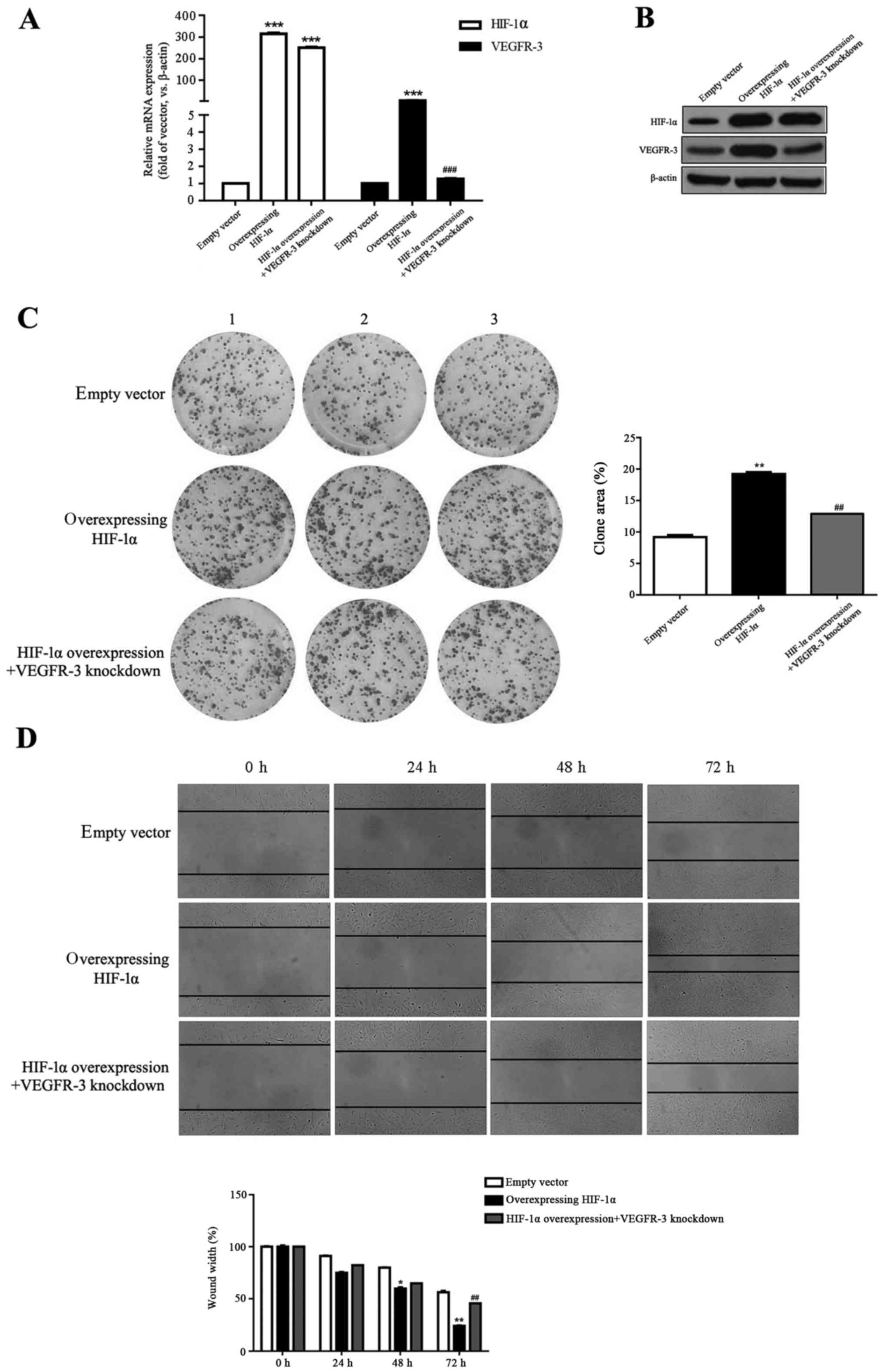
in vitro. RT-qPCR and western blot assays confirmed a
significant increase in HIF-1α mRNA and a marked increase in
protein levels in HIF-1α-overexpressing HLECs compared with the
empty control (P<0.001; Fig. 3A and
B). The significantly increased HIF-1α mRNA and markedly
increased protein levels were maintained in the HIF-1α
overexpression + VEGFR-3 knockdown group compared with the empty
vector control (P<0.001; Fig. 3A
and B). No significant difference in HIF-1α levels was
determined between the HIF-1α and HIF-1α overexpression + VEGFR-3
knockdown groups (P>0.05). VEGFR-3 mRNA levels were
significantly increased and protein levels were markedly increased
in the HIF-1α group compared with the empty vector group
(P<0.001; Fig. 3A and B). For
the HIF-1α overexpression + VEGFR-3 knockdown group, no significant
difference was observed in VEGFR-3 expression compared with the
empty vector group (P>0.05; Fig. 3A
and B). VEGFR-3 mRNA levels were significantly (P<0.001;
Fig. 3A) and protein levels were
markedly (Fig. 3B) decreased in
the HIF-1α overexpression + VEGFR-3 knockdown group compared with
the HIF-1α group.
Overexpression of HIF-1α enhanced colony formation,
as presented by the significant increase in clone area compared
with the empty vector control (P<0.01; Fig. 3C). Interestingly, in the HIF-1α
overexpression + VEGFR-3 knockdown group, there was no significant
difference in clone area compared with the empty vector group
(P>0.05; Fig. 3C). The clone
area in HIF-1α overexpression + VEGFR-3 knockdown group was
significantly decreased compared with the HIF-1α group (P<0.01;
Fig. 3C).
Furthermore, the effect of HIF-1α on HLEC migration
was examined using wound-healing assays. Overexpression of HIF-1α
significantly increased cell migration at 48 and 72 h compared with
empty vector group (P<0.05 and P<0.01, respectively; Fig. 3D). As described above, in the
HIF-1α overexpression + VEGFR-3 knockdown group, no significant
difference in cell migration was observed compared with the empty
vector control (P>0.05; Fig.
3D). Cell migration in the HIF-1α overexpression + VEGFR-3
knockdown group was significantly decreased at 72 h compared with
the HIF-1α group (P<0.01; Fig.
3D). These results suggested that overexpression of HIF-1α
promoted HLEC colony formation and migration by upregulating
VEGFR-3.
HIF-1α knockdown inhibits HLEC colony
formation and migration
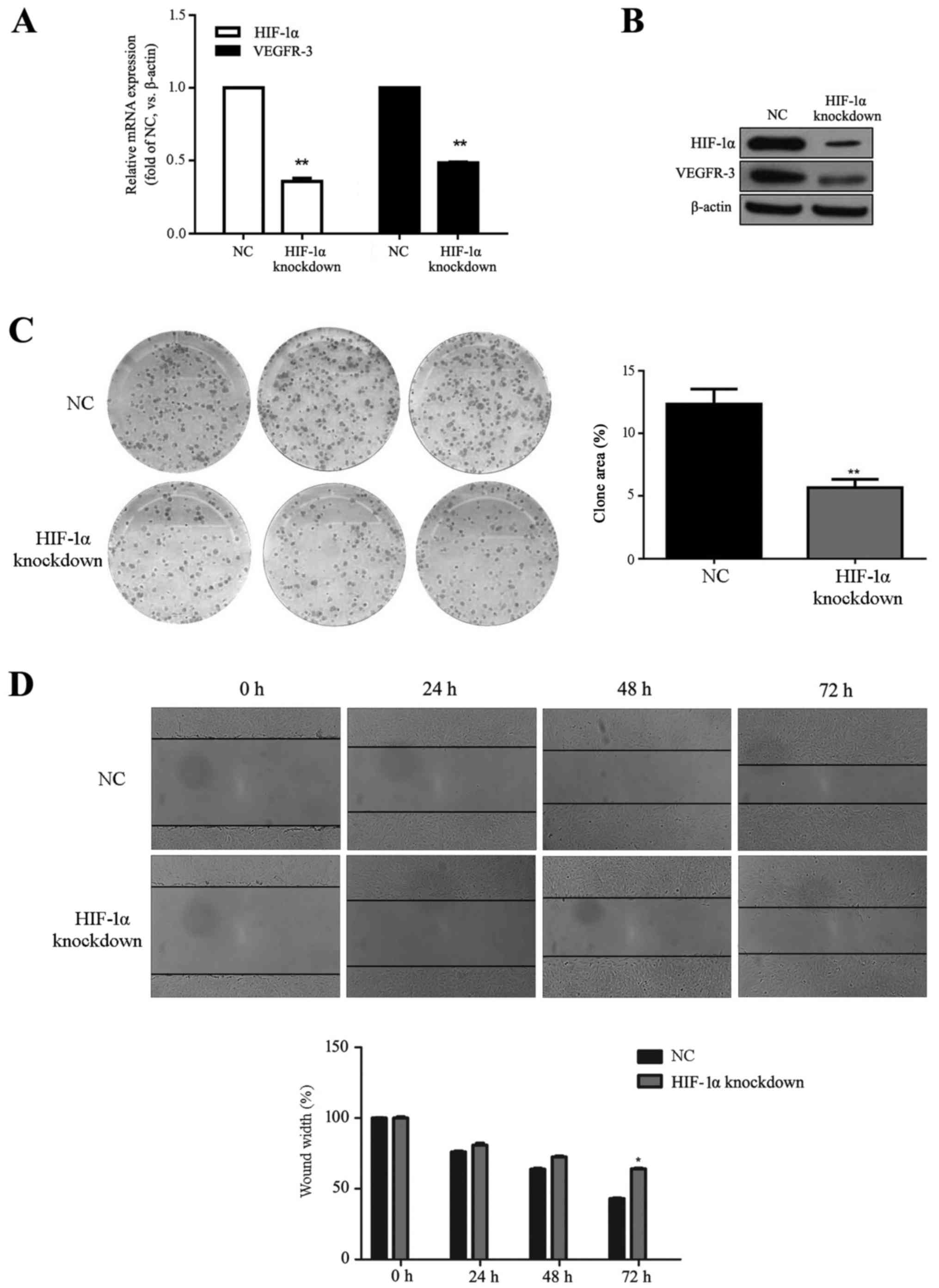
HLECs were transfected with HIF-1α siRNA to observe
knockout effects on colony formation and migration. Transfection
success was determined by RT-qPCR and western blot, demonstrating
significantly decreased mRNA and markedly decreased protein levels
of HIF-1α and VEGFR-3 in the HIF-1α knockdown group compared with
the NC group (P<0.01; Fig. 4A and
B). HIF-1α knockdown significantly decreased clone area
compared with the NC group (P<0.01; Fig. 4C). Cell migration was significantly
reduced at 72 h in the HIF-1α knockdown compared with the NC group
(P<0.05; Fig. 4D). These
results indicated that HIF-1α knockdown reduced HLEC colony
formation and migration.
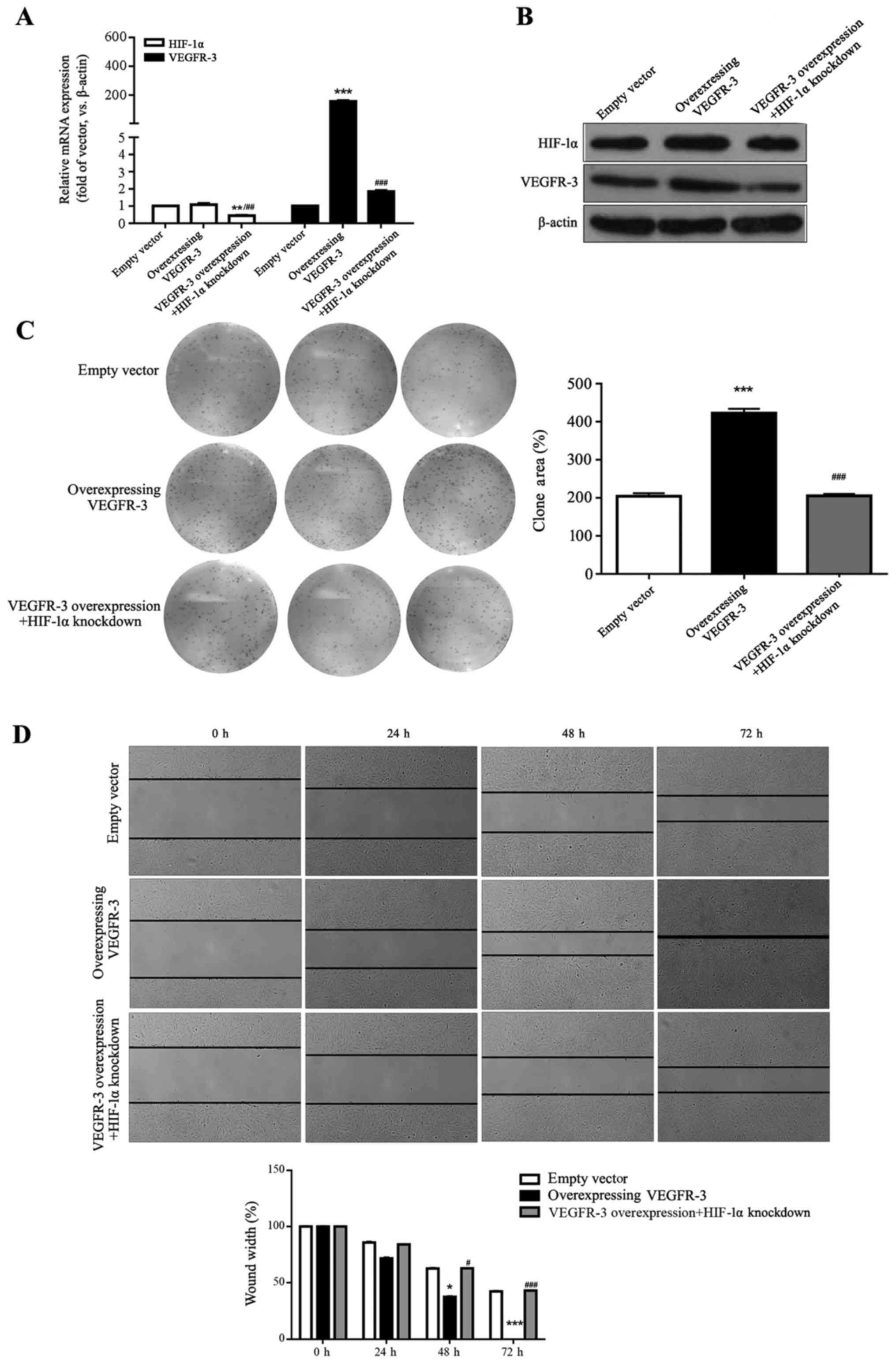
Overexpression of VEGFR-3 promotes HLEC
colony formation and migration
VEGFR-3 overexpression was confirmed by RT-qPCR and
western blot assays, where mRNA levels were significantly and
protein levels were markedly increased compared with the empty
vector control (P<0.001; Fig. 5A
and B). There was no significant difference in VEGFR-3 mRNA
expression in the VEGFR-3 overexpression + HIF-1α knockdown group
compared with empty vector group (P>0.05; Fig. 5A). HIF-1α mRNA and protein levels
were unaffected by VEGFR-3 overexpression; however, mRNA levels
were significantly and protein levels markedly reduced in the
VEGFR-3 overexpression + HIF-1α knockdown group compared with the
empty vector control (P<0.01; Fig.
5A and B). HIF-1α and VEGFR-3 mRNA levels were significantly
and protein levels were markedly decreased in theVEGFR-3
overexpression + HIF-1α knockdown group compared with the VEGFR-3
group (P<0.01 and P<0.001, respectively; Fig. 5A and B).
Colony formation and wound-healing assays were
performed to investigate functions of VEGFR-3 in vitro. As
presented in Fig. 5C, VEGFR-3
overexpression in HLECs significantly increased colony formation
compared with the empty control group (P<0.001). Additionally,
clone area did not change significantly in the VEGFR-3
overexpression + HIF-1α knockdown group compared with the empty
vector control (P>0.05; Fig.
5C). The clone area in the VEGFR-3 overexpression + HIF-1α
knockdown group was significantly decreased compared with the
VEGFR-3 group (P<0.001; Fig.
5C).
VEGFR-3 overexpression significantly promoted HLEC
migration compared with the empty vector control at 48 and 72 h
(P<0.05 and P<0.001, respectively; Fig. 5D) and no significant differences
were observed between the VEGFR-3 overexpression + HIF-1α knockdown
and the empty vector control group (P>0.05; Fig. 5D). Cell migration in the VEGFR-3
overexpression + HIF-1α knockdown group was significantly decreased
at 48 and 72 h compared with the VEGFR-3 group (P<0.05 and
P<0.001, respectively; Fig.
5D). These results indicated that VEGFR-3 levels that were
regulated by HIF-1α expression promoted HLEC colony formation and
migration.
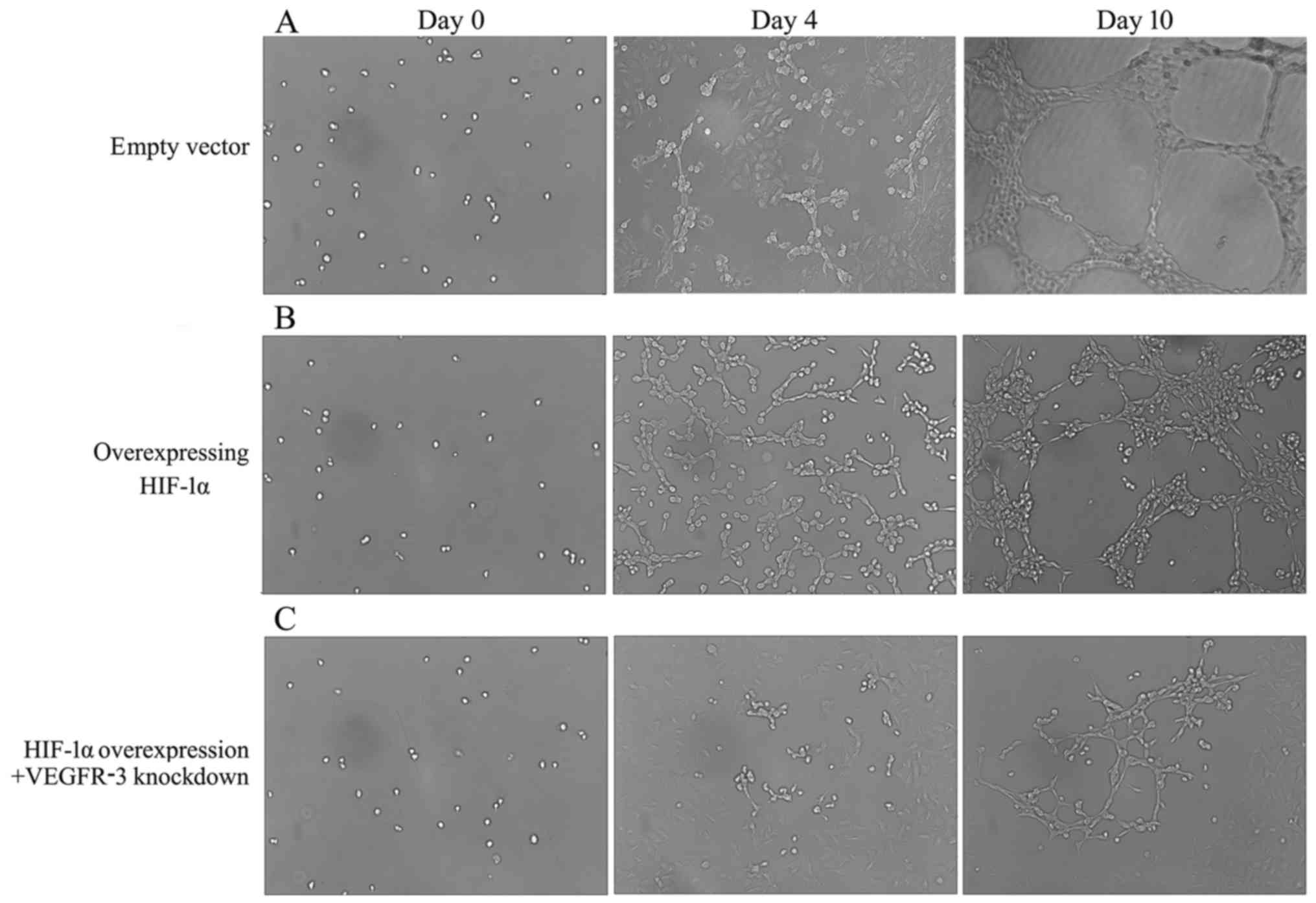
HIF-1α overexpression induces lymphatic
tube malformation in HLECs
The process of lymphangiogenesis was observed in
vitro over 10 days. As presented in Fig. 6, following seeding of cells on the
mixed fibrinogen gel for 4 days, HLECs in the HIF-1α group formed
extensive tubular structures (Fig.
6B) and cells in empty vector group were observed to form cell
clusters (Fig. 6A). Compared with
the empty vector group, fewer cell clusters were observed on day 4
in the HIF-1α overexpression + VEGFR-3 knockdown group (Fig. 6C).
Malformations in the structure and shape of the
lymphatic capillary-type tubules were observed with various
dilatations in the HIF-1α group at day 10 (Fig. 6B). Normal and typical lymphatic
tube structures were exhibited by the empty vector group at the
same time (Fig. 6A). Cells in the
HIF-1α overexpression + VEGFR-3 knockdown group exhibited decreased
formation of tubule walls and a lower density of lymphatic tube
structures compared with the empty vector and the HIF-1α groups
(Fig. 6C). HIF-1α overexpression
induced lymphatic tube malformation in HLECs, potentially by
upregulating VEGFR-3 expression.
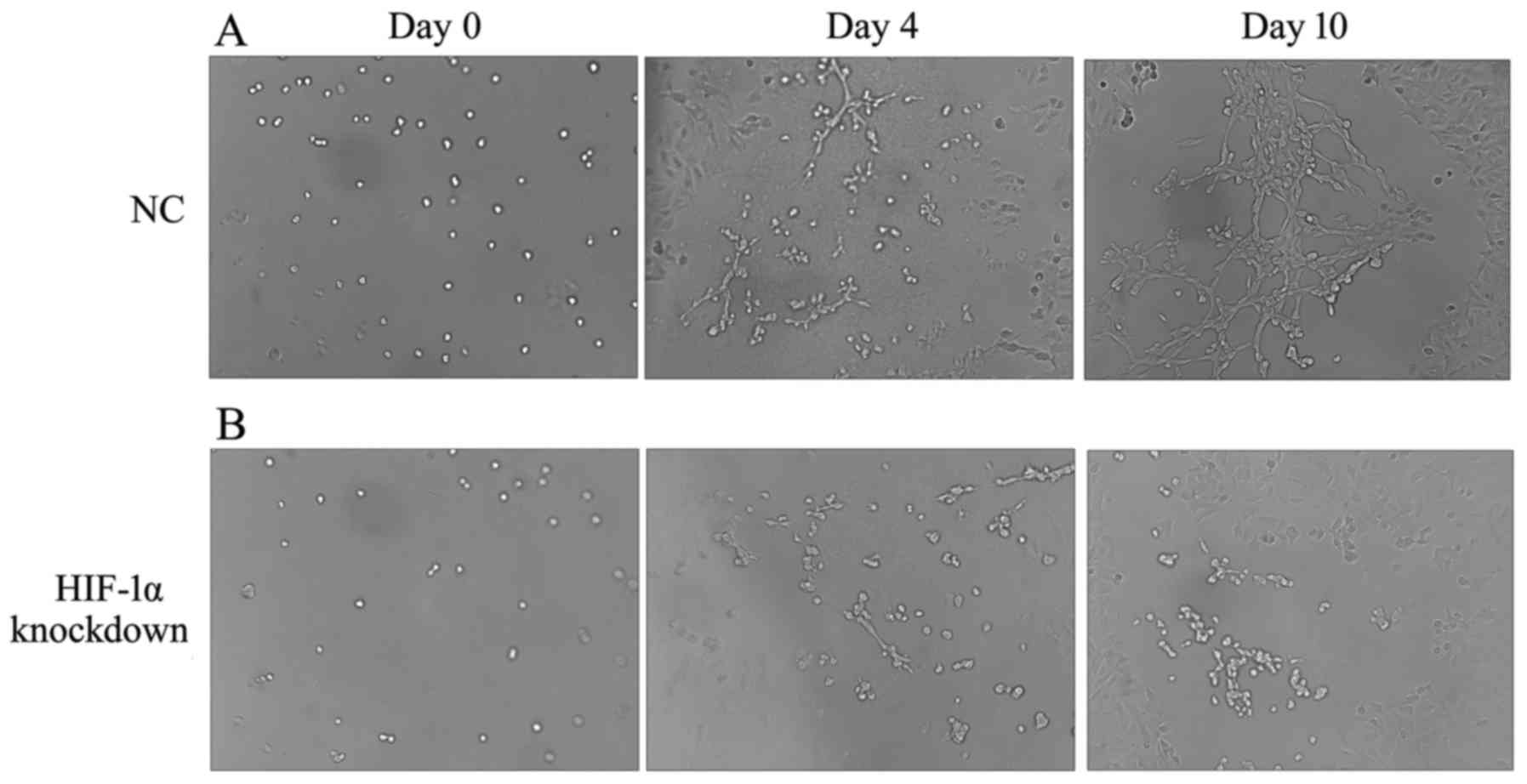
HIF-1α knockdown inhibits lymphatic tube
formation in HLECs
To further investigate the role of HIF-1α in
lymphatic tube formation, HIF-1α knockdown cells were observed.
Cell clusters observed in the HIF-1α knockdown were decreased in
size compared with the NC group at day 4 (Fig. 7A and B). HIF-1α knockdown cells
presented several cell clusters without signs for the formation of
lymphatic tubes at day 10, while typical but small tube structures
were observed in NC group (Fig. 7A and
B). Compared with the empty vector control, smaller dilation of
lymphatic tubes in the NC group may be caused by si-NC
transfection. These findings indicated that the HIF-1α knockdown in
HLECs resulted in the loss of the ability to form lymphatic
tubes.
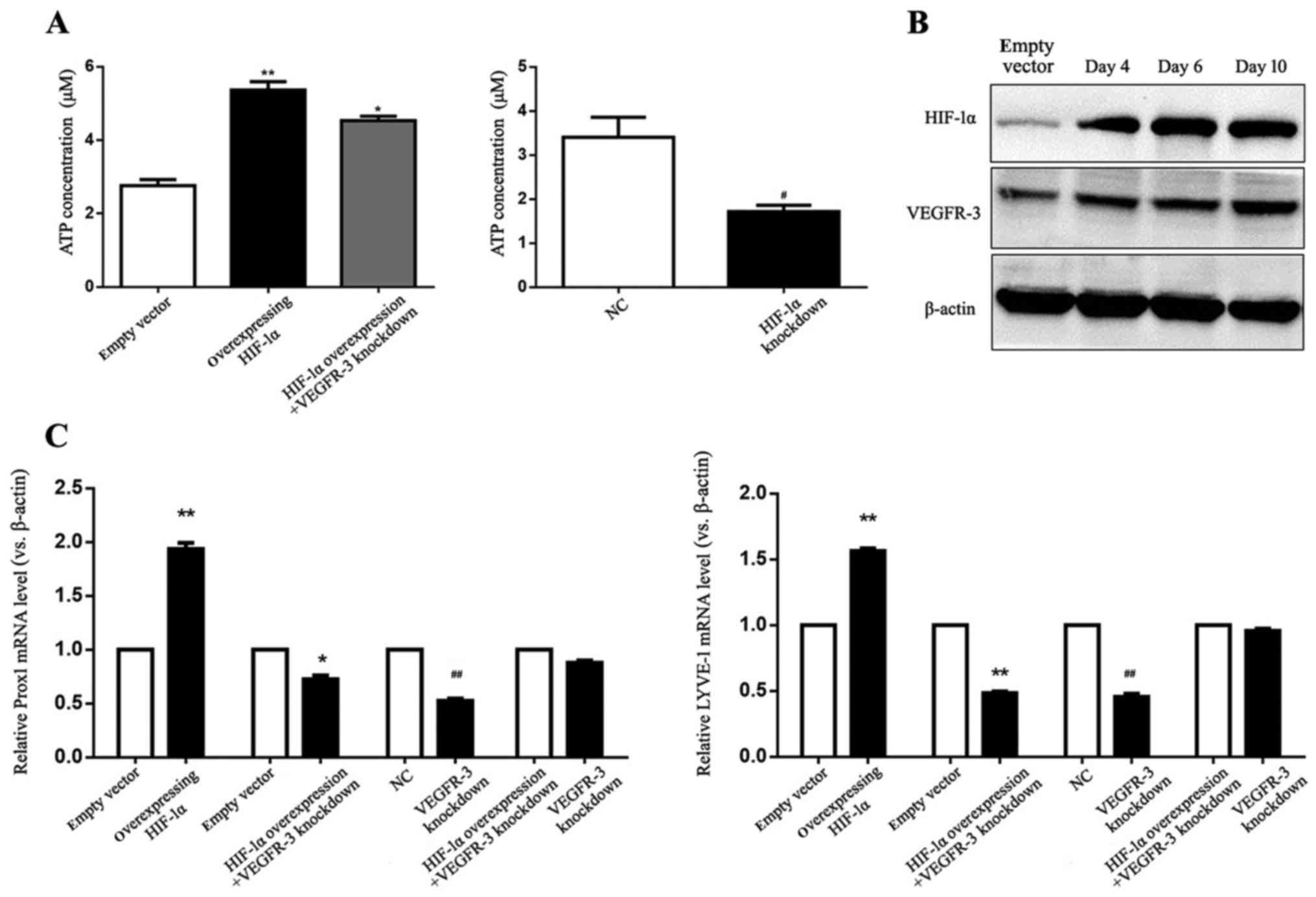
HIF-1α upregulates expression of VEGFR-3
and lymphatic endothelial markers during lymphatic tube
formation
To investigate the role of HIF-1α in HLEC
proliferation during lymphatic tube formation, an enhanced ATP
assay was performed at 48 h post transfection. In
HIF-1α-overexpressing HLECs with or without VEGFR-3 knockdown, ATP
levels increased significantly compared with empty vector group
(P<0.01 and P<0.05, respectively; Fig. 8A). ATP levels in HIF-1α
overexpression + VEGFR-3 knockdown group were significantly
decreased compared with the HIF-1α group (P<0.05; Fig. 8A). In the HIF-1α knockdown group,
ATP levels were significantly decreased compared with the NC group
(P<0.05; Fig. 8A). VEGFR-3 and
HIF-1α protein expression was determined on days 4, 6 and 10 during
HLEC lymphangiogenesis. As presented in Fig. 8B, HIF-1α overexpression increased
VEGFR-3 protein expression in HLECs during lymphatic tube formation
compared with empty vector group (Fig.
8B). To assess the impact of HIF-1α on the expression of
lymphatic endothelial markers, mRNA levels of Prox1 and LYVE-1 at
day 10 during lymphangiogenesis were determined. Prox1 and LYVE-1
mRNA levels were significantly increased in the HIF-1α group
compared with the empty vector control (P<0.01; Fig. 8C). Significantly decreased Prox1
and LYVE-1 expression was observed for the HIF-1α overexpression +
VEGFR-3 knockdown (P<0.05 and P<0.01, respectively) and the
VEGFR-3 knockdown groups compared with the respective empty vector
and NC controls (P<0.01; Fig.
8C). These findings demonstrated that HIF-1α promoted HLEC
proliferation and induced the malformation of lymphatic tubes by
upregulating VEGFR-3 expression and lymphatic endothelial markers
in vitro.
Discussion
According to the 2018 International Society for the
Study of Vascular Anomalies classification of vascular anomalies,
cLM is a primary type of lymphatic anomaly that includes
generalized lymphatic anomaly, lymphatic malformations in
Gorham-Stout disease, primary lymphedema and several other subtypes
(21). The lymphatic channels of
cLM dilate to various degrees and are lined by endothelial cells
with normal lymphatic phenotype (1). The pathogenesis of cLM remains
unclear and molecular events leading to tubular malformation in
lymphatic endothelial cells are not fully understood, preventing
improvements in clinical prevention and treatment.
HIF-1α mediates transcriptional activation of
lymphangiogenesis via regulation of signaling pathways, including
VEGF-C, SHH and TGF-β in certain human tumors (22,23).
Previous studies demonstrated that HIF-1α inhibition impaired
gastric cancer growth and vascular endothelial cells in
angiogenesis (24), while
hypoxia-dependent activation of the nuclear factor-κB/HIF-1α/VEGF
signaling pathway contributed to gastric cancer promotion via
enhancing angiogenesis (25).
HIF-1α and VEGF expression have further been suggested to function
as determinants of tumor angiogenesis in squamous cell carcinoma of
the esophagus (26). Schoppmann
et al (27) indicated that
a significant association between HIF-1α expression and the density
of peritumoral lymphatic microvessels provided evidence for a
potential role of HIF-1α as regulator of tumor-associated
lymphangiogenesis in breast cancer. Additionally, upregulation of
HIF-1α results in activation of the HIF-1α/VEGF signaling pathway
and promotes hypoxia-induced angiogenesis in lung cancer (28). Under hypoxic conditions, certain
cancer cells upregulate VEGF-C protein levels in vitro and
in vivo, inducing lymphatic vessel growth in and around the
hypoxic tumor environments (29-31).
VEGFR-3, which is primarily located in the lymphatic endothelium,
is overexpressed under hypoxic conditions in many carcinoma cell
lines (32,33). In addition, it serves critical
roles during lymphangiogenesis and tumor metastasis in breast
cancer (34). HIF-1α further
regulates VEGFR-3 expression in lymphangiogenesis during invasion
and metastasis of lung adenocarcinoma and non-small cell lung
cancer (35,36). Inspired by this, the current study
investigated the role and biological function of HIF-1α in cLM
progression.
To uncover the potential association between HIF-1α
and certain signaling pathways, mRNA expression of HIF-1α, VEGFR-3,
VEGF-A, VEGF-C, SHH and TGF-β in cLM and adjacent normal tissues
was investigated. HIF-1α and VEGFR-3 expression were significantly
elevated at mRNA levels in cLM tissues compared with the normal
tissues and higher HIF-1α and VEGFR-3 protein levels were detected
in the cLM tissues. Pearson’s correlation analysis revealed a
strong positive correlation between HIF-1α and VEGFR-3 protein
expression in cLM tissues. It was hypothesized that a significant
upregulation of HIF-1α may be accompanied by an increase of VEGFR-3
in cLM and that HIF-1α in conjunction with VEGFR-3 potentially
contribute to the progression of cLM.
As previous studies have revealed that HIF-1α is
involved in lymphangiogenesis via regulation of VEGFR-3, the
biological role of HIF-1α in the functional regulation of lymphatic
tube formation in HLECs was investigated (32,35).
Reports have demonstrated that lymphatic malformations exhibit
cobblestone morphology, which is a characteristic of lymphatic
endothelial cells (37,38). HIF-1α-overexpressing HLECs were
established and co-transfection with si-VEGFR-3 was performed.
Transfection efficiency was evaluated by monitoring GFP expression
and the success of creating an overexpression or knockdown system
was determined by RT-qPCR and western blot. To investigate the role
of VEGFR-3 in HIF-1α-overexpressing HLECs, a HIF-1α overexpression
+ VEGFR-3 knockdown group and a HIF-1α group or empty vector
control group were compared in colony formation, migration, and
lymphangiogenesis assays. The absence of a HIF-1α-overexpression +
si-NC control is a limitation of the current study, as effects of
siRNA transfection on the HIF-1α-overexpression system were not
evaluated. No obvious differences in morphology between the
HIF-1α-overexpressing and empty vector HLECs were observed;
however, it was determined that HIF-1α overexpression significantly
promoted colony formation and HLEC migration. As cells use
glycolysis in the primary mechanism for ATP production under
hypoxic conditions, certain HIF-1α target genes involved in
glycolysis may enhance proliferation and migration abilities of
cells (39,40). Potentially this mechanism supports
how HIF-1α enhanced HLEC colony formation and migration in the
present study. Interestingly, the findings suggested that HIF-1α
overexpression in HLECs accelerated lymphangiogenesis, by reshaping
cells into irregular tubes with different dilatations.
HIF-1α-overexpressing HLECs with VEGFR-3 knockdown
exhibited decreased levels of colony formation, migration and
lymphangiogenesis compared with the HIF-1α-overexpressing cells. It
was confirmed that VEGFR-3 protein levels were increased in HIF-1α
overexpression cells during lymphangiogenesis and VEGFR-3 was
identified as directly associated with HIF-1α. VEGFR-3
overexpression resulted in similar effects on HLEC colony formation
and migration as observed for HIF-1α overexpression and no
significant changes in colony formation and migration were observed
for the VEGFR-3 overexpression + HIF-1 αknockdown group compared
with the empty vector control.
ATP is an indicator for proliferation (41,42).
Results assessing ATP levels during lymphangiogenesis revealed that
HIF-1α overexpression significantly promoted HLEC proliferation.
Furthermore, mRNA expression of Prox1 and LYVE-1, which were
previously described to be upregulated in endothelial cells with
lymphatic malformations (38),
were investigated. It was observed that HIF-1α overexpression
upregulated mRNA expression by targeting VEGFR-3 during the
lymphatic tube formation.
As there is no animal model of cLM currently
available, in vivo mechanisms were not investigated. Further
studies are required to explore whether HIF-1α contributes to
lymphatic tube formations in HLECs via specific signaling pathways
and whether HIF-1α promotes lymphatic malformations in vivo.
Further limitations are originating from the missing controls in
the co-transfection experiments. To exclude potential influences of
the secondary transfection step on the system, future research
should include all relevant controls (43-47).
In conclusion, the current study demonstrated that
HIF-1α and VEGFR-3 were upregulated in cLM tissues. The role of
HIF-1α was investigated in HLEC colony formation, migration and
lymphatic malformations and it was revealed to be associated with
VEGFR-3 upregulation in vitro. By understanding the
molecular function of HIF-1α as a promoter of lymphatic
malformations, HIF-1α may become a viable target of future cLM
therapeutics.
Acknowledgments
Not applicable.
Funding
The current study was supported by the Science and
Technique Development Foundation of Nanjing Medical University
(grant no. 2015NJMU072).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors’ contributions
JZ and WS designed the experiments and conducted the
data analyses. TH and YJ performed the clinical sample preparation
and validated HIF-1α and VEGFR-3 expression in cLM specimens. HC
and YJ performed in vitro assays. JiaC and JieC performed
the statistical analyses. TH wrote and edited the manuscript. All
authors participated in the discussion and revision of the
manuscript. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The protocols of this study were approved by the
Ethics Committee of Children’s Hospital of Nanjing Medical
University (Nanjing, China). Written informed consent was obtained
from all patients or their legal guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wassef M, Blei F, Adams D, Alomari A,
Baselga E, Berenstein A, Burrows P, Frieden IJ, Garzon MC,
Lopez-Gutierrez JC, et al ISSVA Board and Scientific Committee:
Vascular anomalies classification: Recommendations from the
International Society for the study of vascular anomalies.
Pediatrics. 136:e203–e214. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Florez-Vargas A, Vargas SO, Debelenko LV,
Perez-Atayde AR, Archibald T, Kozakewich HP and Zurakowski D:
Comparative analysis of D2-40 and LYVE-1 immunostaining in
lymphatic malformations. Lymphology. 41:103–110. 2008.PubMed/NCBI
|
|
3
|
Defnet AM, Bagrodia N, Hernandez SL,
Gwilliam N and Kandel JJ: Pediatric lymphatic malformations:
Evolving understanding and therapeutic options. Pediatr Surg Int.
32:425–433. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mulliken JB, Burrows PE and Fishman SJ:
Mulliken and Young’s vascular anomalies: hemangiomas and
malformations. 2nd edition. Oxford University Press; Oxford; 2013,
View Article : Google Scholar
|
|
5
|
Fonkalsrud EW: Lymphatic disorders.
Pediatric surgery. Grosfeld JL: Mosby/Elsevier; Philadelphia, PA:
pp. 2137–2146. 2006, View Article : Google Scholar
|
|
6
|
Sun ZJ, Zhang L, Zhang WF, Liu B, Li ZB
and Zhao YF: A possible hypoxia-induced endothelial proliferation
in the pathogenesis of epithelioid hemangioma. Med Hypotheses.
67:1133–1135. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
North PE, Waner M, Mizeracki A and Mihm MC
Jr: GLUT1: A newly discovered immunohistochemical marker for
juvenile hemangiomas. Hum Pathol. 31:11–22. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Revencu N, Boon LM, Mendola A, Cordisco
MR, Dubois J, Clapuyt P, Hammer F, Amor DJ, Irvine AD, Baselga E,
et al: RASA1 mutations and associated phenotypes in 68 families
with capillary malformation-arteriovenous malformation. Hum Mutat.
34:1632–1641. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liao D and Johnson RS: Hypoxia: A key
regulator of angiogenesis in cancer. Cancer Metastasis Rev.
26:281–290. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fulda S and Debatin KM: HIF-1-regulated
glucose metabolism: A key to apoptosis resistance? Cell Cycle.
6:790–792. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Simon F, Bockhorn M, Praha C, Baba HA,
Broelsch CE, Frilling A and Weber F: Deregulation of HIF1-alpha and
hypoxia-regulated pathways in hepatocellular carcinoma and
corresponding non-malignant liver tissue - influence of a modulated
host stroma on the prognosis of HCC. Langenbecks Arch Surg.
395:395–405. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tao J, Li T, Li K, Xiong J, Yang Z, Wu H
and Wang C: Effect of HIF-1alpha on VEGF-C induced
lymphangiogenesis and lymph nodes metastases of pancreatic cancer.
J Huazhong Univ Sci Technolog Med Sci. 26:562–564. 2006. View Article : Google Scholar
|
|
13
|
Katsuta M, Miyashita M, Makino H, Nomura
T, Shinji S, Yamashita K, Tajiri T, Kudo M, Ishiwata T and Naito Z:
Correlation of hypoxia inducible factor-1alpha with lymphatic
metastasis via vascular endothelial growth factor-C in human
esophageal cancer. Exp Mol Pathol. 78:123–130. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Secker GA and Harvey NL: VEGFR signaling
during lymphatic vascular development: From progenitor cells to
functional vessels. Dev Dyn. 244:323–331. 2015. View Article : Google Scholar
|
|
15
|
Tammela T, Saaristo A, Lohela M, Morisada
T, Tornberg J, Norrmén C, Oike Y, Pajusola K, Thurston G, Suda T,
et al: Angiopoietin-1 promotes lymphatic sprouting and hyperplasia.
Blood. 105:4642–4648. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liang X, Yang D, Hu J, Hao X, Gao J and
Mao Z: Hypoxia inducible factor-alpha expression correlates with
vascular endothelial growth factor-C expression and
lymphangiogenesis/angiogenesis in oral squamous cell carcinoma.
Anticancer Res. 28(3A): 1659–1666. 2008.PubMed/NCBI
|
|
17
|
Mizukami Y, Li J, Zhang X, Zimmer MA,
Iliopoulos O and Chung DC: Hypoxia-inducible factor-1-independent
regulation of vascular endothelial growth factor by hypoxia in
colon cancer. Cancer Res. 64:1765–1772. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Boussat S, Eddahibi S, Coste A, Fataccioli
V, Gouge M, Housset B, Adnot S and Maitre B: Expression and
regulation of vascular endothelial growth factor in human pulmonary
epithelial cells. Am J Physiol Lung Cell Mol Physiol.
279:L371–L378. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shen W, Weiping S, Cui J, Chen J and Zou
J: Management of cystic lymphangioma in the head and neck region:
Endoscopic cautery and postoperative intratumoral negative
pressure. J Craniofac Surg. 21:1884–1886. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Perkins JA: New frontiers in our
understanding of lymphatic malformations of the head and neck:
Natural history and basic research. Otolaryngol Clin North Am.
51:147–158. 2018. View Article : Google Scholar
|
|
22
|
Ji RC: Hypoxia and lymphangiogenesis in
tumor microenvironment and metastasis. Cancer Lett. 346:6–16. 2014.
View Article : Google Scholar
|
|
23
|
Chen S, Zhang M, Xing L, Wang Y, Xiao Y
and Wu Y: HIF-1α contributes to proliferation and invasiveness of
neuroblastoma cells via SHH signaling. PLoS One. 10:e01211152015.
View Article : Google Scholar
|
|
24
|
Stoeltzing O, McCarty MF, Wey JS, Fan F,
Liu W, Belcheva A, Bucana CD, Semenza GL and Ellis LM: Role of
hypoxia-inducible factor 1alpha in gastric cancer cell growth,
angiogenesis, and vessel maturation. J Natl Cancer Inst.
96:946–956. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nam SY, Ko YS, Jung J, Yoon J, Kim YH,
Choi YJ, Park JW, Chang MS, Kim WH and Lee BL: A hypoxia-dependent
upregulation of hypoxia-inducible factor-1 by nuclear factor-κB
promotes gastric tumour growth and angiogenesis. Br J Cancer.
104:166–174. 2011. View Article : Google Scholar
|
|
26
|
Kimura S, Kitadai Y, Tanaka S, Kuwai T,
Hihara J, Yoshida K, Toge T and Chayama K: Expression of
hypoxia-inducible factor (HIF)-1alpha is associated with vascular
endothelial growth factor expression and tumour angiogenesis in
human oesophageal squamous cell carcinoma. Eur J Cancer.
40:1904–1912. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schoppmann SF, Fenzl A, Schindl M,
Bachleitner-Hofmann T, Nagy K, Gnant M, Horvat R, Jakesz R and
Birner P: Hypoxia inducible factor-1alpha correlates with VEGF-C
expression and lymphangiogenesis in breast cancer. Breast Cancer
Res Treat. 99:135–141. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu H and Zhang S: Hypoxia inducible
factor-1α/vascular endothelial growth factor signaling activation
correlates with response to radiotherapy and its inhibition reduces
hypoxia-induced angiogenesis in lung cancer. J Cell Biochem.
119:7707–7718. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Morfoisse F, Kuchnio A, Frainay C,
Gomez-Brouchet A, Delisle MB, Marzi S, Helfer AC, Hantelys F, Pujol
F, Guillermet-Guibert J, et al: Hypoxia induces VEGF-C expression
in metastatic tumor cells via a HIF-1α-independent
translation-mediated mechanism. Cell Reports. 6:155–167. 2014.
View Article : Google Scholar
|
|
30
|
Kuwai T, Kitadai Y, Tanaka S, Onogawa S,
Matsutani N, Kaio E, Ito M and Chayama K: Expression of
hypoxia-inducible factor-1alpha is associated with tumor
vascularization in human colorectal carcinoma. Int J Cancer.
105:176–181. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Koshikawa N, Iyozumi A, Gassmann M and
Takenaga K: Constitutive upregulation of hypoxia-inducible
factor-1alpha mRNA occurring in highly metastatic lung carcinoma
cells leads to vascular endothelial growth factor overexpression
upon hypoxic exposure. Oncogene. 22:6717–6724. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Simiantonaki N, Jayasinghe C,
Michel-Schmidt R, Peters K, Hermanns MI and Kirkpatrick CJ:
Hypoxia-induced epithelial VEGF-C/VEGFR-3 upregulation in carcinoma
cell lines. Int J Oncol. 32:585–592. 2008.PubMed/NCBI
|
|
33
|
Okada K, Osaki M, Araki K, Ishiguro K, Ito
H and Ohgi S: Expression of hypoxia-inducible factor (HIF-1alpha),
VEGF-C and VEGF-D in non-invasive and invasive breast ductal
carcinomas. Anticancer Res. 25:3003–3009. 2005.PubMed/NCBI
|
|
34
|
Liu ZY, Qiu HO, Yuan XJ, Ni YY, Sun JJ,
Jing W and Fan YZ: Suppression of lymphangiogenesis in human
lymphatic endothelial cells by simultaneously blocking VEGF-C and
VEGF-D/VEGFR-3 with norcantharidin. Int J Oncol. 41:1762–1772.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Su JL, Yen CJ, Chen PS, Chuang SE, Hong
CC, Kuo IH, Chen HY, Hung MC and Kuo ML: The role of the
VEGF-C/VEGFR-3 axis in cancer progression. Br J Cancer. 96:541–545.
2007. View Article : Google Scholar
|
|
36
|
Teng X, Li D and Johns RA: Hypoxia
up-regulates mouse vascular endothelial growth factor D promoter
activity in rat pulmonary microvascular smooth-muscle cells. Chest.
121(Suppl): 82S–83S. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
O TM and Lou MS: Zinc effect on human
lymphatic malformation cells in vitro. Int J Pediatr
Otorhinolaryngol. 80:33–38. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Boscolo E, Coma S, Luks VL, Greene AK,
Klagsbrun M, Warman ML and Bischoff J: AKT hyper-phosphorylation
associated with PI3K mutations in lymphatic endothelial cells from
a patient with lymphatic malformation. Angiogenesis. 18:151–162.
2015. View Article : Google Scholar :
|
|
39
|
Semenza GL: HIF-1 mediates metabolic
responses to intra-tumoral hypoxia and oncogenic mutations. J Clin
Invest. 123:3664–3671. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hong SS, Lee H and Kim KW: HIF-1alpha: A
valid therapeutic target for tumor therapy. Cancer Res Treat.
36:343–353. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Oliveira SL, Trujillo CA, Negraes PD and
Ulrich H: Effects of ATP and NGF on Proliferation and Migration of
Neural Precursor Cells. Neurochem Res. 40:1849–1857. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen JB, Liu WJ, Che H, Liu J, Sun HY and
Li GR: Adenosine-5′-triphosphate up-regulates proliferation of
human cardiac fibroblasts. Br J Pharmacol. 166:1140–1150. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Song X, Rui C, Meng L, Zhang R, Shen R,
Ding H, Li J, Li J and Long W: Long non-coding RNA RPAIN regulates
the invasion and apoptosis of trophoblast cell lines via complement
protein C1q. Oncotarget. 8:7637–7646. 2017.
|
|
44
|
Xia H, Li Y and Lv X: MicroRNA-107
inhibits tumor growth and metastasis by targeting the BDNF-mediated
PI3K/AKT pathway in human non-small lung cancer. Int J Oncol.
49:1325–1333. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xie F, Huang Q, Liu CH, Lin XS, Liu Z, Liu
LL, Huang DW and Zhou HC: MiR-1271 negatively regulates AKT/MTOR
signaling and promotes apoptosis via targeting PDK1 in pancreatic
cancer. Eur Rev Med Pharmacol Sci. 22:678–686. 2018.PubMed/NCBI
|
|
46
|
Wang J, Yang S, Ge W, Wang Y, Han C and Li
M: MiR-613 suppressed the laryngeal squamous cell carcinoma
progression through regulating PDK1. J Cell Biochem. 119:5118–5125.
2018. View Article : Google Scholar
|
|
47
|
Dai J, Wang J, Yang L, Xiao Y and Ruan Q:
miR-125a regulates angiogenesis of gastric cancer by targeting
vascular endothelial growth factor A. Int J Oncol. 47:1801–1810.
2015. View Article : Google Scholar : PubMed/NCBI
|