Introduction
Gastric cancer (GC) is the most common type of
gastrointestinal tumor worldwide (1). Recent research has demonstrated that
its mortality rate is the third leading cause of mortality in
China, and the incidence of GC remains high (2). Therefore, it is urgent to identify
chemotherapeutic agents for the treatment of GC and to improve
treatment strategies worldwide.
Numerous components derived from natural substances
can inhibit tumor proliferation (3-5).
Investigation into natural substances provides a prospective method
to understand the mechanism of tumorigenesis. Solamargine, a
natural glycoalkaloid compound present in a traditional herbal
medicine called Solanum nigrum L. (6), is reported to possess a variety of
bioactivities, including antiviral, antitumor and anti-inflammatory
properties (7). Previously,
numerous studies have investigated the inhibitory effects of
Solamargine on tumorigenesis (8-10).
Preliminary studies of its role and its potential mechanisms in
lung cancer (8), hepatocellular
carcinoma (11), breast (12), prostate (13) and ovarian cancers (14), and various tumor cell lines
(15) have been reported; however,
the effects of Solamargine on GC remain unknown. It was suggested
that the inhibitory effects of Solamargine are dependent on the
feedback regulation of extracellular signal-regulated kinase
(Erk)1/2 mitogen-activated protein kinase (MAPK) (16,17).
Whether the potential mechanism of Solamargine involves MAPK
regulation requires further investigation.
Long non-coding RNA (lncRNA) refers to transcripts
>200 nucleotides and do not possess protein coding functions
(6). Accumulating research has
demonstrated that the accurate regulation of signaling pathways by
lncRNA serves a pivotal role in the malignant transformation of
cells (18). The study of lncRNA
p53 induced transcript (lncPINT) in pancreatic cancer demonstrated
that its low expression may be an indicator of poor prognosis
(19). Marín-Béjar et al
(7) revealed that the expression
of lncPINT was low in colorectal cancer and its overexpression
served a crucial role in tumor progression. Additionally, lnc
nuclear paraspeckle assembly transcript 1 (lncNEAT1) was
differentially expressed in a variety of solid tumors (20-22);
several studies have reported that lncNEAT1 is associated with the
prognosis of tumors (23,24). To gain insight into the potential
mechanism of the effects of Solamargine on the regulation of
lncRNA, further investigation should be conducted.
The present study aimed to determine whether
Solamargine is effective against GC. Additionally, analysis of the
underlying mechanisms may provide potential therapeutic targets to
improve treatment strategies in GC. Furthermore, the anti-tumor
effects of Solamargine demonstrated in primary GC cells from
patients may contribute to developments into the treatment of
gastric cancer.
Materials and methods
Drug and chemicals
Solamargine was obtained from MedChem Express USA
(Monmouth Junction, NJ, US). The drug was dissolved in dimethyl
sulfoxide (DMSO; Sigma-Aldrich; Merck KGaA) and prepared as a 10 mM
stock solution, which was stored at −80°C and freshly diluted by
cell culture medium to the final concentrations (0.15625, 0.3125,
0.625, 1.25, 2.5, 5, 7.5, 10 and 20 µM) immediately prior to
use. U0126 (10 µM) was purchased from Selleck Chemicals
(Houston, TX, USA) and freshly diluted by cell culture medium to
the final concentrations (10 µM). GC cell lines (SGC7901 and
BGC823) were treated with Erk1/2 MAPK inhibitor U0126 (10
µM) for 0, 12 and 24 h, or 7.5 µM for 36 h at 37°C
and 5% CO2.
Cell viability assay and drug sensitivity
assay
The GC cell lines AGS, BGC823, SGC7901, HGC27 and
MGC803 were obtained from Chinese Academy of Medical Science
(Beijing, China). BGC823, SGC7901 and HGC27 commonly used in our
research center were chosen in the further analysis. Primary GC
cells were obtained from four patients (3 males and 1 female) with
GC who underwent radical gastrectomy at Peking University Beijing
Cancer Hospital in 2017 (Beijing, China); the age ranged from 34 to
67 years old. Informed consent was provided by all patients. Cell
culture medium was Dulbecco's modified Eagle's medium (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, US) supplemented with
10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) at
37°C and 5% CO2. Cells were cultured in cell culture
medium containing Solamargine (0.15625, 0.3125, 0.625, 1.25, 2.5,
5, 7.5, 10 and 20 µM, for the control, equivalent volume of
DMSO) for 88 h at 37°C and 5% CO2. The IncuCyte ZOOM
Live-Cell Analysis system (Essen BioScience, Inc., Ann Arbor, MI,
USA) which automatically acquires images of living cells and
determines the confluence at appropriate time intervals (every 4
h), was employed for cell viability analysis. The viability of
cells was monitored by an IncuCyte® Live Cell Analysis
system, where viability was assessed by determining the confluence
of cells. The half-maximal inhibitory concentration
(IC50) was calculated using GraphPad Prism statistical
software (version 5.0, GraphPad Software, Inc., La Jolla, CA,
USA).
Western blot analysis
Protein was extracted from cultured cells and lysed
with radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology, Haimen, China), phenylmethanesulfonyl
fluoride (Beyotime Institute of Biotechnology) and phosphatase
inhibitor cocktail I (MedChemExpress, Monmouth Junction, NJ, USA).
The protein concentration was measured via the BCA method (BCA
Protein Assay kit, Beyotime Institute of Biotechnology), prior to
each lysate being boiled for 4 min. Equal quantities (20 µg)
of protein were added. Proteins were separated by 12% SDS-PAGE, and
electrophoresed at 120 V for 2 h. Separated proteins were
transferred onto a polyvinylidene difluoride membrane at 250 V for
2 h on ice. Following protein transfer, the membranes were blocked
with 5% non-fat dry milk in 1X Tris-buffered saline with Tween-20
(50 mmol/l Tris, pH 7.5; 150 mmol/l NaCl; 0.1% Tween-20) at room
temperature for 1 h. The membranes were subsequently incubated with
antibodies (rabbit anti-human, polyclonal antibodies, 1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA) against Erk1/2 MAPK
(cat. no. 4695), pErk1/2 MAPK (cat. no. 9101), poly (ADP-ribose)
polymerase (PARP; cat. no. 9542), cleaved PARP (cat. no. 5625),
caspase-7 (cat. no. 9492), cleaved caspase-7 (cat. no. 8438) and
GAPDH (cat. no. 5174) at 4°C overnight and then with a secondary
polyclonal rabbit antibody (goat anti-rabbit; 1:3,000; cat. no.
TA130015; OriGene Technologies, Inc., Beijing, China) for 1 h at
room temperature. Finally, a chemiluminescent agent (Thermo Fisher
Scientific, Inc.) was added for detection. ImageJ 1.41 software
(National Institutes of Health, Bethesda, MD, USA) was used for
densitometric analysis.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from cultured cells using
TRIzol® Reagent (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocols. First-strand cDNA was
synthetized by RT using the GoScript™ Reverse System Kit according
to the manufacturer's protocols (Promega Corporation, Madison, WI,
USA). Subsequently, the reverse transcribed single-stranded cDNA
was amplified by SYBR1 Premix Ex Taq II (Takara Bio, Inc.) on an
ABI Prism 7500 HT sequence detection system (Applied Biosystems;
Thermo Fisher Scientific, Inc.). Each 20-µl PCR reaction
mixture contained 1 µl cDNA product, 0.8 µl specific
forward/universal primer mix, and 10 µl SYBR-Green 2X
Universal PCR Master Mix (cat. no. Q141-02; Vazyme, Piscataway, NJ,
USA). The thermocycling conditions for qPCR comprised a
pre-amplification step of denaturation followed by annealing and
extension, and the specific conditions were as follows: 10 min at
95°C, followed by 40 cycles of 95°C for 3 sec, 60°C for 30 sec, and
95°C for 15 sec. Each sample was tested in triplicate. The raw data
were normalized to the data of β-actin and are presented as the
relative expression of lncNEAT1_2 or lncPINT. Data analysis was
performed using the 2−ΔΔCq method (25). The primers employed in the present
study were as follows: NEAT1_2 forward, 5′-GGCCAGAGCTTTGTT
GCTTC-3′, reverse, 5′-GGTGCGGGCACTTACTTACT-3′, PINT forward,
5′-GAACGAGGCAAGGAGCTAAA-3′, reverse, 5′-AGCAAGGCAGAGAAACTCCA-3′;
and β-actin forward, 5′-CGTGACATTAAGGAGAAGCTG-3′, and reverse,
5′-CTAGAAGCATTTGCGGTGGAC-3′.
Cell cycle analysis
Cells were cultured in cell culture medium
containing 10 µM Solamargine, or for the control group,
cells were treated with DMSO for 6 or 16 h, at 37°C and 5%
CO2. The cells were collected, washed and re-suspended
in PBS following trypsinization by 0.25% trypsin for 2 min at room
temperature. and fixed in 70% ethanol at -20°C overnight.
Subsequently, the samples were washed with PBS and then incubated
with propidium iodide (PI) staining buffer (Dojindo Molecular
Technologies, Inc) at room temperature for 15 min prior to analysis
with a flow cytometer (BD Pharmingen; BD Biosciences, San Jose, CA,
US) using Modfit LT 4.1 software (Verity Software House, Inc.,
Topsham, ME, USA).
Annexin V-fluorescein isothiocyanate
(FITC)/PI apoptosis assay
Cells were cultured in medium containing Solamargine
(for the control, equivalent volume of DMSO) for 48 h, the cells
were then collected and washed with PBS (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C. The cells were re-suspended in Annexin V
Binding Solution, followed by the addition of Annexin V-FITC and PI
solution (Annexin V-FITC Apoptosis Detection Kit, Dojindo Molecular
Technologies, Inc.) for 20 min in the dark at room temperature.
Finally, Annexin V Binding solution was added and cells were
analyzed with BD Accuri™ C6 software by flow cytometry (BD
Biosciences). Quantification of the flow cytometry results
demonstrated the fractions of cells in the early and late stage of
apoptosis.
Transient transfection assay
LncNEAT1_2-target small interfering (si)RNA
(5′-GUCUGUGUGGAAGGAGGA ATT-3′) and nonspecific scrambled siRNA
(5′-UUCUCCGAA CGUGUCACGUTT-3′) were designed and synthesized by
Shanghai GenePharma Co., Ltd. (Shanghai, China). Cells (SGC 7901
and BGC 823) were employed according to manufacturer's protocols; 5
µl siRNA or negative control siRNA, and 6 µl
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) were used for transfection. After 6 h, the
transfection solution was discarded and cell culture medium
containing serum was added; cells were cultured for 24 h prior to
the subsequent experiments. To evaluate the efficiency of
transfection, the expression of lncNEAT1_2 was investigated by
RT-qPCR analysis as aforementioned.
Tumor xenograft
The present study was performed in strict accordance
with the recommendations of the Guidelines for the Care and Use of
Laboratory Animals of the Peking University Cancer Hospital and
Institute (26). Female specific
pathogen-free BALB/c nude mice weighing 18-20 g (6-8-weeks-old)
were employed and housed under 21±2°C, 50-60% humidity with normal
air and with a 12 h light/dark cycle. Mice were provided 5 g food
and 6-8ml water/mouse/day. A total of 10 experimental mice were
randomly divided into two groups (5 per group). BCG 823 is mostly
used to establish tumor xenograft for research in our
center. Mice of each group were administered a subcutaneous
injection of BGC823 GC cells (5×105 cells per mouse)
into the left hind leg. After 10 days, the experimental group was
treated with 10 mg/kg Solamargine once daily by intragastric
administration, while the control group was administered PBS for 8
days corresponding to the time for the tumors in control group to
reach about 600-800 mm3. Tumor growth was monitored
every 2 days (total 18 days) by measuring the width (b) and length
(a) of the tumors with calipers. The tumor volume (V) was
calculated by the formula: V=l/2 a x b2. Mice were sacrificed for
the collection of tumor samples.
Terminal deoxynucleotidyl
transferase-mediated dUTP-biotin nick end labeling (TUNEL)
assay
Paraffin-embedded mouse tumor tissues fixed with 10%
formalin solution for 12 h were cut and mounted on slides (4
µm section) at room temperature. First, the tissue sections
were dewaxed and rehydrated in a descending alcohol series.
Subsequently, the slides were treated In Situ Cell Death Detection
kit, TMR red (Roche Applied Science, Rotkreuz, Switzerland)
according to the manufacturer's protocols. Briefly, the slides were
treated with 0.1 M citrate buffer, pH 6.0 with microwave
irradiation (750 watts) for 1 min. Then, the slides were immersed
in 0.1 M Tris-HCl (pH 7.5) containing 3% BSA and 20% normal bovine
serum (Gibco; Thermo Fisher Scientific, Inc.) at room temperature
for 30 min. In addition, 50 µl TUNEL reaction mixture per
section was added to cover the slides and incubated for another 60
min at >37°C in a humidified atmosphere in the dark. Finally,
the slides were counterstained for 6 min with DAPI at room
temperature (Guangzhou Ribobio, Co., Ltd., Guangzhou, China). The
sections were evaluated under a fluorescence microscope
(magnification, ×63, Lm780, Zeiss AG, Oberkochen, Germany).
Apoptotic cells were quantified by measuring the average cell
numbers in five random high-power fields of each of sample for
analysis.
Haematoxylin and eosin (H&E)
staining
Paraffin-embedded mouse tumor tissues were fixed in
10% formalin solution for 12 h at room temperature were cut and
mounted on slides (4 µm thick). The tissue sections were
dewaxed and rehydrated in a descending alcohol series.
Subsequently, the slides were stained with hematoxylin stain for 1
min at room temperature. Subsequently, the samples were washed in
water for 10 min and after differentiation in acid alcohol, the
slides were stained with eosin for 1 min at room temperature.
High-quality images were obtained using by Aperio CS2 image capture
device (magnification, ×5 and 20). Necrotic areas were quantified
(Aperio ImageScope software, version 12.1, Aperio Technologies;
Leica Microsystems, Inc., Buffalo Grove, IL, USA) by measuring the
necrotic area (%) in three slides of each sample for analysis.
Statistical analysis
Every experiment was repeated at least three times,
and the data were expressed as the mean ± standard deviation. The
differences between groups were assessed by a Student's t-test or
one-way analysis of variance followed by Tukey's post-hoc test. All
statistical analyses were performed using SPSS software (version
22.0, IBM Corp., Armonk, NY, USA) and GraphPad Prism statistical
software (version 5.0). Comparisons were conducted with the
corresponding controls. P<0.05 was considered to indicate a
statistically significant difference.
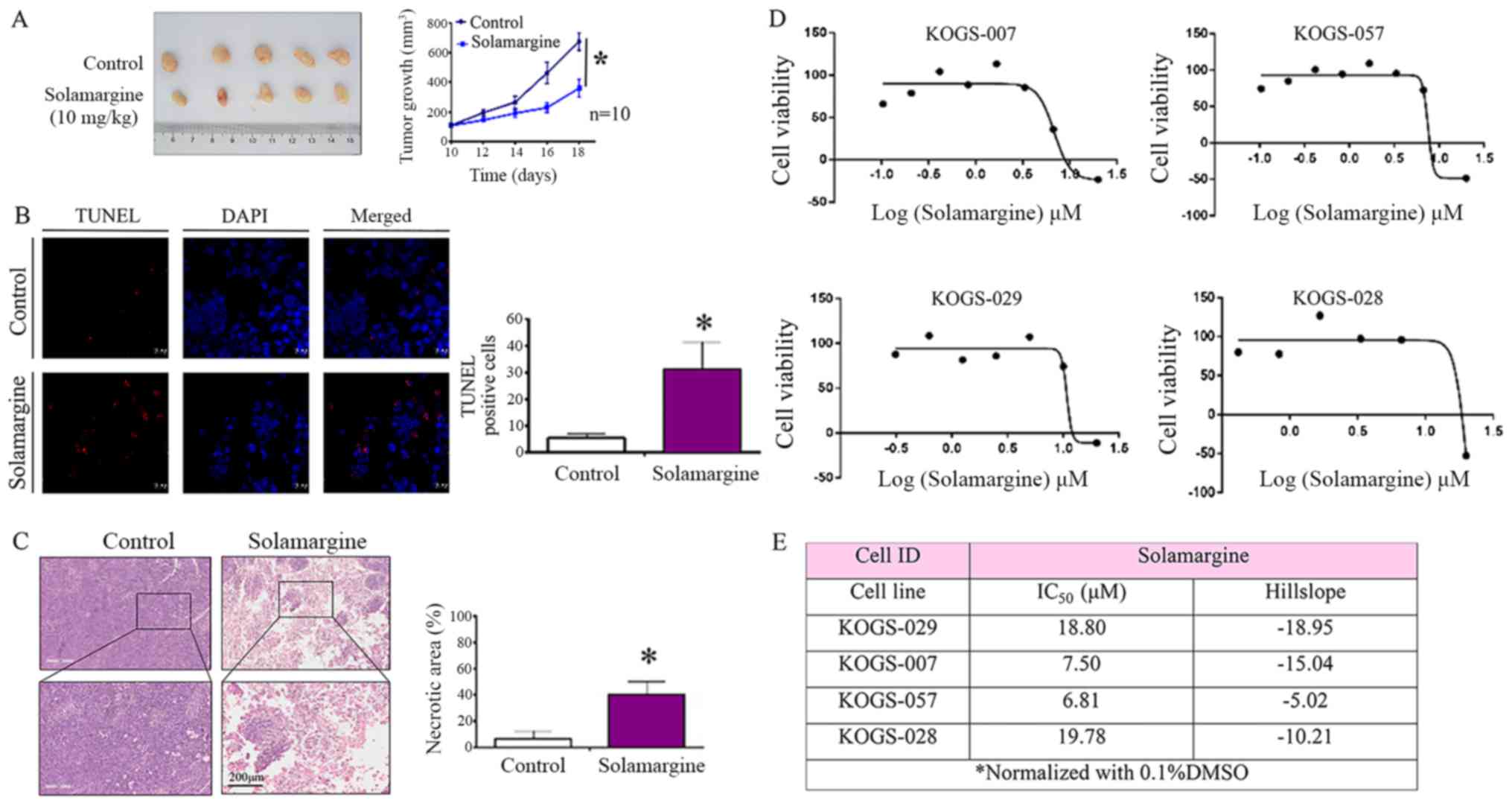
Results
Solamargine suppresses the viability of
GC cells and induces alterations in morphology
Notable morphological changes were observed in GC
cell lines cultured with Solamargine compared with in the control
(Fig. 1A). The volume of cells was
reduced, the intercellular junctions were not visible, the nuclei
were concentrated; few cells were not firmly adhered and large
amounts of cell debris were visible. To verify the effects of
Solamargine on viability, the IncuCyte ZOOM Live-Cell Analysis
System which can automatically acquire images of living cells and
analysis the confluence based on the image to reflect the cell
viability was employed. The results of the present study revealed
that Solamargine suppressed cell viability in five GC cell lines
(AGS, BGC823, SGC7901, HGC27 and MGC803) in a dose-dependent manner
and IC50 values are presented in Fig. 1B. Compared with in the control
group, the viability of GC cells in the Solamargine group gradually
decreased with increasing drug concentrations (P<0.05).
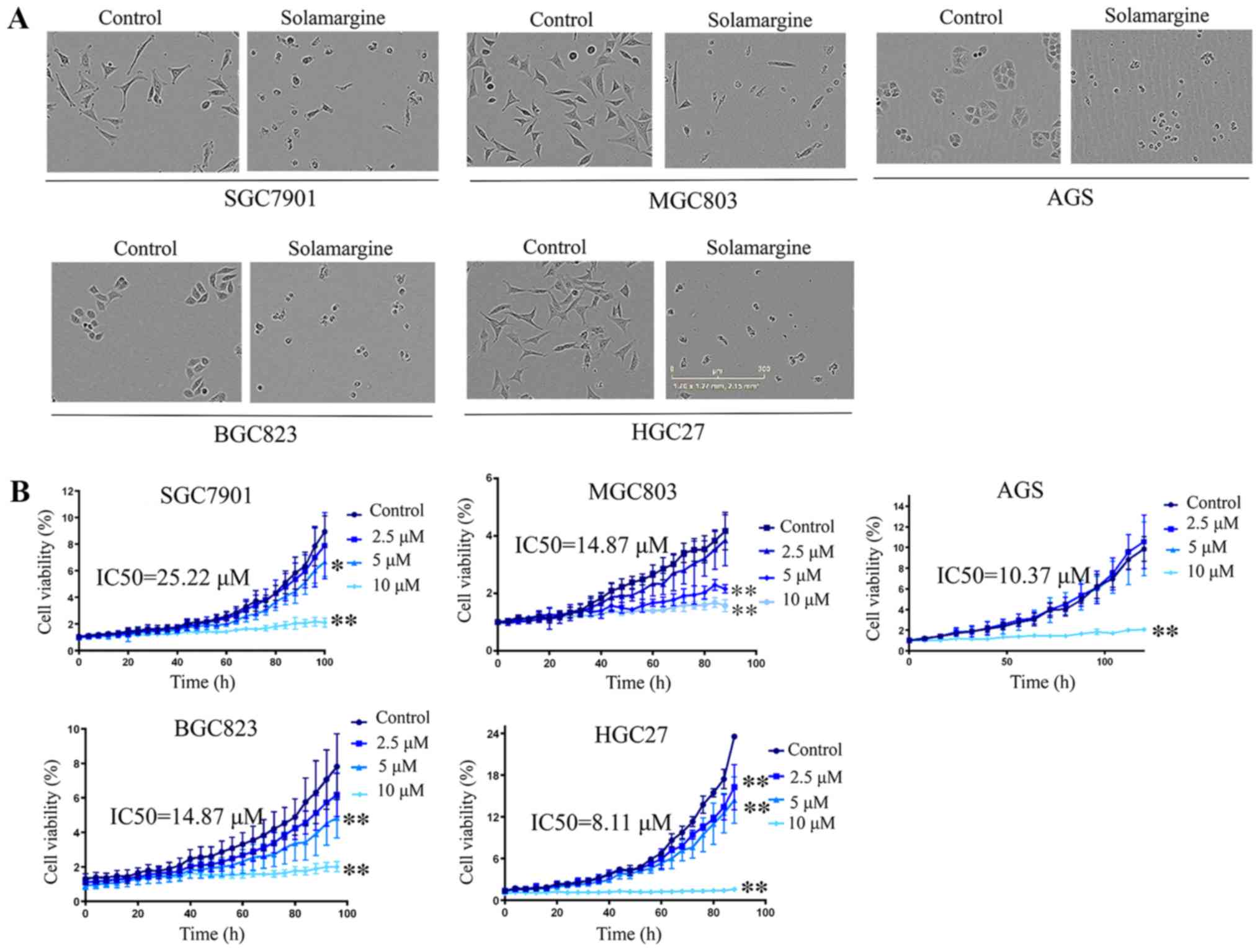 | Figure 1Effects of Solamargine on the
morphology and viability of GC cells. (A) Effects of Solamargine on
cellular morphology. GC cell lines (AGS, BGC823, SGC7901, HGC27 and
MGC803) were treated with Solamargine (10 µM) for 48 h.
Magnification, ×10. (B) Solamargine decreased the viability of GC
cells. GC cell lines (AGS, BGC823, SGC7901, HGC27, and MGC803) were
treated with different concentrations of Solamargine (0, 2.5, 5 and
10 µM). Subsequently, the cell viability was determined with
the IncuCyte ZOOM Live-Cell Analysis System every 8 h.
*P<0.05, **P<0.01 vs. control. The
IC50 for 6 GC cell lines were calculated by GraphPad
Prism statistical software (version 5.0). GC, gastric cancer;
IC50, half-maximal inhibitory concentration. |
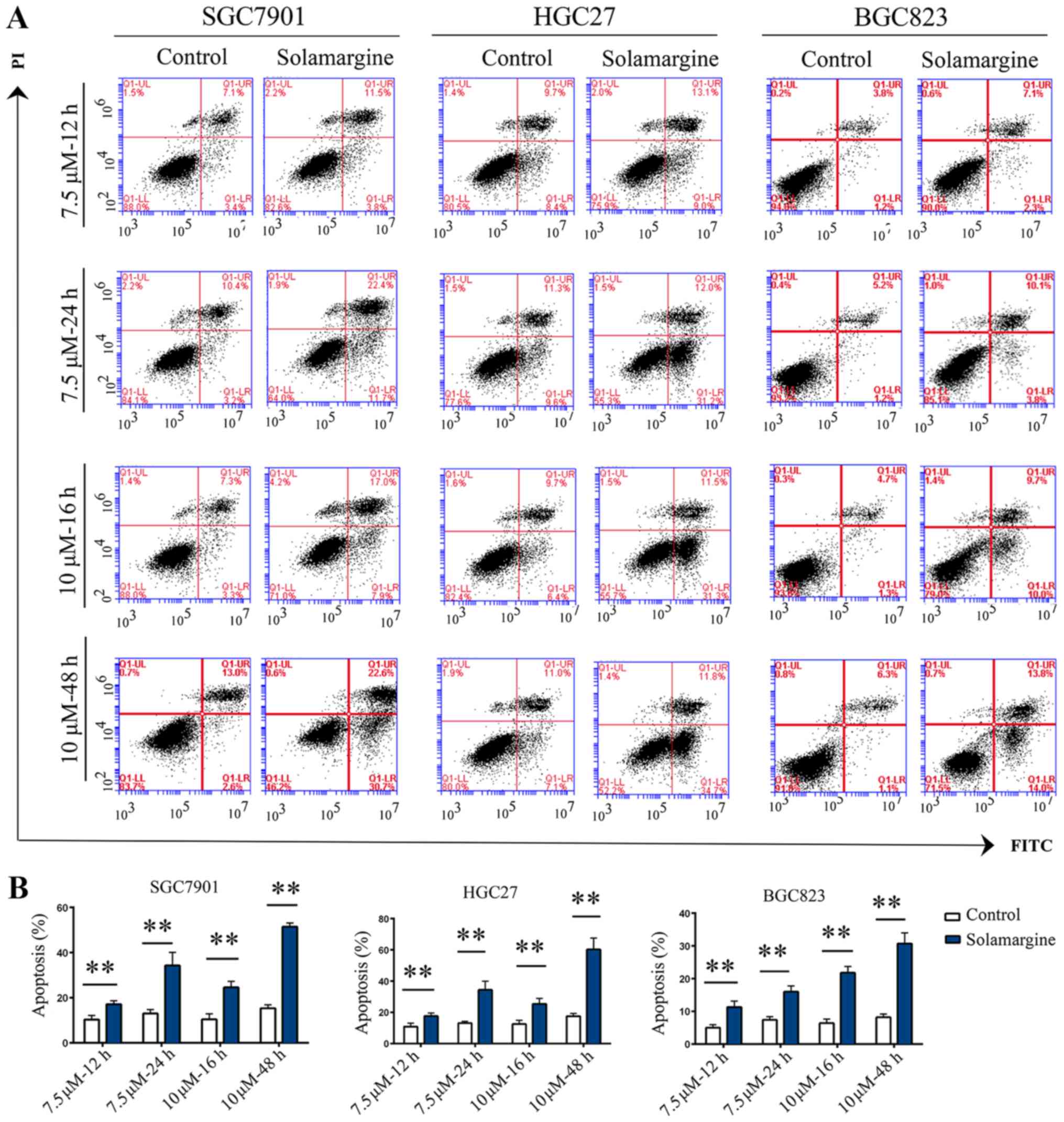
Solamargine promotes the apoptosis of GC
cells
Based on the inhibitory effects and alterations in
morphology induced by Solamargine, the apoptotic effect of
Solamargine on GC cells was analyzed by Annexin V-FITC/PI
double-staining and western blotting. The results demonstrated that
the rate of apoptosis in the Solamargine group was significantly
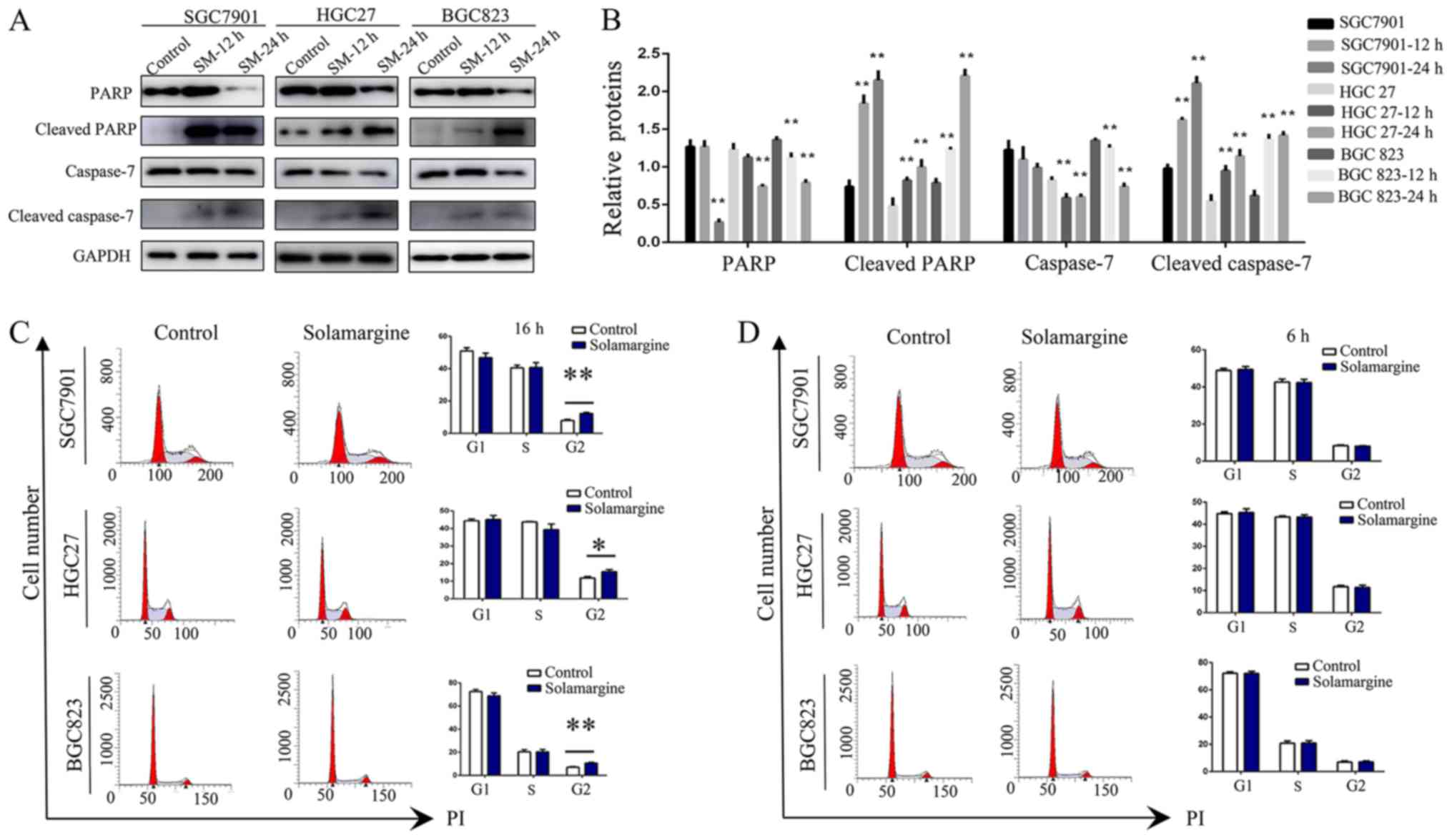
higher than that of the control group (Fig. 2). Considering that caspase
activation is the key event for apoptosis (27,28),
the expression levels of full-length and cleaved caspase proteins
were measured by western blotting (Fig. 3A and B). Treatment with Solamargine
(7.5 µM, 12 or 24 h) significantly enhanced the cleavage of
caspase-7 and PARP compared with in the corresponding controls,
indicating that Solamargine may promote the apoptosis of GC
cells.
Solamargine increases the proportion of
GC cells in G2/M phase
In addition, the effects of Solamargine on the cell
cycle of GC cells was investigated. The results revealed a
significant increase in the number of in SGC7901, HGC27 and BGC823
cells in G2/M phase treated with Solamargine (10 µM) for 16
h compared with in the control (Fig.
3C). Additionally, to exclude interference by other factors,
alterations resulting from 6 h treatment with Solamargine in the
corresponding GC cell lines were analyzed. The results revealed no
significant difference in the number of GC cells exposed to
Solamargine and the corresponding control at this time point
(Fig. 3D). In summary, the present
study reported that Solamargine increased the number of GC cells in
G2/M phase.
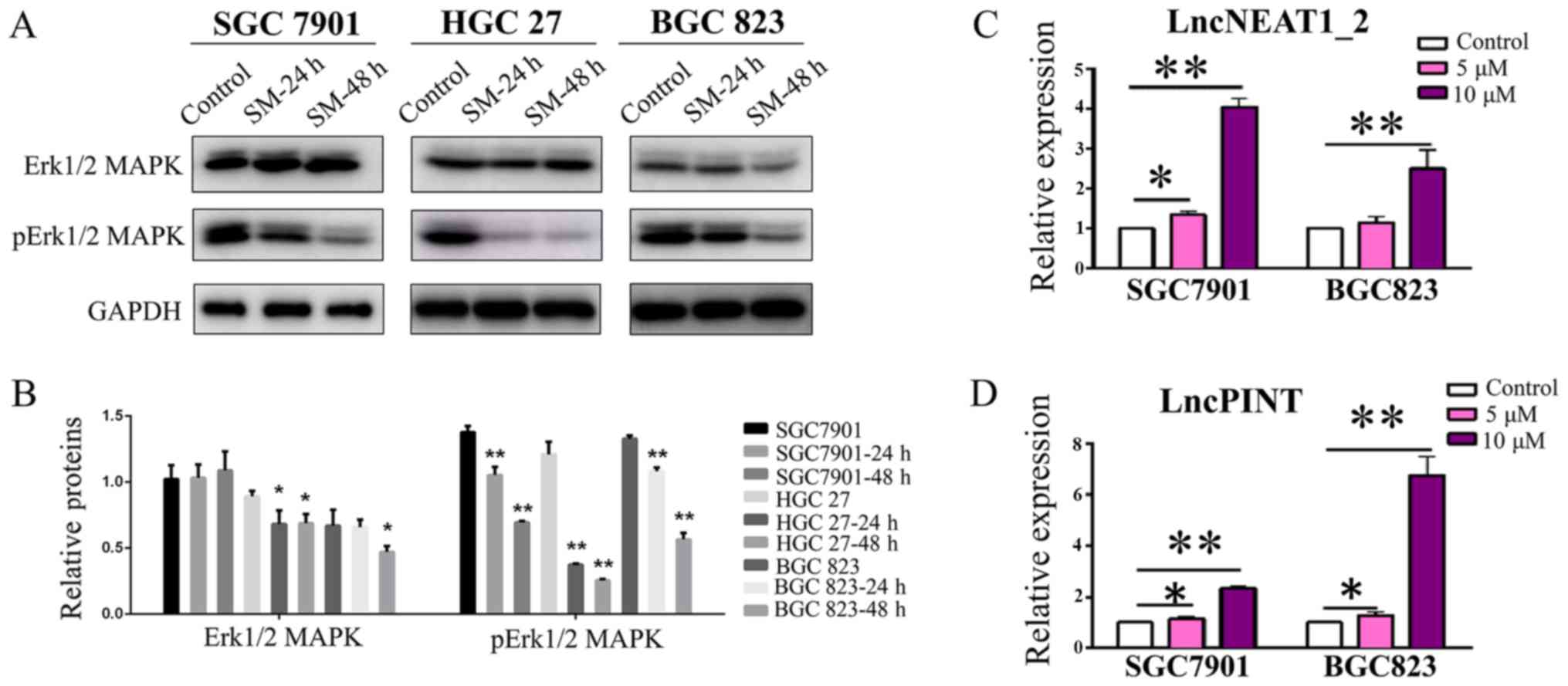
Solamargine inhibits the phosphorylation
of Erk1/2 MAPK
The MAPK signaling pathway, an important signaling
pathway within eukaryotic cells, serves a key role in the
development of tumors and has been reported as a novel target for
tumor therapy (17-19). The present study investigated the
effects of Solamargine on MAPK activity. As presented in Fig. 4A and B, the expression levels of
Erk1/2 and p-Erk1/2 MAPK were significantly reduced in the cell
lines in a time-dependent manner following Solamargine treatment,
compared with in the corresponding control; however, no
significance was observed in the expression levels of Erk1/2 MAPK
in SGC7901 cells. These findings indicated that the MAPK signaling
pathway is involved in the regulatory effects of Solamargine.
Solamargine increases the expression of
lncPINT and lncNEAT1_2
As aforementioned, numerous studies have reported
lncPINT and lncNEAT1_2 to be closely associated with MAPKs
(29,30). Based on the findings of the present
study that Solamargine inhibited the phosphorylation of MAPKs,
whether the expression of lncPINT and lncNEAT1_2 may be affected
was investigated. The results demonstrated that the expression
levels of lncPINT and lncNEAT1_2 were significantly increased in
the Solamargine-treated group compared with in the control group of
SGC7901 cells (5 µM, P<0.05; 10 µM, P<0.01;
Fig. 4C and D). Additionally, the
upregulated expression levels appeared to be dose-dependent;
however, treatment with 5 µM Solamargine revealed a
non-significant increase in the expression of lncNEAT1_2 in BGC823
cells. According to the findings of the present study, Solamargine
inhibited the phosphorylation of MAPKs and upregulated the
expression of lncPINT and lncNEAT1_2 in a dose-dependent
manner.
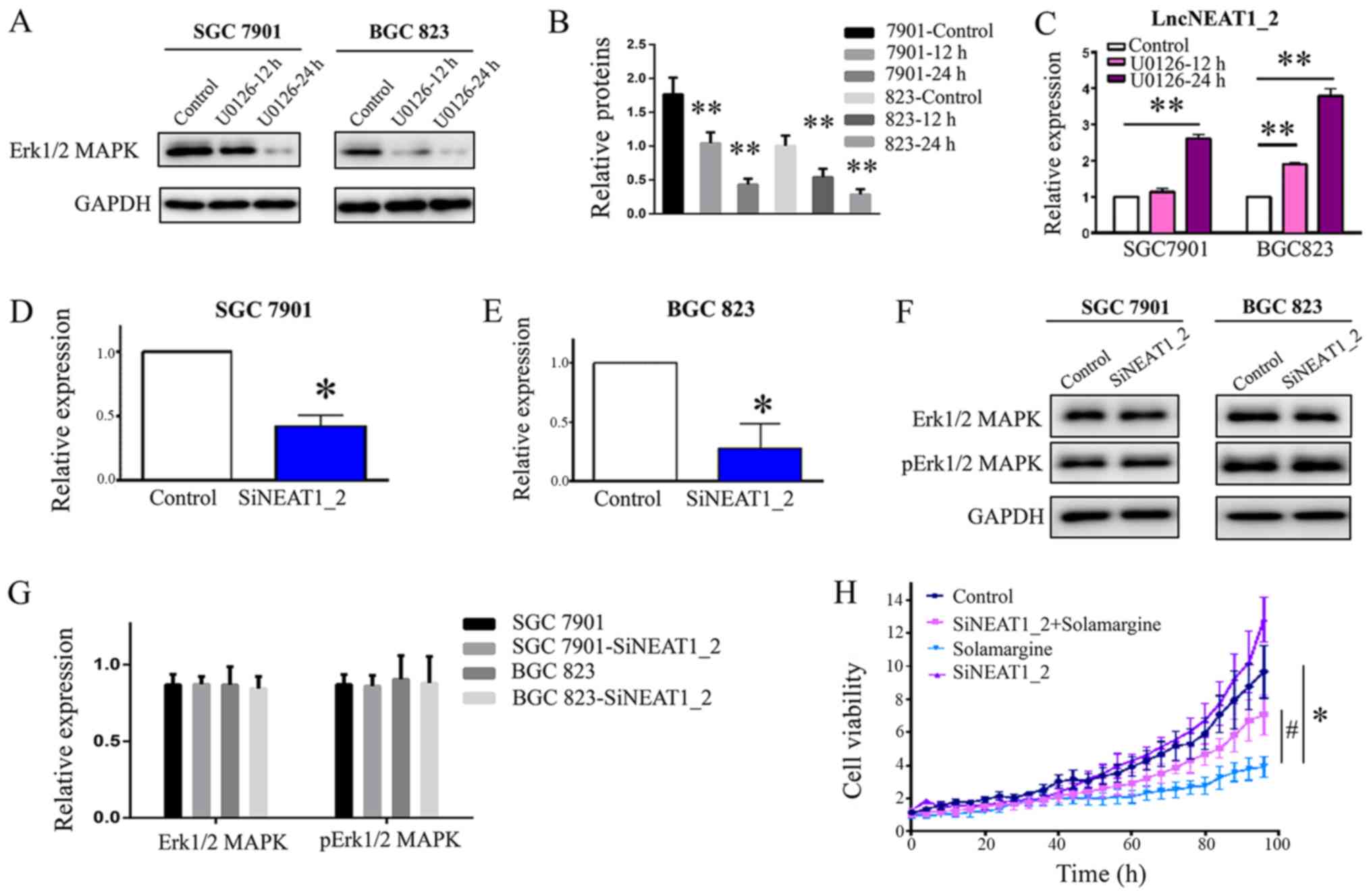
Inhibition of Erk1/2 MAPK increases the
expression of lncNEAT1_2
The present study demonstrated that Solamargine
reduced the phosphorylation of Erk1/2 MAPK and increased the
expression of lncNEAT1_2. LncPINT and one splice variant of
lncNEAT1 were reported to be regulated in an Erk-dependent manner
(20). To understand the potential
mechanism by which Solamargine affects GC, the interactions between
Erk1/2 MAPK and lncNEAT1_2 were investigated. The results revealed
that treatment with an exogenous inhibitor of Erk1/2 MAPK (U0126)
reduced the expression of Erk1/2 MAPK and significantly enhanced
the expression of lncNEAT1_2 in a time-dependent manner (Fig. 5A-C), which was consistent with the
effects of Solamargine. Knockdown of lncNEAT1_2 (Fig. 5D-G) had no notable effect on the
expression of Erk1/2 MAPK or its phosphorylation. Collectively, the
results suggested that Erk1/2 MAPK is upstream of lncNEAT1_2 and
that Solamargine increased the expression of lncNEAT1_2 via the
inhibition of Erk1/2 MAPK signaling.
Knockdown of lncNEAT1_2 attenuates the
inhibitory effects of Solamargine on GC cells
The aforementioned findings indicated that
Solamargine regulated the expression of lncPINT and lncNEAT1_2.
Furthermore, to confirm the observed inhibition, the effects of
Solamargine on GC cell viability were investigated by
downregulating the expression of lncNEAT1_2 in GC cells; the
IncuCyte ZOOM Live-Cell Analysis System was employed. The results
demonstrated that knockdown of lncNEAT1_2 significantly attenuated
the inhibitory effects of Solamargine on GC cells (Fig. 5H). The results suggested that
lncNEAT1_2 is a potentially important therapeutic target for
Solamargine.
In vivo antitumor activity of
Solamargine
To further assess the activity of Solamargine in
vivo, alterations in tumor growth in response to Solamargine
were investigated within a tumor xenograft model. As presented in
Fig. 6A, the tumor volume in the
Solamargine group was significantly reduced compared with in the
control group. Based on the present study, Solamargine served a
crucial role in promoting the apoptosis in GC cells. A TUNEL assay
and H&E staining were performed using excised tumor sections.
The results revealed a significant increase in the number of early
apoptotic cells in the Solamargine group (Fig. 6B). Furthermore, the results of
H&E staining suggested that the necrotic area in the
Solamargine group was significantly increased compared with in the
control (Fig. 6C). In summary, the
results indicated that Solamargine exhibited an antitumor effect
and promoted the apoptosis of GC in vivo.
Solamargine sensitivity of primary GC
cells from patients
The cell lines employed in the present study
exhibited cancer-associated characteristics without the
heterogeneity observed in primary tumors (31). To study the responsiveness of
patients to Solamargine treatment, cell viability assays were
performed on primary GC cells. Primary cells from four patients who
underwent radical gastrectomy were treated with appropriate amounts
of Solamargine for over 72 h. A drug sensitivity assay was
performed and Solamargine was observed to suppress the viability of
GC cells from all four patients (Fig.
6D); the IC50 values were determined (Fig. 6E). The result of the present study
indicated that the effects of Solamargine on primary GC cells
obtained from patients were similar to that of GC cell lines.
Discussion
A previous study reported that Solamargine exhibited
an inhibitory effect on a variety of tumor cell lines; however, the
role of Solamargine in GC cells requires further investigation
(9,32-34).
Studies of Solamargine with in other cancer types demonstrated
variations in the IC50 values from 0.91 to 26.66
µM (32). The present study
observed a significant inhibitory effect on GC in response to 10
µM Solamargine by using the IncuCyte ZOOM Live-Cell system.
Based on the aforementioned analysis and results, concentrations of
0-10 µM Solamargine were applied in the present study. To
the best of our knowledge, the present study is the first to report
that Solamargine induced the apoptosis and suppressed the viability
of GC cell lines.
A few molecular pathways can inf luence the
anti-tumor activity of Solamargine, including the tumor necrosis
factor receptor (35), B-cell
lymphoma 2 (Bcl-2)/Bcl-2-associated X (36) and MAPK signaling pathways (16,33).
MAPKs are a widely conserved family of serine/threonine protein
kinases (37). Previously, studies
have reported numerous biological functions of cells to be
regulated by the MAPK signaling pathway, including proliferation,
apoptosis and metastasis (15,38-42).
In addition, its activation was closely associated with
carcinogenesis and tumor development (43). Targeting the members of the MAPK
family was therefore a prospective approach to the management of
incurable GC. The present study demonstrated that Solamargine
inhibited GC by suppressing the phosphorylation of Erk1/2, one of
the key members of the MAPK family, in a dose-dependent manner. The
results of the present study suggested that the MAPK network is a
crucial signaling pathway involved in the inhibitory effects of
Solamargine on GC.
LncNEAT1 is expressed as two splicing variants that
differ in length, namely, NEAT1_1 and NEAT1_2 (23). A previous study reported that the
two splicing variants exhibited differing effects on tumor
progression (44). Interestingly,
by using the murine hippocampus as a model, Blüthgen et al
(45) revealed that the induction
of lncPINT and the induction of one splicing variant of lncNEAT1
were Erk-dependent (46,47). MAPK is an important signaling
pathway within living organisms in response to extracellular
stimuli (31). The present study
proposed that lncPINT and lncNEAT1 were Erk-dependent in GC as
reported in murine hippocampus (46). Based on the reports that
Solamargine inhibited the MAPK pathway, the roles of lncPINT and
lncNEAT1_2 were investigated in the present study to determine the
potential mechanism underlying the inhibitory effects of
Solamargine. The results demonstrated that Solamargine inhibited
the phosphorylation of Erk1/2 MAPK in GC and increased the
expression of lncPINT and lncNEAT1_2. In addition, downregulating
the expression of lncNEAT1_2 significantly reduced the inhibitory
effects of Solamargine on GC. To the best of our knowledge, the
present study is the first to reveal that Solamargine suppressed
the viability of GC cells mainly via the overexpression of lncPINT
and lncNEAT1_2. Thus, a novel strategy for investigating the
mechanism of Solamargine and insight into clinical anticancer
treatment against GC were reported; however to understand the
mechanism of this regulatory axis, further investigation is
required.
At present, based on the scientific evidence
available, Solamargine may be considered as a promising treatment
against cancer; however, its anticancer potential in vivo
and within patients remains unclear (11,12,48).
In the present study, the effects of Solamargine on GC were also
observed in vivo via nude mice and in primary cells from
patients. The present study reported that experiments in
vivo and in primary cells from patients also suggested that
Solamargine exerted an inhibitory effect on tumor growth in GC. In
future studies, other modes of administration with increased
efficiency, such as intravenous administration, may also be
studied. In addition, patient-derived tumor xenografts can be
employed to verify the effects of Solamargine, and ultimately,
clinical trials may be performed to assess its efficacy.
Collectively, the results of the present study
suggest that Solamargine exerted an inhibitory effect on GC and
that this effect was achieved by inducing the expression of lncPINT
and lncNEAT1_2 via the inhibition of Erk1/2 MAPK phosphorylation.
Therefore, Solamargine may be considered as a potential therapeutic
agent for the treatment of GC and may be applied in future clinical
studies.
Funding
This study was supported by the 'San Ming' Project
(grant no. SZSM201612051, Shenzhen, China).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding authors on reasonable
request.
Authors' contributions
JJ contributed to the design of the study. LiaZ and
GL wrote the manuscript. RF performed the experiments. XW analyzed
the data. YH, HD, BD, SA, LiZ and ZS provided the Solamargine and
technical assistance. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The collection of primary GC cells from patients who
underwent radical gastrectomy for the present study was approved by
the Ethics Committee of Peking University Cancer Hospital and
Institute (Beijing, China). Informed consent was provided by all
patients. The animal study was approved by Peking University Cancer
Hospital Research Ethics Committee (Beijing, China) and was
conducted in strict accordance with the Guidelines for the Care and
Use of Laboratory Animals of the Peking University Cancer Hospital
and Institute (26).
Patient consent for publication
Not applicable.
Competing interests
ZS has ownership interest (including patents) in K2
Oncology.
Acknowledgments
Not applicable.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xi J, Feng J, Li Q, Li X and Zeng S: The
long non-coding RNA lncFOXO1 suppresses growth of human breast
cancer cells through association with BAP1. Int J Oncol.
50:1663–1670. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xu Y, Zheng Y, Liu H and Li T: Modulation
of IGF2BP1 by long non-coding RNA HCG11 suppresses apoptosis of
hepato-cellular carcinoma cells via MAPK signaling transduction.
Int J Oncol. 51:791–800. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang R and Xia T: Long non-coding RNA
XIST regulates PDCD4 expression by interacting with miR-21-5p and
inhibits osteosarcoma cell growth and metastasis. Int J Oncol.
51:1460–1470. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Burger T, Mokoka T, Fouché G, Steenkamp P,
Steenkamp V and Cordier W: Solamargine, a bioactive steroidal
alkaloid isolated from Solanum aculeastrum induces non-selective
cytotoxicity and P-glycoprotein inhibition. BMC Complement Altern
Med. 18:1372018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Marín-Béjar O, Mas AM, González J,
Martinez D, Athie A, Morales X, Galduroz M, Raimondi I, Grossi E,
Guo S, et al: The human lncRNA LINC-PINT inhibits tumor cell
invasion through a highly conserved sequence element. Genome Biol.
18:2022017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen Y, Tang Q, Wu J, Zheng F, Yang L and
Hann SS: Inactivation of PI3-K/Akt and reduction of SP1 and p65
expression increase the effect of solamargine on suppressing EP4
expression in human lung cancer cells. J Exp Clin Cancer Res.
34:1542015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ding X, Zhu FS, Li M and Gao SG: Induction
of apoptosis in human hepatoma SMMC-7721 cells by solamargine from
Solanum nigrum L. J Ethnopharmacol. 139:599–604. 2012. View Article : Google Scholar
|
|
10
|
Sun L, Zhao Y, Li X, Yuan H, Cheng A and
Lou H: A lysosomal-mitochondrial death pathway is induced by
sola-margine in human K562 leukemia cells. Toxicol In Vitro.
24:1504–1511. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xie X, Zhu H, Zhang J, Wang M, Zhu L, Guo
Z, Shen W and Wang D: Solamargine inhibits the migration and
invasion of HepG2 cells by blocking epithelial-to-mesenchymal
transition. Oncol Lett. 14:447–452. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shiu LY, Chang LC, Liang CH, Huang YS,
Sheu HM and Kuo KW: Solamargine induces apoptosis and sensitizes
breast cancer cells to cisplatin. Food Chem Toxicol. 45:2155–2164.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xiang S, Zhang Q, Tang Q, Zheng F, Wu J,
Yang L and Hann SS: Activation of AMPKα mediates additive effects
of solamargine and metformin on suppressing MUC1 expression in
castration-resistant prostate cancer cells. Sci Rep. 6:367212016.
View Article : Google Scholar
|
|
14
|
Wu YH, Chiu WT, Young MJ, Chang TH, Huang
YF and Chou CY: Solanum Incanum Extract Downregulates Aldehyde
Dehydrogenase 1-Mediated Stemness and Inhibits Tumor Formation in
Ovarian Cancer Cells. J Cancer. 6:1011–1019. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cui CZ, Wen XS, Cui M, Gao J, Sun B and
Lou HX: Synthesis of solasodine glycoside derivatives and
evaluation of their cytotoxic effects on human cancer cells. Drug
Discov Ther. 6:9–17. 2012.PubMed/NCBI
|
|
16
|
Zhou Y, Tang Q, Zhao S, Zhang F, Li L, Wu
W, Wang Z and Hann S: Targeting signal transducer and activator of
transcription 3 contributes to the solamargine-inhibited growth and
-induced apoptosis of human lung cancer cells. Tumour Biol.
35:8169–8178. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen Y, Tang Q, Xiao Q, Yang L and Hann
SS: Targeting EP4 downstream c-Jun through ERK1/2-mediated
reduction of DNMT1 reveals novel mechanism of solamargine-inhibited
growth of lung cancer cells. J Cell Mol Med. 21:222–233. 2017.
View Article : Google Scholar
|
|
18
|
Schmitt AM and Chang HY: Long Noncoding
RNAs in Cancer Pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li L, Zhang GQ, Chen H, Zhao ZJ, Chen HZ,
Liu H, Wang G, Jia YH, Pan SH, Kong R, et al: Plasma and tumor
levels of Linc-pint are diagnostic and prognostic biomarkers for
pancreatic cancer. Oncotarget. 7:71773–71781. 2016.PubMed/NCBI
|
|
20
|
Yang X, Qu S, Wang L, Zhang H, Yang Z,
Wang J, Dai B, Tao K, Shang R, Liu Z, et al: PTBP3 splicing factor
promotes hepatocellular carcinoma by destroying the splicing
balance of NEAT1 and pre-miR-612. Oncogene. 37:6399–6413. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang H, Cai Y, Zheng L, Zhang Z, Lin X
and Jiang N: Long noncoding RNA NEAT1 regulate papillary thyroid
cancer progression by modulating miR-129-5p/KLK7 1expression. J
Cell Physiol. 233:6638–6648. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu B and Shan G: Functions of long
noncoding RNAs in the nucleus. Nucleus. 7:155–166. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu X, Li Z, Zheng H, Chan MT and Wu WK:
NEAT1: A novel cancer-related long non-coding RNA. Cell Prolif.
50:502017.
|
|
24
|
Mello SS, Sinow C, Raj N, Mazur PK,
Bieging-Rolett K, Broz DK, Imam JFC, Vogel H, Wood LD, Sage J, et
al: Neat1 is a p53-inducible lincRNA essential for transformation
suppression. Genes Dev. 31:1095–1108. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
26
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals. 8th
edition. National Academies Press (US); Washington, DC: 2011
|
|
27
|
Julien O and Wells JA: Caspases and their
substrates. Cell Death Differ. 24:1380–1389. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Choi EO, Park C, Hwang HJ, Hong SH, Kim
GY, Cho EJ, Kim WJ and Choi YH: Baicalein induces apoptosis via
ROS-dependent activation of caspases in human bladder cancer 5637
cells. Int J Oncol. 49:1009–1018. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pedersen AE, Bregenholt S, Johansen B,
Skov S and Claesson MH: MHC-I-induced apoptosis in human B-lymphoma
cells is dependent on protein tyrosine and serine/threonine
kinases. Exp Cell Res. 251:128–134. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cross TG, Scheel-Toellner D, Henriquez NV,
Deacon E, Salmon M and Lord JM: Serine/threonine protein kinases
and apoptosis. Exp Cell Res. 256:34–41. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Correa BRS, Hu J, Penalva LOF, Schlegel R,
Rimm DL, Galante PAF and Agarwal S: Patient-derived conditionally
reprogrammed cells maintain intra-tumor genetic heterogeneity. Sci
Rep. 8:40972018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kalalinia F and Karimi-Sani I: Anticancer
Properties of Solamargine: A Systematic Review. Phytother Res.
31:858–870. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Al Sinani SS, Eltayeb EA, Coomber BL and
Adham SA: Solamargine triggers cellular necrosis selectively in
different types of human melanoma cancer cells through extrinsic
lysosomal mitochondrial death pathway. Cancer Cell Int. 16:112016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Del Bufalo D, Biroccio A, Trisciuoglio D,
Bruno T, Floridi A, Aquino A and Zupi G: Bcl-2 has differing
effects on the sensitivity of breast cancer cells depending on the
antineoplastic drug used. Eur J Cancer. 38:2455–2462. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hsu SH, Tsai TR, Lin CN, Yen MH and Kuo
KW: Solamargine purified from Solanum incanum Chinese herb triggers
gene expression of human TNFR I which may lead to cell apoptosis.
Biochem Biophys Res Commun. 229:1–5. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xie X, Zhu H, Yang H, Huang W, Wu Y, Wang
Y, Luo Y, Wang D and Shao G: Solamargine triggers hepatoma cell
death through apoptosis. Oncol Lett. 10:168–174. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Morrison DK and Davis RJ: Regulation of
MAP kinase signaling modules by scaffold proteins in mammals. Annu
Rev Cell Dev Biol. 19:91–118. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kyriakis JM and Avruch J: Mammalian MAPK
signal transduction pathways activated by stress and inflammation:
A 10-year update. Physiol Rev. 92:689–737. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yoda Y, Takeshima H, Niwa T, Kim JG, Ando
T, Kushima R, Sugiyama T, Katai H, Noshiro H and Ushijima T:
Integrated analysis of cancer-related pathways affected by genetic
and epigenetic alterations in gastric cancer. Gastric Cancer.
18:65–76. 2015. View Article : Google Scholar
|
|
40
|
Xie J, Jin B, Li DW, Shen B, Cong N, Zhang
TZ and Dong P: ABCG2 regulated by MAPK pathways is associated with
cancer progression in laryngeal squamous cell carcinoma. Am J
Cancer Res. 4:698–709. 2014.PubMed/NCBI
|
|
41
|
Deng J, Qian Y, Geng L, Xie H, Wang Y,
Jiang G, Zhou L, Zhang M and Zheng S: Involvement of ERK and JNK
pathways in IFN-γ-induced B7-DC expression on tumor cells. J Cancer
Res Clin Oncol. 137:243–250. 2011. View Article : Google Scholar
|
|
42
|
Wang Z, Wang W, Xu S, Wang S, Tu Y, Xiong
Y, Mei J and Wang C: The role of MAPK signaling pathway in the
Her-2-positive meningiomas. Oncol Rep. 36:685–695. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Peluso I, Yarla NS, Ambra R, Pastore G and
Perry G: MAPK signalling pathway in cancers: Olive products as
cancer preventive and therapeutic agents. Semin Cancer Biol. Sep
11–2017.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wu Y, Yang L, Zhao J, Li C, Nie J, Liu F,
Zhuo C, Zheng Y, Li B, Wang Z, et al: Nuclear-enriched abundant
transcript 1 as a diagnostic and prognostic biomarker in colorectal
cancer. Mol Cancer. 14:1912015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Blüthgen N, van Bentum M, Merz B, Kuhl D
and Hermey G: Profiling the MAPK/ERK dependent and independent
activity regulated transcriptional programs in the murine
hippocampus in vivo. Sci Rep. 7:451012017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Csibi A, Fendt SM, Li C, Poulogiannis G,
Choo AY, Chapski DJ, Jeong SM, Dempsey JM, Parkhitko A, Morrison T,
et al: The mTORC1 pathway stimulates glutamine metabolism and cell
proliferation by repressing SIRT4. Cell. 153:840–854. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shukla V, Coumoul X, Wang RH, Kim HS and
Deng CX: RNA interference and inhibition of MEK-ERK signaling
prevent abnormal skeletal phenotypes in a mouse model of
craniosyn-ostosis. Nat Genet. 39:1145–1150. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Nishie K, Norred WP and Swain AP:
Pharmacology and toxicology of chaconine and tomatine. Res Commun
Chem Pathol Pharmacol. 12:657–668. 1975.PubMed/NCBI
|