Introduction
Ovarian cancer is the most lethal type of cancer
among all frequent gynecological malignancies, not only in China
but globally as well (1). The
majority of patients are diagnosed with epithelial ovarian cancer
(EOC) in the advanced stages of the disease and subsequent
platinum/taxane chemoresistance occurs following prolonged
treatment, which is responsible for the high mortality associated
with EOC (2). Despite the fact
that serum CA125 examination and ultrasonography are clinically
accepted methods for the diagnosis of ovarian cancer, these are not
satisfactory and do not fulfill the requirements for the early
detection of ovarian cancer due to poor sensitivity and specificity
(3). Cytoreductive surgery and
subsequent platinum/taxane-based post-operative adjuvant
chemotherapy have made some progress in improving the survival rate
of patients with ovarian cancer; however, eventual recurrence and
acquired chemoresistance are still inevitable. Thus, the
identification of more sensitive diagnostic biomarkers and the
establishment of novel therapeutic strategies is of utmost
importance.
Exosomes are spherical and bilayered proteolipids
with a diameter of 30–100 nm. They are abundant in various cellular
components with diverse bioactivities, including proteins, lipids
and nuclear acids (4,5). A variety of cells can release
exosomes, such as epithelial cells (6,7),
immune cells (8–10), reticulocytes (11), mast cells (12) and tumor cells (13,14).
In addition, exosomes have also been found in various bodily
fluids, including amniotic fluid (15), urine (16), malignant ascites (17), plasma (18), bile (19) and saliva (20). They have been proposed as a
potential source of diagnostic and prognostic biomarkers, and have
been proven to play important roles in intercellular communication
(21). Moreover, certain common
exosomal proteins, such as TSG101, Alix, CD9, CD81, CD63, GTPase
active proteins and cytoskeletal proteins, including actin, as well
as tubulin proteins have been found to be crucial for the
biogenesis, secretion and translocation of exosomes (22). Exosomes may also transport some
tumor-associated proteins, particularly those that originate from
cancer cells (23). Therefore,
exosomes in urine, saliva, plasma and other bodily fluids may be
utilized for the identification of diagnostic and prognostic
biomarkers in a non-invasive manner (24).
As regards ovarian cancer, exosomes have been
isolated from plasma/serum or ascites of patients with ovarian
cancer in some studies. In one study, the expression levels of 8
microRNAs (miRNAs or miRs; miR-21, miR-141, miR-200a, miR-200c,
miR-200b, miR-203, miR-205 and miR-214) were found to be elevated
in exosomes isolated from the serum of women with various stages of
ovarian cancer as compared to those from the serum of women with
benign diseases, suggesting that miRNAs of circulating tumor
exosomes can potentially be used as alternative diagnostic markers
(25). Peng et al proved
the origin of ascites-derived exosomes from patients with ovarian
cancer; two genes, namely FasL and TRAIL, were
identified in the exosome and were found to be responsible for the
apoptosis of different types of immune cells (26). Exosomes containing claudin 4 have
also been isolated from patients with EOC and found to be a
promising diagnostic marker as compared to CA125 (27). Therefore, the analysis of the
biological characteristics of ovarian cancer-derived exosomes may
be of great value to the diagnosis of ovarian cancer, monitoring
the therapeutic efficacy, and exploring their roles in tumor
progression and metastasis.
Regardless of the advances achieved in the
above-mentioned studies, the results, however, have only
highlighted the expression of limited individual molecules in
exosomes. The systemic proteomic analysis of ovarian cancer-derived
exosomes from patient plasma has seldom been reported, at least to
the best of our knowledge. Additionally, exosomes isolated from
clinical samples ought to possess greater significance and closer
association with the real-life situations of patients. Therefore,
in this study, we utilized exosomes from EOC patient plasma samples
and performed the first comprehensive proteomic analysis (to the
best of our knowledge) by liquid chromatography tandem mass
spectrometry (LC-MS/MS) with tandem mass tagging (TMT) to identify
potential biomarkers and investigate their clinical values. It
should be noted that, to the best of our knowledge, this is the
first research using the TMT technique to analyze the exosomal
proteins. TMT is a type of chemical label for the quantification
and identification of proteins, peptides, nucleic acids and other
biological macromolecules (28).
The chemical structures of TMTs are identical, but each contains
isotopes replaced at diverse regions. The biomolecule sequence
information is obtained during the fragmentation process and the
amount of the biomolecules is collected in the meantime, which
renders TMT an ideal tool for the high-throughput characterization
of the exosomal proteins (29).
Materials and methods
Subjects and materials
The study protocol was approved by the Ethics
Committee of Fudan University Shanghai Cancer Center (Shanghai,
China). All patients who provided signed informed consent forms in
this study were of Chinese origin and from the Department of
Gynecological Oncology at the Fudan University Shanghai Cancer
Center. For proteomics analysis, plasma samples from 3 patients
with ovarian cancer were obtained as the experimental group and 6
non-cancerous samples served as the controls between July, 2017 to
August, 2017. Another cohort of 80 female candidates (40 patients
and 40 healthy volunteers) was recruited for the verification of
specific biomarkers screened from the proteomics analysis from
June, 2016 to May, 2017. Detailed information of the 9 patients (3
with EOC and 6 with benign disease) for proteomics and 40 patients
with EOC is summarized in Tables I
and II, respectively. Blood was
collected into EDTA tubes and spun at 2,000 × g for 10 min at 4°C
to obtain plasma. The plasma was stored at -80°C for further
analysis. The exosome isolation kit (exoEasy kit) was purchased
from Qiagen GmbH (Hilden, Germany). TSG101 (sc-7964) and CD81
(sc-7637) antibodies were purchased from Santa Cruz Biotechnology
(Santa Cruz, TX, USA). Calnexin (cat. no. 2433) was purchased from
Cell Signaling (Cell Signaling Technology, Danvers, MA, USA).
Enzyme-linked immunosorbent assay (ELISA) kits for gelsolin (GSN;
EH0875), lipopolysaccharide binding protein (LBP; EH1560),
fibrinogen alpha chain (FGA; EH3065) and fibrinogen gamma chain
(FGG; EH0693) were obtained from Finetest (Fine Biotech, Wuhan,
China). Other materials, if not specifically stated, were all
purchased from Sigma-Aldrich (St. Louis, MO, USA).
 | Table IClinical characteristics of the
patients recruited for proteomics analysis. |
Table I
Clinical characteristics of the
patients recruited for proteomics analysis.
| No. | Age (years) | FIGO stage | Grading | Lymph node
metastasis | Histological
subtype | CA125 | Post-operative
residual tumor | Recurrence at time
of analysis | Survival status at
time of analysis |
|---|
| EOC1 | 46 | IIIC | G3 | NA | High-grade serous
carcinoma | 533 | None | NA | NA |
| EOC2 | 67 | III | G3 | N1 | High-grade serous
carcinoma | 7378 | <0.5 cm | Yes | Yes |
| EOC3 | 52 | IV | G2 | N1 | Moderately
differentiated squamous cell carcinoma | 420.4 | NA | Yes | Yes |
| B1 | 44 | | G1 | N1 | Cervical myoma | 14.79 | None | Yes | Yes |
| B2 | 47 | | G1 | N0 | Cervical myoma | 11.79 | None | No | Yes |
| B3 | 45 | | G1 | N0 | Hysteromyoma | 13.48 | None | No | Yes |
| B4 | 54 | | G1 | N0 | Hysteromyoma | 14.8 | None | No | Yes |
| B5 | 44 | | G1 | N0 | Uterine
leiomyoma | 12.24 | None | No | Yes |
| B6 | 53 | | G1 | N0 | Uterine
leiomyoma | 11.42 | None | No | Yes |
| Table IIClinical characteristics of patients
with epithelial ovarian cancer recruited for ELISA. |
Table II
Clinical characteristics of patients
with epithelial ovarian cancer recruited for ELISA.
| No. | Age (years) | FIGO stage | Grading | Histological
subtype | CA125 | Post-operative
residual tumor | Recurrent disease
at time of analysis | Survival status at
time of analysis |
|---|
| EOC1 | 51 | IIIB | G3 | HGSC | 1366 | NA | No | Yes |
| EOC2 | 58 | IIIC | G3 | HGSC | 734.7 | <0.5 cm | No | Yes |
| EOC3 | 40 | IIIC | G3 | HGSC | 89.92 | None | No | Yes |
| EOC4 | 80 | IIIC | G3 | LGSC | 600 | >2 cm | Yes | NA |
| EOC5 | 56 | IIIC | G3 | HGSC | 1201 | NA | No | Yes |
| EOC6 | 56 | IIIB | G3 | HGSC | 1327 | None | No | Yes |
| EOC7 | 63 | IIIC | G3 | HGSC | 962.4 | NA | No | Yes |
| EOC8 | 72 | IIIC | G3 | HGSC | 71.29 | <0.5 cm | No | Yes |
| EOC9 | 49 | IIIB | G3 | HGSC | 381.5 | None | No | Yes |
| EOC10 | 39 | IIIC | G3 | HGSC | 861 | NA | No | Yes |
| EOC11 | 40 | IIIC | G3 | HGSC | >5000 | None | No | Yes |
| EOC12 | 69 | IV | G3 | HGSC | >1000 | <0.2 cm | Yes | Yes |
| EOC13 | 61 | IIIC | N.A. | N.A. | 453.2 | NA | No | Yes |
| EOC14 | 63 | IIIC | G3 | HGSC | 2253 | <1 cm | No | Yes |
| EOC15 | 61 | IV | G3 | Adenocarcinoma | 154 | NA | No | Yes |
| EOC16 | 53 | IV | G3 | HGSC | 893.9 | <1 cm | No | Yes |
| EOC17 | 61 | IIIC | G3 | HGSC | 256.5 | <0.5 cm | NA | NA |
| EOC18 | 52 | IV | G3 | HGSC | N.A. | NA | Yes | Yes |
| EOC19 | 60 | IIIC | G3 | HGSC | 513.7 | NA | No | Yes |
| EOC20 | 57 | III | G3 | HGSC | 1137 | <0.2 cm | No | Yes |
| EOC21 | 47 | IIIC | G3 | HGSC | 465.7 | None | No | Yes |
| EOC22 | 64 | IVB | G3 | HGSC | >5,000 | NA | No | Yes |
| EOC23 | 50 | IVB | G3 | HGSC | 2276 | None | No | Yes |
| EOC24 | 66 | IV | G3 | HGSC | 553.9 | None | No | Yes |
| EOC25 | 58 | IIIC | G3 | HGSC | >5,000 | NA | No | Yes |
| EOC26 | 75 | IV | G3 | HGSC | 1660 | NA | NA | NA |
| EOC27 | 64 | IIIC | G3 | HGSC | 511.1 | None | NA | NA |
| EOC28 | 56 | IIIA | G3 | HGSC | 8.28 | None | No | Yes |
| EOC29 | 54 | IIIC | G3 | HGSC | 317.7 | None | No | Yes |
| EOC30 | 51 | IIIC | G3 | HGSC | 474.7 | <2 cm | NA | Yes |
| EOC31 | 68 | IIIA | G3 | HGSC | 2661 | None | Yes | Yes |
| EOC32 | 67 | III | G3 | HGSC | 295.1 | None | NA | Yes |
| EOC33 | 45 | IIIB | G3 | HGSC | 1487 | <0.3 cm | NA | NA |
| EOC34 | 47 | IIIC | G3 | HGSC | 921.2 | NA | NA | NA |
| EOC35 | 71 | IIIC | G3 | HGSC | 157.4 | 0.1–0.2 cm | No | Yes |
| EOC36 | 56 | IIIC | G3 | HGSC | 425.2 | <1 cm | NA | Yes |
| EOC37 | 48 | IIIC | G3 | HGSC | 276 | NA | No | Yes |
| EOC38 | 50 | IIIC | G3 | HGSC | 1556 | None | No | Yes |
| EOC39 | 50 | IIIC | G3 | HGSC | 1423 | NA | No | Yes |
| EOC40 | 44 | IIIC | G3 | HGSC | 17.98 | None | No | Yes |
Exosome isolation and purification
Exosomes were isolated from plasma using the exoEasy
Maxi kit as described in the manufacturer's manual. Briefly, 1
volume of buffer XBP was added into 1 volume of plasma. The mix was
topped up onto the exoEasy spin column and centrifuged at 500 × g
for 1 min at 4°C. The flow-through was discarded and 10 ml buffer
XWP was added and centrifuged at 5,000 × g for 5 min at 4°C to
remove residual buffer from the column. The spin column was then
transferred to a fresh collection tube and 1 ml Buffer XE was added
to the membrane and incubated for 1 min. The elute was collected by
centrifugation at 500 × g for 5 min at 4°C, which contained the
purified exosome fraction. After measuring the total protein
concentration of the purified exosomes using BCA assays (Thermo
Fisher Scientific, Waltham, MA, USA), the exosome preparations were
stored at −80°C and resuspended in PBS for transmission electron
microscopy (TEM), western blot analysis (WB), nanoparticle tracking
analysis (NTA) and dynamic light scattering (DLS) analysis to
verify the nature of the isolated particles.
NTA and DLS analysis
NTA was performed to measure the size and the
concentration of the isolated exosomes using NanoSight NS300
(Malvern, Westborough, MA, USA) according to the operating
instructions without any changes. In addition, DLS was conducted
using Zetasizer Nano (Malvern) referring to the operating
instructions without further modifications.
TEM
The isolated exosomes were processed at room
temperature for TEM. The samples were diluted 1:5 in 1X PBS prior
to fixation with 2.0 % glutaraldehyde (G5882, Sigma-Aldrich).
Following fixation, a 75-mesh grid (G2075C; Agar Scientific, Essex,
UK) was laid on a drop of sample for 10 min; the grid was then
rinsed 10 times with MiliQ H2O (1 min per rinse).
Subsequently, the grid was firstly laid on a drop of uranyl acetate
(pH 7.0, 2624; SPI-CHEM, West Chester, PA, USA) for 10 min. After
rinsing with Milli-Q H2O and methylcellulose uranyl (pH
4.0), the grid was incubated at room temperature for 10 min on a
drop of methylcellulose uranyl (pH 4.0, M-6385, Sigma-Aldrich). The
exosome samples were eventually analyzed with a FEI Tecnai™ T12
electron microscope (Thermo Fisher Scientific).
Western blot analysis
Exosomes were prepared as described above. RIPA
buffer (#9806, Cell Signaling Technology) was used to extract the
proteins from the exosomes. In brief, 500 µl of RIPA buffer
were added to the exosome samples and this was maintained for 1 h
at room temperature. The mixtures were then centrifuged at 12,000 ×
g for 5 min at 4°C. The supernatants were collected as the exosomal
protein extracts. Subsequently, 5X SDS-loading buffer was added to
dissolve the proteins from exosomes, diluted to 1X SDS-loading
buffer, and then heated at 95°C for 5 min. The samples were
centrifuged at 12,000 × g for 5 min at 4°C to remove insoluble
precipitates. Supernatants were subsequently loaded onto SDS-PAGE
(3% stacking gel, 12% running gel; Bio-Rad, Munich, Germany),
running in a Mini Protean 2 electrophoresis system (Bio-Rad). The
protein was transferred onto a polyvinylidene fluoride membrane
(Bio-Rad) in transfer buffer. After being blocked with 5% non-fat
milk in TBST for 1 h at room temperature, the membrane was
incubated with the primary antibody, including CD81 (1:1,000),
TSG101 (1:1,000), and calnexin (1:1,000) overnight at 4°C. The
membrane was rinsed with 1X TBST and this was repeated 3 times (5
min per rinse). The membrane was then labeled with HRP-conjugated
secondary antibody (1:2,000; cat. no. 7074; Cell Signaling
Technology) for 1 h at room temperature. Again, the membrane was
rinsed with 1X TBST and this was repeated 3 times (5 min per
rinse). The final products were detected using an enhanced and
freshly prepared chemiluminescence (ECL) system (Thermo Fisher
Scientific). Cell lysates were also characterized in order to
compare with exosome samples.
Proteomics of exosomes
LC-MS/MS was performed for proteomics analysis. In
brief, LC-MS/MS was used to analyze the purified proteins from the
exosome fractions. After extracting the protein from the exosome
samples as mentioned above, the fractions were resolved in two
lanes of a 10% SDS-PAGE gel respectively and subsequently stained
with Coomassie blue staining solution (B6529, Sigma-Aldrich) at
room temperature for 1 h. Each lane was excised and divided into 6
sections equally for in-gel trypsin digestion; following reduction
and alkylation, each section was digested with trypsin overnight.
Following digestion, the solution was centrifuged at 12,000 × g at
room temperature for 20 min to collect the dissolved peptides. TEAB
buffer was added and the solution was centrifuged at 12,000 × g at
room temperature for 20 min. The filtrates were collected and
lyophilized to obtain the dry powder. The peptide samples were
dissolved in TEAB buffer and mixed with anhydrous acetonitrile and
vortexed for 1 min. TMT reagents (Thermo Fisher Scientific) were
added to the resulting solution and maintained for 1 h at room
temperature. Subsequently, 5% hydroxylamine was added to terminate
the reaction and the samples were ready for MS. The peptides were
analyzed with a Q-Exactive LC-MS/MS (Thermo Fisher Scientific). The
raw data were converted to mascot generic (mgf) files using
Proteome Discoverer version 2.2 (Thermo Fisher Scientific) and the
mgf files were then searched against the Uniprot human proteome
database using an in-house Mascot Server version 2.4.1 (Matrix
Science, London, UK).
Bioinformatics analysis
An online website, Database for Annotation,
Visualization and Integrated Discovery (DAVID, http://david.abcc.ncifcrf.gov/) is an open
database which provides thorough functional annotation tools for
researchers to understand versatile biological and functional
meanings behind numerous genes (30). Gene Ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway enrichment
analysis were performed for the identified differentially expressed
genes (DEGs) using the DAVID database (30). A P-value <0.05 was set to
distinguish statistically significant enrichment results. The
functional interactions between proteins can be plotted to
illustrate the molecular mechanisms and signaling pathways of
cellular processing. A protein-protein interaction (PPI) network of
DEGs was constructed using an open-access bioinformatics tool, the
Search Tool for the Retrieval of Interacting Genes (STRING,
http://string.embl.de/) database (31) and subsequently was visualized using
Cytoscape (32). The confidence
score ≥0.7 was set as the cut-off criterion and the Molecular
Complex Detection (MCODE) was performed to screen modules of PPI
network with a degree cut-off of 2, a node score cut-off of 0.2, a
k-core of 2, and a max. depth of 100 (33). In order to supplement the evidence
of the identified genes as potential prognostic markers,
progression-free survival (PFS) and overall survival (OS) were
simulated on a Kaplan Meier-plotter (KM plotter, http://kmplot.com/analysis/), which is capable of
assessing the effects of >50,000 genes on survival using
>10,000 cancer samples, including breast, ovarian, lung and
gastric cancer (GC) samples (34).
The patients with ovarian cancer were split into 2 groups according
to the expression of a particular gene (high vs. low). The hazard
ratio (HR) with 95% confidence intervals and log-rank P-value were
calculated and displayed on the webpage.
Procoagulation activity mediated by
plasma exosomes in patients with EOC
Exosomes associated procoagulant activity was
measured follows: Briefly, 20 µl of each sub-fraction-PBS
solution were incubated for 15 min in 100 µl incubation
buffer containing 10 mM HEPES, 25 µM negatively charged
phospholipids which was composed of dioleoylphosphatidylserine and
dioleoylphosphatidylcholine at a 1:9 ratio (840035P and 850375P;
Avanti Polar Lipids, Alabaster, AL, USA), 137 mM NaCl, 4 mM KCl, 6
mM CaCl2, 5 mg/ml bovine serum albumin and 5 U/ml
hirudin (pH 7.45, H0393; Sigma-Aldrich). Subsequently, 40 µl
of 281 nM Factor X (HFX1010; Enzyme Research Laboratories, Ltd.,
Swansea, UK) was added following 40 µl 5.63 nM Factor VII
(HFVII1007; Enzyme Research Laboratories, Ltd.) or blank buffer and
25 µl Factor Xa chromogenic substrate S2765 (82141339;
Chromogenix, Milano, Italy) to commence the reaction. The rate of
the chromophoric group p-nitroaniline formation was recorded for 90
min at 405 nm and the rate of Factor Xa generation was calculated.
The procoagulant activity was expressed as the rate of Factor Xa
generation. Recombinant human tissue factor (B4212,
Dade® Innovin; Siemens Healthcare Diagnostics, Marburg,
Germany) was used as the positive control.
ELISA for GSN, LBP, FGA, and FGG for
prognostic verification
The levels of all 4 markers, namely GSN, LBP, FGA
and FGG were measured according to the manufacturer's instructions
(Fine Biotech). Briefly, 200 µl of Assay Diluent were added
to each well. Subsequently, 50 µl of Standard, control, or
sample was also added to each well. The plate was incubated at room
temperature for 2 h on a horizontal orbital microplate shaker. Each
well was aspirated and washed, repeating the process 2 times for a
total of 3 washes. Subsequently, 200 µl of Conjugate were
added to each well and the plate was incubated at room temperature
for 2 h on the shaker again. Each well was aspirated and washed 3
times. A total of 200 µl Substrate Solution were then added
to each well and incubated at room temperature for 30 min on the
benchtop free from light. Eventually, 50 µl of Stop Solution
was added to each well. UV absorption was measured at 450 nm within
30 min using a plate reader (Thermo Fisher Scientific). Wavelength
correction was set to 540 or 570 nm.
Statistical analysis
Receiver Operating Characteristic curve (ROC)
analysis was performed to evaluate the efficiency of GSN, LBP, FGA
and FGG as diagnostic markers. The result of procoagulation assay
was analyzed using a Student's t-test as each case was independent
from each other. All statistical analyses were performed using
GraphPad Prism® software (GraphPad Prism®
Version 6).
Results
Isolation and characterization of plasma
exosomes in ovarian cancer
The exosomes were isolated from the patient plasma
samples with a commercially available exosome isolation kit. The
isolation procedure was referred to the manual without further
modifications. Western blot analysis was conducted to detect the
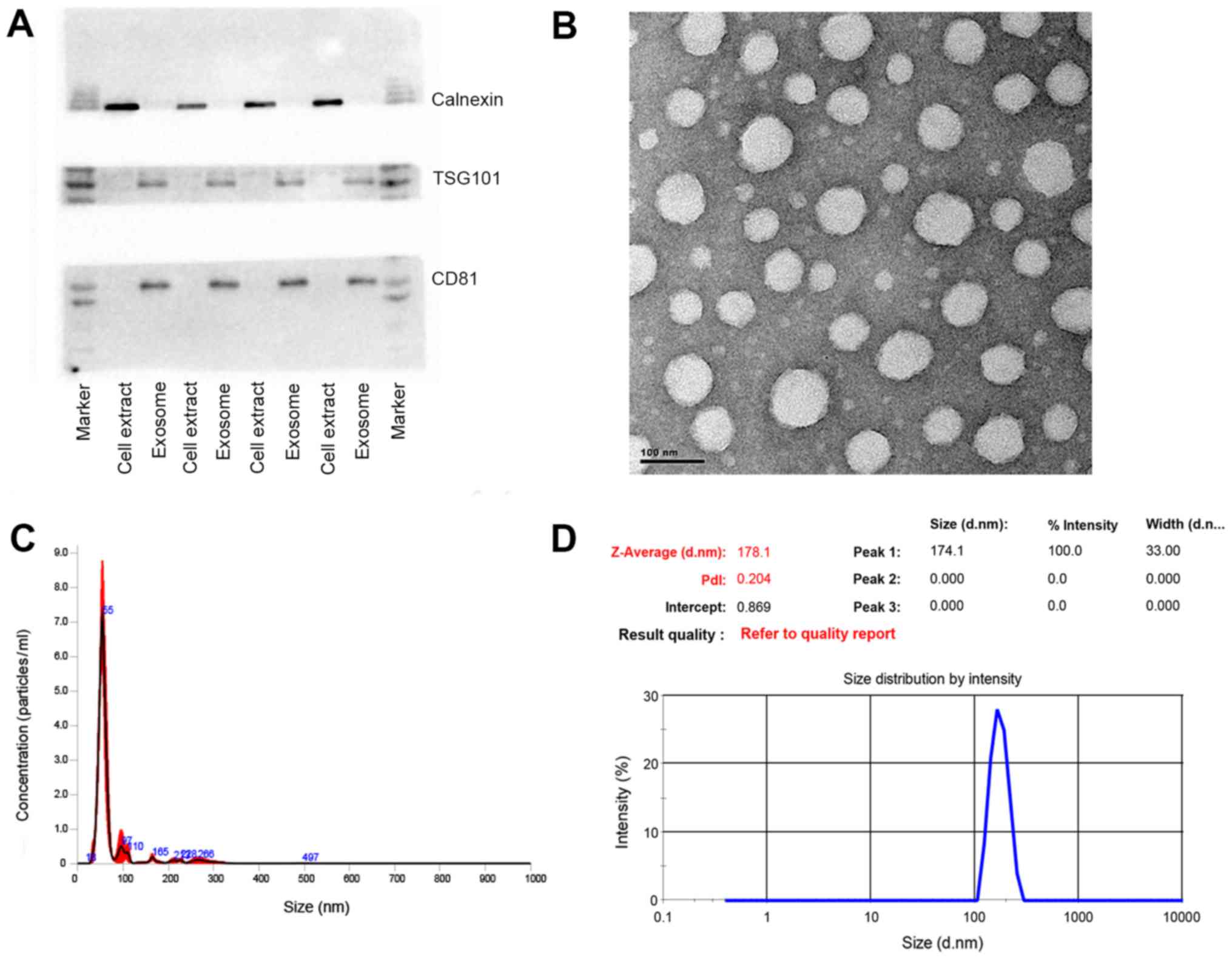
exosomal protein markers. As shown in Fig. 1A, TSG101 and CD81, as exosomal
protein markers that are involved in exosome biogenesis, maturation
and secretion, were detected in all the plasma exosome samples. On
the contrary, calnexin, an endoplasmic reticular protein, was only
found to be present in the cell extracts, proving that the isolated
exosomes were highly purified and not contaminated by redundant
intracellular components. TEM was utilized to further characterize
the size and morphology of the exosomes. A number of spherical
particles were detected on the copper meshes. The size remained
within the range of 30–100 nm and approximately 70 nm in diameter
(Fig. 1B). Other methodologies
were applied to confirm the size as well. For instance, NTA, as a
conventional method of characterizing exosomes, was exploited to
measure the size and concentration based on the tracking of
Brownian movement (35). The mean
size was approximately 70 nm and the concentration was
approximately 1.53×108 following a 1,000-fold dilution
(Fig. 1C). Another technique, DLS,
was also applied to measure the size that was greatly larger due to
the aquated membrane covering exosomes (Fig. 1D). Another crucial parameter, the
polydispersity index (PDI) was also evaluated by this technique to
characterize the size distribution of the exosomes. The PDI was
approximately 0.22, showing relatively even size distribution of
exosomes, which could also be confirmed by the sharp single peak in
NTA analysis. Taken together, these results demonstrated that the
exosomes were successfully isolated from the plasma with high
purity and well characterized by various methods.
Proteomics analysis of exosomes derived
from patient plasma samples
To determine the protein profile of the exosomes,
total exosomal proteins were separated by SDS-PAGE on a 12% gel.
After every band was cut into sections and subjected to trypsin
digestion in 1.5 ml Eppendorf tubes respectively overnight, the
extracted peptides were analyzed by LC-MS/MS. A total of 262
proteins in the cancerous plasma exosomes and 257 proteins in the
non-cancerous plasma exosomes were identified from the protein
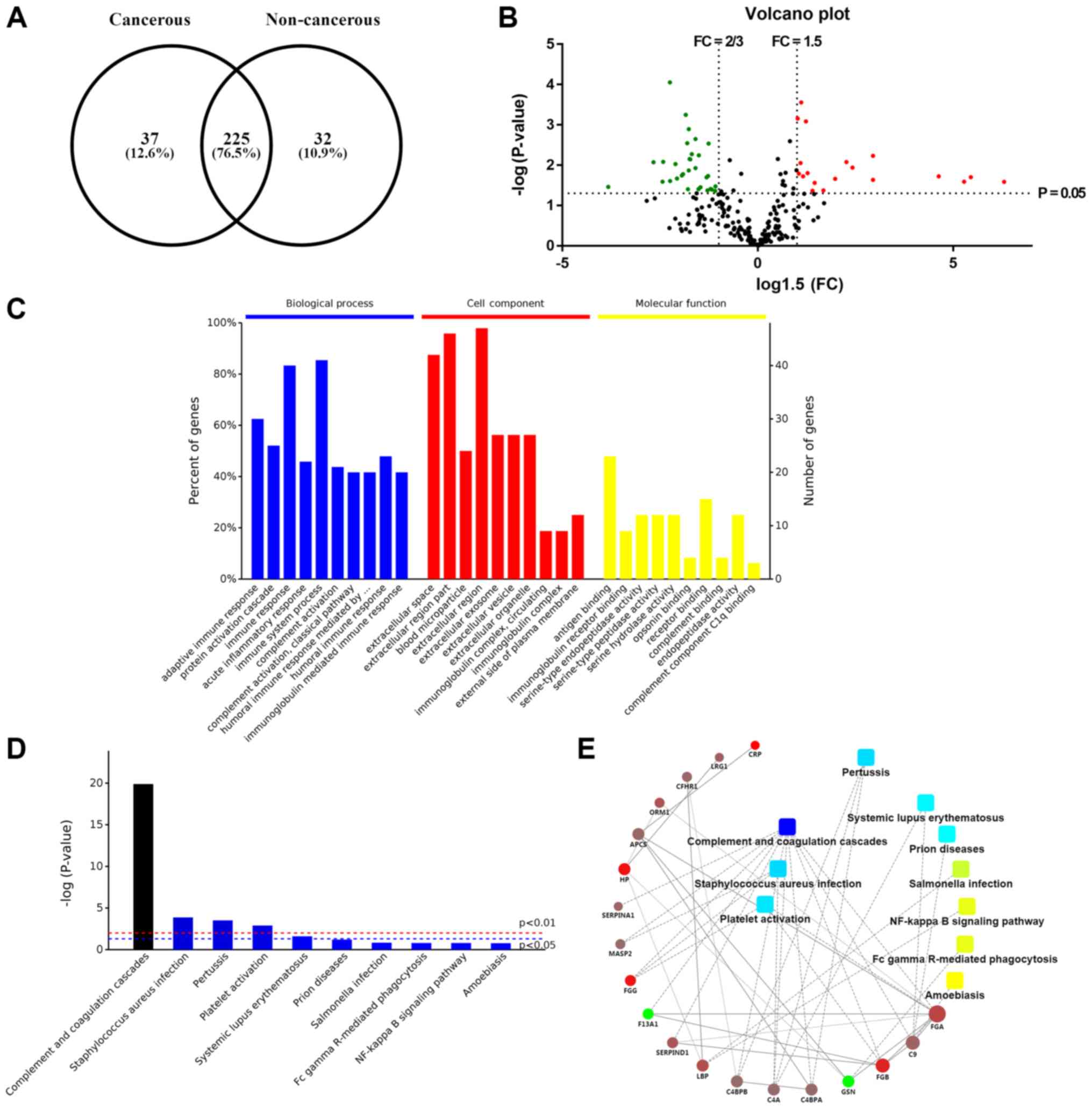
database. In total, 225 proteins were present in both samples
(Fig. 2A). Based on the protein
information following crude screening, the fold change (FC) value
and P-value were calculated from the t-test. An FC >1.5 or FC
<2/3 and P-value <0.05 were set as the criteria for the
selection of upregulated or downregulated genes. Eventually, 50
genes were screened as DEGs, which were depicted in a volcano plot
(Fig. 2B). The red dots represent
overexpressed genes, which contained 19 genes; while the green dots
represent downregulated genes, 31 in sum.
Annotation of identified genes
GO analysis was performed for the classification of
various molecular functions. The identified genes were categorized
into several groups, including biological process, cellular
components and molecular functions. In Fig. 2C, the 10 most statistically
significant items in biological process, cellular components and
molecular functions are listed in sequence. For biological process,
the exosomal genes were mainly involved in immune response, protein
activation cascade, complement activation, and so on, which may
indicate that plasma exosomes contribute to the proper functioning
of immune system. The aberrant regulation of body immunity could
partially involve the tumorigenesis and metastasis of EOC. For
instance, in one study, exosomes were revealed to be able to
facilitate the intercellular communication in tumor
microenvironment, thereby remodeling normal macrophages to a
tumor-activated phenotype with the assistance of hypoxia-inducible
factors (36). As for the GO
analysis of cellular components, these DEGs were mainly associated
with extracellular space, extracellular region part, blood
microparticle, extracellular region, extracellular exosome,
extracellular vesicle, etc., confirming that these proteins were
originated from plasma exosomes. GO enrichment analysis in
molecular functions was also conducted for these 50 genes. Some of
the genes involved in the following molecular functions: antigen
binding, immunoglobulin receptor binding, serine-type endopeptidase
activity, and so on. These enriched molecular functions were
closely relevant to corresponding biological process the other way
around, such as immune response, indicating that the molecular
function of each gene mutually interacted and participated in these
meaningful biological processes. Additionally, other processes may
also be related to particular functions of the exosomes, such as
complement binding and receptor binding.
KEGG pathway enrichment and PPI
network
As can be seen from Fig. 2D, the top 10 pathways that contain
these exosomal proteins, among which complement and coagulation
cascades and platelet activation were closely related to abnormal
coagulation function of patients with EOC, which could be an
explanation for the hypercoagulability in a number of patients with
EOC. Pathways of Staphylococcus aureus infection, pertussis
and systemic lupus erythematosus may result from immune
dysfunction, which was in accordance with the results of GO
enrichment analysis. Most other pathways ought to be involved in
tumor growth, progression and apoptosis, including Fc γ
Receptor-mediated phagocytosis and the NF-κB signaling pathway.
Either direct or indirect interactions between DEGs were plotted in
the PPI network (Fig. 2E). It is
worth noting that FGG and FGA were relevant to both complement and
coagulation cascades, and platelet activation, implying that
exosomal FGG and FGA may be key genes regulating the coagulation
cascade in patients with EOC. Moreover, GSN and LBP were linked to
Fc γ Receptor-mediated phagocytosis and the NF-κB signaling
pathway, respectively, which made it more explicit that GSN and LBP
originated from exosomes played a role in tumor apoptosis.
Exosome-associated hypercoagulability in
patients with EOC
In clinical practice, patients with EOC often suffer
from venous thromboembolism (VTE) due to the aberrant activation of
platelet and coagulation dysfunction (37). Most of them remain in a
hypercoagulable state and the status of coagulation has emerged as
an indicator for EOC. As shown in Table III, among the 50 DEGs, 10 genes
participated in the complement and coagulation cascade, namely
coagulation factor XIII A chain (F13A1), coagulation factor IX
(F9), serpin family A member 1 (SERPINA1), FGA, FGB, FGG,
complement C9 (C9), complement component 4 binding protein alpha
(C4BPA), complement C8 alpha chain (C8A) and complement
component 4 binding protein beta (C4BPB), the majority of which
were upregulated in patients with EOC, suggesting that the
overexpression of these exosomal genes mediates the coagulation
cascade, as well as platelet activation and induces blood clotting.
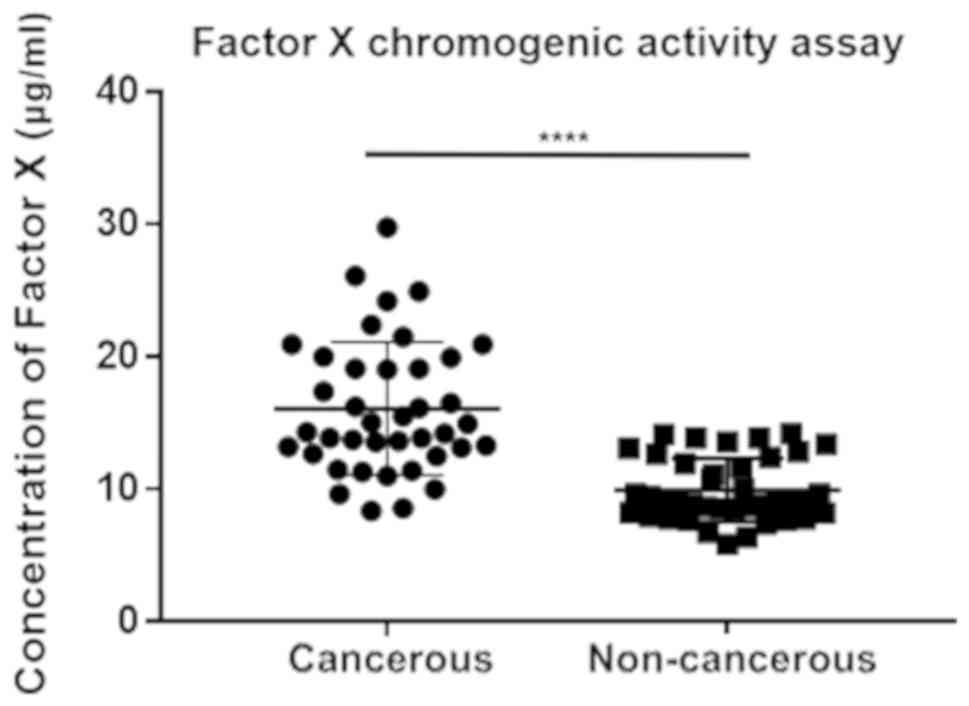
Therefore, the procoagulation assay was conducted to verify the
discrepant influences of cancerous and non-cancerous plasma
exosomes on the rate of clotting (Fig.
3). The EOC group exhibited a much higher degree of coagulation
(P<0.0001), which proved the promotion and acceleration effects
on coagulation by EOC exosomes and confirmed the estimated pathways
and the subsequent regulations.
| Table IIIInformation of identified
differentially expressed genes (DEGs). |
Table III
Information of identified
differentially expressed genes (DEGs).
| Accession no. | Gene ID | Fold change | P-value | Expression |
|---|
| P01706 | IGLV2-11 | 0.218133124 | 0.046131844 | Downregulated |
| P01705 | IGLV2-23 | 0.301249245 | 0.032502181 | Downregulated |
| P04211 | IGLV7-43 | 0.337616894 | 0.003289896 | Downregulated |
| A0A075B6K0 | IGLV3-16 | 0.342103003 | 0.027922644 | Downregulated |
| A0A075B6S2 | IGKV2D-29 | 0.361179729 | 0.018761308 | Downregulated |
| P01780 | IGHV3-7 | 0.413239637 | 0.000846937 | Downregulated |
| A0A0B4J1V0 | IGHV3-15 | 0.413556853 | 0.003187854 | Downregulated |
| P01701 | IGLV1-51 | 0.466774595 | 0.017830268 | Downregulated |
| P01859 | IGHG2 | 0.47139263 | 0.017141917 | Downregulated |
| P06396 | GSN | 0.47577625 | 0.000550377 | Downregulated |
| P06312 | IGKV4-1 | 0.480672648 | 0.002873312 | Downregulated |
| P00488 | F13A1 | 0.481024159 | 0.005927534 | Downregulated |
| P01860 | IGHG3 | 0.481916606 | 0.027922715 | Downregulated |
| A0A0B4J1V2 | IGHV2-26 | 0.48266588 | 0.014221922 | Downregulated |
| A0A075B7D0 | IGHV1OR15-1 | 0.486125479 | 0.032529504 | Downregulated |
| A0A0B4J1X8 | IGHV3-43 | 0.488511091 | 0.006554509 | Downregulated |
| A0A0B4J1Y9 | IGHV3-72 | 0.493082012 | 0.001689489 | Downregulated |
| A0A0C4DH35 | IGHV3-35 | 0.494979343 | 0.006757808 | Downregulated |
| P05452 | CLEC3B | 0.507434994 | 0.004828297 | Downregulated |
| A0A075B6I9 | IGLV7-46 | 0.511105442 | 0.007728489 | Downregulated |
| S4R460 | IGHV3OR16-9 | 0.520150846 | 0.010756891 | Downregulated |
| A0A075B6R9 | IGKV2D-24 | 0.52141676 | 0.035994038 | Downregulated |
| Q9NQ79 | CRTAC1 | 0.533552214 | 0.009071347 | Downregulated |
| P0DOY2 | IGLC2 | 0.540646515 | 0.025332541 | Downregulated |
| A0A0A0MS15 | IGHV3-49 | 0.567732527 | 0.014566172 | Downregulated |
| A0A0B4J1U7 | IGHV6-1 | 0.593352797 | 0.033839926 | Downregulated |
| A0A075B7B8 | IGHV3OR16-12 | 0.604561348 | 0.003882883 | Downregulated |
| A0A0B4J231 | IGLL5 | 0.608827162 | 0.049914959 | Downregulated |
| P01834 | IGKC | 0.641872888 | 0.032179959 | Downregulated |
| P01742 | IGHV1-69 | 0.646993729 | 0.042670996 | Downregulated |
| F8W1S1 | KRT74 | 0.652950133 | 0.045547508 | Downregulated |
| Q99784 | OLFM1 | 1.520266138 | 0.016352632 | Upregulated |
| P0C0L4 | C4A | 1.523579037 | 0.011662874 | Upregulated |
| Q03591 | CFHR1 | 1.528039639 | 0.000491116 | Upregulated |
| P36980 | CFHR2 | 1.539803521 | 0.02091088 | Upregulated |
| P02743 | APCS | 1.57664451 | 0.01704065 | Upregulated |
| P06702 | S100A9 | 1.644481596 | 0.01755607 | Upregulated |
| E7ETH0 | CFI | 1.732825958 | 0.031482969 | Upregulated |
| P02748 | C9 | 1.867280995 | 0.024739231 | Upregulated |
| P01009 | SERPINA1 | 1.885728943 | 0.033000428 | Upregulated |
| P02750 | LRG1 | 1.939799893 | 0.045335361 | Upregulated |
| P05546 | SERPIND1 | 2.289112647 | 0.020819823 | Upregulated |
| P02763 | ORM1 | 2.538489431 | 0.007998879 | Upregulated |
| P18428 | LBP | 2.616390498 | 0.012556488 | Upregulated |
| A0A096LPE2 | SAA2-SAA4 | 3.287876685 | 0.005920009 | Upregulated |
| P02671 | FGA | 3.31200379 | 0.02307751 | Upregulated |
| P02675 | FGB | 6.618243962 | 0.018788218 | Upregulated |
| P02679 | FGG | 8.57650775 | 0.025580522 | Upregulated |
| P00738 | HP | 10.72525182 | 0.018597109 | Upregulated |
| P02741 | CRP | 11.99122018 | 0.026727478 | Upregulated |
Identification of diagnostic and
prognostic biomarkers based on the results of proteomics
analysis
As mentioned above, 50 DEGs were identified and
sorted according to FC value with statistical significance, among
which 19 genes were all enriched in GO analysis, KEGG pathway
analysis, as well as the PPI network. In total, 4 genes (LBP,
FGG, FGA and GSN) were eventually selected as the
potential diagnostic and prognostic biomarkers for the reason that
they were all involved in two categories of function enrichment,
namely coagulation and apoptosis related pathways. The proteomics
analysis revealed that the protein levels of FGA and GSN were
upregulated. FGG and LBP, on the contrary, were downregulated.
Another cohort of patients (40 with EOC and 40 non-cancerous
subjects) was enrolled in the validation assay of these 4 genes.
ELISA was conducted and the sensitivity and specificity were
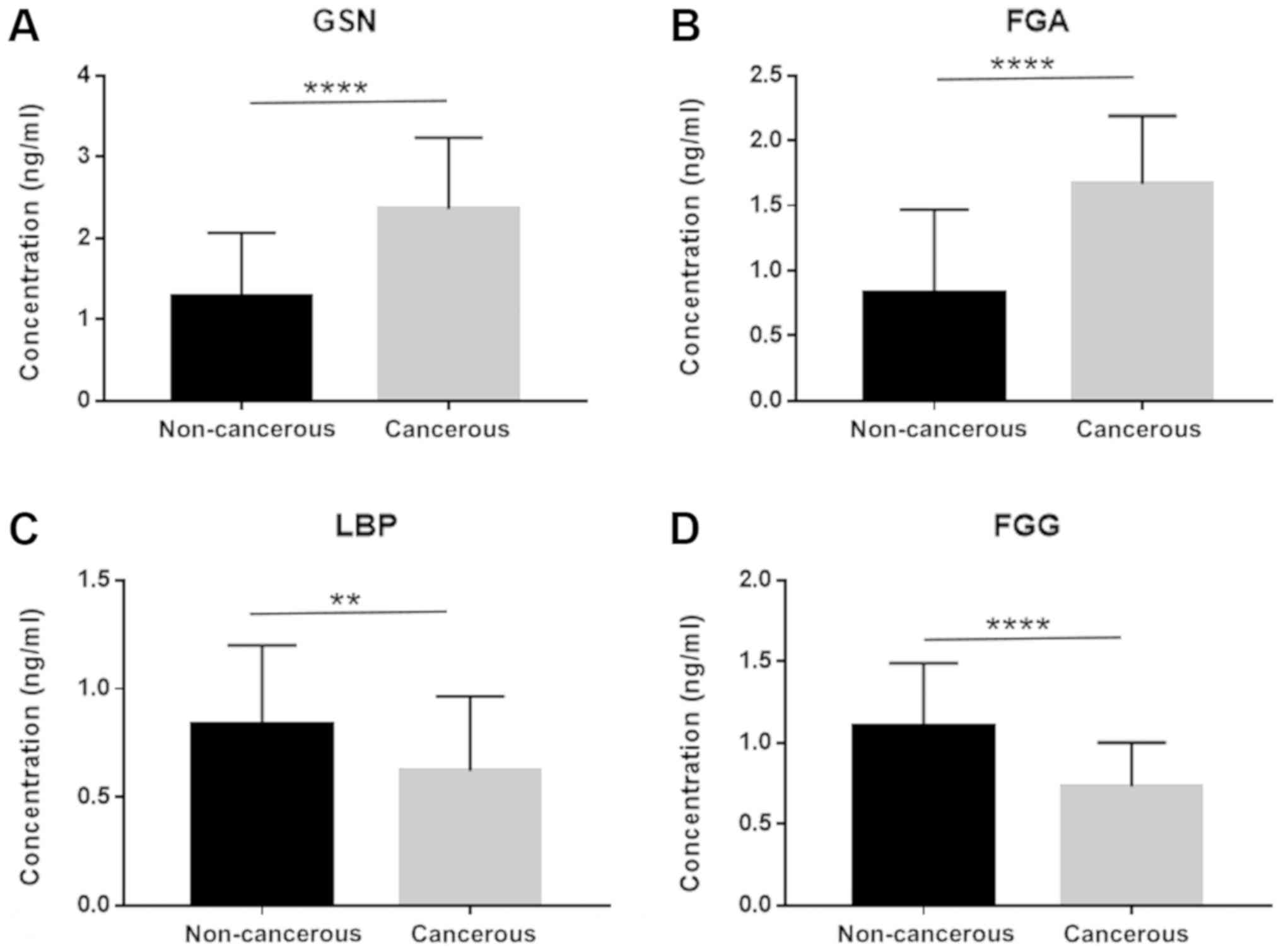
accessed by ROC analysis. The expression of these 4 genes in
exosomes derived from the 80 patient plasma samples is shown in
Fig. 4. The FGA and GSN levels
were significantly elevated in the cancer group, which was
consistent with what was observed in the proteomics analysis.
Similarly, the FGG and LBP levels were downregulated in the cancer
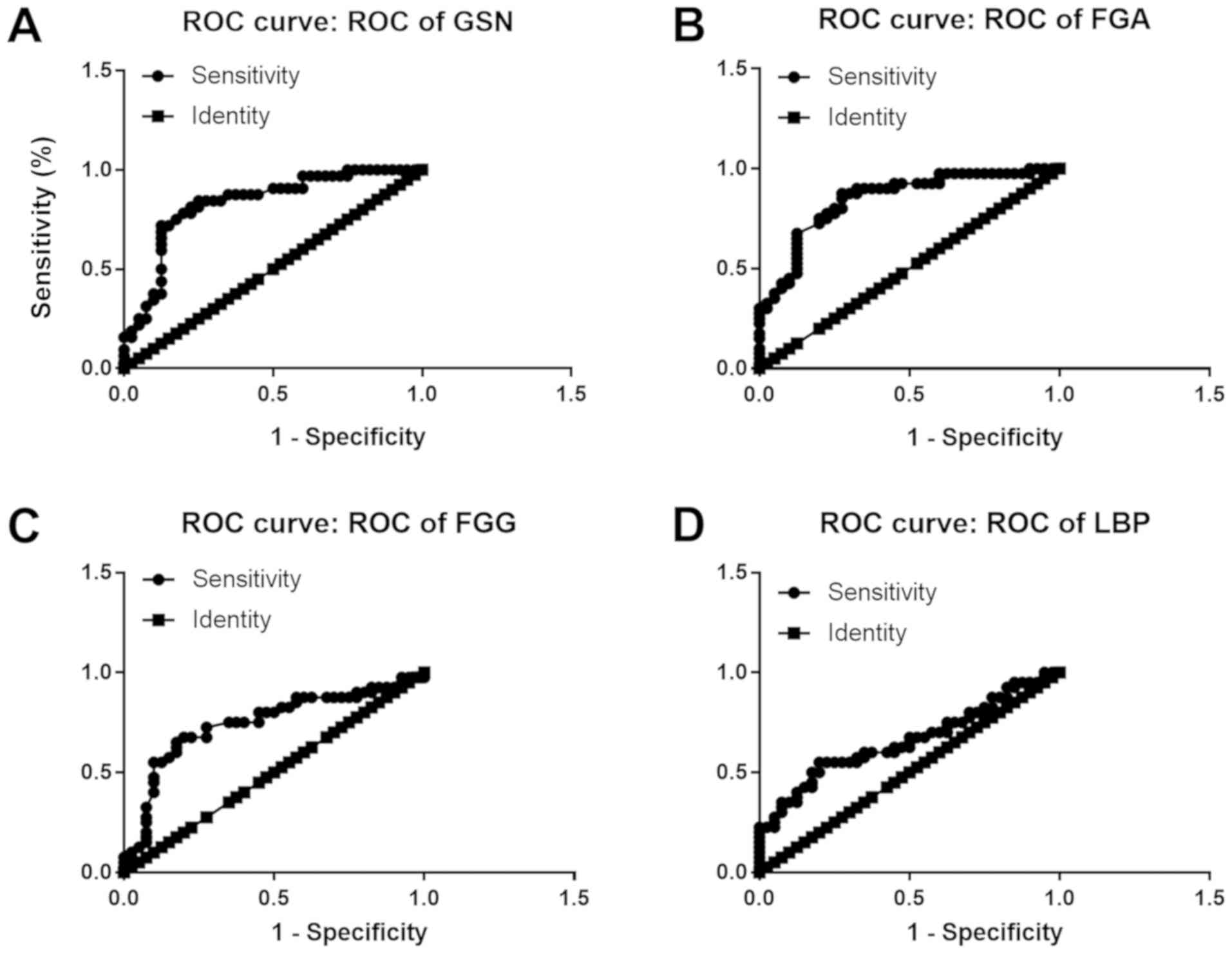
patient cohort. FGA conferred the highest area under the curve
(AUC) among all 4 candidates, which was approximately 0.8459
(Fig. 5B). The lowest AUC belonged
to LBP, approximately 0.6588 (Fig.
5D). Thus, FGA emerged as a promising biomarker for the
diagnosis of EOC. For prognostic biomarker screening, the OS and
PFS of patients as regards FGG and LBP expression were estimated on
the Kaplan Meier-plotter, which is an open-access online database
for estimating the progression-free survival (PFS) and overall
survival (OS) of over 50,000 genes on survival using >10,000
cancer samples, including breast, ovarian, lung and gastric cancer
(34). The patients with ovarian
cancer were divided into 2 groups according to the expression of a
particular gene (high vs. low). The hazard ratio (HR) with 95%
confidence intervals and log-rank P-value were calculated. As can
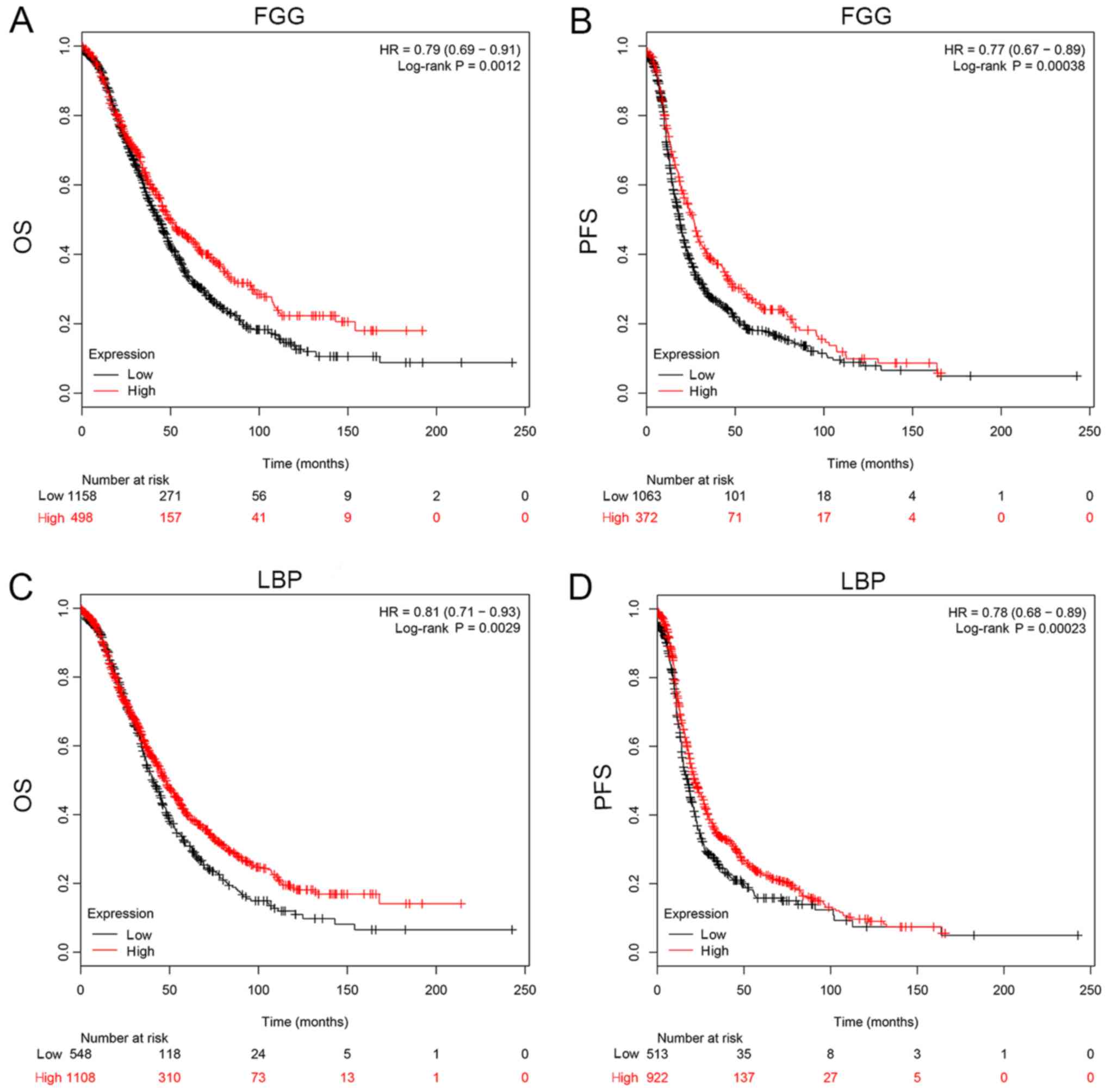
be seen from Fig. 6, FGG (HR, 0.79
for OS and 0.77 for PFS) and LBP (HR, 0.81 for OS and 0.78 for PFS)
could both be applied to predict the prognosis of patients with
EOC. It was found that a high mRNA expression of FGG or LBP was
associated with a shorter PFS and OS for patients with EOC,
reflecting a worse prognosis. However, it should be noted that the
potential for these genes as prognostic biomarkers was only
investigated on a Kaplan Meier-plotter that contains thousands of
cases. In fact, the survival curve was derived from the results of
gene expression, but not from exosomal gene expression. Further
studies are required for the validation and verification of these
genes in exosomes as prognostic indicators.
Discussion
Exosomes belong to extracellular vesicles with
30–100 nm in diameter and are secreted by various types of cells.
Researchers nowadays pay more attention to the studies of exosomes
to investigate the important roles in intercellular communications.
Despite tremendous research being made on exosomes in a variety of
types of cancer, these studies have shed light into the pivotal
roles of several exosomal molecules in tumorigenesis, evasion,
metastasis and recurrence (5–12).
However, there were few reports on the systemic proteomic analysis
of exosomes, particularly for plasma exosomes in ovarian cancer, as
compared to other types of cancer (38–40).
It is of great importance to obtain a more comprehensive proteomic
profile of the plasma exosomes in ovarian cancer, when developing
some novel diagnostic and prognostic biomarkers based on exosomes.
Furthermore, investigating the protein composition of exosomes
would supplement the understanding of the mechanism of their
biogenesis and the functional roles in this most lethal
gynecological malignancy.
In this study, exosomes were enriched from EOC
patient plasma with spherical morphology and mostly 30–100 nm in
diameter. TEM, NTA and DLS were also utilized to confirm the size,
concentration and size distribution of the exosomes. Western blot
analysis was performed to examine the expression of typical
exosomal proteins, including TSG101 and CD81. The high purity of
the exosomes was not only ensured during isolation procedures, but
was also verified by an endoplasmic reticular marker, calnexin,
which was not observed in exosome stains. Based on these
multifaceted characterizations, plasma exosomes with a high purity
were obtained, which laid the foundation for the subsequent
proteomics analysis. In total, 262 exosomal proteins were detected
from EOC and non-cancerous samples by performing an LC-MS/MS in
combination with TMT, which is the first ovarian cancer exosome
study based on this technique, at least to the best of our
knowledge. A total of 50 DEGs were screened out with distinct
annotation to cellular components, biological process, and
molecular function respectively. Among these, 19 genes were plotted
in the PPI network to explain their interaction and involved
signaling pathways. Two particular categories of pathway should be
paid more attention to, including coagulation and apoptosis-related
pathways. In this study, we demonstrated the procoagulation
activity of exosomes derived from EOC patient plasma. The
bioinformatics analysis provided an explanation for
hypercoagulation that occurs in patients with EOC at the molecular
level. FGG, FGA, C9, C4BPA, C4BPB and C4A, as well as other genes
were found to be relevant to coagulation in this study.
Among all DEGs, 4 genes, namely GSN, FGG, LBP and
FGA were selected as diagnostic markers. FGA ought to be the most
promising diagnostic biomarker based on the results of ROC
analysis. FGA has been found to serve as a diagnostic marker in
several studies. For instance, Davalieva et al reported the
non-invasive biomarkers in urine with higher specificity than
prostate-specific antigen (PSA) in patients with prostate cancer; a
number of genes, including FGG and FGA were observed to be present
with different abundances, which could be potential biomarker
candidates (41). In another
study, the serum levels of the proteins, alpha 2-HS glycoprotein
(AHSG), FGA and apolipoprotein A1 (APOA-I), were identified as
diagnostic biomarkers for gastric cancer (GC); ELISA data suggested
that the serum levels of FGA, AHSG and APOA-I in patients with GC
differed significantly as compared to the healthy volunteers;
moreover, the serum levels of these three proteins were associated
with TNM stages and could reflect tumor burden (42). The FGG gene mediates the formation
of the fibrinogen γ chain, a subunit of the fibrinogen protein.
This protein is important for coagulation, which is needed to
attenuate excessive bleeding following injury (43). Besides being the diagnostic
factors, FGA and FGG have also been proven to serve as novel
prognostic biomarkers for GC. The elevated expression of FGA and
FGG predicted poorer prognosis of GC patients according to the
survival analysis (44).
In prostate cancer (PC), GSN has also been
demonstrated to interact with androgen receptor (AR) in a
dose-dependent manner, which can enhance the bioactivity of AR. The
blockage of the interaction between GSN and AR may thus emerge as a
promising therapeutic target for the treatment of PC (45). Another study focused on the
prognostic value of cytoskeleton-associated proteins for ovarian
cancer (46). In silico analysis
was performed using the cancer genome atlas (TCGA) and 17
cytoskeletal proteins, including GSN, playing a role in tumor
progression (46). GSN has also
been shown to be involved in TGF-β1 induced epithelial to
mesenchymal transition (EMT) in breast cancer (47). An increased GSN expression was
shown to result in alterations of cell proliferation and cell
cycle, due to the modification of the actin filament assembly by
GSN. The expression of typical markers for EMT process, such as
N-cadherin, E-cadherin, and vimentin was altered by the silencing
of GSN, suggesting that TGF-β1 triggered aberrant expression of GSN
could affect the EMT process in breast cancer cells (47). A high level of GSN was found to be
involved in chemoresistance in gynecological cancer cells compared
with their sensitive cell types. Cisplatin-induced GSN
downregulation is associated with its cleavage and apoptosis. In
resistant cells, GSN was highly expressed and cisplatin failed to
abolish the downstream interaction, leading to the attenuated
apoptosis. In addition, the overexpression of GSN was closely
associated with a more aggressive behavior and a shorter OS, as
well as PFS. These findings are in agreement with the notion that
GSN is a crucial role in chemosensitivity in gynecological cell,
which could be exploited as diagnostic and prognostic markers
(48).
LBP has been studied in other types of cancer to
explore its potential as the diagnostic and/or prognostic
biomarkers. An increased level of LBP has been shown to be closely
associated with colorectal (CRC) and gallbladder cancer (49,50).
Gut microbiota dysbiosis impairs the intestinal barrier function
and elevates plasma lipopolysaccharide levels, thereby promoting
endotoxemia and contributing to the development of CRC. The
reduction in plasma LBP levels may be a crucial parameter for
patients newly diagnosed with CRC (51). Additionally, LBP may also be useful
in renal cancer (52). Cox
regression analysis was performed to explore the survival in
association with age, sex, clinicopathological parameters and LBP
expression. A high expression of LBP indicates conventional renal
cell carcinoma patients at a high risk of post-operative
progression that may require optimized active surveillance and
prompt regime (52). Circulating
exosomes containing LBP emerged as a biomarker for non-small cell
lung cancer (NSCLC) (53). LBP was
found to be well distinguished between patients with metastatic and
patients with non-metastatic NSCLC, implying LBP as a promising and
effective indicator of metastatic NSCLC (53).
Due to the heterogeneity and frequent relapse of
ovarian cancer, it is of great importance to discover biomarkers
for early diagnosis of ovarian cancer. The study by Liang et
al demonstrated that the exosomal proteins were highly enriched
in signaling pathways associated with carcinogenesis. A number of
proteins were overexpressed in both tissue and exosomes, including
tubulin beta-3 chain (TUBB3), epithelial cell surface antigen
(EpCAM), claudin 3 (CLDN3), proliferation cell nuclear antigen
(PCNA), epidermal growth factor receptor (EGFR), apolipoprotein E
(APOE), fatty acid synthase (FASN), etc., which may serve as
potential diagnostic markers and therapeutic targets for the
treatment of ovarian cancer (38).
In one study, EpCAM was demonstrated to be associated with the
remote metastases in advanced endometrial cancer (54). In clinical practice, however, EpCAM
was not a conventional biomarker to be evaluated, implying that the
sensitivity and the specificity for ovarian cancer is not
sufficient enough. A more promising way is to discover other
biomarkers with high sensitivity and specificity.
Apart from ovarian cancer, proteomics analysis has
been conducted in a number of other types of cancer. In one study,
exosomes were enriched from the serum of patients with prostate
cancer and proteomic profiling was performed using LC-MS/MS so as
to reveal distinct proteins across different ethnicities (55). Large quantities of novel proteins
were discovered that appeared to be ethnicity-specific in prostate
cancer, including Iroquois homeobox protein 5 (IRX5), mitochondrial
tumor suppressor 1 isoform 4 (MTS1) and trinucleotide repeat
containing 6B isoform 3 (TNR6B). The purpose of that study was to
find drug targets for the prostate cancer patients with particular
ethnicity. However, those authors failed to recruit a validation
cohort to verify the feasibility of these potential biomarkers
which should be further improved (55).
Furthermore, proteomics analysis has also been
performed for the identification of biomarkers in exosomes derived
from patients with pancreatic cancer. An et al (56) reported the proteomic results of
exosomes from the sera of ten patients with locally advanced
pancreatic cancer. The quantitative analysis was conducted using
the iTRAQ method. Each sample contained 700 to 800 exosomal
proteins, several of which may be relevant to metastasis and
chemoresistance of pancreatic cancer. The exosomes at various time
points were also collected to compare the alterations of proteomic
profiles during the treatment period. In total, 8 proteins were
identified to show universal treatment-specific changes, supporting
the importance of tumor-derived exosomes in pancreatic cancer
progression and metastasis. OBSL1 and PLF4 displayed the highest
treatment response among 8 proteins, which could be favorable
candidates for biomarker development (56).
Recently, Chen et al (57) presented a quantitative proteomics
analysis of exosomes purified from the colorectal cancer (CRC)
patient serum samples. A total of 918 proteins were detected and
725 of these were found in the Exocarta proteins list. In
comparison with normal volunteers, 36 proteins were upregulated and
22 proteins were downregulated in the serum exosomes of patients
with CRC. Bioinformatics analysis revealed that upregulated
proteins were involved in processes relevant to metastasis; while
the downregulated proteins mainly contributed to tumorigenesis and
cell survival (57). Some of these
differently expressed proteins may be promising diagnostic and/or
prognostic indicators; however, no further verification analysis
was performed to establish the association between clinical
characteristics and the proteomics results, which should be further
elucidated for the development and application in clinical
practice.
Notably, this study covered several important
clinical needs in ovarian cancer. On the one hand, this study is
the first (to the best of our knowledge) to propose exosomal GSN,
FGG, FGA and LBP to be the diagnostic biomarkers of ovarian cancer,
which was verified and validated by the ELISA. The diagnostic
molecular signature identified in exosomes would perfectly
supplement the current diagnostic procedure based on the tissue
samples, reducing the number of invasive biopsies needed. This
proteomics approach paves the way for the identification of
proteomic signatures in exosomes that are more relevant to clinical
characteristics. On the other hand, from the Kaplan Meier-plotter,
we found the promising prognostic values of FGG and LBP. However,
the survival results from that website were concluded based on the
gene expression in tissue samples, but not in exosomes, attenuating
the reliability of exosomal FGG and LBP as the potential prognostic
biomarkers. Therefore, the survival data should be summarized
according to the exosomal expression of FGG and LBP in future
studies, which could complement the limitations of the current
study. In fact, the patients have been under observations to
acquaint their updated follow-up statuses. Upon completion, the
survival curves will be constructed to enhance the values of
exosomal FGG and LBP as the prognostic factors. Given that ovarian
cancer is closely associated with the high challenge of metastatic
and chemoresistant characteristics, our exosomal proteomic results
may provide some new aspects and directions for future studies. Due
to the limited sample size of this study, further research should
be further conducted to reveal more prevalent and insightful
markers. Additionally, we only explored the diagnostic and
prognostic potentials of the individual exosomal protein. The
combined utilization of our findings and clinical markers, such as
CA125, could emerge as an encouraging perspective for a more
in-depth investigation.
In conclusion, in this study, we successfully
isolated and purified exosomes from plasma in patients with EOC,
which were characterized by various approaches to prove the high
quality of the isolated exosomes. To the best of our knowledge,
this study is the first to use LC-MS/MS combined with TMT for the
proteomics analysis of ovarian cancer-derived exosomes. The
subsequent GO analysis, KEGG pathway and PPI network described the
mutual interactions of DEGs, indicating that some of DEGs were
involved in the mediation of coagulation cascade relevant to
hypercoagulable state in patients with EOC. The procoagulation
assay also proved that exosomes enriched from EOC patient plasma
could enhance and accelerate the coagulation process. In total, 4
genes emerged as promising diagnostic and 2 as potential prognostic
indicators. The common proteins associated with exosome biogenesis
may provide some new information on understanding the mechanisms of
exosome secretion in ovarian cancer. Further studies should be
conducted to validate some candidate prognostic markers in one
separate cohort of patients and confirm the functional roles of
exosomes in the malignant disease. More patients should be included
to reduce bias and enhance the reproducibility and reliability of
the study, particularly for proteomic analysis. Limited sample size
may compromise the robustness.
Funding
This study was supported by the National Natural
Science Foundation of China (no. 81602270).
Availability of data and materials
The datasets and materials used and/or analyzed in
this study are available from the corresponding author on
reasonable request.
Authors' contributions
WZ and XO conceived and designed, and conducted and
drafted the manuscript. XW revised the initial design of the study,
incorporated the ELISA for validation, and provided suggestions and
solutions to the difficulties encountered in the project. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The present study and experimental procedures were
approved by the Human Research Ethics Committee of Fudan University
Shanghai Cancer Center (Shanghai, China). Written informed consent
was obtained from the patients or patients' families.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Special and sincere thanks were given to Polaris
Biology (Shanghai, China) for their heartful and prompt assistance
in this study.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jelovac D and Armstrong DK: Recent
progress in the diagnosis and treatment of ovarian cancer. CA
Cancer J Clin. 61:183–203. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Seidman JD, Horkayne-Szakaly I, Haiba M,
Boice CR, Kurman RJ and Ronnett BM: The histologic type and stage
distribution of ovarian carcinomas of surface epithelial origin.
Int J Gynecol Pathol. 23:41–44. 2004. View Article : Google Scholar
|
|
4
|
Melo SA, Luecke LB, Kahlert C, Fernandez
AF, Gammon ST, Kaye J, LeBleu VS, Mittendorf EA, Weitz J, Rahbari
N, et al: Glypican-1 identifies cancer exosomes and detects early
pancreatic cancer. Nature. 523:177–182. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hoshino A, Costa-Silva B, Shen TL,
Rodrigues G, Hashimoto A, Tesic Mark M, Molina H, Kohsaka S, Di
Giannatale A, Ceder S, et al: Tumour exosome integrins determine
organotropic metastasis. Nature. 527:329–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Riches A, Campbell E, Borger E and Powis
S: Regulation of exosome release from mammary epithelial and breast
cancer cells - a new regulatory pathway. Eur J Cancer.
50:1025–1034. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Skogberg G, Lundberg V, Berglund M,
Gudmundsdottir J, Telemo E, Lindgren S and Ekwall O: Human thymic
epithelial primary cells produce exosomes carrying
tissue-restricted antigens. Immunol Cell Biol. 93:727–734. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pitt JM, Charrier M, Viaud S, André F,
Besse B, Chaput N and Zitvogel L: Dendritic cell-derived exosomes
as immunotherapies in the fight against cancer. J Immunol.
193:1006–1011. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lugini L, Cecchetti S, Huber V, Luciani F,
Macchia G, Spadaro F, Paris L, Abalsamo L, Colone M, Molinari A, et
al: Immune surveillance properties of human NK cell-derived
exosomes. J Immunol. 189:2833–2842. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cai Z, Yang F, Yu L, Yu Z, Jiang L, Wang
Q, Yang Y, Wang L, Cao X and Wang J: Activated T cell exosomes
promote tumor invasion via Fas signaling pathway. J Immunol.
188:5954–5961. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Barrès C, Blanc L, Bette-Bobillo P, André
S, Mamoun R, Gabius HJ and Vidal M: Galectin-5 is bound onto the
surface of rat reticulocyte exosomes and modulates vesicle uptake
by macrophages. Blood. 115:696–705. 2010. View Article : Google Scholar
|
|
12
|
Xiao H, Lässer C, Shelke GV, Wang J,
Rådinger M, Lunavat TR, Malmhäll C, Lin LH, Li J, Li L, et al: Mast
cell exosomes promote lung adenocarcinoma cell proliferation - role
of KIT-stem cell factor signaling. Cell Commun Signal.
12:642014.PubMed/NCBI
|
|
13
|
Welton JL, Khanna S, Giles PJ, Brennan P,
Brewis IA, Staffurth J, Mason MD and Clayton A: Proteomics analysis
of bladder cancer exosomes. Mol Cell Proteomics. 9:1324–1338. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Choi DS, Lee JM, Park GW, Lim HW, Bang JY,
Kim YK, Kwon KH, Kwon HJ, Kim KP and Gho YS: Proteomic analysis of
microvesicles derived from human colorectal cancer cells. J
Proteome Res. 6:4646–4655. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xiao GY, Cheng CC, Chiang YS, Cheng WT,
Liu IH and Wu SC: Exosomal miR-10a derived from amniotic fluid stem
cells preserves ovarian follicles after chemotherapy. Sci Rep.
6:231202016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
McKiernan J, Donovan MJ, O'Neill V,
Bentink S, Noerholm M, Belzer S, Skog J, Kattan MW, Partin A,
Andriole G, et al: A novel urine exosome gene expression assay to
predict high-grade prostate cancer at initial biopsy. JAMA Oncol.
2:882–889. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Andre F, Schartz NE, Movassagh M, Flament
C, Pautier P, Morice P, Pomel C, Lhomme C, Escudier B, Le Chevalier
T, et al: Malignant effusions and immunogenic tumour-derived
exosomes. Lancet. 360:295–305. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Inal JM, Kosgodage U, Azam S, Stratton D,
Antwi-Baffour S and Lange S: Blood/plasma secretome and
microvesicles. Biochim Biophys Acta. 1834:2317–2325. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Severino V, Dumonceau JM, Delhaye M, Moll
S, Annessi-Ramseyer I, Robin X, Frossard JL and Farina A:
Extracellular vesicles in bile as markers of malignant biliary
stenoses. Gastroenterology. 153:495–504e8. 2017. View Article : Google Scholar
|
|
20
|
Sharma S, Gillespie BM, Palanisamy V and
Gimzewski JK: Quantitative nanostructural and single-molecule force
spectroscopy biomolecular analysis of human-saliva-derived
exosomes. Langmuir. 27:14394–14400. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lau C, Kim Y, Chia D, Spielmann N, Eibl G,
Elashoff D, Wei F, Lin YL, Moro A, Grogan T, et al: Role of
pancreatic cancer-derived exosomes in salivary biomarker
development. J Biol Chem. 288:26888–26897. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Menay F, Herschlik L, De Toro J, Cocozza
F, Tsacalian R, Gravisaco MJ, Di Sciullo MP, Vendrell A, Waldner CI
and Mongini C: Exosomes isolated from ascites of T-cell
lymphoma-bearing mice expressing surface CD24 and HSP-90 induce a
tumor-specific immune response. Front Immunol. 8:2862017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Purushothaman A, Bandari SK, Liu J, Mobley
JA, Brown EE and Sanderson RD: Fibronectin on the surface of
myeloma cell-derived exosomes mediates exosome-cell interactions. J
Biol Chem. 291:1652–1663. 2016. View Article : Google Scholar :
|
|
24
|
Wen SW, Sceneay J, Lima LG, Wong CS,
Becker M, Krumeich S, Lobb RJ, Castillo V, Wong KN, Ellis S, et al:
The biodistri-bution and immune suppressive effects of breast
cancer-derived exosomes. Cancer Res. 76:6816–6827. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Taylor DD and Gercel-Taylor C: MicroRNA
signatures of tumor-derived exosomes as diagnostic biomarkers of
ovarian cancer. Gynecol Oncol. 110:13–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Peng P, Yan Y and Keng S: Exosomes in the
ascites of ovarian cancer patients: Origin and effects on
anti-tumor immunity. Oncol Rep. 25:749–762. 2011.
|
|
27
|
Li J, Sherman-Baust CA, Tsai-Turton M,
Bristow RE, Roden RB and Morin PJ: Claudin-containing exosomes in
the peripheral circulation of women with ovarian cancer. BMC
Cancer. 9:2442009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dayon L, Hainard A, Licker V, Turck N,
Kuhn K, Hochstrasser DF, Burkhard PR and Sanchez JC: Relative
quantification of proteins in human cerebrospinal fluids by MS/MS
using 6-plex isobaric tags. Anal Chem. 80:2921–2931. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Thompson A, Schäfer J, Kuhn K, Kienle S,
Schwarz J, Schmidt G, Neumann T, Johnstone R, Mohammed AK and Hamon
C: Tandem mass tags: A novel quantification strategy for
comparative analysis of complex protein mixtures by MS/MS. Anal
Chem. 75:1895–1904. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar
|
|
31
|
Szklarczyk D, Franceschini A, Wyder S,
Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos
A, Tsafou KP, et al: STRING v10: Protein-protein interaction
networks, integrated over the tree of life. Nucleic Acids Res 43D:.
D447–D452. 2015.
|
|
32
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gyorffy B, Lánczky A and Szállási Z:
Implementing an online tool for genome-wide validation of
survival-associated biomarkers in ovarian-cancer using microarray
data from 1287 patients. Endocr Relat Cancer. 19:197–208. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wright M: Nanoparticle tracking analysis
for the multiparameter characterization and counting of
nanoparticle suspensions. Methods Mol Biol. 906:511–524.
2012.PubMed/NCBI
|
|
36
|
Chen X, Zhou J, Li X and Wang X, Lin Y and
Wang X: Exosomes derived from hypoxic epithelial ovarian cancer
cells deliver microRNAs to macrophages and elicit a tumor-promoted
phenotype. Cancer Lett. 435:80–91. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Matsuo K, Hasegawa K, Yoshino K, Murakami
R, Hisamatsu T, Stone RL, Previs RA, Hansen JM, Ikeda Y, Miyara A,
et al: Venous thromboembolism, interleukin-6 and survival outcomes
in patients with advanced ovarian clear cell carcinoma. Eur J
Cancer. 51:1978–1988. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liang B, Peng P, Chen S, Li L, Zhang M,
Cao D, Yang J, Li H, Gui T, Li X, et al: Characterization and
proteomic analysis of ovarian cancer-derived exosomes. J
Proteomics. 80:171–182. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yi H, Zheng X, Song J, Shen R, Su Y and
Lin D: Exosomes mediated pentose phosphate pathway in ovarian
cancer metastasis: A proteomics analysis. Int J Clin Exp Pathol.
8:15719–15728. 2015.
|
|
40
|
Sinha A, Ignatchenko V, Ignatchenko A,
Mejia-Guerrero S and Kislinger T: In-depth proteomic analyses of
ovarian cancer cell line exosomes reveals differential enrichment
of functional categories compared to the NCI 60 proteome. Biochem
Biophys Res Commun. 445:694–701. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Davalieva K, Kiprijanovska S, Komina S,
Petrusevska G, Zografska NC and Polenakovic M: Proteomics analysis
of urine reveals acute phase response proteins as candidate
diagnostic biomarkers for prostate cancer. Proteome Sci. 13:22015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shi F, Wu H, Qu K, Sun Q, Li F, Shi C, Li
Y, Xiong X, Qin Q, Yu T, et al: Identification of serum proteins
AHSG, FGA and APOA-I as diagnostic biomarkers for gastric cancer.
Clin Proteomics. 15:182018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tokutomi K, Tagawa T, Korenaga M, Chiba M,
Asai T, Watanabe N, Takeoka S, Handa M, Ikeda Y and Oku N:
Decoration of fibrinogen γ-chain peptide on adenosine
diphosphate-encapsulated liposomes enhances binding of the
liposomes to activated platelets. Int J Pharm. 407:151–157. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Duan S, Gong B, Wang P, Huang H, Luo L and
Liu F: Novel prognostic biomarkers of gastric cancer based on gene
expression microarray: COL12A1, GSTA3, FGA and FGG. Mol Med Rep.
18:3727–3736. 2018.PubMed/NCBI
|
|
45
|
Nishimura K, Ting HJ, Harada Y, Tokizane
T, Nonomura N, Kang HY, Chang HC, Yeh S, Miyamoto H, Shin M, et al:
Modulation of androgen receptor transactivation by gelsolin: A
newly identified androgen receptor coregulator. Cancer Res.
63:4888–4894. 2003.PubMed/NCBI
|
|
46
|
Schiewek J, Schumacher U, Lange T, Joosse
SA, Wikman H, Pantel K, Mikhaylova M, Kneussel M, Linder S,
Schmalfeldt B, et al: Clinical relevance of cytoskeleton associated
proteins for ovarian cancer. J Cancer Res Clin Oncol.
144:2195–2205. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen ZY, Wang PW, Shieh DB, Chiu KY and
Liou YM: Involvement of gelsolin in TGF-beta 1 induced epithelial
to mesenchymal transition in breast cancer cells. J Biomed Sci.
22:902015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Abedini MR, Wang PW, Huang YF, Cao M, Chou
CY, Shieh DB and Tsang BK: Cell fate regulation by gelsolin in
human gynecologic cancers. Proc Natl Acad Sci USA. 111:14442–14447.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chen R, Luo FK, Wang YL, Tang JL and Liu
YS: LBP and CD14 polymorphisms correlate with increased colorectal
carcinoma risk in Han Chinese. World J Gastroenterol. 17:2326–2331.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Van Dyke AL, Kemp TJ, Corbel AF, Zhu B,
Gao YT, Wang BS, Rashid A, Shen MC, Hildesheim A, Hsing AW, et al:
Lipopolysaccharide-pathway proteins are associated with gallbladder
cancer among adults in Shanghai, China with mediation by systemic
inflammation. Ann Epidemiol. 26:704–709. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
González-Sarrías A, Núñez-Sánchez MA,
Ávila-Gálvez MA, Monedero-Saiz T, Rodríguez-Gil FJ, Martínez-Díaz
F, Selma MV and Espín JC: Consumption of pomegranate decreases
plasma lipopolysaccharide-binding protein levels, a marker of
metabolic endotoxemia, in patients with newly diagnosed colorectal
cancer: A randomized controlled clinical trial. Food Funct.
9:2617–2622. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kovacs G, Peterfi L, Farkas N, Javorhazy
A, Pusztai C and Szanto A: Expression of inflammatory
lipopolysaccharide binding protein (LBP) predicts the progression
of conventional renal cell carcinoma - a short report. Cell Oncol
(Dordr). 40:651–656. 2017. View Article : Google Scholar
|
|
53
|
Wang N, Song X, Liu L, Niu L, Wang X, Song
X and Xie L: Circulating exosomes contain protein biomarkers of
metastatic non-small-cell lung cancer. Cancer Sci. 109:1701–1709.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Hsu YT, Osmulski P, Wang Y, Huang YW, Liu
L, Ruan J, Jin VX, Kirma NB, Gaczynska ME and Huang TH:
EpCAM-regulated transcription exerts influences on nanomechanical
properties of endometrial cancer cells that promote
epithelial-to-mesenchymal transition. Cancer Res. 76:6171–6182.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Turay D, Khan S, Diaz Osterman CJ, Curtis
MP, Khaira B, Neidigh JW, Mirshahidi S, Casiano CA and Wall NR:
Proteomic profiling of serum-derived exosomes from ethnically
diverse prostate cancer patients. Cancer Invest. 34:1–11. 2016.
View Article : Google Scholar :
|
|
56
|
An M, Lohse I, Tan Z, Zhu J, Wu J,
Kurapati H, Morgan MA, Lawrence TS, Cuneo KC and Lubman DM:
Quantitative proteomic analysis of serum exosomes from patients
with locally advanced pancreatic cancer undergoing
chemoradiotherapy. J Proteome Res. 16:1763–1772. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen Y, Xie Y, Xu L, Zhan S, Xiao Y, Gao
Y, Wu B and Ge W: Protein content and functional characteristics of
serum-purified exosomes from patients with colorectal cancer
revealed by quantitative proteomics. Int J Cancer. 140:900–913.
2017. View Article : Google Scholar
|