Introduction
Breast cancer is the most frequent type of cancer
among women and is the second leading cause of cancer-associated
mortality in the female population of the United States (1). In the last two decades, the prognosis
of patients with breast cancer has improved due to lifestyle
modification, early detection, prophylactic mastectomy and advanced
therapies (2). However, the
curative effects of conventional chemotherapeutic agents are not
satisfactory, and certain patients with advanced-stage breast
cancer exhibit resistance to existing chemotherapeutic drugs
(3). The risk factors of breast
cancer are multiple and complicated, and the molecular mechanisms
underlying its pathogenesis remain to be elucidated. Therefore,
there has been increasing interest in identifying alternative
chemotherapeutic drugs with novel anticancer mechanisms.
Harmine (HM) is a β-carboline alkaloid that was
originally isolated from seeds of Peganum harmala and
Banisteriopsis caapi in 1847 (4). HM is widely distributed in various
medicinal plants and has long been used in folk medicine in the
Middle East and Asia (5). Studies
have demonstrated that HM exhibits significant antitumor activities
in vitro and in vivo (6), including inhibiting proliferation
(6), migration (7) and invasion (8), promoting apoptosis (9) and preventing tumorigenesis. HM
inhibits the growth of several types of cancer, including lung
(10), gastric (9), breast (11) and hepatic cancer (12). It arrests the cell cycle at the
G0/G1 phase (13) and decreases
the cyclin-dependent kinase activity (14). HM induces autophagy and apoptosis
through the protein kinase B (Akt)/mammalian target of rapamycin
(mTOR) and extracellular signal-regulated kinase (ERK)1/2 signaling
pathways, increases the expression of pro-apoptotic factors,
including P53, caspases 3/8/9, B-cell lymphoma 2 (Bcl-2)-associated
X protein (Bax), and BH3-interacting domain death agonist (Bid),
and reduces the level of pro-inflammatory cytokines, including
TNF-α, IL-6 and granulocyte-macrophage colony-stimulating factor,
in melanoma and gastric cancer (7). HM induces autophagy via increasing
LC3-II and downregulating P62 in a dose-dependent manner in B16
cells (15). It can also suppress
the expression of pro-metastatic genes, including matrix
metalloproteinase-9, ERK and vascular endothelial growth factors,
to inhibit melanoma cell invasion (16).
Transcriptional co-activator with PDZ-binding motif
(TAZ) was initially identified as a 14-3-3 binding protein
(17). Due to the lack of a
DNA-binding domain, TAZ is not a transcription factor, but it can
function as a transcriptional regulator though its transactivation
domain. In addition to its interaction with 14-3-3, it has been
reported to interact with multiple proteins, including thyroid
transcription factor-1 (18),
myogenic differentiation 1 (19),
Smads (20), core-binding factor
α1/Runt-related transcription factor 2 (21), transcriptional enhancer factor-1
(22), peroxisome
proliferator-activated receptor (23) and TEA domains (24). Serving as a transcriptional
coactivator, TAZ is important in osteoblastic, myogenic and
adipogenic differentiation (25).
TAZ was initially identified as an oncogenic protein in non-small
cell lung cancer in 2011 (26).
Accumulating studies have confirmed that the expression of TAZ is
elevated in various types of human cancer, including colorectal
cancer (27), glioblastoma
(28) and breast cancer (29,30).
The overexpression of TAZ leads to cancer cell behavior, including,
but not limited to, growth-factor-independent proliferation
(31) and resistance to
chemotherapeutics (32). It
promotes epithelial-mesenchymal transition (30), cell proliferation, migration,
invasion, tumorigenesis and tumor formation in xenograft models
(33), suggesting an oncogenic
role of TAZ in the development of human cancer. Studies have shown
that TAZ knockdown in MCF7 and Hs578T cells inhibited cell
migration and invasion. The knockdown of TAZ in MCF7 cells also
suppressed cell growth in vitro and in vivo (34). These data suggest that TAZ promotes
cell proliferation, migration and invasion in breast cancer.
Considering the crucial roles of TAZ in breast
cancer, the present study hypothesized that HM induces anticancer
activity in breast cancer through the regulation of TAZ. The
present study first showed that HM inhibited proliferation and
migration, and promoted apoptosis in breast cancer cell lines,
confirming its anticancer activity. Subsequently, it was found that
the overexpression of TAZ was able to rescue HM-induced cellular
toxicity. The results of the present study showed that HM exerted
anticancer activity through inhibiting the expression and activity
of TAZ.
Materials and methods
Preparation and chemical analysis of
HM
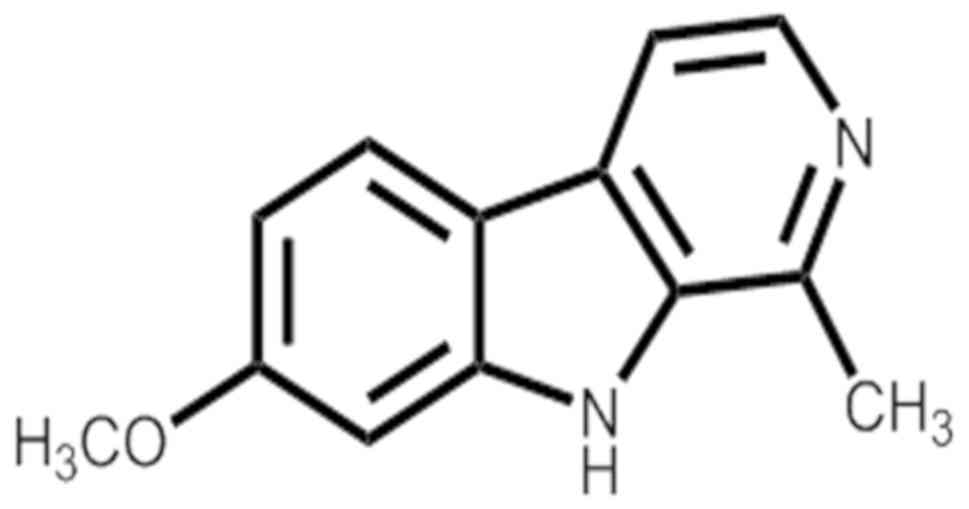
HM (Fig. 1) was
separated from the seeds of Peganum nigellastrum Bunge by
column chromatography on AB-8 Macroporous Resin eluted with
gradient ethyl alcohol and water. Furthermore, HM was isolated by
preparative high-performance liquid chromatography (Fig. S1). Structure identification and
the purity (>98%) of this compound (Figs. S2-8) were identified by
spectroscopic methods and compared with the reported spectral
data.
Cell culture and HM treatment
The MDA-MB-231 and MCF-7 human breast cancer cell
lines were obtained from the Shanghai Institute of Biochemistry and
Cell Biology (Shanghai, China). The cells were cultured in DMEM
(cat. no. C11965500BT; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% FBS (cat. no. 10100154; Gibco; Thermo
Fisher Scientific, Inc.) and 100 μg/ml streptomycin at 37°C
in an incubator with 5% CO2. A 200-mM stock solution of
HM was prepared in DMSO (cat. no. V900090; Merck KGaA, Darmstadt,
Germany) and diluted in culture media as necessary, to achieve the
final desired experimental concentrations.
Cell transfection
The pCMV6-TAZ expression vector and pCMV6-control
vector were purchased from OriGene Technologies, Inc. (Rockville,
MD, USA; cat. no. Rc204082). MDA-MB-231 and MCF-7 cells were
cultured in 96-well plates at a density of 5×103 cells per well for
24 h at 37°C, and then transfected with pCMV6-TAZ or pCMV6-control
using Lipofectamine 3000 (cat. no. L3000-015; Thermo Fisher
Scientific, Inc.) for 2 days. The transfected cells were visualized
using a fluorescence microscope.
CCK-8 assay
The cells were plated in 96-well plates at a density
of 5×103 cells per well. Following 24 h of incubation,
the cells were left untreated or treated with HM at 50, 100 or 150
μM for 24, 48 or 72 h, followed by the addition of 10
μl CCK-8 to each well. The absorbance was measured at 450 nm
using EnSpire Multimode plate reader (PerkinElmer, Inc., Waltham,
MA, USA). The optical density (OD) was calculated for cell
viability assay. Cell viability (%) = OD (treated) / OD (control)
×100%.
Wound healing assay
Cell migration was measured using a wound healing
assay. The MDA-MB-231 and MCF7 cells were cultured in 6-well
plates. A clean pipette tip was used to inflict a ‘wound’ when
cells formed a confluent monolayer, and the cultured cells in DMEM
were supplemented with 2% FBS. The cells were left untreated or
treated with HM at 50, 100 or 150 μM. Images of the wound
margins were captured using an inverted light microscope (Olympus
Corporation, Tokyo, Japan) formed 0 h at this time point. Following
incubation for 24 h, images of the same region of cells were
captured for measurement. The wound healing rate was calculated
using the following formula: (average wound margin in 0 h - average
wound margin in 24 h) / average wound margin in 0 h.
DAPI staining
The cells were left untreated or treated with HM at
50, 100 or 150 μM for 24 h, washed twice with phosphate
buffer saline (PBS), fixed in methanol for 10 min and finally
stained with DAPI (cat. no. C1002; Beyotime Institute of
Biotechnology, Haimen, China) for 10 min at room temperature.
Images were then captured by fluorescent microscopy (Olympus
Corporation).
Flow cytometry
Cell apoptosis was analyzed using a PE Annexin V
apoptosis detection kit І (cat. no. 559763; BD Biosciences, USA).
The cells were seeded in 6-well plates with 50, 100 and 150
μM HM for 24 h, and stained according to the manufacturer's
instructions. The apoptotic cells were immediately detected by a
FACS Caliber II Sorter and the Cell Quest FACS system (BD
Biosciences). Data were analyzed using FlowJo software (version
7.6.5; Tree Star, Inc., Ashland, OR, USA).
Western blotting
The cells were lysed in cell lysis buffer (cat. no.
P0013B; Beyotime Institute of Biotechnology) and centrifuged
(12,000 × g, 10 min, 4°C) to collect the supernatants following
treatment with HM (50, 100 or 150 μM) for 24 h. The protein
concentration was measured using a BCA Protein Assay kit (cat. no.
CW0014S; CWBio, Beijing, China), and an equal quantity (20
μg per well) of protein was separated by 10% SDS-PAGE and
transferred onto nitrocellulose membranes. The membranes were
blocked using 5% nonfat skim milk for 1 h at room temperature and
then incubated with primary antibodies overnight for 4°C. The
primary antibodies included anti-TAZ (1:1,000; cat. no. 70148; Cell
Signaling Technology, Inc., Danvers, MA, USA), anti-Bax (1:1,000;
cat. no. ab-32503; Abcam, Cambridge, MA, USA), anti-Bcl-2 (1:1,000;
cat. no. ab-32124; Abcam), anti-Erk (1:500; cat. no. sc-514302;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA),
anti-phosphorylated (p-)Erk (1:500; cat. no. sc-81492; Santa Cruz
Biotechnology, Inc.), anti-Akt (1:500; cat. no. sc-24500; Santa
Cruz Biotechnology, Inc.), anti-p-Akt (1:500; cat. no. sc-7985;
Santa Cruz Biotechnology, Inc.) and anti-β-actin (1:1,000; cat. no.
sc-47778; Santa Cruz Biotechnology, Inc.). Following washing with
TBST three times, for 10 min each time, goat anti-rabbit IgG
(1:10,000; cat. no. sc-2040; Santa Cruz Biotechnology, Inc.) or
goat anti-mouse IgG antibody (1:10,000; cat. no. sc-2005; Santa
Cruz Biotechnology, Inc.) conjugated with horseradish peroxidase
was incubated for 1 h at room temperature. The protein bands were
visualized using the enhanced ECL Select Western Blotting Detection
Reagent (cat. no. RPN2235; GE Healthcare Life Sciences, Chalfont,
UK).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted from untreated and treated
cells using TRIzol reagent (cat. no. 15596018; Thermo Fisher
Scientific, Inc.) and cDNA was synthesized using the ReverTra Ace
qPCR RT kit (cat. no. FSQ-101; Toyobo Life Science, Osaka, Japan).
The qPCR was performed using UltraSYBR mixture (cat. no. CW-2601;
CWBio) on an ABI StepOne Plus QPCR system (Thermo Fisher
Scientific, Inc.). The thermocycling steps were as follows: Initial
denaturation at 95°C for 10min, followed by 40 cycles at 95°C for
15 sec, 58°C for 1 min, and a final extension step at 72°C for 5
min. The 2-ΔΔCq method was used to calculate changes in
relative mRNA expression levels (35). The primer sequences used were as
follows: TAZ, forward 5'-GTCACCAACAGTAGCTCAGATC-3' and reverse
5'-AGT GATTACAGCCAGGTTAGAAAG-3'; β-actin, forward 5'-AC
TCTTCCAGCCTTCCTTCC-3' and reverse 5'-CGTCATACT CCTGCTTGCTG-3'.
Xenograft mice model
A total of 20 six-week-old male athymic nude BALB/c
mice (weight, 16-18 g) purchased from Beijing Vital River
Laboratory Animal Technology (Beijing, China) were housed under
specific pathogen-free conditions (24-26°C; 12-h light/dark cycle;
free access to food and water). The mice were subcutaneously
inoculated with MCF-7 cells (1×107) resuspended in 0.1
ml DMEM. When the tumor size reached 100 mm3, the mice
were randomly divided into four groups (n=5/group) to receive one
of the following treatments: Normal saline (NS; intraperitoneal
injection) or 20, 40 or 80 mg/kg/day of HM (5 days per week for 2
weeks, intraperitoneal injection). Tumor length (L) and width (W)
were measured at 2-day intervals using a Vernier caliper, and tumor
volume (V) was calculated using the following formula: V
(mm3) = L (mm) × W2 (mm2) × 0.5.
After 4 weeks injection, all mice were sacrificed and tumor tissues
were removed. Each mouse bore a single tumor, the maximum diameter
of a single subcutaneous tumor reached 15.98 mm, the maximum tumor
volume reached 1,189 mm3. Tumor tissues from the mice
were dissected for western blotting, RT-qPCR analysis and
immunohistochemistry (IHC). The animal experiments were approved by
the Committee on the Ethics of Animal Experiments of the Tongji
Medical College, Huazhong University of Science and Technology
(Wuhan, China; IACUC no. 837).
IHC analysis
IHC was performed to detect the levels of TAZ,
Bcl-2, Bax, p-Erk and p-Akt in mouse tumor tissues. The samples
were embedded in paraffin and sliced into thin sections (5 μm). The
sections were dewaxed in xylene and sequentially rehydrated with
100, 95, 80 and 75% ethanol. The slides were blocked with BSA for 2
h at room temperature. The slides were incubated with primary
antibodies anti-TAZ, Bcl-2, Bax, p-Erk and p-Akt (1:200) overnight
at 4°C. Following washing three times with PBS for 10 min each
time, biotinylated goat anti-rabbit secondary antibody (1:200; cat.
no. A0279; Beyotime Institute of Biotechnology) was incubated for 1
h at 37°C, and diaminobenzidine tetrachloride (cat. no. P0203;
Beyotime Institute of Biotechnology) was used for staining at 37°C
for 10 min. Data were analyzed using a light microscope (Olympus
Corporation).
Statistical analysis
Statistical analysis was performed by Student's
t-test for comparison between two groups or one-way ANOVA for
comparison between more than two groups. The least-significant
difference post hoc test was used following ANOVA. All statistical
analyses were performed using SPSS 17.0 software (SPSS, Inc.,
Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
HM inhibits proliferation and migration,
and induces apoptosis in MDA-MB-231 and MCF-7 cells
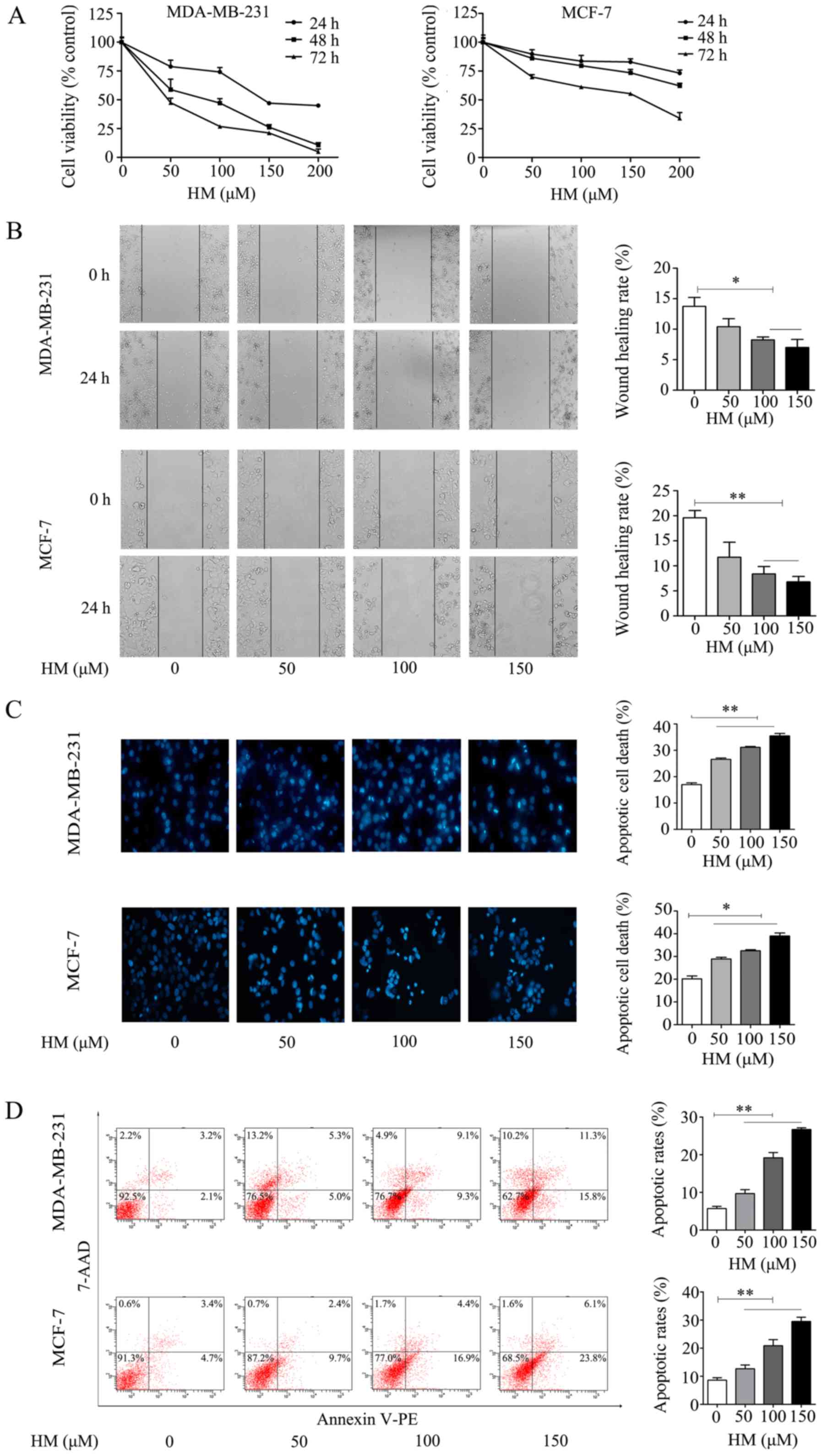
The anti-proliferative effect of HM on breast cancer
cells was evaluated via CCK-8 assay. The MDA-MB-231 and MCF-7 cells
were treated with different concentrations of HM for 24, 48 and 72
h, respectively. As shown in Fig.
2A, HM inhibited cell proliferation in a dose- and
time-dependent manner. In addition to inhibiting cell
proliferation, HM suppressed the migration of breast cancer cells.
The MDA-MB-231 and MCF-7 cells were exposed to different
concentrations of HM for 24 h, as shown in Fig. 2B, and the results revealed a
dose-dependent inhibition of cell migration. Apoptotic
morphological changes were determined by DAPI staining. Chromatin
condensation and nuclear fragmentation, which are characteristics
of apoptosis, were evident in the MDA-MB-231 and MCF-7 cells
treated with HM (50, 100 or 150 μM) for 24 h. As shown in
Fig. 2C, a significant increase of
chromatin condensation and nuclear fragmentation was observed in
the HM-treated cells, when compared with that of control (no HM
added) cells. To further confirm the pro-apoptotic effect of HM,
cell apoptosis was determined by flow cytometry using a Annexin V
apoptosis detection kit. Following treatment of the MDA-MB-231
cells with HM at 50, 100 or 150 μM for 24 h, the apoptotic
population increased from 5.7 to 26.7% in a dose-dependent manner.
Similar results were observed in MCF-7 cells (Fig. 2D). In combination, these results
indicated that HM had an anticancer effect on the breast cancer
cell lines.
HM decreases the expression level of TAZ
in MDA-MB-231 and MCF-7 cells
Increasing studies have suggested that TAZ is an
oncogene in human cancer cells and serves an important role in the
development of tumors (17). It
has been reported that TAZ is expressed at high levels in breast
cancer cell lines, promotes cell proliferation, migration and
tumorigenesis, and inhibits apoptosis in breast cancer (29,30,34).
To further understand the mechanism underlying the anticancer
activities induced by HM, the present study examined whether TAZ
was involved in the anticancer mechanism. The relative mRNA
expression of TAZ was significantly decreased, compared with that
in the control group (Fig. 3A).
Following 48 h of HM treatment (50, 100 or 150 μM), the
protein expression of TAZ was reduced in the MDA-MB-231 and MCF-7
cells. The activation of MAPK kinase/ERK and PI3K/AKT has been
reported to regulate cancer cell proliferation and migration
through various pathways (36,37).
In the present study, proliferation- and migration-related
proteins, including p-Erk and p-Akt, were decreased, whereas
anti-apoptotic protein Bcl-2 was decreased and pro-apoptotic Bax
was increased (Fig. 3B).
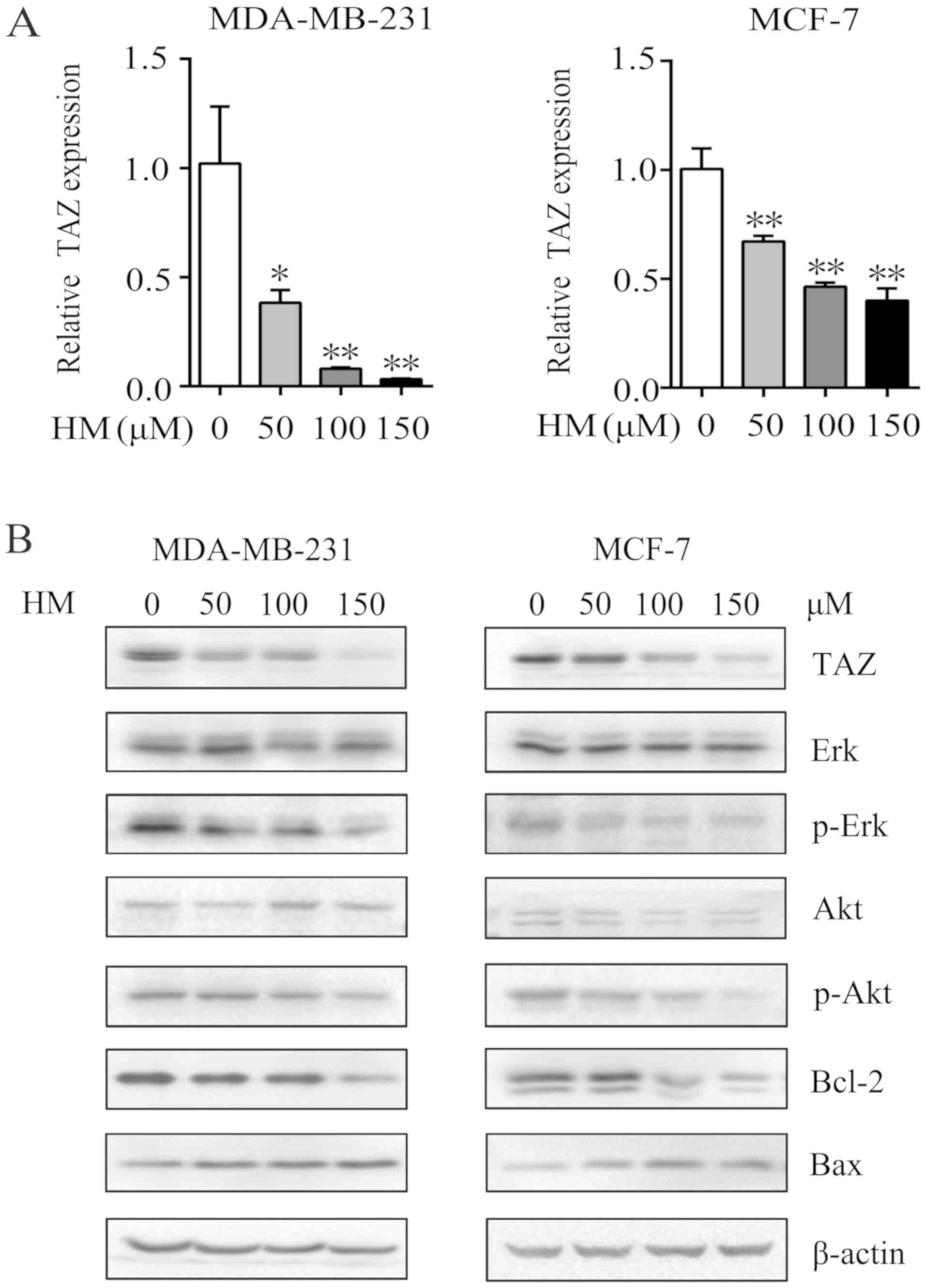 | Figure 3HM mediates antitumor effects via the
downregulation of TAZ in MDA-MB-231 and MCF-7 cells. (A) MDA-MB-231
and MCF-7 cells were treated with HM (50, 100 or 150 μM) for
24 h and the relative mRNA transcription levels were determined via
reverse transcription-quantitative polymerase chain reaction
analysis. (B) MDA-MB-231 and MCF-7 cells were treated with HM (50,
100 or 150 μM) for 48 h, and the expression of
proliferation, migration and apoptosis-related proteins, Akt,
p-Akt, Erk, p-Erk, Bcl-2 and Bax, were detected by western
blotting. β-actin was used as an internal standard. Data are
presented as the mean ± SD of three independent experiments.
*P<0.05 and **P<0.01 vs. 0 μM. HM,
harmine; TAZ, transcriptional co-activator with PDZ-binding motif;
Akt, protein kinase B; Erk, extracellular signal-regulated kinase;
Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X protein; p-,
phosphorylated. |
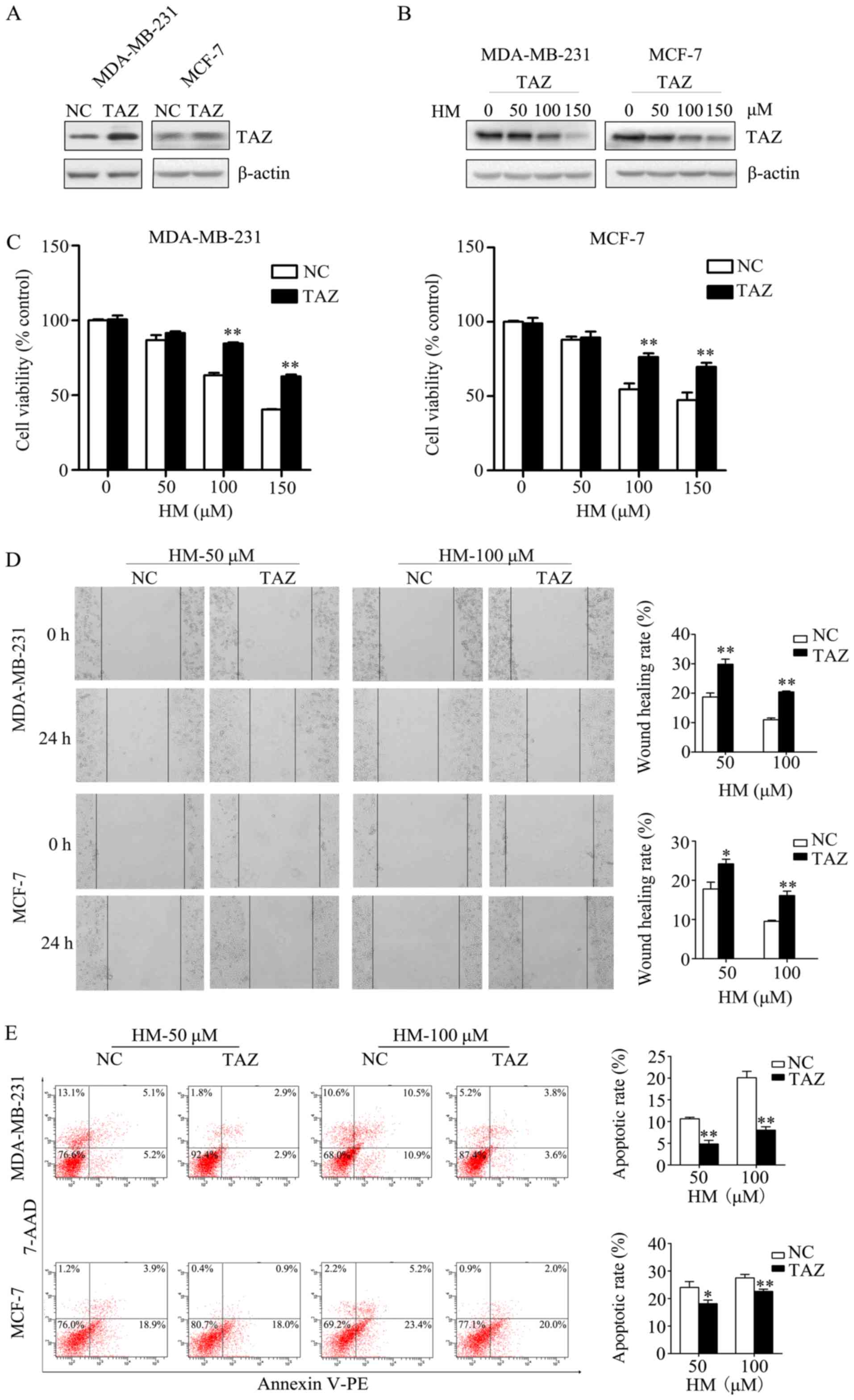
Overexpression of TAZ inhibits the
anticancer activity of HM
In order to validate that HM-induced anticancer
activity via targeting TAZ, the effect of increased TAZ on
HM-mediated anticancer activity was measured. The MDA-MB-231 and
MCF-7 cells were transfected with either a TAZ-expressing plasmid
(pCMV6-TAZ) or a control plasmid (pCMV6-control). At 2 days
post-transfection, western blotting was performed to check the
transfection efficiency of TAZ, the expression of TAZ was increased
in the TAZ overexpression group (Fig.
4A). The expression level of TAZ in the TAZ overexpression
group under HM treatment (50, 100 or 150 μM) was decreased
in a dose-dependent manner (Fig.
4B). The transfected cells were treated with HM (50, 100 or 150
μM) for 48 h. When compared with negative control cells, the
overexpression of TAZ in breast cancer cells was found to prevent
the HM-induced inhibition of cell proliferation and migration
(Fig. 4C and D). HM-induced cell
apoptosis was also inhibited by the overexpression of TAZ (Fig. 4E).
HM inhibits the growth of breast
xenograft tumors in vivo
The MCF-7 cells were subcutaneously injected into
athymic nude mice to establish a breast xenograft tumor model, and
tumor growth was recorded. Different doses of HM (20, 40 or 80
mg/kg/day) were administered, and HM was shown to reduce tumor
volume and weight in a dose- and time- dependent manner (Fig. 5A-D). Tumor tissues were used to
evaluate the level of TAZ by IHC, RT-qPCR and western blot
analyses. It was found that HM decreased the level of TAZ, compared
with that in the control group (Fig.
5E-G). The IHC results showed that the proliferation and
expression of metastasis-related proteins, including p-Akt and
p-Erk, were decreased compared with the control group (Fig. 5H). These results indicated that HM
inhibited the development of breast cancer in vivo.
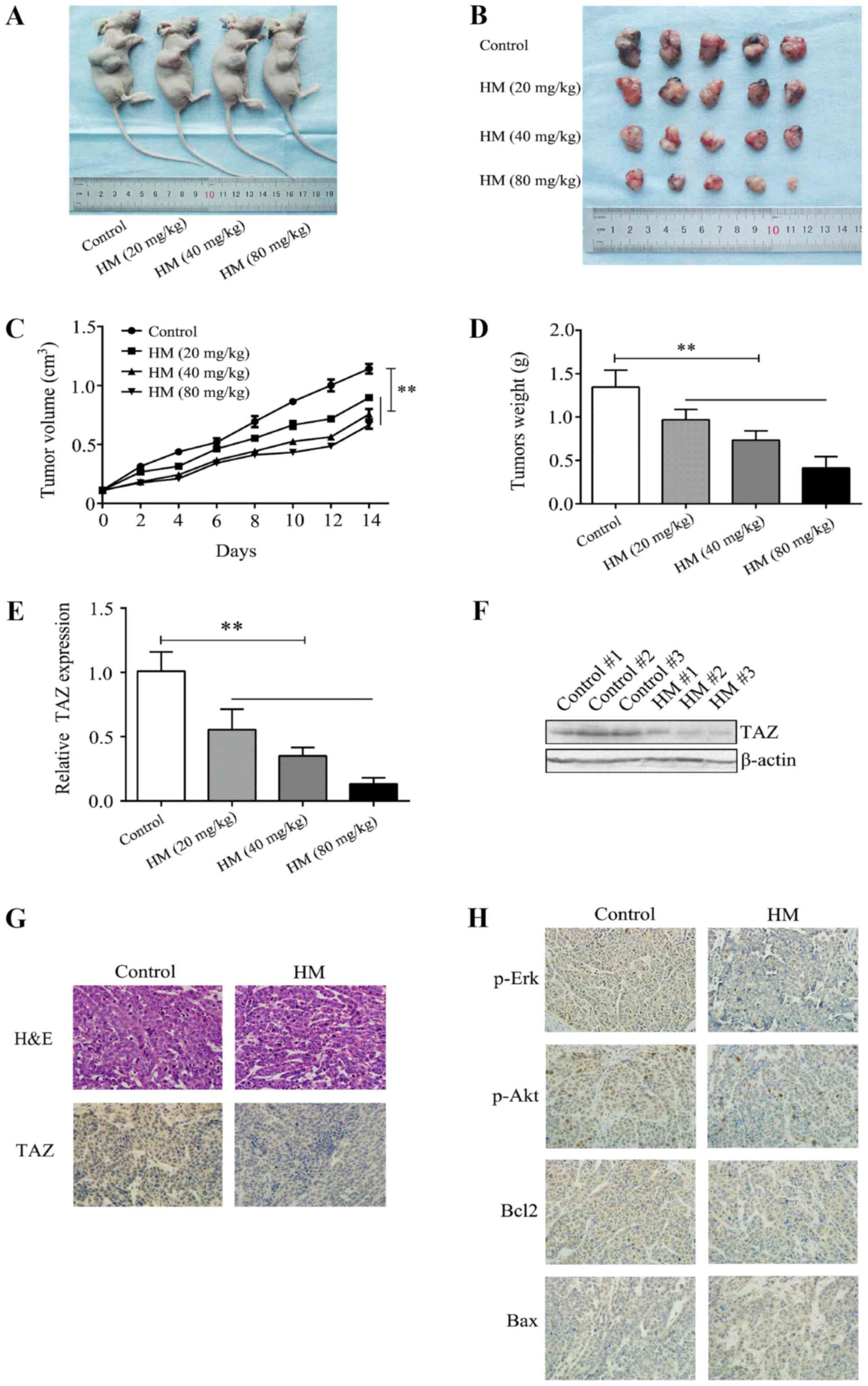 | Figure 5HM inhibits breast cancer tumor
growth in vivo. (A) Mice were sacrificed 2 weeks after being
injected with different doses of HM. (B) Tumors were dissected and
images were captured. (C) MCF-7 cells were injected into the nude
mice. Tumor volume was measured at 2-day intervals and the growth
curves were drafted. (D) Tumors were weighted at the end of
treatment. (E) Relative mRNA expression of TAZ in tumor tissues
from xenograft mice. (F) Representative images from western blot
analysis for TAZ protein expression in tumor tissues from xenograft
mice. (G and H) Tumor tissues were analyzed by H&E and
immunohistochemistry for TAZ, Bax, Bcl-2, p-Erk and p-Akt.
Magnification, ×400. **P<0.01, with comparisons
indicated by brackets. HM, harmine; H&E, hematoxylin and eosin;
TAZ, transcriptional co-activator with PDZ-binding motif; Akt,
protein kinase B; Erk, extracellular signal-regulated kinase;
Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associatedX protein; p-,
phosphorylated. |
Discussion
Although current therapeutic strategies can lead to
considerable anticancer activity, the development of novel
antitumor agents with less toxicity remains of interest. HM is a
natural β-carboline alkaloid isolated from Peganum harmala,
which was been previously used in folk medicine as an anticancer
therapy (5). Studies have shown
the anticancer activities of HM through the activation of apoptosis
and autophagy (38); however, the
effect of HM targeting TAZ in breast cancer has not been previously
reported. The present study demonstrated the anticancer effects of
HM on breast cancer and revealed a novel target of HM that is
important for its activity.
Previous studies have reported the antitumor effects
of HM on hepatocellular carcinoma (39), gastric cancer (9) and neuroblastoma (40). In the present study, the anticancer
activity of HM was demonstrated using MDA-MB-231 and MCF-7 cells
in vitro and MCF-7 cell-derived xenograft tumor in
vivo. In vitro, HM was found to inhibit proliferation
and migration in a dose- and time- dependent manner. Consistent
with this result, it was observed that HM significantly increased
cell apoptosis. Following treatment with HM, cellular morphological
changes, including cell rounding, shrinkage and karyopyknosis, were
observed. In vivo, HM also inhibited tumor growth, in terms
of tumor volume and weight, in a dose-dependent manner. These
results suggest that HM may be used in combination with other
chemical drugs for the inhibition of breast cancer growth and
metastasis.
To reveal the underlying molecular mechanism through
which HM induces anticancer activity in human breast cancer, the
expression of TAZ was investigated following HM treatment. The
Hippo pathway serves an important role in mammalian developmental
stages (41-44). TAO1-3, mammalian Ste20-like kinase
1/2, MAPK kinase kinase kinase 1-4/6/7, large tumor suppressor 1/2
and nuclear Dbf2-related 1/2 are the core kinases, whereas
yes-associated protein (YAP) and TAZ are the primary downstream
effectors of the Hippo pathway in mammals. When phosphorylated by
these core kinases, YAP is sequestered and degraded in the
cytoplasm (45). The Hippo pathway
coordinates cell proliferation and apoptosis in response to a
variety of signals by regulating the transcriptional coactivators
YAP and TAZ (46). Although it has
been reported that YAP functions as a tumor suppressor in breast
cancer (47), it has been
described that TAZ is an oncogenic protein in certain types of
human cancer, including non-small cell lung and breast cancer
(26,34). It was also found in the present
study that the level of TAZ was decreased in vitro following
treatment with HM, and that the expression of TAZ was reduced in
the MCF-7 xenograft tumor. These results revealed that HM may
function by targeting TAZ. To further verify this, experiments were
performed to reveal that the antiproliferative and pro-apoptotic
activities of HM were inhibited by overexpressing TAZ.
Furthermore, several growth factors associated with
proliferation and metabolism were examined in HM-treated MDA-MB-231
and MCF-7 cells. The PI3K/Akt pathway is commonly recognized for
its critical role in autophagy (48), metabolism (49) and metastasis (50). The elevated phosphory-lation of Akt
led to higher tumor recurrence and poorer overall survival
(51). Erk belongs to a subclass
of MAPKs that includes serine/threonine kinases. Upon activation of
the MAPK pathway, Erk is phosphorylated and regulates various
cellular processes, including proliferation, differentiation,
apoptosis and transformation (52). In the present study, it was found
that HM treatment increased the levels of p-Akt and p-Erk in
vitro and in vivo. The Bcl-2 family of proteins is
divided into two subfamilies; Bcl-2 and Bcl-xl are anti-apoptotic
proteins, whereas Bax, Bad and Bid are pro-apoptotic proteins
(53). The results of the present
study indicated a decreased level of Bcl-2 and increased level of
Bax in HM-treated cells, which is consistent with the
immunohistochemical results obtained from the MCF-7 xenograft
tumor. All the above results confirmed the antiproliferative and
pro-apoptotic function of HM on breast cancer.
In conclusion, the present study demonstrated the
anticancer (antiproliferative, antimetastatic and pro-apoptotic)
activity of HM on breast cancer cell lines. The study also
identified a novel target of HM, TAZ, in antagonizing the role of
HM on breast cancer cell lines, however, the anticancer molecular
mechanisms of HM require further in-depth investigation. Therefore,
HM may serve as an anticancer agent in the chemoprevention and
treatment of breast cancer, although future clinical and
pharmacological studies are required to confirm this.
Supplementary Materials
Funding
This study was partially sponsored by the National
Natural Science Foundation of China (grant no. 81301084 to CP;
grant no. 81800296 to SC), the Natural Science Foundation of Hubei
Province in China (grant no. 2017CFB357 to SC) and the Health
Commission Foundation of Hubei Province in China (grant no.
WJ2019M029 to JHu).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SC, CP, YD, JHe and JHu conceived and designed the
experiments. YD, JHe, TZ and GY performed the experiments. XS and
TY analyzed data. YD and JHe wrote the manuscript.
Ethics approval and consent to
participate
Animal studies were approved by the Committee on the
Ethics of Animal Experiments of the Tongji Medical College,
Huazhong University of Science and Technology (IACUC no. 837).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
HM
|
harmine
|
|
FBS
|
fetal bovine serum
|
|
MAPKs
|
mitogen-activated protein kinases
|
|
Bcl-2
|
B-cell lymphoma 2
|
|
Bax
|
Bcl-2-associated X protein
|
|
Akt
|
protein kinase B
|
|
Erk
|
extracellular signal-regulated protein
kinase
|
Acknowledgments
The authors would like to thank Mrs. Weiqun Chen
(Central Laboratory, The Central Hospital of Wuhan) for kindly
providing technical assistance in cell experiments and Mr.
Shunchang Zhou (Department of Experimental Zoology, Tongji Medical
College) for animal care.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chang L, Weiner LS, Hartman SJ, Horvath S,
Jeste D, Mischel PS and Kado DM: Breast cancer treatment and its
effects on aging. J Geriatr Oncol. 10:346–355. 2019. View Article : Google Scholar
|
|
3
|
Nathanson KL and Domchek SM: Therapeutic
approaches for women predisposed to breast cancer. Annu Rev Med.
62:295–306. 2011. View Article : Google Scholar
|
|
4
|
Patel K, Gadewar M, Tripathi R, Prasad SK
and Patel DK: A review on medicinal importance, pharmacological
activity and bioanalytical aspects of beta-carboline alkaloid
‘Harmine’. Asian Pac J Trop Biomed. 2:660–664. 2012. View Article : Google Scholar
|
|
5
|
Berrougui H, Martín-Cordero C, Khalil A,
Hmamouchi M, Ettaib A, Marhuenda E and Herrera MD: Vasorelaxant
effects of harmine and harmaline extracted from Peganum harmala L.
seeds in isolated rat aorta. Pharmacol Res. 54:150–157. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen Q, Chao R, Chen H, Hou X, Yan H, Zhou
S, Peng W and Xu A: Antitumor and neurotoxic effects of novel
harmine derivatives and structure-activity relationship analysis.
Int J Cancer. 114:675–682. 2005. View Article : Google Scholar
|
|
7
|
Zhang H, Sun K, Ding J, Xu H, Zhu L, Zhang
K, Li X and Sun W: Harmine induces apoptosis and inhibits tumor
cell proliferation, migration and invasion through down-regulation
of cyclo-oxygenase-2 expression in gastric cancer. Phytomedicine.
21:348–355. 2014. View Article : Google Scholar
|
|
8
|
Dai F, Chen Y, Song Y, Huang L, Zhai D,
Dong Y, Lai L, Zhang T, Li D, Pang X, et al: A natural small
molecule harmine inhibits angiogenesis and suppresses tumour growth
through activation of p53 in endothelial cells. PLoS One.
7:e521622012. View Article : Google Scholar
|
|
9
|
Li C, Wang Y, Wang C, Yi X, Li M and He X:
Anticancer activities of harmine by inducing a pro-death autophagy
and apoptosis in human gastric cancer cells. Phytomedicine.
28:10–18. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Abe A and Yamada H: Harmol induces
apoptosis by caspase-8 activation independently of Fas/Fas ligand
interaction in human lung carcinoma H596 cells. Anticancer Drugs.
20:373–381. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hashemi Sheikh, Shabani S, Seyed Hasan,
Tehrani S, Rabiei Z, Tahmasebi Enferadi S and Vannozzi GP: Peganum
harmala L.'s anti-growth effect on a breast cancer cell line.
Biotechnol Rep (Amst). 8:138–143. 2015. View Article : Google Scholar
|
|
12
|
Zhang L, Zhang F, Zhang W, Chen L, Gao N,
Men Y, Xu X and Jiang Y: Harmine suppresses homologous
recombination repair and inhibits proliferation of hepatoma cells.
Cancer Biol Ther. 16:1585–1592. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hamsa TP and Kuttan G: Harmine activates
intrinsic and extrinsic pathways of apoptosis in B16F-10 melanoma.
Chin Med. 6:112011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Song Y, Kesuma D, Wang J, Deng Y, Duan J,
Wang JH and Qi RZ: Specific inhibition of cyclin-dependent kinases
and cell proliferation by harmine. Biochem Biophys Res Commun.
317:128–132. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zou N, Wei Y, Li F, Yang Y, Cheng X and
Wang C: The inhibitory effects of compound Muniziqi granule against
B16 cells and harmine induced autophagy and apoptosis by inhibiting
Akt/mTOR pathway. BMC Complement Altern Med. 17:5172017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hamsa TP and Kuttan G: Harmine inhibits
tumour specific neo-vessel formation by regulating VEGF, MMP, TIMP
and pro-inflammatory mediators both in vivo and in vitro. Eur J
Pharmacol. 649:64–73. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou X and Lei QY: Regulation of TAZ in
cancer. Protein Cell. 7:548–561. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Park KS, Whitsett JA, Di Palma T, Hong JH,
Yaffe MB and Zannini M: TAZ interacts with TTF-1 and regulates
expression of surfactant protein-C. J Biol Chem. 279:17384–17390.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jeong H, Bae S, An SY, Byun MR, Hwang JH,
Yaffe MB, Hong JH and Hwang ES: TAZ as a novel enhancer of
MyoD-mediated myogenic differentiation. FASEB J. 24:3310–3320.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Varelas X, Samavarchi-Tehrani P, Narimatsu
M, Weiss A, Cockburn K, Larsen BG, Rossant J and Wrana JL: The
Crumbs complex couples cell density sensing to Hippo-dependent
control of the TGF-β-SMAD pathway. Dev Cell. 19:831–844. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cui CB, Cooper LF, Yang X, Karsenty G and
Aukhil I: Transcriptional coactivation of bone-specific
transcription factor Cbfa1 by TAZ. Mol Cell Biol. 23:1004–1013.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mahoney WM Jr, Hong JH, Yaffe MB and
Farrance IK: The transcriptional co-activator TAZ interacts
differentially with transcriptional enhancer factor-1 (TEF-1)
family members. Biochem J. 388:217–225. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hong JH, Hwang ES, McManus MT, Amsterdam
A, Tian Y, Kalmukova R, Mueller E, Benjamin T, Spiegelman BM, Sharp
PA, et al: TAZ, a transcriptional modulator of mesenchymal stem
cell differentiation. Science. 309:1074–1078. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang H, Liu CY, Zha ZY, Zhao B, Yao J,
Zhao S, Xiong Y, Lei QY and Guan KL: TEAD transcription factors
mediate the function of TAZ in cell growth and
epithelial-mesenchymal transition. J Biol Chem. 284:13355–13362.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hong JH and Yaffe MB: TAZ: A
beta-catenin-like molecule that regulates mesenchymal stem cell
differentiation. Cell Cycle. 5:176–179. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou Z, Hao Y, Liu N, Raptis L, Tsao MS
and Yang X: TAZ is a novel oncogene in non-small cell lung cancer.
Oncogene. 30:2181–2186. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang L, Shi S, Guo Z, Zhang X, Han S, Yang
A, Wen W and Zhu Q: Overexpression of YAP and TAZ is an independent
predictor of prognosis in colorectal cancer and related to the
proliferation and metastasis of colon cancer cells. PLoS One.
8:e655392013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bhat KP, Salazar KL, Balasubramaniyan V,
Wani K, Heathcock L, Hollingsworth F, James JD, Gumin J, Diefes KL,
Kim SH, et al: The transcriptional coactivator TAZ regulates
mesenchymal differentiation in malignant glioma. Genes Dev.
25:2594–2609. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou X, Wang S, Wang Z, Feng X, Liu P, Lv
XB, Li F, Yu FX, Sun Y, Yuan H, et al: Estrogen regulates Hippo
signaling via GPER in breast cancer. J Clin Invest. 125:2123–2135.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cordenonsi M, Zanconato F, Azzolin L,
Forcato M, Rosato A, Frasson C, Inui M, Montagner M, Parenti AR,
Poletti A, et al: The Hippo transducer TAZ confers cancer stem
cell-related traits on breast cancer cells. Cell. 147:759–772.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang N, Morrison CD, Liu P, Miecznikowski
J, Bshara W, Han S, Zhu Q, Omilian AR, Li X and Zhang J: TAZ
induces growth factor-independent proliferation through activation
of EGFR ligand amphiregulin. Cell Cycle. 11:2922–2930. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xu W, Wei Y, Wu S, Wang Y, Wang Z, Sun Y,
Cheng SY and Wu J: Up-regulation of the Hippo pathway effector TAZ
renders lung adenocarcinoma cells harboring EGFR-T790M mutation
resistant to gefitinib. Cell Biosci. 5:72015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yuen HF, McCrudden CM, Huang YH, Tham JM,
Zhang X, Zeng Q, Zhang SD and Hong W: TAZ expression as a
prognostic indicator in colorectal cancer. PLoS One. 8:e542112013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chan SW, Lim CJ, Guo K, Ng CP, Lee I,
Hunziker W, Zeng Q and Hong W: A role for TAZ in migration,
invasion, and tumori-genesis of breast cancer cells. Cancer Res.
68:2592–2598. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
36
|
Chen H, Zhu G, Li Y, Padia RN, Dong Z, Pan
ZK, Liu K and Huang S: Extracellular signal-regulated kinase
signaling pathway regulates breast cancer cell migration by
maintaining slug expression. Cancer Res. 69:9228–9235. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pal I and Mandal M: PI3K and Akt as
molecular targets for cancer therapy: Current clinical outcomes.
Acta Pharmacol Sin. 33:1441–1458. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Geng X, Ren Y, Wang F, Tian D, Yao X,
Zhang Y and Tang J: Harmines inhibit cancer cell growth through
coordinated activation of apoptosis and inhibition of autophagy.
Biochem Biophys Res Commun. 498:99–104. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cao MR, Li Q, Liu ZL, Liu HH, Wang W, Liao
XL, Pan YL and Jiang JW: Harmine induces apoptosis in HepG2 cells
via mitochondrial signaling pathway. Hepatobiliary Pancreat Dis
Int. 10:599–604. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Uhl KL, Schultz CR, Geerts D and Bachmann
AS: Harmine, a dual-specificity tyrosine phosphorylation-regulated
kinase (DYRK) inhibitor induces caspase-mediated apoptosis in
neuroblastoma. Cancer Cell Int. 18:822018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sasaki H: Roles and regulations of Hippo
signaling during preimplantation mouse development. Dev Growth
Differ. 59:12–20. 2017. View Article : Google Scholar
|
|
42
|
He J, Bao Q, Yan M, Liang J, Zhu Y, Wang C
and Ai D: The role of Hippo/yes-associated protein signalling in
vascular remodelling associated with cardiovascular disease. Br J
Pharmacol. 175:1354–1361. 2018. View Article : Google Scholar
|
|
43
|
Patel SH, Camargo FD and Yimlamai D: Hippo
Signaling in the Liver Regulates Organ Size, Cell Fate, and
Carcinogenesis. Gastroenterology. 152:533–545. 2017. View Article : Google Scholar :
|
|
44
|
Gregorieff A and Wrana JL: Hippo
signalling in intestinal regeneration and cancer. Curr Opin Cell
Biol. 48:17–25. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Watt KI, Harvey KF and Gregorevic P:
Regulation of Tissue Growth by the Mammalian Hippo Signaling
Pathway. Front Physiol. 8:9422017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kim W and Jho EH: The history and
regulatory mechanism of the Hippo pathway. BMB Rep. 51:106–118.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yuan M, Tomlinson V, Lara R, Holliday D,
Chelala C, Harada T, Gangeswaran R, Manson-Bishop C, Smith P,
Danovi SA, et al: Yes-associated protein (YAP) functions as a tumor
suppressor in breast. Cell Death Differ. 15:1752–1759. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yu X, Long YC and Shen HM: Differential
regulatory functions of three classes of phosphatidylinositol and
phosphoinositide 3-kinases in autophagy. Autophagy. 11:1711–1728.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tang D, Chen QB, Xin XL and Aisa HA:
Anti-diabetic effect of three new norditerpenoid alkaloids in vitro
and potential mechanism via PI3K/Akt signaling pathway. Biomed
Pharmacother. 87:145–152. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lynch JT, McEwen R, Crafter C, McDermott
U, Garnett MJ, Barry ST and Davies BR: Identification of
differential PI3K pathway target dependencies in T-cell acute
lymphoblastic leukemia through a large cancer cell panel screen.
Oncotarget. 7:22128–22139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li YH, Fu HL, Tian ML, Wang YQ, Chen W,
Cai LL, Zhou XH and Yuan HB: Neuron-derived FGF10 ameliorates
cerebral ischemia injury via inh+ibiting NF-κB-dependent
neuroinflammation and activating PI3K/Akt survival signaling
pathway in mice. Sci Rep. 6:198692016. View Article : Google Scholar
|
|
52
|
Fujimori Y, Inokuchi M, Takagi Y, Kato K,
Kojima K and Sugihara K: Prognostic value of RKIP and p-ERK in
gastric cancer. J Exp Clin Cancer Res. 31:302012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Roos DH, Puntel RL, Lugokenski TH, Ineu
RP, Bohrer D, Burger ME, Franco JL, Farina M, Aschner M, Rocha JB,
et al: Complex methylmercury-cysteine alters mercury accumulation
in different tissues of mice. Basic Clin Pharmacol Toxicol.
107:789–792. 2010. View Article : Google Scholar : PubMed/NCBI
|