Introduction
Colorectal cancer (CRC) is the fourth most commonly
diagnosed cancer type worldwide, with estimated 1.1 million new
cases and 551,000 deaths in 2018 (1). Despite the observation that the
incidence of CRC has been gradually decreased and integrated
therapy has greatly improved the prognosis of patients with CRC in
the recent years, the overall survival of patients with CRC remains
poor, and the five-year survival is ~60% (2,3).
Therefore, there is an urgent need to develop innovative detection
and therapeutic approaches for patients with CRC.
β-catenin-dependent Wnt signaling, also known as
canonical Wnt signaling, has a pivotal role in mediating cancer
cell proliferation, metastasis and stemness maintenance (4-6).
Upon activation, such as following Wnt ligand binding, β-catenin,
the major effector of Wnt/β-catenin signaling, translocates into
the nucleus of cells to form a complex with co-regulators of
transcription factors, thus promoting the transcription of multiple
oncogenes (7). Overactivation of
Wnt/β-catenin signaling has been observed in several types of
cancer, including CRC (8).
Activation of Wnt/β-catenin signaling is essential for the
initiation of colorectal cancer, as well as promotes
chemoradiotherapy resistance (9,10).
The activity of Wnt/β-catenin signaling is tightly controlled by
several positive and negative regulators (11). Adenomatous polyposis coli (APC) is
one of the well-studied negative regulators of Wnt/β-catenin
signaling (12). A APC truncating
mutation has been observed in a large proportion of patients with
CRC (13), suggesting that
Wnt/β-catenin signaling may be a promising target for the treatment
of patients with CRC. Altered expression of non-coding RNA and
transcription factors also contributes to the downregulation of APC
in cancer cells (14,15).
microRNAs (miRNAs) are small, non-coding RNA
molecules which were first discovered in Caenorhabditis
elegans (16).
Mechanistically, miRNAs function via base-pairing with sequences on
the 3′-untranslated region (3′-UTR) of target gene mRNAs (17). Dysregulation of miRNAs leads to
aberrant expression of target genes, resulting in the disruption of
signaling networks in cells. Accumulating evidence suggests that
miRNAs are involved in almost all physiological and pathological
conditions (18,19). In CRC, multiple miRNAs have been
identified as oncogenes or tumor suppressors via regulating key
genes in oncogenic signaling pathways (20). For example, miR-144 was reported as
a regulator of cell proliferation and rapamycin sensitivity in CRC
through directly targeting mTOR (21). miRNA microarray analysis of normal,
adenoma and carcinoma tissues discovered several dysregulated
miRNAs (22), however, their roles
have not been studied yet. Through bioinformatic analysis,
miR-501-3p was discovered as a differentially expressed miRNA in
CRC. miR-501-3p has been previously confirmed as an oncogenic miRNA
in cervical cancer and hepatocellular carcinoma (23,24).
However, the role of miR-501-3p in CRC remains unknown.
The current study focused on the role of this
differentially expressed miRNA, miR-501-3p, in CRC cells. Reverse
transcription-quantitative PCR (RT-qPCR) suggested that miR-501-3p
was overexpressed in tumor tissues of patients with CRC. In
addition, downregulation of miR-501-3p inhibited cell
proliferation, cell cycle progression and sphere formation in CRC
cells. Furthermore, miR-501-3p regulated CRC progression via
targeting APC to activate Wnt/β-catenin signaling.
Materials and methods
Patient samples
A total of 30 patients with CRC who underwent
surgical resection at the Fourth People's Hospital of Shanxi from
September 2015 to January 2018 were included in the present study.
None of the patients received chemotherapy or radiotherapy prior to
surgery. Written consents were provided by all participants and all
patients agreed to the use of their tissue samples in the current
study. The experiments were performed under the supervision of the
Ethics Committee of the Fourth People's Hospital of Shanxi. There
were 19 males and 11 females, aged 35-68 years. Exclusion criteria
were as follows: Patients with a history of other malignancies or
other serious active diseases recently; and patients with drug
hepatitis, alcoholic liver disease or autoimmune liver disease. The
normal and tumor tissues were immediately stored at −80°C, until
use for the following experiments.
Cell culture
CRC cell lines HCT116, Caco2, LOVO, HT29 and the
fetal colon epithelial cell line FHC were purchased from the Cell
Center in Shanghai Institutes for Biological Sciences, Chinese
Academy of Sciences (Shanghai, China). All cells were cultured in
DMEM (Invitrogen; Thermo Fisher Scientific, Inc.) supplemented with
10% FBS (Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin (Sigma-Aldrich; Merck KGaA) in a 95% humid
incubator with 5% CO2.
RNA extraction and RT-qPCR
Total RNA was extracted from tissues and cells using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
following the manufacturer's protocol. The concentration of RNA was
determined using a NanoDrop 2000 (Thermo Fisher Scientific, Inc.).
RNA was reverse transcribed into cDNA using SMART MMLV Reverse
Transcriptase (Takara Bio, Inc.). qPCR was performed on a CFX96
Real-Time PCR Detection System (Bio-Rad Laboratories, Inc.) with TB
Green Premix Ex Taq (Takara Bio, Inc.). The qPCR conditions were as
follows: Predenature at 98°C for 30 sec, followed by 35 cycles of
denature at 98°C for 5 sec and elongation and annealing at 60°C for
30 sec. GAPDH and U6 were used as internal controls for mRNA and
miRNA, respectively. Relative gene expression was calculated using
the 2−ΔΔCq method (25). The primer sequences were as
follows: APC, forward 5′-AAG CAT GAA ACC GGC TCA CAT-3′ and reverse
5′-CAT TCG TGT AGT TGA ACC CTG A-3′; β-catenin, forward 5′-ATG GAC
AGT ATG CAA TGA CTC G-3′ and reverse 5′-TAG CAG ACA CCA TCT GAG GAG
A-3′; cyclin D1, forward 5′-GCT GCG AAG TGG AAA CCA TC-3′ and
reverse 5′-CCT CCT TCT GCA CAC ATT TGA A-3′; c-Myc, forward 5′-GGC
TCC TGG CAA AAG GTC A-3′ and reverse 5′-CTG CGT AGT TGT GCT GAT
GT-3′; GAPDH, forward 5′-GGA GCG AGA TCC CTC CAA AAT-3′ and reverse
5′-GGC TGT TGT CAT ACT TCT CAT GG-3′; stem-loop primer, 5′-CTC AAC
TGG TGT CGT GGA GTC GGC AAT TCA GTT GAG AGA ATC-3′; miR-501-3p,
forward 5′-GCC GAG AAT GCA CCC GGG CA-3′ and reverse 5′-CTC AAC TGG
TGT CGT GGA-3′; U6, forward 5′-CTC GCT TCG GCA GCA C-3′ and reverse
5′-AAC GCT TCA CGA ATT TGC G-3′.
Protein extraction and western
blotting
Primary antibodies against APC (cat. no. 2504;
1:1,000), β-catenin (cat. no. 8480; 1:1,000), phosphorylated (p-)
β-catenin (cat. no. 9561; 1:1,000), cyclin D1 (cat. no. 2978;
1:1,000) and c-Myc (cat. no. 9402; 1:1,000) were purchased from
Cell Signaling Technology, Inc. The antibodies targeting histone H1
(H1; cat. no. ab71594; 1:10,000) and GAPDH (cat. no. ab8245;
1:5,000) were purchased from Abcam. Horseradish
peroxidase-conjugated mouse (cat. no. SA00001-1; 1:10,000) and
rabbit (cat. no. SA00001-2; 1:10,000) secondary antibodies were
products of ProteinTech Group, Inc. Protein lysates were prepared
with RIPA lysis buffer (Thermo Fisher Scientific, Inc.) following
the manufacturer's protocol. The concentration of lysates was
determined with BCA Protein Assay kit (Thermo Fisher Scientific,
Inc.). A total of 20 µg lysate sample was loaded in each
well of 8% SDS gels and separated by electrophoresis. The proteins
were transferred to PVDF membranes and incubated with 5% non-fat
milk for 30 min at room temperature. Afterwards, the membrane was
incubated with primary antibody and secondary antibody for 1 h at
room temperature. The blots were developed with enhanced
chemiluminescence ECL Western Blotting Substrate (Thermo Fisher
Scientific, Inc.). The relative protein expression was analyzed
with Image J software version 1.51J8 (National Institutes of
Health).
Transfection of miR-501-3p mimic and
inhibitor
miR-501-3p mimic (5′-AAU CCU UUG UCC CUG GGU GAG
A-3′), miR-negative control (NC) mimic (5′-UCA CAA CCU CCU AGA AAG
AGU AGA-3′), miR-501-3p inhibitor (5′-UCU CAC CCA GGG ACA AAG GAU
U-3′) and miR-NC inhibitor (5′-CAG UAC UUU UGU GUA GUA CAA-3′) were
synthesized and purchased from Shanghai GenePharma Co., Ltd. For
transfection, 50 nM miR-501-3p mimic or miR-NC mimic or miR-501-3p
inhibitor or miR-NC inhibitor was transfected into the indicated
cells using Lipofectamine 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) following the manufacturer's protocol. At 48 h
post-transfection, the cultured cells were harvested for subsequent
experiments.
Cell proliferation assay
To determine the cell proliferation ability, a Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.) was
used. Briefly, the cells were seeded in 6-well plates and
transfected with miR-501-3p inhibitor or miR-NC inhibitor or miR-NC
inhibitor + APC small interfering (si)RNA. On the next day, the
cells were harvested and re-seeded in 96-well plates at a
concentration of 10,000 cells per well. Thereafter, 10 µl
CCK8 solution was added for 2 h into each well at 0, 24, 48, 72 h
post-cell adhesion. The medium was then transferred into new
96-well plates and the absorbance of each well was determined using
a microplate reader.
Cell cycle assay
At 48 h post-transfection, the cells were fixed in
75% ethanol at −20°C overnight. On the next day, the cells were
washed in cold PBS and stained with propidium iodide (Invitrogen;
Thermo Fisher Scientific, Inc.) for 30 min at room temperature.
Finally, the cells were analyzed using a FACSCalibur flow cytometer
(BD Biosciences) and FlowJo software version 10.4 (FlowJo LLC).
Sphere formation assay
Briefly, after transfection, the cells were
harvested by trypsinization, and the cell suspensions were washed
with PBS three times. The cells were resuspended with sphere
condition media (DMEM/F12; Thermo Fisher Scientific, Inc.)
supplemented with B27 (Thermo Fisher Scientific, Inc.) and
recombinant epidermal growth factor (20 ng/ml; PeproTech, Inc.) at
a final concentration of 1,000 cells/ml. These cell suspensions (1
ml) were seeded in a 96-well ultra-low attachment plate (Corning
Inc.). At day 7 of cell culture, the number of spheres (diameter
>50 µm) was counted in three random fields using an
inverted light microscope (magnification, ×20).
Immunofluorescence
HCT116 cells were plated on a slide glass in 96-well
plates. Following transfection with miR-501-3p inhibitor or miR-NC
inhibitor in combination with control siRNA or APC siRNA, HCT116
cells were fixed with 4% formaldehyde for 1 h at room temperature,
washed with PBS containing 0.5% triton, and blocked with 5% bovine
serum (Sigma Aldrich; Merck KGaA) for 1 h at room temperature. The
cells were then incubated with β-catenin antibody (cat. no. 8480;
1:100; Cell Signaling Technology, Inc.) at 4°C overnight. On the
next day, the cells were washed with PBS and incubated with Alexa
Fluor 488-conjugated secondary antibody (cat. no. A-11034;
1:100,000; Thermo Fisher Scientific, Inc.) at room temperature for
2 h. After washing with PBS, Vectashield antifade mounting medium
with DAPI (Vector Laboratories, Inc.) was used. Cells were observed
and images were captured with a fluorescence microscope
(magnification, ×40; Leica Microsystems GmbH).
Cycloheximide (CHX) chase assay
To detect the stability of proteins, a CHX chase
assay was performed. Following transfection with miR-501-3p
inhibitor or miR-NC inhibitor for 48 h, the cells were treated with
CHX (10 µg/µl) for 0, 2 or 4 h, respectively. The
cells were then harvested for western blotting, as
aforementioned.
Extraction of nuclear and cytoplasmic
protein
The NE-PER™ Nuclear and Cytoplasmic Extraction
Reagents kit (Thermo Fisher Scientific, Inc.) was used to extract
nuclear and cytoplasmic proteins from cells, following the
manufacturer's protocol. H1 and GAPDH were used as controls for the
nuclear and cytoplasmic extracts, respectively.
Dual luciferase reporter assay
For construction of the pGL3-APC 3'UTR-WT reporter
plasmid, the 3'UTR sequence of the APC gene containing the putative
binding sites for miR-501-3p was amplified from cDNA generated from
HCT116 cells and ligated into the pGL3 plasmid (Promega
Corporation) with T4 DNA ligase (New England Biolabs, Inc.).
Briefly, HCT116 cells were seeded in 24-well plates. On the next
day, miR-501-3p mimic or miR-NC mimic was co-transfected with
pGL3-APC 3'UTR-WT or pGL3-APC 3'UTR-Mut into HCT116 cells using
Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific, Inc.).
After 24 h, the relative luciferase activity of HCT116 cells was
determined with a Dual Luciferase Reporter System Assay kit
(Promega Corporation) via normalization of the firefly luciferase
activity to Renilla luciferase activity.
Silencing of APC
APC siRNA (5′-GGA TCA GCC TAT TGA TTA T-3′) and
control siRNA (5′-TTC TCC GAA CGT GTC ACG T-3′) were synthesized
and purchased from Shanghai GenePharma Co., Ltd. For silencing of
APC, the cells were transfected with 50 nM APC siRNA using
Lipofectamine RNAiMax (Invitrogen; Thermo Fisher Scientific, Inc.).
At 48 h post-transfection, the cells were subjected to subsequent
experiments.
Statistical analysis
All experiments were repeated at least three times.
Data were analyzed with GraphPad Prism 5.0 (GraphPad Software,
Inc.) and presented as mean ± SD. The correlation between
miR-501-3p and APC expression levels was analyzed by Pearson's
correlation analysis. The patients were divided into high and low
expression groups according to the median expression value of
miR-501-3p, and Chi-square test was used to compare the
pathological data from the collected samples. Two groups were
compared with Student's t-test. Three groups were analyzed with
one-way ANOVA followed by Newman-Keuls test. P<0.05 was
considered to indicate a statistically significant difference.
Results
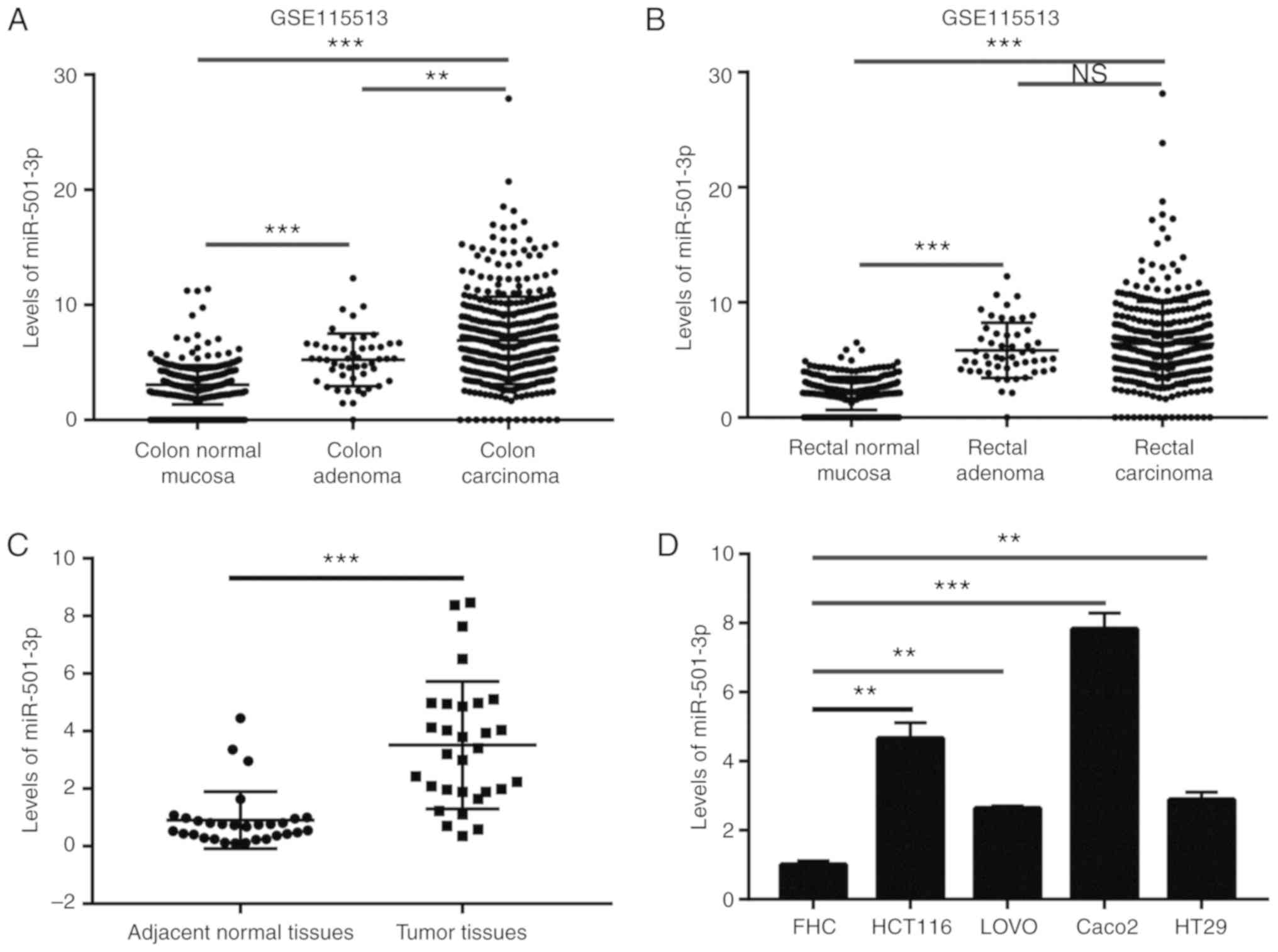
miR-501-3p is upregulated in CRC tissues
and cells
To investigate the potential role of miR-501-3p in
CRC, the expression levels of miR-501-3p in colon normal mucosa,
colon adenoma, colon carcinoma, rectal normal mucosa, rectal
adenoma and rectal carcinoma were analyzed from a GEO dataset
(GSE115513) (22). The results
indicated that miR-501-3p levels were elevated in adenoma and
carcinoma compared with normal tissues (Fig. 1A and B). To confirm this
observation, RT-qPCR was used to detect the miR-501-3p expression
levels in 30 pairs of normal and tumor tissues from patients with
CRC collected for the present study. The results demonstrated that
miR-501-3p was significantly overexpressed in tumor tissues
compared to normal tissues (Fig.
1C). Additionally, high expression of miR-501-3p was associated
with increased TNM stage (III/IV) in patients with CRC (Table I). In vitro, miR-501-3p was
significantly overexpressed in CRC cell lines (HCT116, LOVO, Caco2
and HT29) compared with a colon epithelial cell line FHC (Fig. 1D). These data suggested that
miR-501-3p might be involved in the progression of CRC.
 | Table IAssociation between miR-501-3p
expression and clinicopathological features of patients with
colorectal cancer. |
Table I
Association between miR-501-3p
expression and clinicopathological features of patients with
colorectal cancer.
| Clinicopathological
features | Number of
cases | miR-501 -3p
expression
| P-value |
|---|
| High (n=15) | Low (n=15) |
|---|
| Age | | | | 1.000 |
| >50 | 17 | 9 | 8 | |
| ≤50 | 13 | 6 | 7 | |
| Sex | | | | 0.427 |
| Male | 21 | 9 | 12 | |
| Female | 9 | 6 | 3 | |
| Tumor size
(mm) | | | | 0.272 |
| >5 | 14 | 9 | 5 | |
| ≤5 | 16 | 6 | 10 | |
| TNM stage | | | | 0.008 |
| I-II | 12 | 2 | 10 | |
| III-IV | 18 | 13 | 5 | |
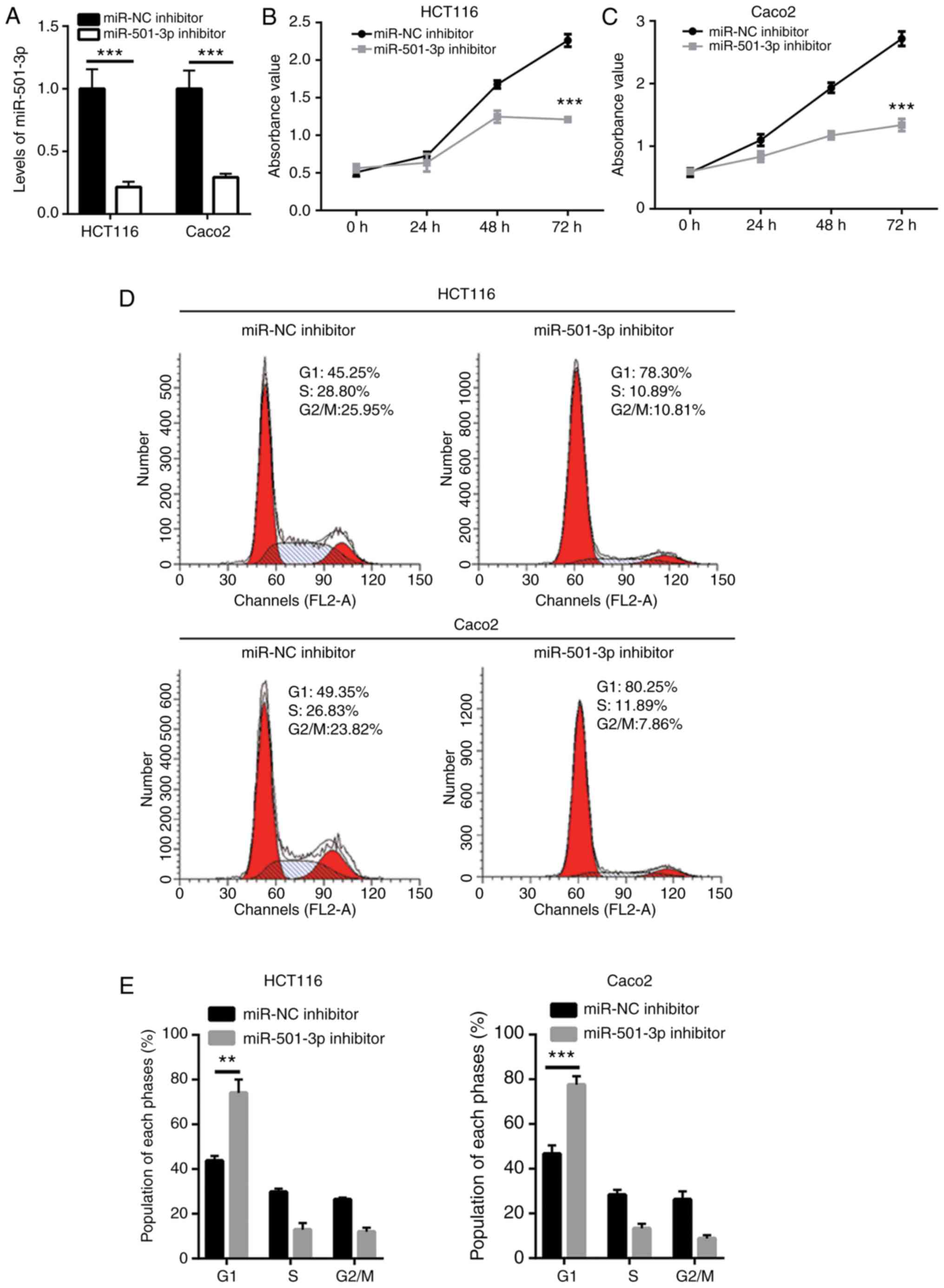
Downregulation of miR-501-3p inhibits CRC
cell proliferation and stemness
To explore the function of miR-501-3p in CRC, CRC
cells were transfected in vitro with a miR-501-3p inhibitor.
Transfection of mR-501-3p inhibitor effectively decreased the
miR-501-3p expression levels in HCT116 and Caco2 cells (Fig. 2A). The cell proliferation assay
indicated that miR-501-3p inhibitor significantly suppressed cell
proliferation in both HCT116 and Caco2 cells (Fig. 2B and C). Uncontrolled cell cycle
progression leads to fast cell proliferation in cancer cells.
Results from flow cytometry analysis revealed that downregulation
of miR-501-3p induced an accumulation of cells in the G1 phase in
HCT116 and Caco2 cells (Fig. 2D and
E), suggesting that miR-501-3p might regulate cell
proliferation via cell cycle control. Cancer stem cells are cancer
cells that exhibit strong self-renewing ability and are associated
with cancer initiation and drug resistance. The sphere formation
assay revealed that downregulation of miR-501-3p significantly
reduced the number of spheres formed by HCT116 and Caco2 cells
(Fig. 2F and G), indicating that
miR-501-3p could maintain the stemness of CRC cells.
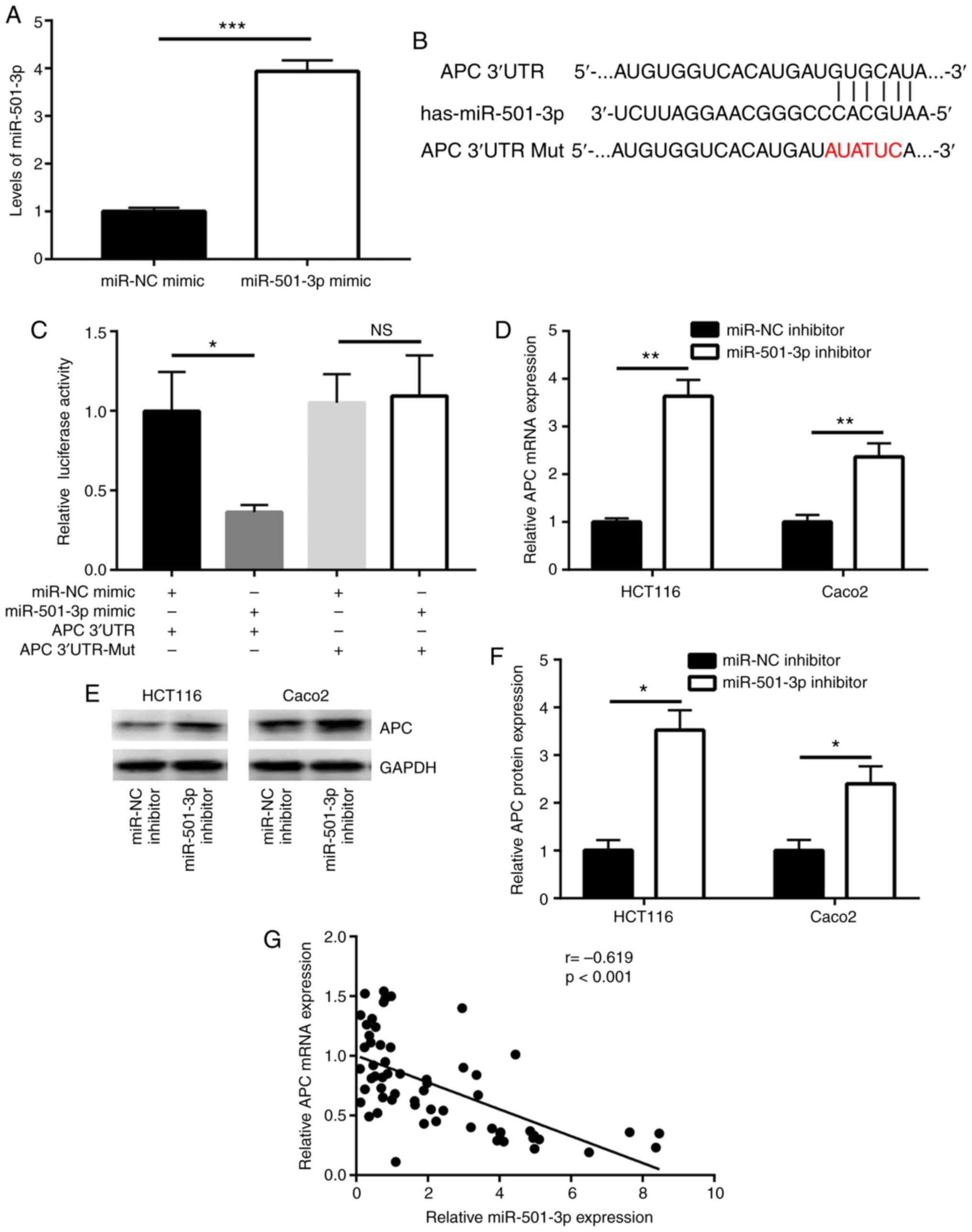
APC is a direct target gene of miR-501-3p
in CRC cells
miRNAs function via directly binding to the 3'UTR of
target mRNAs to downregulate their expression. Using TargetScan
software (26), several mRNAs were
predicted to be potential target genes of miR-501-3p. Among them,
the 3'UTR of APC, a well-known negative regulator of Wnt/β-catenin
signaling, was found to harbor a complementary binding site for
miR-501-3p (Fig. 3B). To further
investigate whether APC is a direct target of miR-501-3p, mimics
transfection and a dual luciferase assay were employed. First, the
efficacy of miR-501-3p overexpression by mimic transfection was
confirmed (Fig. 3A). In the dual
luciferase reporter assay, overexpression of miR-501-3p
significantly reduced the relative luciferase activity in HCT116
cells transfected with APC 3'UTR, but not the mutant APC 3'UTR
(Fig. 3C). In addition, inhibition
of miR-501-3p significantly increased the APC mRNA expression
levels in HCT116 and Caco2 cells (Fig.
3D). Western blot analysis revealed that miR-501-3p inhibition
also significantly increased the APC protein expression levels in
these two cell lines (Fig. 3E and
F). Of note, using RT-qPCR to examine the mRNA expression
levels of APC, a strong negative correlation was observed between
miR-501-3p and APC mRNA expression in tumor and normal tissues from
the 30 patients with CRC (Fig.
3G).
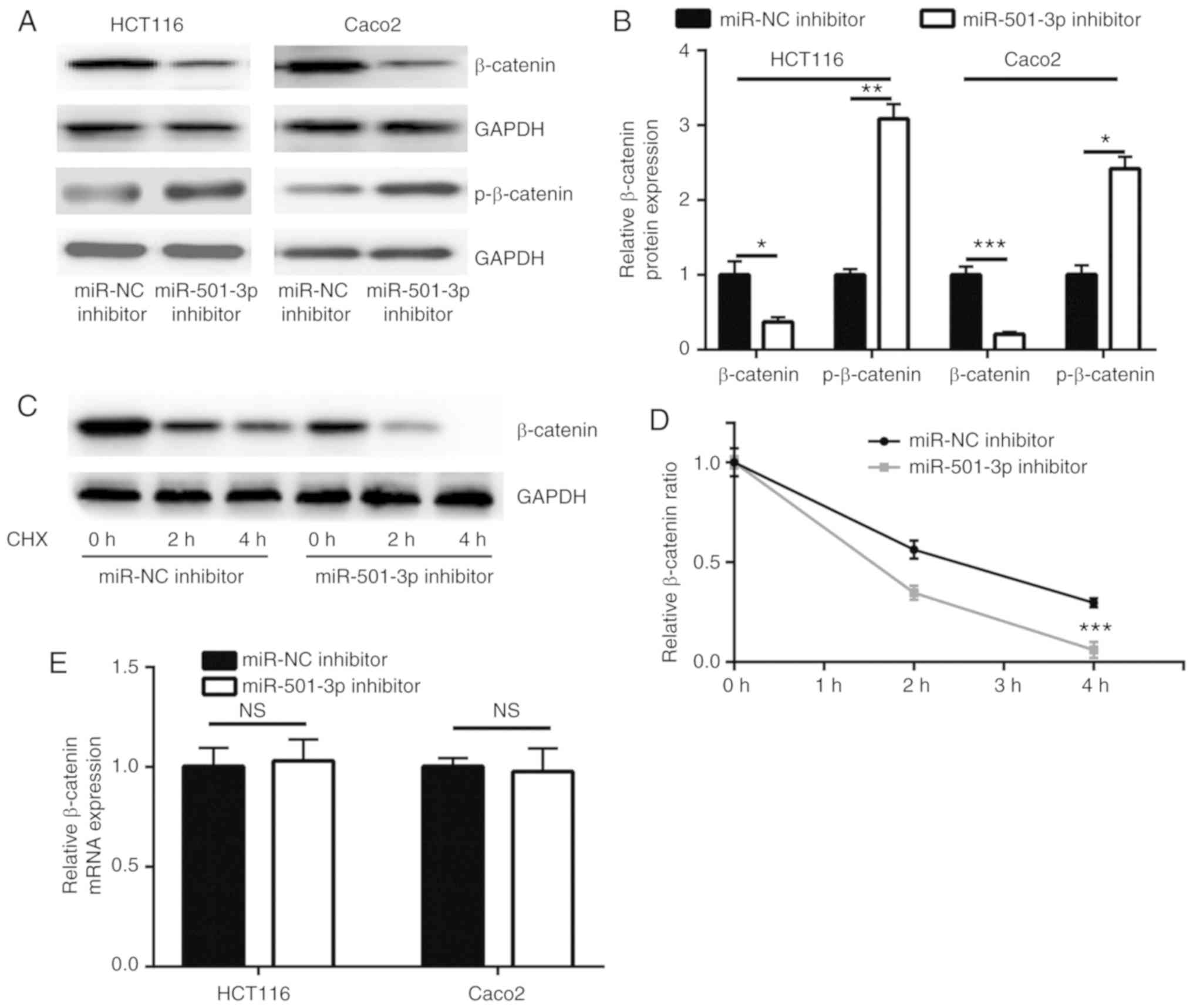
miR-501-3p inhibition downregulates
β-catenin expression via targeting APC in CRC cells
Consistent with the upregulation of APC, miR-501-3p
inhibition significantly decreased β-catenin protein expression
levels and increased p-β-catenin levels, in both HCT116 and Caco2
cells (Fig. 4A and B). APC
promotes the degradation of β-catenin protein via forming a complex
with glycogen synthase kinase 3β and other proteins to
phosphorylate β-catenin (28). In
a CHX chase assay, inhibition of miR-501-3p significantly
destabilized the β-catenin protein in HCT116 cells (Fig. 4C and D). RT-qPCR results indicated
that the mRNA expression levels of β-catenin were not altered
following miR-501-3p inhibition (Fig.
4E), suggesting that miR-501-3p regulated the stability of the
β-catenin protein rather than its gene transcription. The
subcellular localization of β-catenin is tightly controlled by APC
and associated with its transcriptional activity (28). To investigate whether APC was
involved in the regulation of β-catenin by miR-501-3p, APC siRNA
was transfected into HCT116 cells to downregulate APC expression
(Fig. 5A and B). While the
miR-501-3p inhibitor increased the APC protein expression levels,
APC silencing by siRNA decreased the APC protein expression levels
in HCT116 cells (Fig. 5C and D).
To explore whether miR-501-3p regulated β-catenin cellular
localization, cytoplasmic and nuclear protein extracts were
separately prepared. Western blotting demonstrated that
transfection with the miR-501-3p inhibitor significantly increased
the proportion of β-catenin in the cytoplasm and decreased the
proportion of β-catenin in the nucleus, compared with the cells
transfected with the negative control (Fig. 5E and F). By contrast, APC silencing
reversed the translocation of β-catenin in HCT116 cells (Fig. 5E and F), suggesting the
inactivation of Wnt/β-catenin signaling. The translocation of
β-catenin from the nucleus to the cytoplasm following miR-501-3p
inhibition was further confirmed by immunofluorescence (Fig. 5G); APC silencing reversed this
effect (Fig. 5G). These results
indicated that miR-501-3p regulated Wnt/β-catenin signaling via
targeting APC in CRC cells.
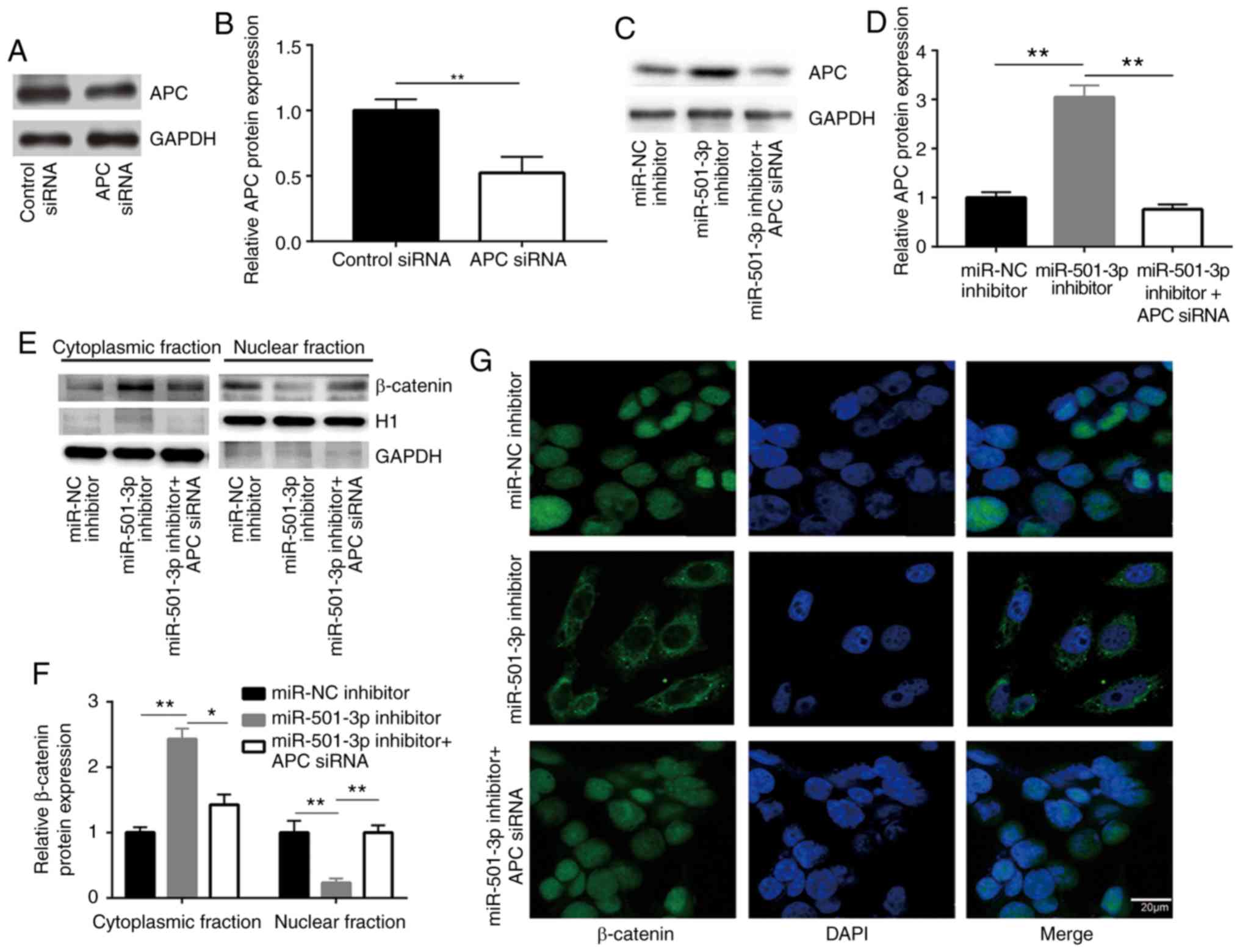 | Figure 5Downregulation of miR-501-3p promotes
translocation of β-catenin via targeting APC in colorectal cancer
cells. (A) Western blotting confirmed that transfection of APC
siRNA significantly decreased APC protein expression levels in
HCT116 cells. (B) Quantitative analysis of APC protein expression
in A. (C) Western blotting confirmed that miR-501-3p inhibitor
increased APC protein expression, and this was effectively reversed
following APC siRNA transfection in HCT116. (D) Quantitative
analysis of APC protein expression levels in C. (E) Western
blotting revealed that miR-501-3p downregulation resulted in an
increase of β-catenin protein in the cytoplasmic fraction and a
decrease of β-catenin in the nuclear fraction, while silencing of
APC reversed the altered subcellular localization of β-catenin in
HCT116 cells. H1 and GAPDH were used as internal controls for the
nuclear and the cytoplasmic protein extracts, respectively. (F)
Quantitative analysis of β-catenin protein expression in E. (G)
Immunofluorescence analysis revealed that miR-501-3p downregulation
induced translocation of β-catenin from the nucleus to the
cytoplasm, and silencing of APC reversed this translocation in
HCT116 cells. Scale bar, 20 µm. *P<0.05 and
**P<0.01, with comparisons indicated by lines. miR,
microRNA; APC, adenomatous polyposis coli; si, small interfering;
H1, histone H1. |
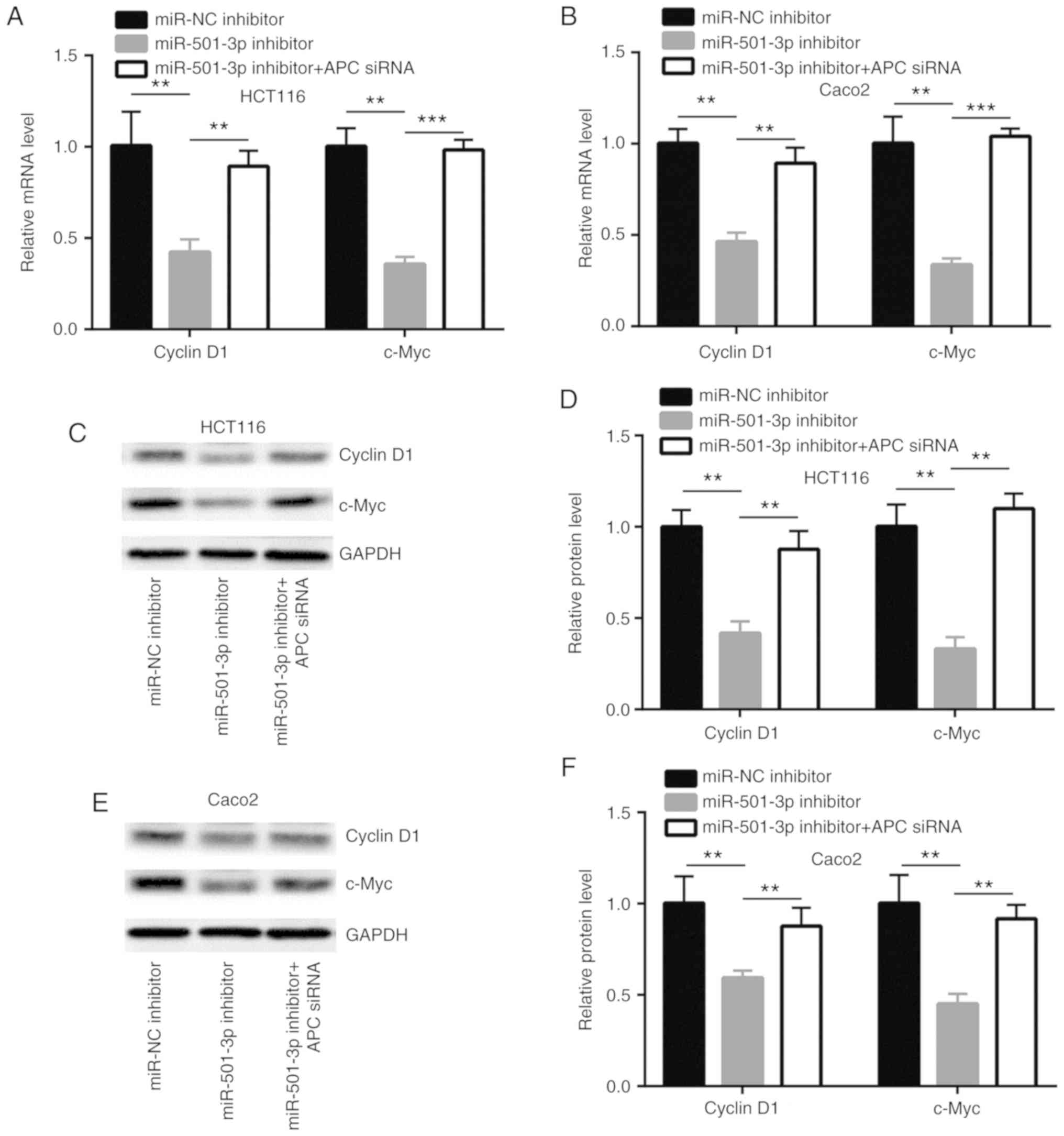
miR-501-3p regulates Wnt/β-catenin
signaling-related gene expression via targeting APC in CRC
cells
Wnt/β-catenin signaling directly activates several
key genes which are involved in cell proliferation, cell cycle and
stemness to control cancer progression (29). In HCT116 and Caco2 cells,
transfection with miR-501-3p inhibitor significantly decreased the
mRNA expression levels of cyclinD1 and c-Myc, two Wnt/β-catenin
pathway target genes; this effect was effectively reversed by APC
silencing (Fig. 6A and B). In
addition, the protein expression levels of cyclinD1 and c-Myc were
also reduced with miR-501-3p inhibition, which was again reversed
following APC silencing (Fig.
6C-F).
APC is pivotal for miR-501-3p-mediated
CRC cell proliferation and stemness
Next, the present study aimed to confirm whether APC
was critical for the function of miR-501-3p in CRC cells. Analysis
of cell cycle phase distribution revealed that an accumulation of
cells in the G1 phase induced by the miR-501-3p inhibitor was also
rescued after APC silencing (Fig. 7A
and B). In addition, transfection with the miR-501-3p inhibitor
induced a significant decrease of sphere formation ability, which
was recovered following APC silencing (Fig. 7C and D). Furthermore, the cell
proliferation assay suggested that APC silencing reversed the
miR-501-3p inhibitor-induced cell growth arrest in HCT116 cells
(Fig. 7E). The present results
demonstrated that miR-501-3p regulated CRC cell proliferation and
stemness via regulation of APC expression.
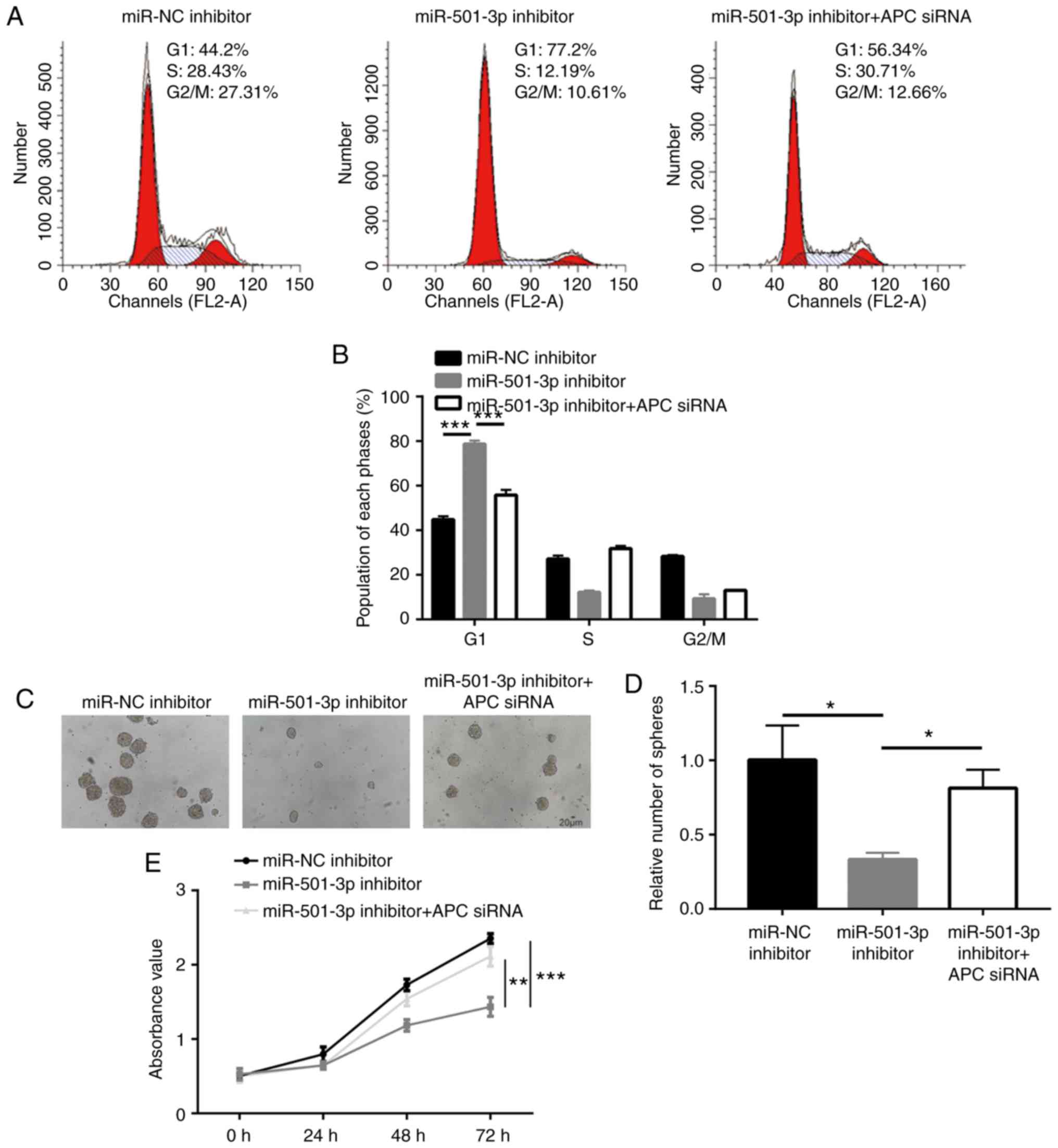 | Figure 7miR-501-3p/APC regulate colorectal
cancer cell proliferation, cell cycle progression and sphere
formation. (A) Representative images of cell cycle distribution in
HCT116 cells transfected with miR-NC inhibitor or miR-501-3p
inhibitor or miR-501-3p inhibitor + APC siRNA. (B) Quantitative
analysis of cell cycle distribution indicated that miR-501-3p
downregulation induced cell accumulation in the G1 phase, and this
effect was reversed after APC silencing. (C) Representative images
of sphere formation in HCT116 cells transfected with miR-NC
inhibitor or miR-501-3p inhibitor or miR-501-3p inhibitor + APC
siRNA. Scale bar, 20 µm. (D) Quantitative analysis of the
sphere formation assay. (E) Downregulation of miR-501-3p induced
cell growth arrest, and this was reversed after APC silencing.
*P<0.05, **P<0.01 and
***P<0.001, with comparisons indicated by lines. miR,
microRNA; APC, adenomatous polyposis coli; NC, negative control;
si, small interfering. |
Discussion
Accumulating evidence has suggested that miRNAs are
involved in the initiation and development of cancer (30). Several miRNAs have been discovered
as differentially expressed between tumor and normal tissues, or
serum from healthy volunteers and from patients with cancer
(31-33). Several have been experimentally
validated as oncogenes or tumor suppressors via targeting key genes
that are involved in cancer progression (34,35).
Compared with normal colonic mucosa, miR-501-3p is one of the most
significantly upregulated miRNAs in colorectal carcinomas, as
reported by Slattery et al (22). The current study further confirmed
the upregulation of miR-501-3p in CRC tumors and cell lines
compared with normal. Notably, the present results demonstrated
that miR-501-3p promoted CRC cell proliferation and stemness via
targeting APC to activate Wnt/β-catenin signaling.
miR-501-3p is involved in carcinogenesis and
Alzheimer's disease (23,36). In cervical cancer and
hepatocellular carcinoma, miR-501-3p is overexpressed in tumor
tissues compared with normal tissues, and promotes cell
proliferation, migration and invasion via targeting CYLD lysine 63
deubiquitinase (23,24). The present study demonstrated that
downregulation of miR-501-3p inhibited CRC cell proliferation, cell
cycle progression and sphere formation in CRC cells. The results of
functional assays suggested that miR-501-3p might accelerate cell
cycle progression to promote cell proliferation; in addition, they
emphasized a role of miR-501-3p in mediating cancer cell stemness
for the first time. Cancer stem cells are pivotal for tumor
initiation and drug resistance (37,38);
therefore, it would be interesting to examine the potential role of
miR-501-3p in regulating drug sensitivity in the future. Overall,
in agreement with previous studies, the present data support an
oncogenic role of miR-510-3p in CRC.
Among several oncogenic signaling pathways, the
Wnt/β-catenin signaling pathway is the most well-characterized
driver of CRC (13). Several
negative regulators of the Wnt/β-catenin signaling pathway, such as
APC and axin 1/2, are frequently mutated, resulting in loss of
function in CRC (39). In
addition, almost all the key components of Wnt/β-catenin signaling
are reported to be regulated by miRNAs in CRC. For example, miR-7
targets the transcription factor YY1 to inactivate Wnt/β-catenin
signaling and to inhibit CRC cell proliferation (40). miR-23b inhibits CRC cell
proliferation via targeting Frizzled class receptor 7, a receptor
of Wnt/β-catenin signaling (41).
By contrast, miR-135a/b targets APC to promote CRC progression
(42). Several miRNAs, such as
miR-3607, have been shown to target APC in other types of cancer
(43). In the present study, it
was predicted and confirmed that APC was a target gene of
miR-501-3p in CRC cells. Inhibition of miR-501-3p resulted in
downregulation of β-catenin protein expression, as well as its
translocation from the nucleus to the cytoplasm, inactivating
Wnt/β-catenin signaling in CRC cells. As target genes of
Wnt/β-catenin signaling, CyclinD1 and c-Myc are major regulators of
the G0/G1 cell cycle checkpoint and stemness (44,45).
The present data showing that cyclinD1 and c-Myc were downregulated
following miR-501-3p inhibition, suggested that miR-501-3p might
regulate cell proliferation, cell cycle and sphere formation via
APC. In addition, it was demonstrated that the inhibition of cell
proliferation, cell cycle and sphere formation induced by
miR-501-3p downregulation were reversed by APC silencing. The
current data revealed miR-501-3p as a novel regulator of
Wnt/β-catenin signaling in CRC.
In conclusion, the present results demonstrated that
miR-501-3p promoted the proliferation and stemness of CRC cells
through targeting APC and activating Wnt/β-catenin signaling. These
findings enrich the current understanding on the molecular
mechanism of miR-501-3p function and the activity of Wnt/β-catenin
signaling in CRC, which may facilitate the research and development
of novel treatment approaches. However, the association between
miR-501-3p and the prognosis of patients with CRC was not included
in the current study; further studies will be needed to assess the
prognostic potential of miR-501-3p.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
FL participated in the design and performance of the
experiments. XG and TX contributed to the collection of samples and
clinical data. TX supervised the study and wrote the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All procedures performed in the present study
involving human specimens were approved by the Ethics Committee of
The Fourth People's Hospital of Shanxi. Written informed consent
was provided by all patients prior to surgery.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD, Fedewa SA, Ahnen DJ,
Meester RGS, Barzi A and Jemal A: Colorectal cancer statistics,
2017. CA Cancer J Clin. 67:177–193. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Araghi M, Soerjomataram I, Jenkins M,
Brierley J, Morris E, Bray F and Arnold M: Global trends in
colorectal cancer mortality: Projections to the year 2035. Int J
Cancer. 144:2992–3000. 2019. View Article : Google Scholar
|
|
4
|
Jiang S, Miao D, Wang M, Lv J, Wang Y and
Tong J: MiR-30-5p suppresses cell chemoresistance and stemness in
colorectal cancer through USP22/Wnt/β-catenin signaling axis. J
Cell Mol Med. 23:630–640. 2019. View Article : Google Scholar
|
|
5
|
Claessen MM, Schipper ME, Oldenburg B,
Siersema PD, Offerhaus GJ and Vleggaar FP: WNT-pathway activation
in IBD-associated colorectal carcinogenesis: Potential biomarkers
for colonic surveillance. Cell Oncol. 32:303–310. 2010.PubMed/NCBI
|
|
6
|
Cheng H, Sun X, Li J, He P, Liu W and Meng
X: Knockdown of Uba2 inhibits colorectal cancer cell invasion and
migration through downregulation of the Wnt/β-catenin signaling
pathway. J Cell Biochem. 119:6914–6925. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bienz M and Clevers H: Linking colorectal
cancer to Wnt signaling. Cell. 103:311–320. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Emons G, Spitzner M, Reineke S, Möller J,
Auslander N, Kramer F, Hu Y, Beissbarth T, Wolff HA, Rave-Fränk M,
et al: Chemoradiotherapy resistance in colorectal cancer cells is
mediated by Wnt/β-catenin signaling. Mol Cancer Res. 15:1481–1490.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Myant KB, Cammareri P, McGhee EJ, Ridgway
RA, Huels DJ, Cordero JB, Schwitalla S, Kalna G, Ogg EL, Athineos
D, et al: ROS production and NF-κB activation triggered by RAC1
facilitate WNT-driven intestinal stem cell proliferation and
colorectal cancer initiation. Cell Stem Cell. 12:761–773. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Peifer M and Polakis P: Wnt signaling in
oncogenesis and embryogenesis-a look outside the nucleus. Science.
287:1606–1609. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sparks AB, Morin PJ, Vogelstein B and
Kinzler KW: Mutational analysis of the APC/beta-catenin/Tcf pathway
in colorectal cancer. Cancer Res. 58:1130–1134. 1998.PubMed/NCBI
|
|
13
|
Sawa M, Masuda M and Yamada T: Targeting
the Wnt signaling pathway in colorectal cancer. Expert Opin Ther
Targets. 20:419–429. 2016. View Article : Google Scholar
|
|
14
|
Olsen AK, Coskun M, Bzorek M, Kristensen
MH, Danielsen ET, Jørgensen S, Olsen J, Engel U, Holck S and
Troelsen JT: Regulation of APC and AXIN2 expression by intestinal
tumor suppressor CDX2 in colon cancer cells. Carcinogenesis.
34:1361–1369. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shu Z, Chen L and Ding D: miR-582-5P
induces colorectal cancer cell proliferation by targeting
adenomatous polyposis coli. World J Surg Oncol. 14:2392016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee RC, Feinbaum RL and Ambros V: The C.
elegans heter-ochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Alvarez-Garcia I and Miska EA: MicroRNA
functions in animal development and human disease. Development.
132:4653–4662. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ha M and Kim VN: Regulation of microRNA
biogenesis. Nat Rev Mol Cell Biol. 15:509–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Thomas J, Ohtsuka M, Pichler M and Ling H:
MicroRNAs: Clinical relevance in colorectal cancer. Int J Mol Sci.
16:28063–28076. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Iwaya T, Yokobori T, Nishida N, Kogo R,
Sudo T, Tanaka F, Shibata K, Sawada G, Takahashi Y, Ishibashi M, et
al: Downregulation of miR-144 is associated with colorectal cancer
progression via activation of mTOR signaling pathway.
Carcinogenesis. 33:2391–2397. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Slattery ML, Herrick JS, Pellatt DF,
Stevens JR, Mullany LE, Wolff E, Hoffman MD, Samowitz WS and Wolff
RK: MicroRNA profiles in colorectal carcinomas, adenomas and normal
colonic mucosa: Variations in miRNA expression and disease
progression. Carcinogenesis. 37:245–261. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sanches JGP, Xu Y, Yabasin IB, Li M, Lu Y,
Xiu X, Wang L, Mao L, Shen J, Wang B, et al: miR-501 is upregulated
in cervical cancer and promotes cell proliferation, migration and
invasion by targeting CYLD. Chem Biol Interact. 285:85–95. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Y, Chai Y, Zhang J and Tang J: A
Function variant at miR-501 alters susceptibility to hepatocellular
carcinoma in a Chinese Han population. Cell Physiol Biochem.
38:2500–2508. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
26
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:e050052015. View Article : Google Scholar :
|
|
27
|
Moreno-Bueno G, Hardisson D, Sánchez C,
Sarrió D, Cassia R, García-Rostán G, Prat J, Guo M, Herman JG,
Matías-Guiu X, et al: Abnormalities of the APC/beta-catenin pathway
in endometrial cancer. Oncogene. 21:7981–7990. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gumbiner BM: Carcinogenesis: A balance
between beta-catenin and APC. Curr Biol. 7:R443–R446. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tetsu O and McCormick F: Beta-catenin
regulates expression of cyclin D1 in colon carcinoma cells. Nature.
398:422–426. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gomase VS and Parundekar AN: MicroRNA:
Human disease and development. Int J Bioinform Res Appl. 5:479–500.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bailey ST, Westerling T and Brown M: Loss
of estrogen-regulated microRNA expression increases HER2 signaling
and is prognostic of poor outcome in luminal breast cancer. Cancer
Res. 75:436–445. 2015. View Article : Google Scholar :
|
|
32
|
Wang Y, Gu J, Roth JA, Hildebrandt MA,
Lippman SM, Ye Y, Minna JD and Wu X: Pathway-based serum microRNA
profiling and survival in patients with advanced stage non-small
cell lung cancer. Cancer Res. 73:4801–4809. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yokoi A, Matsuzaki J, Yamamoto Y, Yoneoka
Y, Takahashi K, Shimizu H, Uehara T, Ishikawa M, Ikeda SI, Sonoda
T, et al: Integrated extracellular microRNA profiling for ovarian
cancer screening. Nat Commun. 9:43192018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu C, Liu R, Zhang D, Deng Q, Liu B, Chao
HP, Rycaj K, Takata Y, Lin K, Lu Y, et al: MicroRNA-141 suppresses
prostate cancer stem cells and metastasis by targeting a cohort of
pro-metastasis genes. Nat Commun. 8:142702017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yu J, Lei R, Zhuang X, Li X, Li G, Lev S,
Segura MF, Zhang X and Hu G: MicroRNA-182 targets SMAD7 to
potentiate TGFβ-induced epithelial-mesenchymal transition and
metastasis of cancer cells. Nat Commun. 7:138842016. View Article : Google Scholar
|
|
36
|
Hara N, Kikuchi M, Miyashita A, Hatsuta H,
Saito Y, Kasuga K, Murayama S, Ikeuchi T and Kuwano R: Serum
microRNA miR-501-3p as a potential biomarker related to the
progression of Alzheimer's disease. Acta Neuropathol Commun.
5:102017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
James MI, Iwuji C, Irving G, Karmokar A,
Higgins JA, Griffin-Teal N, Thomas A, Greaves P, Cai H, Patel SR,
et al: Curcumin inhibits cancer stem cell phenotypes in ex vivo
models of colorectal liver metastases, and is clinically safe and
tolerable in combination with FOLFOX chemotherapy. Cancer Lett.
364:135–141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Boman BM, Fields JZ, Bonham-Carter O and
Runquist OA: Computer modeling implicates stem cell overproduction
in colon cancer initiation. Cancer Res. 61:8408–8411.
2001.PubMed/NCBI
|
|
39
|
Novellasdemunt L, Antas P and Li VS:
Targeting Wnt signaling in colorectal cancer. A review in the
theme: Cell signaling: Proteins, pathways and mechanisms. Am J
Physiol Cell Physiol. 309:C511–C521. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang N, Li X, Wu CW, Dong Y, Cai M, Mok
MT, Wang H, Chen J, Ng SS, Chen M, et al: microRNA-7 is a novel
inhibitor of YY1 contributing to colorectal tumorigenesis.
Oncogene. 32:5078–5088. 2013. View Article : Google Scholar
|
|
41
|
Zhang H, Hao Y, Yang J, Zhou Y, Li J, Yin
S, Sun C, Ma M, Huang Y and Xi JJ: Genome-wide functional screening
of miR-23b as a pleiotropic modulator suppressing cancer
metastasis. Nat Commun. 2:5542011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nagel R, le Sage C, Diosdado B, van der
Waal M, Oude Vrielink JA, Bolijn A, Meijer GA and Agami R:
Regulation of the adenomatous polyposis coli gene by the miR-135
family in colorectal cancer. Cancer Res. 68:5795–5802. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lin Y, Gu Q, Sun Z, Sheng B, Qi C, Liu B,
Fu T, Liu C and Zhang Y: Upregulation of miR-3607 promotes lung
adenocar-cinoma proliferation by suppressing APC expression. Biomed
Pharmacother. 95:497–503. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu L, Zhang H, Shi L, Zhang W, Yuan J,
Chen X, Liu J, Zhang Y and Wang Z: Inhibition of Rac1 activity
induces G1/S phase arrest through the GSK3/cyclin D1 pathway in
human cancer cells. Oncol Rep. 32:1395–1400. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lee SH, Chen TY, Dhar SS, Gu B, Chen K,
Kim YZ, Li W and Lee MG: A feedback loop comprising PRMT7 and
miR-24-2 interplays with Oct4, Nanog, Klf4 and c-Myc to regulate
stemness. Nucleic Acids Res. 44:10603–10618. 2016. View Article : Google Scholar : PubMed/NCBI
|