Introduction
At present, skin melanoma (SM) remains the most
frequently occurring and aggressive type of cutaneous cancer, and
it is estimated that, in addition to the markedly increased
incidence of SM over the course of the last decade, there will be
96,480 newly diagnosed cases of SM in the USA in 2019 (1). Despite the continuing improvements
that have been made in terms of diagnosis and treatment, the
recommended maintenance schedules, from radical resection to
molecular-targeted drugs, at present are only effective in a subset
of patients (2,3). The tumors are often found to have
metastasized to distant sites or visceral organs in patients even
at an early stage of diagnosis, and the 5-year overall survival
rate remains extremely disappointing for this subset of patients,
with a median survival time of 6-9 months (4). Therefore, in order to establish novel
treatment therapies for SM, it is important to explore in detail
the underlying molecular mechanisms and identify novel molecules to
counteract the progression of SM.
Long non-coding RNAs (lncRNAs) are a class of
non-protein-coding transcripts (>200 nucleotides in length) that
have been reported to serve pivotal roles in physiological and
pathophysiological processes associated with the reproductive,
metabolic, endocrine, cardiovascular and nervous systems (5-7). Of
greater importance considering the present study, lncRNAs have been
also identified as crucial regulators in the carcinogenesis and
progression of multiple types of cancer, where they function as
oncogenes or tumor suppressors, depending on the cancer type and
the prevailing circumstances (8-10).
In the case of SM, an increasing body of evidence has indicated
that lncRNAs are also involved in the regulation of the expression
and function of established oncogenes or tumor suppressors, either
post-transcriptionally or via chromatin regulation (11-13).
Nevertheless, only a small number of lncRNAs have been functionally
characterized at present, and the mechanisms underlying their
biological functions are yet to be fully elucidated.
Long intergenic noncoding RNA 00961 (Linc00961),
located in chromosome 9 and 1,546 nucleotides in length, was
reported to be an encoder of small regulatory polypeptide of amino
acid response (SPAAR), and is involved in muscle regeneration
(14,15). Furthermore, recent studies have
also identified Linc00961 as a tumor suppressor in multiple types
of cancers, including glioma (16)
and non-small cell lung cancer (NSCLC) (17). However, the expression profile,
biological function and clinical significance of Linc00961 have not
been fully investigated.
The aims of the present study were to identify the
expression and function of Linc00961 in SM. It was revealed that
Linc00961 was downregulated in SM, and functioned as a competitive
endogenous RNA (ceRNA) to positively regulate the expression of
phosphate and tension homology deleted on chromosome 10 (PTEN) by
sponging microRNA (miRNA/miR)-367 in A375 and SL-MEL-28 cells.
Collectively, these results demonstrated the tumor suppressor role
of Linc00961 in the carcinogenesis and progression of SM.
Materials and methods
Clinical specimens
The present study was performed in accordance with
the principles of the Declaration of Helsinki and approved by the
Ethical Committee of the First Affiliated Hospital of Xi'an
Jiaotong University. Pathologically diagnosed SM or benign nevi
(BN) patients were enrolled in this study. Tissues from a total of
13 patients with SM (aged 34-68 years; 5 male and 8 female) and 13
BN tissues from different cohorts of patients (aged 29-67 years; 5
male and 8 female) were collected at the First Affiliated Hospital
of Xi'an Jiaotong University between January 2010 and August 2017.
All the patients enrolled in the present study provided their
informed consent for the use of their resected tissues in this
research.
Cell culture
Human A375 (cat. no. GCC-ME0001CS), SK-MEL-28 (cat.
no GCC-ME0003CS) and A2058 (cat. no. GCC-ME0004CS) SM cell lines,
and human epidermal HEMa-LP melanocytes (cat. no. GCC-NC0027CS)
were all purchased from Shanghai GeneChem Co., Ltd., where they
were characterized by mycoplasma detection, isozyme detection and
DNA fingerprinting. HEMa-LP cells were cultured in
Invitrogen® Medium 254 (Thermo Fisher Scientific, Inc.)
supplemented with human melanocyte growth supplement (Thermo Fisher
Scientific, Inc.), and A375, A2058, and SK-MEL-28 cells were
cultured in Gibco® Dulbecco's Modified Eagle's medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10%
fetal bovine serum (CellMax) and 100 U/ml penicillin and
streptomycin (CellMax) at 37°C in a humidified atmosphere of 5%
CO2.
Bioinformatics analysis
The expression of Linc00961 in the Gene Expression
Omnibus (https://www.ncbi.nlm.nih.gov/geo/) dataset
(GSM3071633) (18), containing
data for 57 patients with SM and 23 patients with BN, was analyzed
using the R2: Genomics Analysis and Visualization Platform
(http://r2.amc.nl). The associations between Linc00961
expression and the overall survival of patients with SM in The
Cancer Genome Atlas (TCGA) database (https://cancergenome.nih.gov/) was analyzed using Gene
Expression Profiling Interactive Analysis (http://gepia.cancer-pku.cn). High and low expression
were predefined in this cohort prior to the present study. The
potential miRNAs containing putative binding sites for Linc00961
were identified using miRDB (version 2.0; http://mirdb.org/).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA/miRNA extraction and RT-qPCR were performed as
previously described in a study by the present authors (19). In brief, total RNA was extracted
form A375, A2058, and SK-MEL-28 cells or the tissues of patients
with SM. RNA samples were then reverse transcribed into cDNA
(PrimeScript™ RT Reagent kit; Takara Bio, Inc.). Then, SYBR Green
Mix (Takara Bio, Inc.) was used to detect and quantify Linc00961,
miR-367, β-actin and U6 expression. The thermocycling conditions
were as follows: Denaturation at 95°C for 7 min, followed by 35
cycles of 95°C for 5 sec, 60°C for 30 sec, 72°C for 3 min. The
relative expression level was calculated using the
2-ΔΔCq method (20).
The primers used in this study were purchased from Sangon Biotech
Co., Ltd., and their sequences are presented in Table SI.
Lentivirus vector construction and
transfection
Lentivirus vector overexpressing Linc00961
(Lv-Linc00961) was designed and constructed by Shanghai GeneChem
Co., Ltd. Briefly, the full length of Linc00961 was chemically
synthesized and cloned into the BamHI and AgeI sites
of a CV146 lentivirus core vector. Subsequently, CV146-Linc00961
(20 µg) and lentiviral packaging helper plasmids, Helper 1.0
(15 µg) and Helper 2.0 (10 µg; both Shanghai GeneChem
Co., Ltd.), were co-transfected into 293T cells using
Lipofectamine™ 2000 (Thermo Fisher Scientific, Inc.). The cell
supernatant was collected 48 h later, and subjected to
centrifugation (4°C, 4,000 x g, 10 min) in order to concentrate and
purify Lv-Linc00961. Cells stably expressing Linc00961 were
constructed by transfecting 5×105 TU Lv-Linc00961 or
Lv-control (containing empty CV146 vector) into 2×106
A375 or SK-MEL-28 cells cultured in 1 ml enhanced infection
solution (Shanghai GeneChem Co., Ltd.). Then, 5 µg polybrene
(Shanghai GeneChem Co., Ltd.) was added in each well according to
the manufacturer's protocol (17).
Transfected cells were harvested 72 h later, and subsequently
treated with puromycin (2 µg/ml) in order to isolate the
Linc00961-stably-expressing cells.
Small interfering RNA (siRNA) and miRNA
transfection
PTEN siRNA (sense, 5′-GAG CGU GCA GAU AAU GAC A-3′;
antisense, 3′-CUC GCA CGU CUA UUA CUG U-5′), negative control (NC)
siRNA (sense, 5′-UUC UCC GAA CGU GUC AGG UTT-3′; antisense, 3′-ACC
UGA CAC GUU CGG AGA ATT-5′), miR-367 mimics (5′-AGU GGU AAC GAU UUC
ACG UUA A-3′), and miR-367 NC (5′-GUG GAU AUU GUU GCC AUC A-3′)
were purchased from Shanghai GenePharma Co., Ltd. Transfection was
performed as previously described (19). Briefly, 1×106 A375 or
SK-MEL-28 cells were seeded in 6-well plates, PTEN siRNA (50 nM),
normal control siRNA (50 nM), miR-367 mimics (50 nM) or miR-367 NC
(50 nM) were mixed with 5 µl Lipofectamine 2000 in 500
µl serum-free DMEM. Then, the solutions were added to each
well. The cells were collected at 72 h after transfections for Cell
Counting Kit-8 (CCK-8), flow cytometry (FCM) and Transwell
assays.
CCK-8, FCM and Transwell assays
A375 and SK-MEL-28 cells were transfected with
Lv-Linc00961, Lv-Linc00961 + miR-367 mimics, Lv-Linc00961 + PTEN
siRNA and corresponding controls as aforementioned. Then, the
proliferation of transfected cells was measured using a CCK-8 assay
(MedChemExpress) at 0, 24, 48, 72, 96 and 120 h following
transfection, whereas cell apoptosis was measured using an
Annexin-V/PI kit (BD Biosciences); the percentage of early
apoptotic cells was analyzed using FlowJo 10 software (FlowJo LLC),
and cell migration and invasion were measured using Transwell
(Corning Inc.) and Matrigel (BD Biosciences) assays at 72 h
following transfection. All the procedures for the CCK-8, FCM and
Transwell assays were performed as described in our previous study
(19).
For the Transwell assays, briefly, A375 and
SK-MEL-28 cells were suspended in serum-free DMEM at a density of
4×105 cells/ml. Then, cells (200 µl) were seeded
in the upper chamber coated with (for the invasion assay) or
without (for the migration assay) Matrigel (BD Biosciences). DMEM
with 10% FBS (600 µl) was added to the lower chamber. After
the cells had been allowed to migrate for 24 h or to invade for 48
h, the penetrated cells on the filter were fixed in dried methanol
for 1 min and stained in 4 g/l crystal violet for 10 min (both at
room temperature). The numbers of migrated or invasive cells were
determined from five random fields using a light microscope
(magnification, ×100; Olympus Corporation).
Western blotting
Proteins were extracted from the transfected cells
at 72 h after transfection using RIPA buffer (cat. no. P0013B;
Beyotime Institute of Biotechnology) and quantified using a
bicinchoninic acid protein assay kit (cat. no. P0011; Beyotime
Institute of Biotechnology). Then, 20 µg/lane protein from
different transfected cells was separated via 10% SDS-PAGE and
transferred to PVDF membranes. The PVDF membranes were blocked with
1X Blocker™ BSA in TBS (cat. no. 37520; Thermo Fisher Scientific,
Inc.) for 2 h at room temperature. Then, the PVDF membranes were
incubated overnight at 4°C with rabbit-anti-human PTEN (1:400; cat.
no. 9188; Cell Signaling Technology, Inc.) and β-actin (1:1,000,
cat. no. 4970; Cell Signaling Technology, Inc.) primary antibodies.
After incubating with the horseradish peroxidase-conjugated goat
anti-rabbit IgG secondary antibodies (1:5,000; cat. no. 7074; Cell
Signaling Technology, Inc.) for 2 h at room temperature, the
signals were detected using SignalFire™ Plus ECL Reagent (cat. no.
12630; Cell Signaling Technology, Inc.).
Fluorescence in situ hybridization
(FISH)
Oligonucleotide modified probes for Linc00961 (cat.
no. lnc1CM001) and U6 (cat. no. lnc110101) were purchased from
Guangzhou RiboBio Co., Ltd. SK-MEL-28 cells (1×106) were
seeded in 18-mm confocal dishes. Following overnight incubation,
the cells were fixed with 4% paraformaldehyde for 20 min and
permeabilized with Triton X-100 for 90 sec (both at room
temperature). Then, SK-MEL-28 cells were incubated with
hybridization buffer supplemented with 100 µl Linc00961 and
U6 FISH probes (20 µM) at 37°C overnight in a dark moist
chamber. The following day, cells were washed three times in 2X
saline-sodium citrate buffer and stained with DAPI for 5 min at
room temperature. The images were acquired using a confocal
microscope (magnification, ×400; Olympus Corporation).
In vivo xenograft experiments
For tumorigenesis assays, xenograft tumors were
generated by subcutaneous injection of A375 cells
(5×106) stably expressing Linc00961 or infected with
Lv-control into the hind limbs of female Balb/C athymic nude mice
(nu/nu; Animal Center of Xi'an Jiaotong University) weighing 18-20
g and aged 6 weeks old (n=3/group). All mice were housed and
maintained under specific pathogen-free conditions at 18-22°C, with
20% humidity, a 12 h light:12 h dark cycle, and with commercial rat
food and water ad libitum. All experiments were approved by
the Animal Care and Use Committee of Xi'an Jiaotong University and
performed in accordance with institutional guidelines. Tumor size
and weight were measured using Vernier calipers and an electronic
balance every 7 days. Tumor volume was determined according to the
formula: 0.5x AxB2, where A represents the diameter of
the base of the tumor, and B represents the corresponding
perpendicular value. After 35 days, the mice were weighed, and the
weights of the mice at 35 days were all between 20-22 g. Then, the
mice were anesthetized using isoflurane and sacrificed in a
CO2 chamber for 7 min, where the flow rate displaced no
more than 30% chamber volume/min. Once death was confirmed, the
tumors were collected and weighed. Symptoms such as pain, weight
loss, loss of appetite or weakness were set as humane endpoints for
the present study; however, no animal was sacrificed before the
completion of the 35-day experiment as a result of displaying any
of these symptoms.
TUNEL and Ki-67 staining
The resected xenograft tumors were fixed in 4%
paraformaldehyde for 24 h at room temperature. Then, resected
tumors were desiccated with ascending series of ethanol, cleared in
xylene, dipped in wax, embedded in paraffin and sectioned.
Subsequently, tissue sections (5-µm thick) were
deparaffinized and rehydrated. For TUNEL staining, the slides were
incubated with Proteinase K (20 µg/ml in 10 mM Tris/HCL, pH
7.4) for 30 min at 37°C and 50 µl TUNEL reaction mixture
(Roche Diagnostics) for 60 min at 37°C. Subsequently, the slides
were stained with 200 nM DAPI for 5 min at room temperature to
visualize the nuclei.
For Ki-67 staining, slides were incubated in citric
acid antigen retrieval solution for 100 sec at 110°C in an
autoclave. After washing with PBS for three times, the slides were
incubated with 3% H2O2 for 30 min at room
temperature to blocking endogenous peroxidases. The slides were
blocked with 1X Blocker™ BSA in TBS for 2 h at room temperature.
The mouse-anti-human primary antibody for Ki-67 (1:400; cat. no.
9449; Cell Signaling Technology, Inc.) was added to the slides,
which were incubated overnight at 4°C. Then, biotinylated
goat-anti-mouse IgG secondary antibodies (1:500; cat. no. 14709;
Cell Signaling Technology, Inc.) was added and incubated for 30 min
at room temperature. HRP-conjugated streptavidin was used to attach
peroxidase to biotinylated antibodies, and DAB chromogen was used
for visualization. Then, hematoxylin was used to nuclear
counterstain for 3.5 min at room temperature. The stained slides
were observed under a FluoView FV1000 microscope (magnification,
×200; Olympus Corporation), and 10 random fields per sample were
captured. FV10-ASW Viewer (version 4.2; Olympus Corporation) was
used to analyze images.
Luciferase assay
A375 and SK-MEL-28 cells were seeded in a 96-well
plate at 70% confluence. The 3′-untranslated region (UTR) of
Linc00961 containing miR-367-binding sites was cloned into a
pmirGLO dual-luciferase miRNA target expression vector (Promega
Corporation), yielding pmirGLO-wild type (WT)-Linc00961. Mutations
were introduced into potential miR-367-binding sites of Linc00961
using a QuikChange™ site-directed mutagenesis kit (Stratagene;
Agilent Technologies, Inc.) to construct pmirGLO-mutant
(MUT)-Linc00961. Subsequently, 20 nM miR-367 NC or mimics and 50 ng
pmirGLO-WT/MUT-Linc00961 were co-transfected into 1×105
A375 or SK-MEL-28 cells using Lipofectamine 2000 in 96 well plate.
Cells were collected 48 h after transfection, and the relative
firefly luciferase activities were measured using a dual-luciferase
reporter assay system according to the manufacturer's protocol
(Promega Corporation). Renilla luciferase activity served as
an internal control.
RNA pull-down assay
The procedures for RNA pull-down assay was performed
as described in our previous study (19). Briefly, full length Linc00961 and
Linc00961-MUT transcripts were transcribed from
pmirGLO-WT-Linc00961 and pmirGLO-MUT-Linc00961 in vitro. The
restriction enzyme BamHI was used to linearize the plasmids.
Then, Ribo™ RNA max-T7 RNA polymerase and Biotin RNA Labeling kit
(cat. no. C11002-1; Guangzhou RiboBio Co., Ltd.) was used to
produce biotin-labeled RNAs, according to the manufacturer's
instructions. Subsequently, 5 µg biotin-labeled Linc00961 or
Linc00961-MUT was incubated with 1 mg cell lysates, which were
lysed using NP40 solution (cat. no. P0013F; Beyotime Institute of
Biotechnology) at 4°C for 4 h. A negative control biotinylated
probe was also included in the Biotin Labeling kit. Subsequently,
the RNAs with biotin-labelled Linc00961 or Linc00961-MUT were mixed
with 40 µl Dynabeads® MyOne™ Streptavidin C1
beads (cat. no. 65002; Invitrogen; Thermo Fisher Scientific, Inc.)
and incubated on a rotator at 4°C overnight. Subsequently, the
pulled-down RNA was identified via RT-qPCR analysis, performed as
previously described.
Statistical analysis
IBM SPSS statistical software (version 22.0) was
used to perform statistical analysis. The experiments were repeated
three times, and data were presented as the mean ± standard
deviation. Student's t-test (for analysis of groups) and one-way
ANOVA followed by least significant difference testing (for
analysis of multiple groups) were used for data analysis.
Kaplan-Meier curve and log-rank tests were used to analyze the
survival rates of different groups. Pearson's correlation analysis
was used to analyze the correlations between Linc00961 and miR-367
expression in patient with SM. P<0.05 was considered to indicate
a statistically significant difference.
Results
Linc00961 is downregulated in tissues and
cells of SM
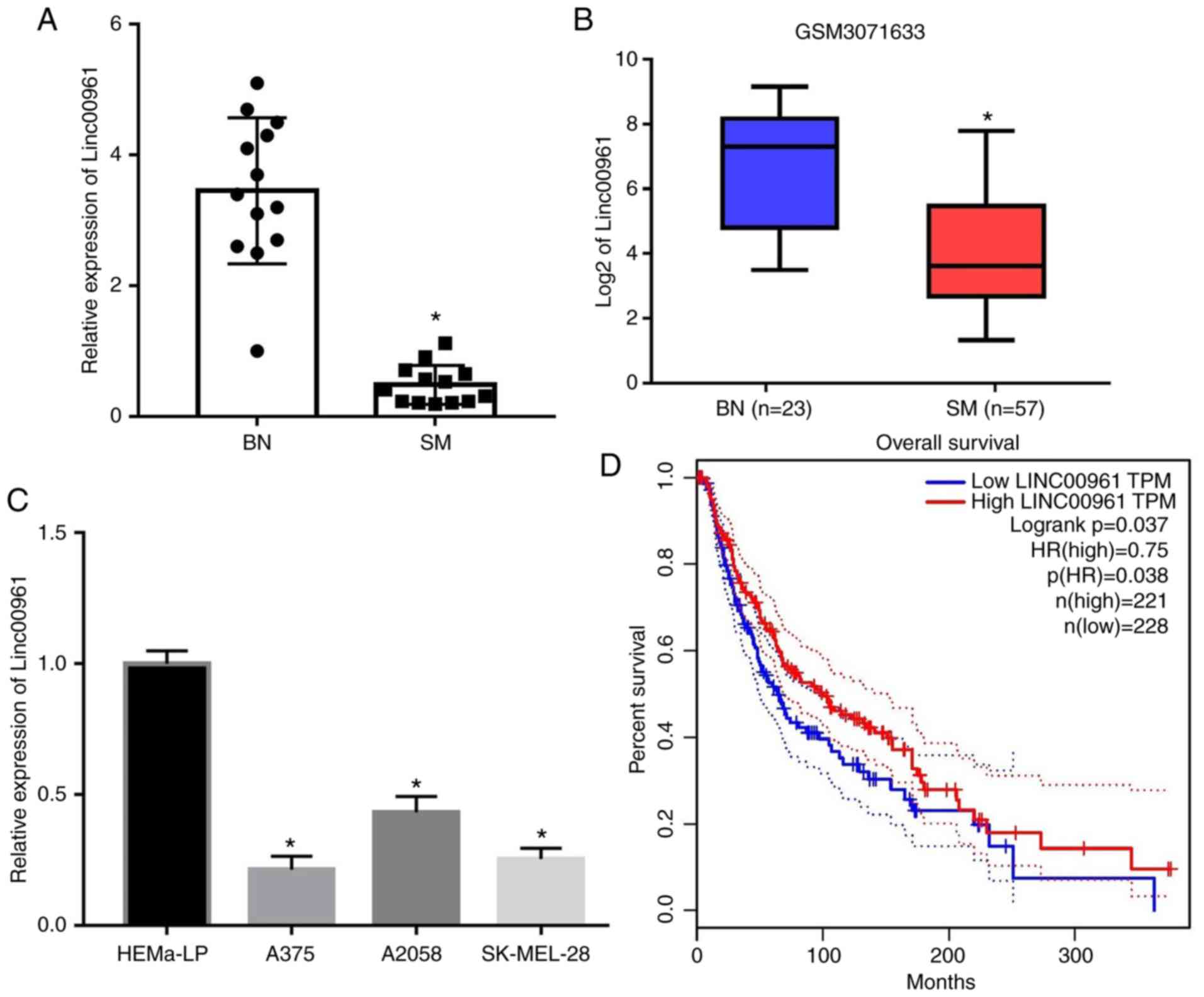
RT-qPCR analysis indicated that Linc00961 was
significantly downregulated in the tissues of 13 patients with SM
compared with 13 patients with BN (P<0.05; Fig. 1A). Furthermore, analysis of the
GSM3071633 dataset also revealed decreased expression levels of
Linc00961 in SM tissues compared with BN tissues (P<0.05;
Fig. 1B). In addition, Linc00961
levels were also significantly downregulated in SM cells (A375,
A2058 and SK-MEL-28) compared with the human epidermal melanocyte
cell line, HEMa-LP (P<0.05; Fig.
1C). Subsequently, the prognostic value of Linc00961 expression
levels for SM was investigated by analyzing TCGA data. It was
revealed that the overall survival of patients with SM exhibiting
high Linc00961 levels was significantly higher compared with those
with low Linc00961 levels (Fig.
1D). These results indicated that Linc00961 was a potential
tumor suppressor in SM. Therefore, in a subsequent experiment, the
Linc00961 levels in A375 and SK-MEL-28 cells were upregulated using
Lv-Linc00961 (P<0.05; Fig.
S1A), and the effects of Linc00961 on the proliferation and
invasion of A375 and SK-MEL-28 cells were further investigated.
Effects of Linc00961 on the proliferation
and apoptosis of A375 and SK-MEL-28 cells in vitro
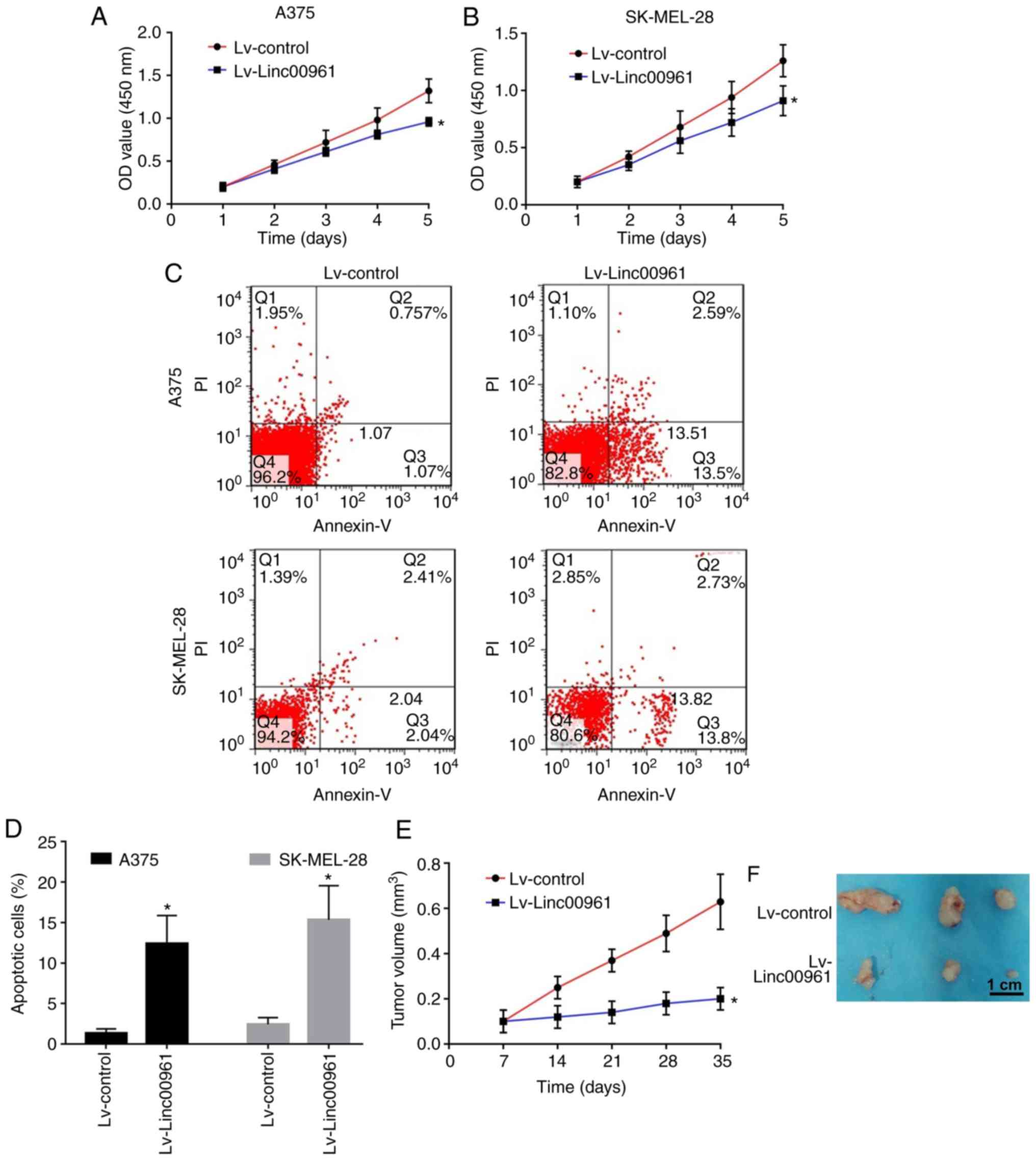
A CCK-8 assay revealed that the proliferation rates
of A375 and SK-MEL-28 cells transfected with Lv-Linc00961 were
significantly reduced compared with those of cells transfected with
Lv-control (P<0.05; Fig. 2A and
B). FCM assays indicated that the early apoptosis of A375 and
SK-MEL-28 cells transfected with Lv-Linc00961 was promoted compared
with in cells transfected with Lv-control (P<0.05; Fig. 2C and D).
Effects of Linc00961 on tumor growth and
apoptosis of nude mice bearing A375 cells in vivo
Subsequently, the effects of Linc00961 on tumor
growth in nude mice injected with A375 cells were determined. As
shown by the growth curves, slower tumor growth and smaller tumor
volumes were observed in the mice injected with A375 cells
transfected with Lv-Linc00961 compared with Lv-control (P<0.05;
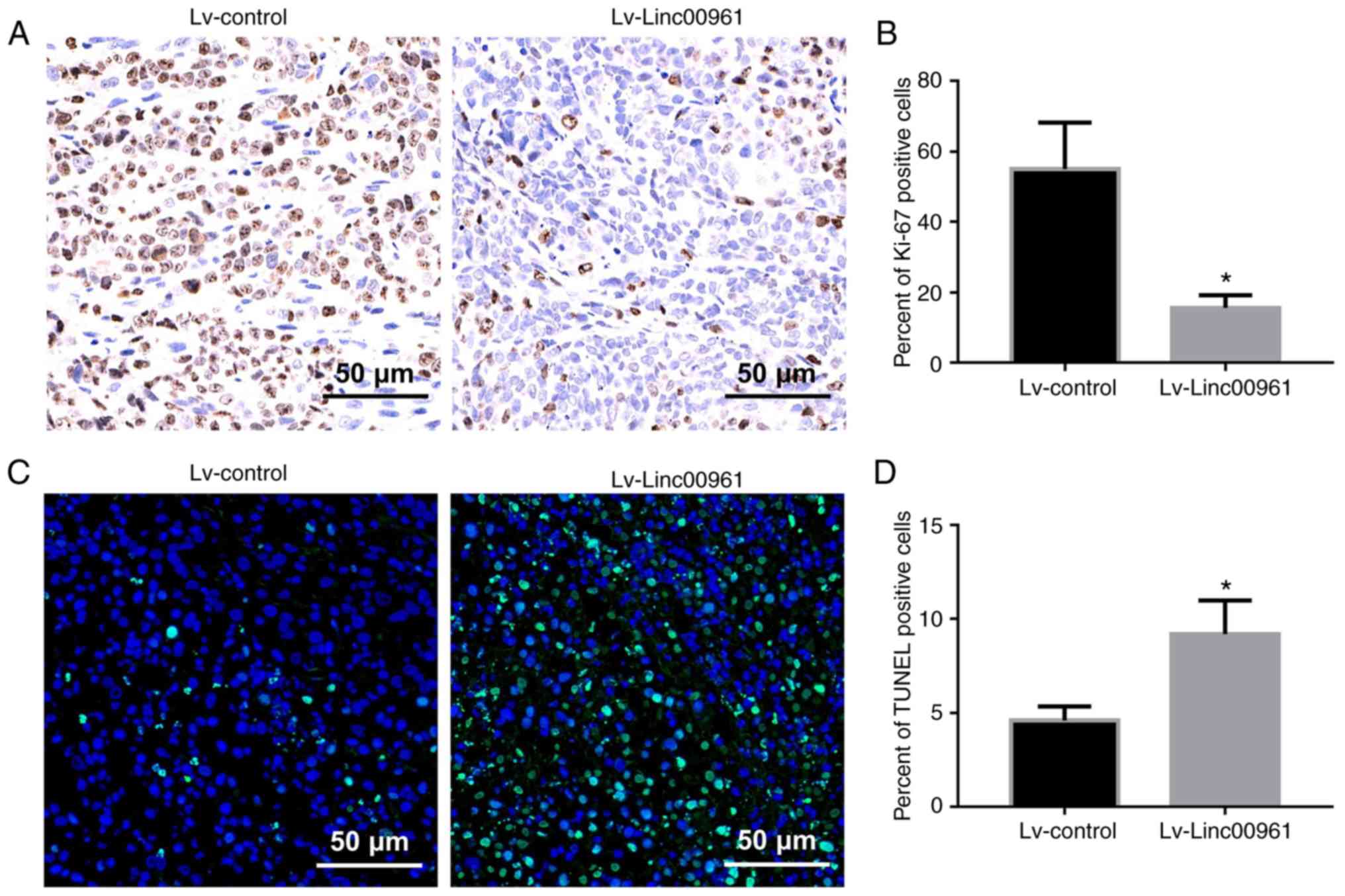
Fig. 2E and F). Furthermore, Ki-67
assays revealed that the percentage of Ki-67 positive cells was
significantly downregulated in mice injected with A375 cells
transfected with Lv-Linc00961 (P<0.05; Fig. 3A and B). However, TUNEL assays
revealed a more intense fluorescence signal indicative of apoptosis
in the tissues of mice injected with A375 cells transfected with
Lv-Linc00961 (P<0.05; Fig. 3C and
D).
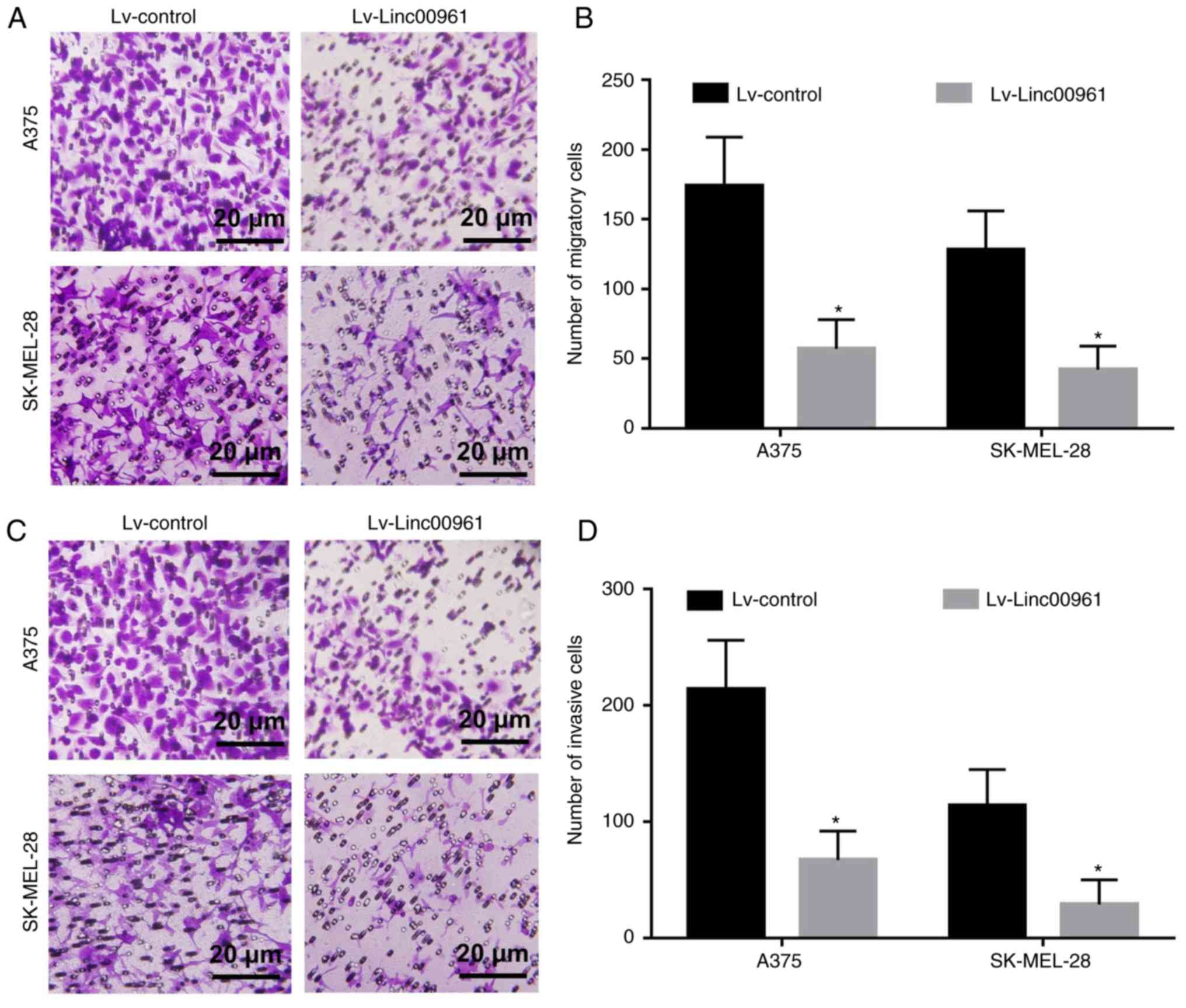
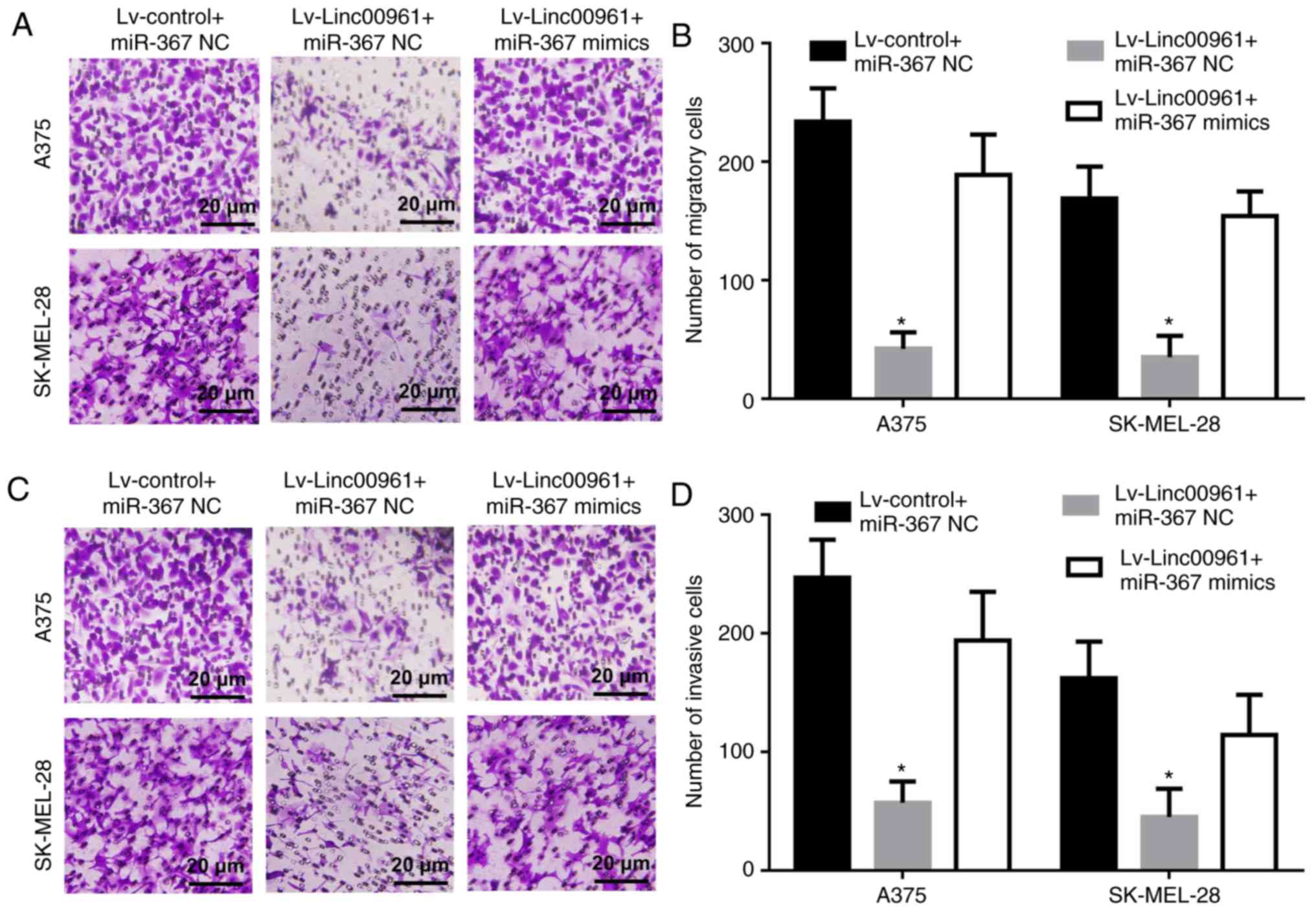
Effects of Linc00961 on the migration and
invasion of A375 and SK-MEL-28 cells in vitro
The effects of Linc00961 on the migration and
invasion of A375 and SK-MEL-28 cells were further explored. Cell
migration assays revealed that the numbers of migrating A375 and
SK-MEL-28 cells transfected with Lv-Linc00961 were reduced compared
with cells transfected with Lv-control (P<0.05; Fig. 4A and B). Cell invasion assays
demonstrated that the numbers of invasive A375 and SK-MEL-28 cells
transfected with Lv-Linc00961 were also decreased compared with
cells transfected with Lv-control (P<0.05; Fig. 4C and D).
Linc00961 is inversely correlated with
miR-367 in SM
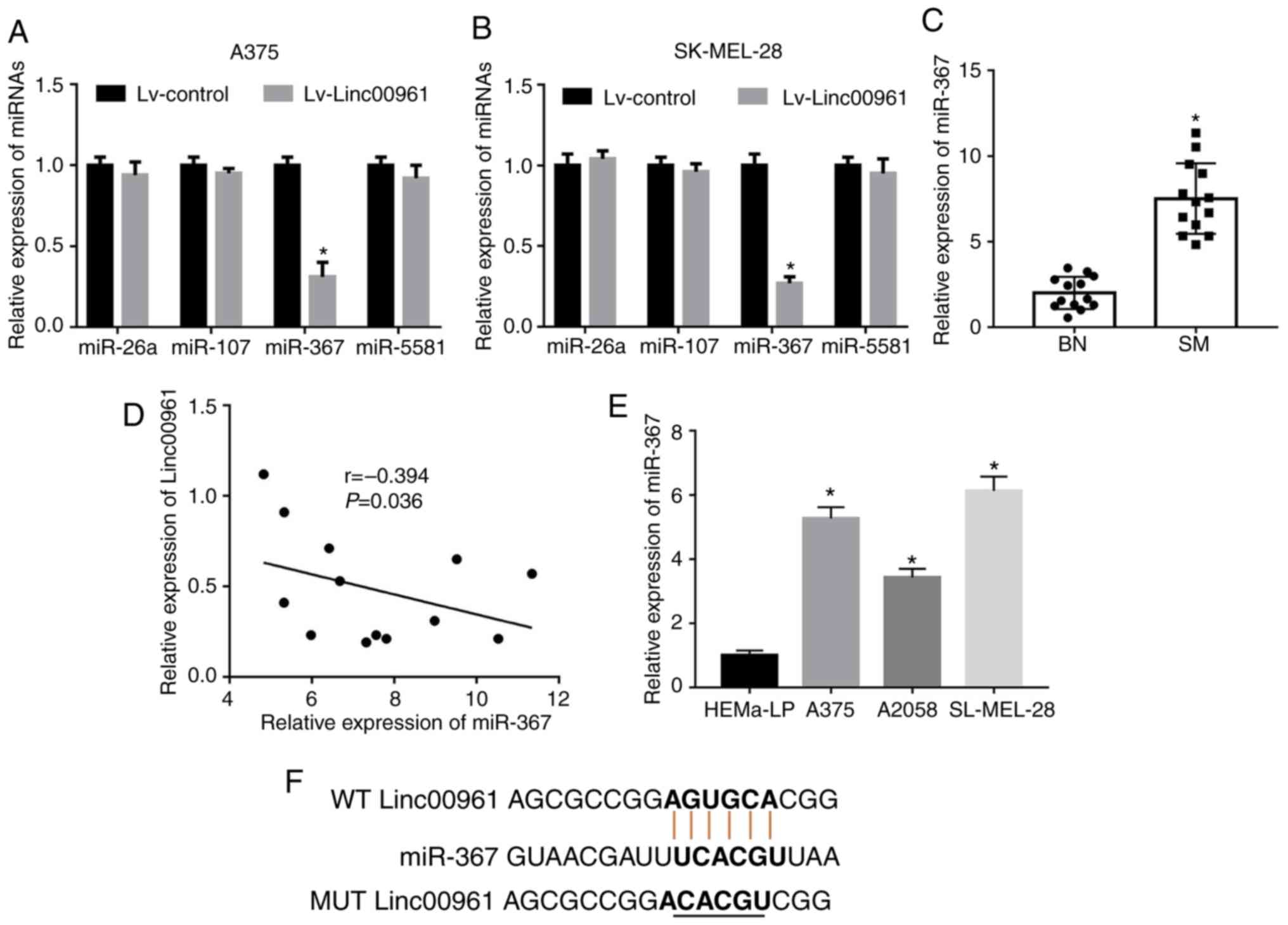
To explore the underlying mechanisms of Linc00961 in
SM cells, miR-26a, miR-107, miR-367 and miR-5581 were identified as
potential targeting miRNAs containing putative binding sites for
Linc00961 via a search of an online bioinformatics database
(miRDB). Subsequently, the expression levels of miR-367 were
revealed to be significantly downregulated following the
upregulation of Linc00961 expression in A375 (P<0.05; Fig. 5A) and SK-MEL-28 cells (P<0.05;
Fig. 5B). However, the expression
levels of miR-26a, miR-107 and miR-5581 were not significantly
altered following overexpression of Linc00961 in A375 (P>0.05;
Fig. 5A) or SK-MEL-28 (P>0.05;
Fig. 5B) cells. The results from
RT-qPCR analysis revealed that miR-367 levels were upregulated in
SM tissues (P<0.05; Fig. 5C),
which were inversely correlated with Linc00961 levels (r=-0.394;
P<0.05; Fig. 5D). Furthermore,
miR-367 expression was also enhanced in SM cells (A375, A2058 and
SK-MEL-28) compared with normal melanocytes (P<0.05; Fig. 5E).
Linc00961 directly binds with miR-367 in
SM cells
To confirm whether Linc00961 could function as a
ceRNA to competitively bind with miR-367, FISH was performed to
identify the subcellular localization of Linc00961 in SK-MEL-28
cells. Linc00961 was revealed to be mainly expressed in cytoplasm
and the nuclei (Fig. S2),
indicating that Linc00961 could both function as a ceRNA and bind
with RNA-binding proteins in SM cells. Subsequently, WT-Linc00961
and MUT-Linc00961 vectors were constructed to further explore the
association between Linc00961 and miR-367 in SM (Fig. 5F). First, it was revealed that
miR-367 expression levels were downregulated in A375 and SK-MEL-28
cells transfected with Lv-Linc00961 compared with A375 cells and
SK-MEL-28 cells transfected with Lv-control (P<0.05; Fig. 6A). Dual-luciferase reporter assays
indicated that the relative luciferase activities in A375
(P<0.05; Fig. 6B) and SK-MEL-28
cells (P<0.05; Fig. 6C)
co-transfected with Linc00961-WT and miR-367 mimics were
significantly decreased compared with cells transfected with
miR-367 NC. By contrast, co-transfecting Linc00961-MUT and miR-367
mimics into A375 (P>0.05; Fig.
6B) and SK-MEL-28 cells (P>0.05; Fig. 6C) did not significantly affect the
relative luciferase activity. Furthermore, biotin-labeled RNA
pulldown assays demonstrated that a bio-Linc00961 probe could
directly pull down miR-367, but the associated
bio-Linc00961-MUT-probe failed to pull down miR-367 in A375
(P<0.05; Fig. 6D and E) or
SK-MEL-28 cells (P<0.05; Fig. 6D
and F). Collectively, these results indicated that Linc00961
could directly sponge miR-367 at special recognition sites in SM
cells.
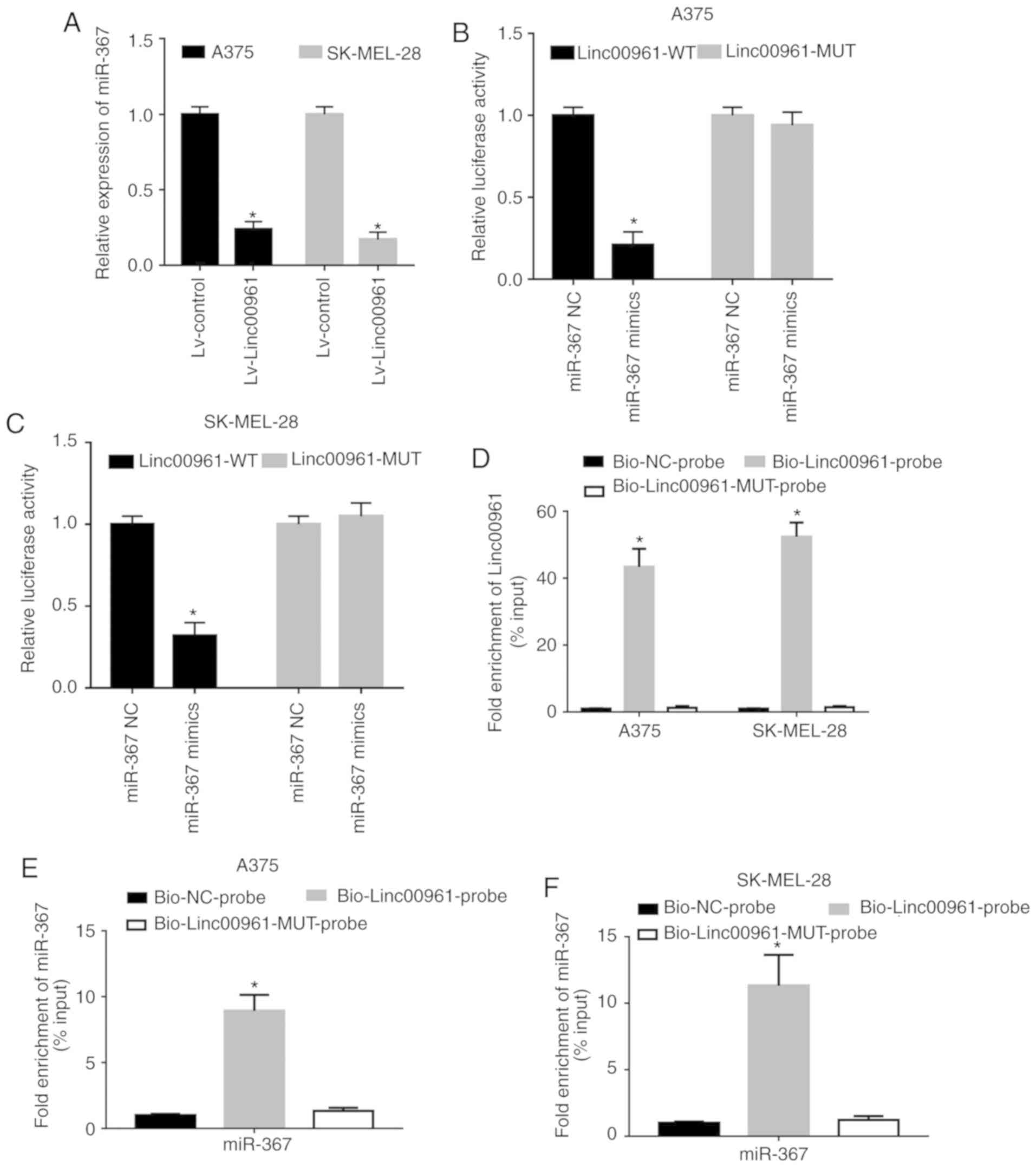 | Figure 6Linc00961 directly sponges miR-367 in
SM cells. (A) Effects of Linc00961 on the expression of miR-367 in
SM cells. Relative luciferase activity in (B) A375 and (C)
SK-MEL-28 cells co-transfected with Linc00961-WT or LINC00961-MUT
luciferase plasmid and miR-367 NC or mimics. (D) Detection of
LINC00961 in RNA pulled down by Bio-Linc00961, Bio-Linc00961-MUT or
Bio-NC probes. Detection of miR-367 in RNA pulled down by
Bio-Linc00961, Bio-Linc00961-MUT or Bio-NC probes in (E) A375 and
(F) SK-MEL-28 cells. Data from three experiments are presented as
the mean ± SD. *P<0.05 vs. control (Lv-control,
miR-367 NC, Bio-NC-probe). Linc00961, long intergenic noncoding RNA
00961; miR, microRNA; SM, skin melanoma; NC, negative control; WT,
wild-type; MUT, mutant; Bio, biotinylated. |
Linc00961 functions as a ceRNA to inhibit
the proliferation and invasion of A375 and SK-MEL-28 cells by
directly sponging miR-367
To investigate whether Linc00961 inhibited the
malignant behaviors of A375 and SK-MEL-28 cells via binding to
miR-367, which was established as an oncogenic miRNA in SM
(21), the combined effects of
overexpression of both Linc00961 and miR-367 on proliferation and
invasion of A375 and SK-MEL-28 cells were examined. miR-367 mimics
were transfected into A375 and SK-MEL-28 cells in order to
upregulate miR-367 expression (P<0.05; Fig. S1B). Subsequently, it was confirmed
that the decreased miR-367 expression level resulted from Linc00961
overexpression, and that recovery of the expression of miR-367 was
identified following co-transfection with miR-367 mimics (Fig. 7A). Subsequently, CCK-8 and FCM
assays were employed to investigate the effects of Lv-Linc00961 on
the proliferation and apoptosis of A375 and SK-MEL-28 cells, and
these inhibitory effects were rescued by co-transfection of miR-367
mimics (P<0.05; Fig. 7B-E).
Similarly, Transwell assays demonstrated that up-regulation of
miR-367 could reverse the inhibitory effects of Lv-Linc00961 on
cell migration and invasion of A375 and SK-MEL-28 cells (P<0.05;
Fig. 8A-D).
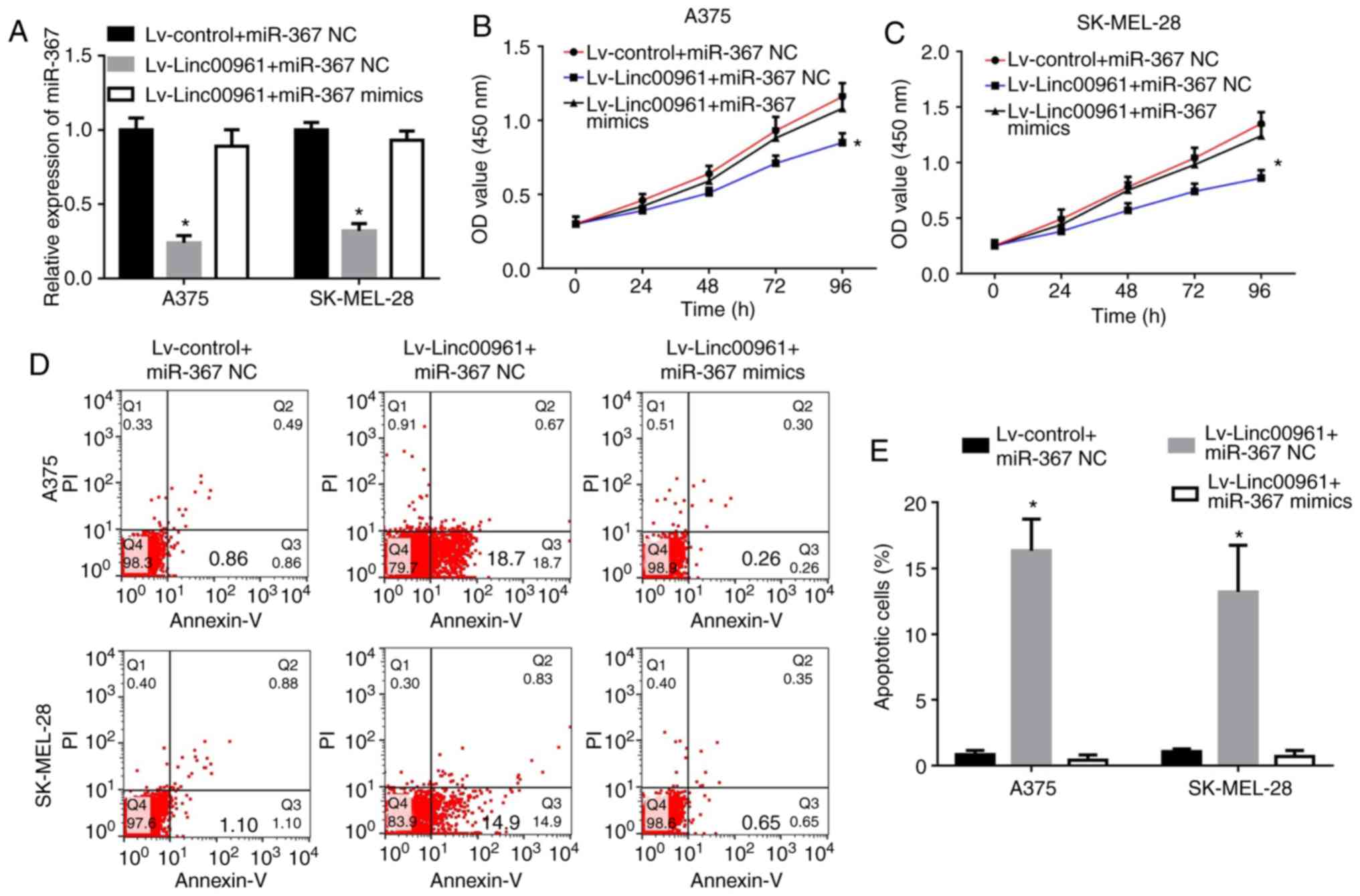 | Figure 7Upregulation of miR-367 reverses the
effects of Linc00961 on the proliferation and apoptosis of skin
melanoma cells. (A) Expression of miR-367 in A375 and SK-MEL-28
cells transfected with Lv-control + miR-367 NC, Lv-Linc00961 +
miR-367 NC or Lv-Linc00961 + miR-367 mimics, as determined via
reverse transcription-quantitative PCR. Proliferation of (B) A375
and (C) SK-MEL-28 cells transfected with Lv-control + miR-367 NC,
Lv-Linc00961 + miR-367 NC or Lv-Linc00961 + miR-367 mimics, as
determined by a Cell Counting Kit-8 assay. (D) Apoptosis of A375
and SK-MEL-28 cells transfected with Lv-control + miR-367 NC,
Lv-Linc00961 + miR-367 NC or Lv-Linc00961 + miR-367 mimics, as
determined via flow cytometry. (E) Statistical analysis of early
apoptotic rates of A375 and SK-MEL-228 cells transfected with
Lv-control + miR-367 NC, Lv-Linc00961 + miR-367 NC or Lv-Linc00961
+ miR-367 mimics. Data from three experiments are presented as the
mean ± SD. *P<0.05 vs. Lv-control + miR-367 NC.
Linc00961, long intergenic noncoding RNA 00961; Lv, lentivirus;
miR, microRNA; NC, negative control; PI, propidium iodide. |
Silencing PTEN ameliorates the inhibitory
effects of Linc00961 on proliferation and invasion of SM cells
Previous studies had indicated that PTEN was a
direct target of miR-367 in SM cells (21,22).
Thus, it was hypothesized that the inhibitory effects of Linc00961
in SM may be mediated via PTEN. To investigate this, the effects of
Linc00961 on the mRNA and protein expression levels of PTEN were
examined, and both were revealed to be upregulated as a consequence
of Linc00961 overexpression, although the mRNA and protein
expression levels were subsequently decreased following
co-transfection of miR-367 mimics (Fig. 9A and B). In functional terms, the
proliferation, migration and invasion of A375 and SK-MEL-28 cells
infected with Lv-Linc000961 were restored following the silencing
of PTEN (Fig. S1C); the effects
on proliferation are presented in Fig.
9C and D, whereas those on migration and invasion are presented
in Figs. S3, and 9F and G. Furthermore, PTEN knockdown
rescued the increase in apoptosis induced by Linc00961
overexpression in A375 and SK-MEL-28 cells (Figs. 9E and S4). Collectively, these results
indicated that Linc00961 inhibited the proliferation and invasion
of A375 and SK-MEL-28 cells by targeting the miR-367/PTEN axis.
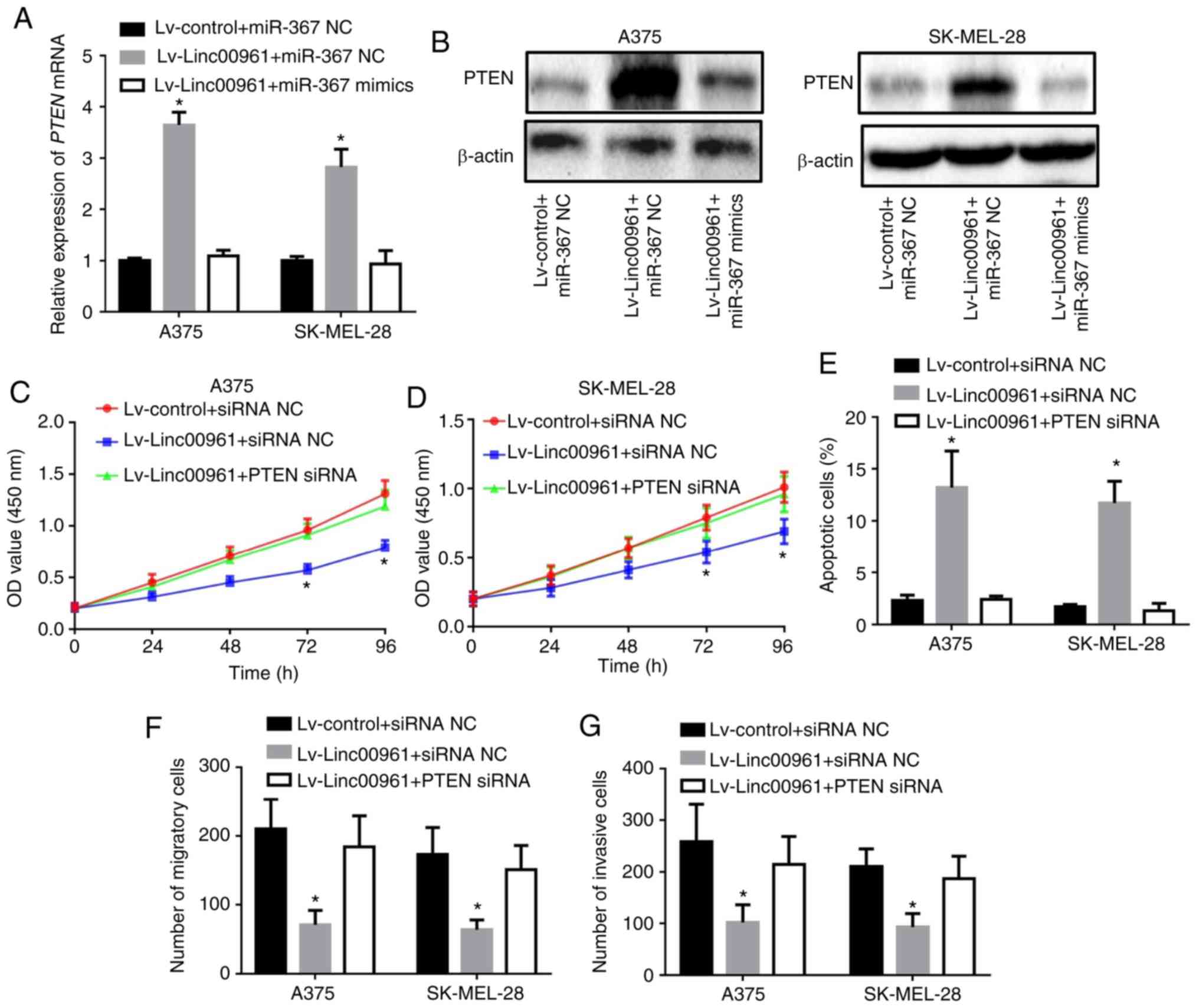 | Figure 9Silencing PTEN reverses the
inhibitory effects of Linc00961 on the proliferation and invasion
of SM cells. (A) mRNA and (B) protein expression of PTEN in A375
and SK MEL-28 cells transfected with Lv-control + miR-367 NC,
Lv-Linc00961 + miR-367 NC or Lv-Linc00961 + miR-367 mimics.
Proliferation of (C) A375 and (D) SK-MEL-28 cells transfected with
Lv-control + siRNA NC, Lv-Linc00961 + siRNA NC or Lv-Linc00961 +
PTEN siRNA, as determined by a Cell Counting Kit-8 assay. (E)
Apoptosis of A375 and SK-MEL-28 cells transfected with Lv-control +
siRNA NC, Lv-Linc00961 + siRNA NC or Lv-Linc00961 + PTEN siRNA, as
determined via flow cytometry. Numbers of (F) migratory and (G)
invasive A375 and SK-MEL-28 cells transfected with Lv-control +
siRNA NC, Lv-Linc00961 + siRNA NC, or Lv-Linc00961 + PTEN siRNA, as
determined via Transwell assays. Data from three experiments are
presented as mean ± SD. *P<0.05 vs. Lv-control +
miR-367 NC or siRNA NC. Linc00961, long intergenic noncoding RNA
00961; PTEN, phosphate and tension homology deleted on chromosome
10; Lv, lentivirus; miR, microRNA; siRNA, small interfering RNA;
NC, negative control. |
Discussion
LncRNAs have been identified as crucial regulators
and biomarkers in SM (13,23). In the present study, it was
revealed that Linc00961 was upregulated in SM tissues and cells
compared with BN tissues and human epidermal melanocytes. In
addition, Linc00961 expression was demonstrated to be positively
associated with the 5-year overall survival rate of patients with
SM by analyzing TCGA data; however, in the present study, as only
13 SM tissues were collected, the sample size was too small to
analyze associations between Linc00961 expression and the
clinicopathological features of melanoma patients. Additionally,
Linc00961 overexpression inhibited the proliferation, migration and
invasion, and promote the apoptosis of SM cells in vitro.
Furthermore, to the best of our knowledge, the first evidence was
provided that Linc00961 is able to sponge miR-367 and upregulate
the expression of PTEN, a direct target of miR-367 in SM (21,22).
Additionally, restoration of miR-367 or silencing of PTEN
expression reversed the inhibitory effects of Linc00961
overexpression on SM cells. Collectively, these results indicated
that Linc00961 may function as a ceRNA to inhibit cell
proliferation and invasion in SM cells by targeting the
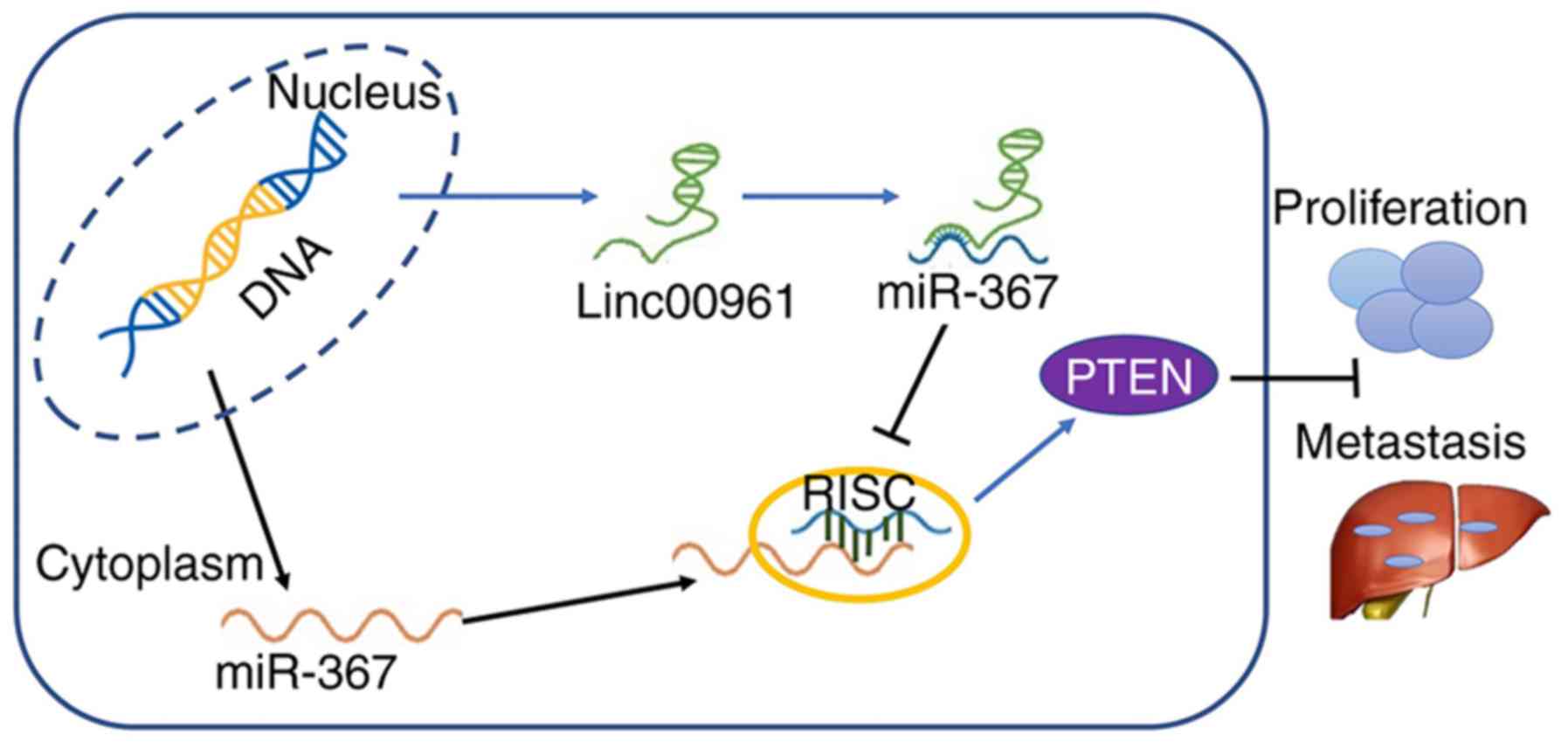
miR-367/PTEN axis (Fig. 10).
A recently published study reported that Linc00961
encoded a novel polypeptide termed SPAAR, which is involved in mTOR
complex 1 activation and muscle regeneration (14,24).
Additionally, the expression profile and potential function of
Linc00961 in genesis and progression of human cancers was
preliminarily established; Linc00961 was shown to suppress the
invasion and migration of NSCLC cells in vitro, and
metastasis in vivo (25).
Furthermore, Linc00961 was also reported to inhibit cell
proliferation and promote cell apoptosis by upregulating the levels
of proliferating cell nuclear antigen and suppressing the
expression of Bax protein in NSCLC (17). Furthermore, Linc00961 was also
downregulated in lung squamous cell carcinoma and demonstrated to
be a capable diagnostic biomarker (26). In glioma, Linc00961 was revealed to
be downregulated in tissues and cell lines, and its expression was
inversely associated with pathological features, poor overall
survival and unfavorable prognosis (16), indicating that Linc00961 functions
as a tumor suppressor in glioma. In renal cell carcinoma (RCC), it
was revealed that Linc00961 suppressed RCC progression by
inhibiting the epithelial-mesenchymal transition signaling pathway
(27). Consistent with these
previous studies, the present study also demonstrated that
Linc00961 functioned as a tumor suppressor gene in the
carcinogenesis and development of SM. Linc00961 was demonstrated to
inhibit the proliferation, and suppress the tumor growth of SM
in vitro and in vivo. It also induced inhibitory
effects on the migration and invasion of SM cells; however, the
underlying mechanism of Linc00961's suppressive role in SM
development is yet to be fully determined.
An increasing body of evidence has revealed that
lncRNAs are able to function as ceRNAs to interfere with miRNAs and
their downstream pathways. CeRNA regulatory networks have been
demonstrated to be key regulators in autoimmune diseases and viral
infections (28-30), and they are also commonly
associated with carcinogenesis and the progression of different
types of cancer (31-33), including gastric cancer and SM
(34). For example, LncRNA HOTAIR
promoted the tumorigenesis and progression of SM via the
miR-152-3p/c-MET/PI3K/AKT pathway (35). LncRNA activated by TGF-β, acting as
a ceRNA against miR-590-5p, promoted the proliferation and invasion
of SM cells (19). In the present
study, miR-367 was identified as a potential targeting miRNA that
possessed putative binding sites for Linc00961. It was subsequently
shown that Linc00961 could directly sponge and bind to miR-367 at
special recognition sites. Thus, the role of miR-367 in Linc00961's
suppressive role in SM was then investigated.
miR-367 has been revealed to be involved in the
carcinogenesis and progression in multiple types of cancer,
including NSCLC and glioma (36,37).
It is associated with the role of oncogenic or tumor-suppressive
genes, depending on the type of cancer involved. For example,
miR-367 promoted tumor growth in NSCLC, was associated with an
unfavorable prognosis, and stimulated Wnt cascade activation by
directly targeting the gene, F-box and WD repeat domain containing
7 (36). Conversely, upregulation
of miR-367 was associated with a favorable prognosis for patients
with high-grade glioma (37). In
the case of melanoma, miR-367 was shown to promote the
proliferation and invasion of cutaneous SM (21) or uveal melanoma (22) by downregulating PTEN expression. In
the present study, it was demonstrated that the suppressive effects
of Linc00961 on malignant phenotypes of SM cells were rescued by
miR-367 overexpression or PTEN knockdown. Therefore, combining the
results observed in previous studies with our present findings
supports the hypothesis that Linc00961 functions as a ceRNA to
inhibit the proliferation and invasion of SM by targeting the
miR-367/PTEN axis.
In conclusion, it was revealed that Linc00961 is a
potential tumor suppressor in SM. Linc00961 could inhibit cell
proliferation, and promote the apoptosis of SM cells in vivo
and in vitro. Furthermore, Linc00961 also suppressed the
migration and invasion of A375 and SK-MEL-28 cells. In
investigating the underlying mechanism, the present study also
demonstrated that Linc00961 functioned as a sponge of miR-367 to
enhance PTEN expression, and that miR-367 overexpression or PTEN
knockdown rescued the inhibitory effects of Linc00961
overexpression on A375 and SK-MEL-28 cells. The present study has
provided novel insight into the regulatory mechanisms of Linc00961
and the miR-367/PTEN axis in SM, and may provide a novel target for
the treatment of SM in the future.
Supplementary Materials
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81802935), Natural Science
Foundation of Shaanxi Provincial (grant no. 2019JM-487) and
Foundation of the First Affiliated Hospital of Xi'an Jiaotong
University (grant no. 2018MS-04).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contribution
LJW conceived and designed the experiments. XM, KHM,
and RG performed the majority of the experiments. YZ and DH were
involved in animal experiments, data analysis and compilation. LJW
drafted the manuscript. All authors have read and approved the
manuscript, and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Care and Use Committee of Xi'an Jiaotong University and performed
in accordance with institutional guidelines. The human study was
performed in accordance with the principles of the Declaration of
Helsinki and approved by the Ethical Committee of the First
Affiliated Hospital of Xi'an Jiaotong University. All the patients
enrolled in the present study provided their informed consent for
the use of their resected tissues in this research.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
We would like to express our gratitude to Dr
Shengjia Shi (Department of Urology, Xijing Hospital, Airforce
Military Medical University, Xi'an, China) for the generous
assistance with the RNA pull-down and luciferase assays.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhou J, Jin B, Jin Y, Liu Y and Pan J: The
antihelminthic drug niclosamide effectively inhibits the malignant
phenotypes of uveal melanoma in vitro and in vivo. Theranostics.
7:1447–1462. 2017. View Article : Google Scholar :
|
|
3
|
Rozeman EA, Dekker TJA, Haanen J and Blank
CU: Advanced melanoma: Current treatment options, biomarkers, and
future perspectives. Am J Clin Dermatol. 19:303–317. 2018.
View Article : Google Scholar
|
|
4
|
Guo W, Wang H, Yang Y, Guo S, Zhang W, Liu
Y, Yi X, Ma J, Zhao T, Liu L, et al: Down-regulated miR-23a
contributes to the metastasis of cutaneous melanoma by promoting
autophagy. Theranostics. 7:2231–2249. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang Y: The novel regulatory role of
lncRNA-miRNA-mRNA axis in cardiovascular diseases. J Cell Mol Med.
22:5768–5775. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yu B and Wang S: Angio-LncRs: LncRNAs that
regulate angio-genesis and vascular disease. Theranostics.
8:3654–3675. 2018. View Article : Google Scholar :
|
|
7
|
Krause HM: New and prospective roles for
lncRNAS in organelle formation and function. Trends Genet.
34:736–745. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Balas SM and Johnson AM: Exploring the
mechanisms behind long noncoding RNAs and cancer. Noncoding RNA
Res. 3:108–117. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rafiee A, Riazi-Rad F, Havaskary M and
Nuri F: Long noncoding RNAs: Regulation, function and cancer.
Biotechnol Genet Eng Rev. 34:153–180. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Anastasiadou E, Faggioni A, Trivedi P and
Slack FJ: The nefarious nexus of noncoding RNAs in cancer. Int J
Mol Sci. 19:pii. E20722018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu X, Zheng H, Tse G, Zhang L and Wu WKK:
CASC2: An emerging tumour-suppressing long noncoding RNA in human
cancers and melanoma. Cell Prolif. 51:e125062018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu X, Zheng H, Tse G, Chan MT and Wu WK:
Long non-coding RNAs in melanoma. Cell Prolif. 51:e124572018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Richtig G, Ehall B, Richtig E,
Aigelsreiter A, Gutschner T and Pichler M: Function and clinical
implications of long non-coding RNAs in melanoma. Int J Mol Sci.
18:pii. E7152017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Matsumoto A, Pasut A, Matsumoto M,
Yamashita R, Fung J, Monteleone E, Saghatelian A, Nakayama KI,
Clohessy JG and Pandolfi PP: mTORC1 and muscle regeneration are
regulated by the LINC00961-encoded SPAR polypeptide. Nature.
541:228–232. 2017. View Article : Google Scholar
|
|
15
|
Tajbakhsh S: lncRNA-encoded polypeptide
SPAR(s) with mTORC1 to regulate skeletal muscle regeneration. Cell
Stem Cell. 20:428–430. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lu XW, Xu N, Zheng YG, Li QX and Shi JS:
Increased expression of long noncoding RNA LINC00961 suppresses
glioma metastasis and correlates with favorable prognosis. Eur Rev
Med Pharmacol Sci. 22:4917–4924. 2018.PubMed/NCBI
|
|
17
|
Huang Z, Lei W, Tan J and Hu HB: Long
noncoding RNA LINC00961 inhibits cell proliferation and induces
cell apoptosis in human non-small cell lung cancer. J Cell Biochem.
119:9072–9080. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kunz M, Löffler-Wirth H, Dannemann M,
Willscher E, Doose G, Kelso J, Kottek T, Nickel B, Hopp L,
Landsberg J, et al: RNA-seq analysis identifies different
transcriptomic types and developmental trajectories of primary
melanomas. Oncogene. 37:6136–6151. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mou K, Liu B, Ding M, Mu X, Han D, Zhou Y
and Wang LJ: lncRNA-ATB functions as a competing endogenous RNA to
promote YAP1 by sponging miR-590-5p in malignant melanoma. Int J
Oncol. 53:1094–1104. 2018.PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Long J, Luo J and Yin X: miR-367 enhances
the proliferation and invasion of cutaneous malignant melanoma by
regulating phosphatase and tensin homolog expression. Mol Med Rep.
17:6526–6532. 2018.PubMed/NCBI
|
|
22
|
Ling JW, Lu PR, Zhang YB, Jiang S and
Zhang ZC: miR-367 promotes uveal melanoma cell proliferation and
migration by regulating PTEN. Genet Mol Res. 16:gmr160390672017.
View Article : Google Scholar
|
|
23
|
Liu XF, Hao JL, Xie T, Pant OP, Lu CB, Lu
CW and Zhou DD: The BRAF activated non-coding RNA: A pivotal long
non-coding RNA in human malignancies. Cell Prolif. 51:e124492018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rion N and Rüegg MA: LncRNA-encoded
peptides: More than translational noise? Cell Res. 27:604–605.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang B, Liu J, Zhang YH, Shen D, Liu S,
Lin F, Su J, Lin QF, Yan S, Li Y, et al: Long noncoding RNA
LINC00961 inhibits cell invasion and metastasis in human non-small
cell lung cancer. Biomed Pharmacother. 97:1311–1318. 2018.
View Article : Google Scholar
|
|
26
|
Chen WJ, Tang RX, He RQ, Li DY, Liang L,
Zeng JH, Hu XH, Ma J, Li SK and Chen G: Clinical roles of the
aberrantly expressed lncRNAs in lung squamous cell carcinoma: A
study based on RNA-sequencing and microarray data mining.
Oncotarget. 8:61282–61304. 2017.PubMed/NCBI
|
|
27
|
Chen D, Zhu M, Su H, Chen J, Xu X and Cao
C: LINC00961 restrains cancer progression via modulating
epithelial-mesenchymal transition in renal cell carcinoma. J Cell
Physiol. 234:7257–7265. 2019. View Article : Google Scholar
|
|
28
|
Li LJ, Zhao W, Tao SS, Leng RX, Fan YG,
Pan HF and Ye DQ: Competitive endogenous RNA network: Potential
implication for systemic lupus erythematosus. Expert Opin Ther
Targets. 21:639–648. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bossi L and Figueroa-Bossi N: Competing
endogenous RNAs: A target-centric view of small RNA regulation in
bacteria. Nat Rev Microbiol. 14:775–784. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cai Y and Wan J: Competing endogenous RNA
regulations in neurodegenerative disorders: Current challenges and
emerging insights. Front Mol Neurosci. 11:3702018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang C, Wu D, Gao L, Liu X, Jin Y, Wang D,
Wang T and Li X: Competing endogenous RNA networks in human cancer:
Hypothesis, validation, and perspectives. Oncotarget.
7:13479–13490. 2016.PubMed/NCBI
|
|
32
|
Qu J, Li M, Zhong W and Hu C: Competing
endogenous RNA in cancer: A new pattern of gene expression
regulation. Int J Clin Exp Med. 8:17110–17116. 2015.
|
|
33
|
Guo LL, Song CH, Wang P, Dai LP, Zhang JY
and Wang KJ: Competing endogenous RNA networks and gastric cancer.
World J Gastroenterol. 21:11680–11687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sarkar D, Leung EY, Baguley BC, Finlay GJ
and Askarian-Amiri ME: Epigenetic regulation in human melanoma:
Past and future. Epigenetics. 10:103–121. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Luan W, Li R, Liu L, Ni X, Shi Y, Xia Y,
Wang J, Lu F and Xu B: Long non-coding RNA HOTAIR acts as a
competing endogenous RNA to promote malignant melanoma progression
by sponging miR-152-3p. Oncotarget. 8:85401–85414. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xiao G, Zhang B, Meng J, Wang J, Xu C,
Tang SC, Li X, Zhang J, Liang R, Ren H and Sun X: miR-367
stimulates Wnt cascade activation through degrading FBXW7 in NSCLC
stem cells. Cell Cycle. 16:2374–2385. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Guan Y, Chen L, Bao Y, Qiu B, Pang C, Cui
R and Wang Y: High miR-196a and low miR-367 cooperatively correlate
with unfavorable prognosis of high-grade glioma. Int J Clin Exp
Pathol. 8:6576–6588. 2015.PubMed/NCBI
|