1. Introduction
Muscle is an important part of the animal body and
skeletal muscle constitutes ~40% of total body weight. The
principal functions of skeletal muscle include maintaining body
structure and posture, controlling motor movement and storing
energy (1). Rhabdomyosarcoma (RMS)
is a skeletal muscle-derived sarcoma occurring predominantly in
children and young adults (2).
There are two main subtypes of RMS: Embryonal RMS (eRMS) and
alveolar RMS (aRMS). eRMS occurs more often in children <10
years old, whereas aRMS prototypically occurs in adolescents in 30%
of RMS cases with a poorer prognosis and a higher rate of
metastasis (3). In addition, aRMS
exhibits typical chromosomal translocations between chromosomes 2
and 13 [t (2;13)(q35;q14)] or chromosomes 1 and 13 [t (1;13)
(q36;q14)], which lead to the production of two fusion genes:
paired box (PAX)3/forkhead box protein O1 (FOXO1) and PAX7/FOXO1,
respectively (4). Furthermore,
although RMS tumors commonly form from within skeletal muscle, they
can also originate from non-muscle sites, including the skull base,
genitourinary tract, biliary tree and salivary glands (5,6).
Previously, microRNAs (miRNAs), a novel class of
small non-coding RNAs, have been demonstrated to act as key
regulators of skeletal muscle cell fate determination and to be
dysregulated in aRMS and eRMS (7).
miRNAs are single-strand RNAs of ~22 nucleotides in length, which
negatively regulate gene expression at the post-transcriptional
level by complementary binding to the 3′ untranslated regions of
target genes and result in mRNA degradation or translation
inhibition (8). To date, emerging
evidence has demonstrated that miRNAs are critical in a
considerable number of physiological and pathological processes,
including proliferation, differentiation, chemoresistance and
tumorigenesis (9–12). It was also reported that
overexpression of selected ‘tumor suppressor’ miRNAs by gain-of
function studies impaired the tumorigenic behavior of RMS cells
(13). In addition, miRNA
expression profiling has been demonstrated to be a promising
approach to discriminate specific variants among RMS subtypes and
further provide useful prognostic information concerning the
alveolar and embryonal forms of RMS (14,15).
These studies suggest that miRNA dysregulation may be involved in
the pathogenesis of RMS.
The present review aimed to evaluate our current
understanding of the regulation of miRNAs in skeletal muscle
development and their deregulation in RMS. Additionally, the
possible therapeutic application and challenges of miRNAs in
clinical practice were discussed.
2. Process of skeletal muscle
development
The skeletal muscle system of vertebrates originates
through a complex and multi-stage process termed myogenesis where
numerous genes are co-operatively involved in the regulation of
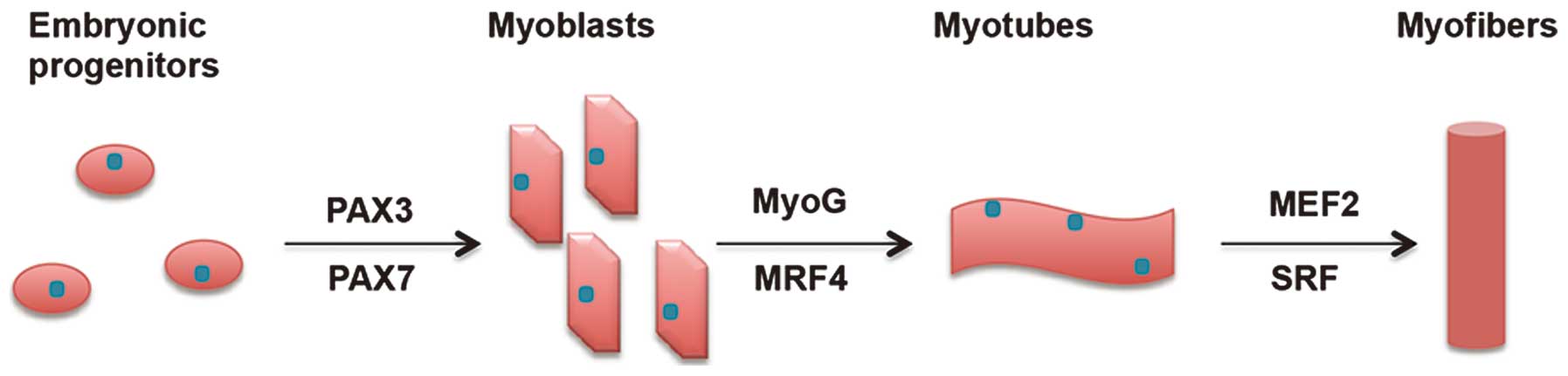
each stage (Fig. 1). This process
begins in the somites of the embryo, which differentiate into
dermomyotome-containing myogenic precursors at the first stage.
Following commitment to a myogenic cell lineage, the myogenic
precursors proliferate and differentiate into myoblasts, followed
by differentiation into myotubes and finally differentiate into
myofibers (16).
The regulatory network leading to the process of
muscle development has been ascribed to a specific class of
transcription factors termed myogenic regulatory factors (MRFs)
(17). The expression of MRFs is
limited to the muscle lineage and results in the activation of a
cascade of events leading to the formation of mature muscle
fibers.
The upstream regulators of early MRFs are
paired-domain- and homeobox-containing proteins, including Pax3 and
Pax7, which are active in embryo-genesis. As myoblasts migrate,
myogenin (MyoG) and MRF4 are expressed and trigger myoblasts to
differentiate into myotubes. Following the terminal differentiation
of myotubes, they act in concert with other factors, including
myocyte enhancer factor 2 (MEF2) and serum response factor (SRF) to
activate genes responsible for muscle fiber architecture and
functionality (18). Besides these
intrinsic signaling pathways, the differentiating muscle cells are
regulated by external stimuli, including transforming growth factor
(TGF)-β or Wnt signaling (19). In
addition, the majority of the aforementioned processes can also be
modulated at the post-transcriptional level by miRNAs, which are
demonstrated to be irreplaceable in skeletal muscle
development.
3. Expression patterns of miRNAs in skeletal
muscle development
During skeletal muscle development, certain miRNAs
are specifically enriched in skeletal muscle cells and others are
differentially expressed in the development process. The temporal
or tissue specific expression patterns of miRNAs have been
determined by miRNA array or high-throughput sequencing approaches
in previous years(20). In one
study, 77 miRNAs were found to be upregulated and 68 miRNAs were
downregulated by microarray in C2C12 myoblast cells, which were
induced to differentiate in horse serum (20). Among the 77 upregulated miRNAs,
miR-133a-1, miR-133a-2, miR-133b and miR-206 were the most
significantly upregulated. Their critical role in skeletal muscle
differentiation was also confirmed by other studies (21-24).
Several other miRNAs, including miR-9-2, miR-122a, miR-703 and
miR-805, were most significantly downregulated, however, few of
them were found to be involved in the differentiation process.
Certain miRNAs, including miR-699a, were downregulated during
skeletal muscle differentiation. These observations demonstrated
that miRNAs were differentially expressed in the process of
skeletal muscle development.
4. Muscle-specific miRNAs in myogenesis
Numerous miRNAs can be highly and specifically
enriched in certain tissues. The miRNAs that are specifically
expressed in skeletal muscle are referred to as myomiRs, which
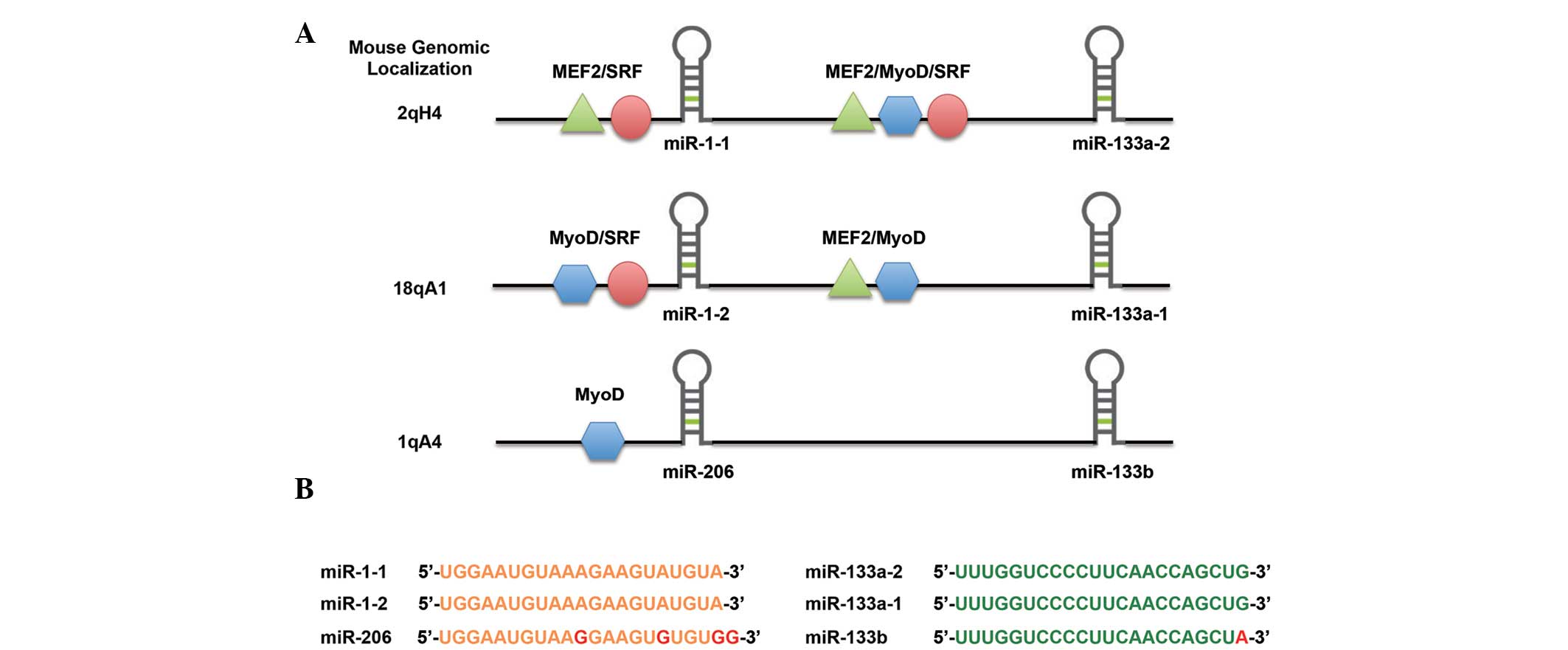
include the miR-1/206 cluster (21,25).
A complete list of myomiRs is provided in Table I and their function is further
described in corresponding paragraphs. The miR-1/206 cluster is
composed of six distinct miRNAs located on three separate
chromosomes in three bicistronic transcripts. miR-1-2 and
miR-133a-1 are located on chromosome 18, miR-1-1 and miR-133a-2 are
located on chromosome 20 and miR-133b along with miR-206 are
located on chromosome 6 (Fig. 2).
In terms of architecture, miR-1-1 and miR-1-2 are identical and
differ from miR-206 by four nucleotides. miR-133a-1 and miR-133a-2
are identical and differ from miR-133b by one nucleotide. During
myogenesis, the myogenic transcription factors MyoD, MEF2 and SRF
directly regulate the expression of miR-1 and miR-133a in skeletal
muscle whereas the expression of miR-206 is controlled by MyoD and
MyoG (26,27).
 | Table IMuscle-specific miRNAs involved in
myogenesis and RMS. |
Table I
Muscle-specific miRNAs involved in
myogenesis and RMS.
| miRNA | Target gene | Function | Reference |
|---|
| miR-1 | HDAC4 | Promotes myoblast
differentiation | 20 |
| Hand2 | Inhibits
cardiomyocyte proliferation | 25,27 |
| PAX3 | Inhibits RMS cell
proliferation | 41 |
| PAX7 | Promotes muscle
cell differentiation | 29 |
| CCND2 | Inhibits RMS cell
proliferation | 41 |
| cMet | Inhibits RMS
development | 40 |
| miR-133 | SRF | Promotes myoblast
proliferation | 20 |
| miR-206 | PAX7 | Inhibits muscle
cell proliferation | 29 |
| PAX3 | Inhibits RMS cell
proliferation | 41 |
| CCND2 | Inhibits RMS cell
proliferation | 41 |
| cMet | Inhibits RMS
development | 40 |
| Cx43 | Promotes myoblast
differentiation | 28 |
| Polα1 | Inhibits muscle
cell proliferation | 21 |
| Fstl1 | Inhibits muscle
cell proliferation | 30 |
| Utrn | Inhibits muscle
cell proliferation | 30 |
The function of the muscle-specific miRNAs in
myogenesis has been examined in detail. It was reported that miR-1
and miR-133a regulate skeletal muscle cell proliferation and
differentiation by targeting histone deacetylase 4 (HDAC4) and SRF,
respectively, thus establishing a negative-feedback loop for
myocyte differentiation (21).
Furthermore, the injection of miR-1 into embryonic cardiomyocytes
of mice led to decreased proliferation of cardiomyocytes, which was
ascribed to the decreased expression of Hand2, a transcription
factor that promotes cardiomyocyte proliferation (26). Consistently, a significant increase
in Hand2 expression and proliferating cardiomyocytes was observed
in an miR-1-2 deficient mouse model (28).
By contrast, miR-133a promotes myoblast
proliferation partly by repressing the expression of SRF, a
critical regulator of muscle cell differentiation (21). The genetic interaction between
miR-133a and SRF results in the upregulation of miR-133a by SRF
leading to the further repression of SRF, thereby constituting a
negative feedback loop. Paradoxically, miR-133a and miR-1 exhibit
opposing effects on skeletal muscle development although they
derive from the same miRNA polycistronic transcript. The primary
function of miR-133a is to promote proliferation and inhibit
differentiation, while the function of miR-1 is to induce the
differentiation of mesodermal progenitors to the muscle lineages.
It has been demonstrated that miR-1 and miR-133 have a specific
role in muscle cell proliferation and differentiation in an
antagonistic manner, with the balance being altered one way or the
other by additional modulators of gene expression (29).
Similar to miR-1 in skeletal muscle, miR-206 has
been demonstrated to promote myoblast differentiation by repressing
the expression of connexin 43 (Cx43), thereby decreasing the
electrical coupling between myofibers via gap junctions, which
inhibits the terminal differentiation of skeletal muscle cells
(30). In addition, miR-206 was
reported to repress the expression of the p180 subunit of DNA
polymerase α1 (22), Pax7
(31), follistatin-like 1
(32) or utrophin (32), thereby suppressing muscle cell
proliferation through inhibiting DNA synthesis.
5. Non-muscle-specific miRNAs in
myogenesis
In addition to muscle-specific miRNAs, numerous
non-muscle-specific miRNAs, referred to as non-myomiRs, are also
important in the regulation of myogenesis (Table II). It has been demonstrated that
these non-myomiRs regulate muscle proliferation and differentiation
through the repression of target genes through multiple processes
(33–39). At the onset of myogenesis, miR-27b
induced improper migration and early differentiation of myoblasts
by targeting the Pax3 protein (33). miR-26a (34) and miR-214 (35,36)
also promote myogenesis by targeting enhancer of zeste 2, another
known inhibitor of myogenesis. It is noted that the timing of
expression of miR-26a and miR-214 differs during myogenesis. Once
muscle differentiation begins, miR-214 is upregulated via
MyoD/MyoG, which promote P21Cip1 and myogenin
expression, while miR-26a increases gradually during the course of
myogenesis.
| Table IINon-muscle-specific miRNAs involved
in myogenesis and RMS. |
Table II
Non-muscle-specific miRNAs involved
in myogenesis and RMS.
| miRNA | Target gene | Function | Reference |
|---|
| miR-27b | PAX3 | Promotes myoblast
differentiation | 31 |
| miR-26a | Ezh2 | Promotes myoblast
differentiation | 32 |
| miR-214 | Ezh2 | Promotes myoblast
differentiation | 33,34 |
| miR-181 | HOX11 | Promotes muscle
cell differentiation | 35 |
| miR-669a | MyoD | Inhibits skeletal
muscle differentiation | 36 |
| miR-669q | | | |
| miR-29 | YY1 | Promotes myoblast
differentiation | 42 |
| PAX3 | Inhibits RMS cell
proliferation | 41 |
| CCND2 | Inhibits RMS cell
proliferation | 41 |
| miR-183 | PTEN | Promotes RMS cell
migration | 43 |
| EGR1 | Promotes RMS cell
migration | 43 |
| miR-203 | P63 | Inhibits RMS cell
proliferation | 44 |
|
miR-9a | E-cadherin | Inhibits RMS cell
migration | 45 |
| miR-450b | TGF-β1 | Inhibits RMS
development | 46 |
Inhibition of homeobox-protein A11 (HOX11) by
miR-181 is another step in muscle differentiation (37). The low expression of HOX11 leads to
increases in MyoD, a target of HOX11, and proper differentiation in
muscle cells. It is consistent with the finding that miR-181 is
upregulated during muscle development whereas it is downregulated
in adult skeletal muscle (38). By
contrast, miR-669a and miR-669q are expressed in the heart muscle
to prevent skeletal muscle differentiation from the beginning
through inhibiting MyoD and its targets, thus ensuring that
skeletal muscle myogenesis occurs in the correct locations
(39).
Taken together, these observations are consistent
with differential expression profiles of miRNAs between myogenesis
and the adult skeletal muscle. The differentially expressed miRNAs
provide a molecular basis for proper regulation of muscle
development, which highlights the complexity of miRNA function.
6. Muscle-specific miRNAs in
rhabdomyosarcoma
RMS is predominantly a pediatric sarcoma that
resembles developing skeletal muscle and accounts for >50% of
soft tissues sarcomas in children (40). Evidence has demonstrated that
myomiRs were significantly downregulated in RMS, indicating a
critical role in the terminal differentiated phenotype of RMS cells
(41). In particular, Rao et
al (29) reported that
overexpression of miR-1 in the RMS cell line, RD, results in muscle
gene expression and cell cycle arrest, whereas miR-133a decreases
the expression of muscle markers. This is consistent with the
distinct roles of miR-1 and miR-133a in normal muscle
differentiation. However, miRNAs suppress cell growth in the RMS
cell line, indicating that cell context is important to the fate of
miRNA regulation.
A similar growth inhibitory effect has also been
confirmed by forced expression of either miR-1 or miR-206 in the
RMS cell line in vitro and in vivo (42,43).
The induction of miR-1/206 precursor led to decreased myogenic
differentiation in cell migration and inhibition of tumorigenic
potential. Furthermore, the results of mRNA profiling prior to and
following miR-206 transfection in RD18 cells revealed that >700
genes were modulated, including c-Met (43). The downregulation of c-Met by
miR-1/206 led to a significant inhibition of RMS development,
suggesting that the targeting of c-Met is one of the underlying
mechanisms responsible for RMS development.
The anti-tumor capacity of the ectopic expression of
the miR-1/206 cluster in RMS was further verified by the
observation that these miRNAs directly regulate the expression of
CCND2, a cell cycle gene (44).
Overexpression of miR-1/206 demonstrated a strong promyogenic
effect in RMS cells and downregulated the protein and transcript
levels of CCND2. Additionally, miR-1/206 significantly
downregulated PAX3 protein expression in the eRMS cell line, JR1,
however, demonstrated no effect on the protein levels of PAX3 in an
aRMS cell line, Rh30 (44). This
finding highlights, once more, the importance of cell context in
determining the response to miRNA modulation.
7. Non-muscle-specific miRNAs in
rhabdomyosarcoma
In addition to myomiRs, numerous non-myomiRs are
implicated in the regulation of RMS development. The deregulation
of miR-29 was reported in a small cohort of aRMS in which nuclear
factor-κB activation led to overexpression of Yin Yang 1, resulting
in sustained downregulation of miR-29b2/miR-29c and inhibition of
myogenesis (45). In addition,
decreased expression of miR-29, as well as miR-1/206, stabilized
the RMS phenotype by targeting PAX3 and CCND2 (44). These findings reiterate that the
RMS state is maintained by the deregulation of multiple miRNAs and
their target genes, supporting a tumor suppressor role for these
miRNAs.
Another miRNA linked to RMS is miR-183, which acts
as an onco-miR in several types of cancer, including RMS, synovial
sarcoma and colon cancer (46).
Knocking down of miR-183 by anti-miR-183 treatment in tumor cells
reduced cell migration in vitro and stimulated the
expression of the tumor suppressor gene phosphatase and tensin
homolog (PTEN), which in turn, promoted early growth response 1
(EGR1) expression, thus reinforcing the repression of cell
migration (46). These results
demonstrated that miR-183 has an oncogenic role through targeting
two tumor suppressor genes, EGR1 and PTEN, and the deregulation of
the fundamental miRNA regulatory network may be central to the
development of several other tumor types.
Additionally, certain other non-myomiRs were
described in the context of cell differentiation, migration or
metastasis in RMS. Re-expression of miR-203 in RMS cells inhibited
their proliferation and migration and promoted terminal myogenic
differentiation by directly targeting p63 (47). miR-9a is another miRNA
capable of inhibiting cell migration, which was found to directly
target E-cadherin and was expressed in higher levels in aRMS than
in eRMS, correlating with their metastatic potentials (48). In addition, in the two cultured
cells and tumor implants, the growth of RMS was significantly
arrested by miR-450b-5p, which was strictly regulated by TGF-β1
(49).
Taken together, these data demonstrate the critical
role of miRNAs in modulating target genes involved in one or more
cellular function/process and the complexity of miRNA regulation
and function in RMS development. In addition, these studies have
revealed that small gene expression alterations, even if only
occurring in one miRNA, may be affect the balance between
pathological and physiological cell fate programs.
8. miRNAs as novel therapeutic targets in
rhabdomyosarcoma
The widespread and crucial roles of miRNAs in RMS
development and progression raise interesting prospects for
exploiting miRNAs as novel therapeutic targets in RMS.
In this regard, various approaches that upregulate
or downregulate miRNAs have been employed to target miRNAs in RMS,
and demonstrate significant efficacy in the treatment of RMS
development following intravenous delivery in vivo (50). In particular, two pre-clinical
studies demonstrated that ectopic expression of miR-206 by
lentiviral vectors leads to cell cycle arrest and myogenic
differentiation of RMS cells, preventing xenograft growth in
vivo by inhibiting the expression of oncogenic c-Met (42,43).
In addition, knockdown of miR-183, an onco-miR in several types of
cancer, by antisense-based miRNA antagonists led to significant
decreases in tumor migration through directly promoting the
expression of EGR1, a regulator of cell migration (46).
Although the upregulation or downregulation of
selected miRNAs is a possible strategy for targeted therapy in RMS,
it must be noted that there remain several challenges regarding
miRNA-based therapy. Viral vectors, though efficient in the
overexpression of miRNA genes, are limited in their clinical
application by immunogenicity and non-specificity. Non-viral
cationic liposomes are attractive for mediating miRNA transfer,
however, their low efficiency in cell transfection also limit their
development. Certain types of nanoparticles have been proposed to
efficiently deliver miRNAs or anti-miRNAs to target tumor sites
(51), implying that they are
alternative tools for introducing miRNAs for the treatment of RMS.
However, additional preclinical data are required to demonstrate
their suitability for the clinic and efficacy in the application of
miRNA therapy. Their relevance and mode of action require further
investigation in genetic models of RMS that more accurately
recapitulate the onset and progression of aRMS and eRMS tumors.
9. Conclusion
miRNAs have emerged as critical regulators in
skeletal muscle development, regeneration and function. They are
also found to be dysregulated in skeletal muscle-associated
diseases, including RMS. Thus, miRNAs are promising biomarkers and
candidates for potential therapeutic intervention, and provide an
avenue to further dissect the mechanisms that may contribute to
genetic and acquired muscle disorders or other associated diseases.
Future studies are required to focus on the identification of
miRNAs involved in skeletal muscle development and on advancing
novel therapies that are able to modulate miRNA activity to treat
muscle-associated diseases.
Acknowledgments
This study was supported by the Project Funded by
the Priority Academic Program Development of Jiangsu Higher
Education Institutions (PAPD), Jiangsu Co-innovation Center for
Prevention and Control of Important Animal Infectious Diseases and
Zoonoses and the National Natural Science Foundation of China
(grant nos. 31101683 and 31272405).
References
|
1
|
Fong AP and Tapscott SJ: Skeletal muscle
programming and re-programming. Curr Opin Genet Dev. 23:568–573.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arndt CA, Rose PS, Folpe AL and Laack NN:
Common musculoskeletal tumors of childhood and adolescence. Mayo
Clin Proc. 87:475–487. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Perez EA, Kassira N, Cheung MC, Koniaris
LG, Neville HL and Sola JE: Rhabdomyosarcoma in children: a SEER
population based study. J Surg Res. 170:e243–e251. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Charytonowicz E, Cordon-Cardo C,
Matushansky I and Ziman M: Alveolar rhabdomyosarcoma: is the cell
of origin a mesenchymal stem cell? Cancer Lett. 279:126–136. 2009.
View Article : Google Scholar
|
|
5
|
Hatley ME, Tang W, Garcia MR, et al: A
mouse model of rhabdomyosarcoma originating from the adipocyte
lineage. Cancer Cell. 22:536–546. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Barr FG: Molecular genetics and
pathogenesis of rhabdomyosarcoma. J Pediatr Hematol Oncol.
19:483–491. 1997. View Article : Google Scholar
|
|
7
|
Novák J, Vinklárek J, Bienertová-Vašků J
and Slabý O: MicroRNAs involved in skeletal muscle development and
their roles in rhabdomyosarcoma pathogenesis. Pediatr Blood Cancer.
60:1739–1746. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang F, Fu XD, Zhou Y and Zhang Y:
Down-regulation of the cyclin E1 oncogene expression by
microRNA-16-1 induces cell cycle arrest in human cancer cells. BMB
Rep. 42:725–730. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang F, Niu G, Chen X and Cao F: Molecular
imaging of microRNAs. Eur J Nucl Med Mol Imaging. 38:1572–1579.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang F, Song X, Li X, et al: Noninvasive
visualization of microRNA-16 in the chemoresistance of gastric
cancer using a dual reporter gene imaging system. PLoS One.
8:e617922013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ciarapica R, Russo G, Verginelli F, et al:
Deregulated expression of miR-26a and Ezh2 in rhabdomyosarcoma.
Cell Cycle. 8:172–175. 2009. View Article : Google Scholar
|
|
14
|
Davicioni E, Anderson JR, Buckley JD,
Meyer WH and Triche TJ: Gene expression profiling for survival
prediction in pediatric rhabdomyosarcomas: a report from the
children’s oncology group. J Clin Oncol. 28:1240–1246. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gougelet A, Perez J, Pissaloux D, et al:
miRNA profiling: How to bypass the current difficulties in the
diagnosis and treatment of sarcomas. Sarcoma. 2011:4606502011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bentzinger CF, Wang YX and Rudnicki MA:
Building muscle: molecular regulation of myogenesis. Cold Spring
Harb Perspect Biol. 4:a0083422012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Buckingham M: Skeletal muscle formation in
vertebrates. Curr Opin Genet Dev. 11:440–448. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Berkes CA and Tapscott SJ: MyoD and the
transcriptional control of myogenesis. Semin Cell Dev Biol.
16:585–595. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stockdale FE: Mechanisms of formation of
muscle fiber types. Cell Struct Funct. 22:37–43. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu L, Zhou L, Chen EZ, et al: A novel
YY1-miR-1 regulatory circuit in skeletal myogenesis revealed by
genome-wide prediction of YY1-miRNA network. PLoS One.
7:e275962012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen JF, Mandel EM, Thomson JM, et al: The
role of microRNA-1 and microRNA-133 in skeletal muscle
proliferation and differentiation. Nat Genet. 38:228–233. 2006.
View Article : Google Scholar
|
|
22
|
Kim HK, Lee YS, Sivaprasad U, Malhotra A
and Dutta A: Muscle-specific microRNA miR-206 promotes muscle
differentiation. J Cell Biol. 174:677–687. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dey BK, Gagan J and Dutta A: miR-206 and
-486 induce myoblast differentiation by downregulating Pax7. Mol
Cell Biol. 31:203–214. 2011. View Article : Google Scholar :
|
|
24
|
Huang MB, Xu H, Xie SJ, Zhou H and Qu LH:
Insulin-like growth factor-1 receptor is regulated by microRNA-133
during skeletal myogenesis. PLoS One. 6:e291732011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
McCarthy JJ: MicroRNA-206: the skeletal
muscle-specific myomiR. Biochim Biophys Acta. 1779:682–691. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao Y, Samal E and Srivastava D: Serum
response factor regulates a muscle-specific microRNA that targets
Hand2 during cardiogenesis. Nature. 436:214–220. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sweetman D, Goljanek K, Rathjen T, et al:
Specific requirements of MRFs for the expression of muscle specific
microRNAs, miR-1, miR-206 and miR-133. Dev Biol. 321:491–499. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao Y, Ransom JF, Li A, et al:
Dysregulation of cardiogenesis, cardiac conduction and cell cycle
in mice lacking miRNA-1-2. Cell. 129:303–317. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rao PK, Missiaglia E, Shields L, et al:
Distinct roles for miR-1 and miR-133a in the proliferation and
differentiation of rhabdomyosarcoma cells. FASEB J. 24:3427–3437.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Anderson C, Catoe H and Werner R: MIR-206
regulates connexin43 expression during skeletal muscle development.
Nucleic Acids Res. 34:5863–5871. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen JF, Tao Y, Li J, et al: microRNA-1
and microRNA-206 regulate skeletal muscle satellite cell
proliferation and differentiation by repressing Pax7. J Cell Biol.
190:867–879. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rosenberg MI, Georges SA, Asawachaicharn
A, Analau E and Tapscott SJ: MyoD inhibits Fstl1 and Utrn
expression by inducing transcription of miR-206. J Cell Biol.
175:77–85. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Crist CG, Montarras D, Pallafacchina G, et
al: Muscle stem cell behavior is modified by microRNA-27 regulation
of Pax3 expression. Proc Natl Acad Sci USA. 106:13383–13387. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wong CF and Tellam RL: MicroRNA-26a
targets the histone methyltransferase Enhancer of Zeste homolog 2
during myogenesis. J Biol Chem. 283:9836–9843. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Feng Y, Cao JH, Li XY and Zhao SH:
Inhibition of miR-214 expression represses proliferation and
differentiation of C2C12 myoblasts. Cell Biochem Funct. 29:378–383.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu J, Luo XJ, Xiong AW, et al:
MicroRNA-214 promotes myogenic differentiation by facilitating exit
from mitosis via down-regulation of proto-oncogene N-ras. J Biol
Chem. 285:26599–26607. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Naguibneva I, Ameyar-Zazoua M, Polesskaya
A, et al: The microRNA miR-181 targets the homeobox protein Hox-A11
during mammalian myoblast differentiation. Nat Cell Biol.
8:278–284. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Naguibneva I, Polesskaya A, Ameyar-Zazoua
M, et al: Micro-RNAs and muscle differentiation. J Soc Biol.
201:367–376. 2007.in French. View Article : Google Scholar
|
|
39
|
Crippa S, Cassano M, Messina G, et al:
miR669a and miR669q prevent skeletal muscle differentiation in
postnatal cardiac progenitors. J Cell Biol. 193:1197–1212. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Slater O and Shipley J: Clinical relevance
of molecular genetics to paediatric sarcomas. J Clin Pathol.
60:1187–1194. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen JF, Callis TE and Wang DZ: microRNAs
and muscle disorders. J Cell Sci. 122(Pt 1): 13–20. 2009.
View Article : Google Scholar :
|
|
42
|
Taulli R, Bersani F, Foglizzo V, et al:
The muscle-specific microRNA miR-206 blocks human rhabdomyosarcoma
growth in xenotransplanted mice by promoting myogenic
differentiation. J Clin Invest. 119:2366–2378. 2009.PubMed/NCBI
|
|
43
|
Yan D, Dong Xda E, Chen X, et al:
MicroRNA-1/206 targets c-Met and inhibits rhabdomyosarcoma
development. J Biol Chem. 284:29596–29604. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li L, Sarver AL, Alamgir S and Subramanian
S: Downregulation of microRNAs miR-1, -206 and -29 stabilizes PAX3
and CCND2 expression in rhabdomyosarcoma. Lab Invest. 92:571–583.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang H, Garzon R, Sun H, et al:
NF-kappaB-YY1-miR-29 regulatory circuitry in skeletal myogenesis
and rhabdomyosarcoma. Cancer Cell. 14:369–381. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sarver AL, Li L and Subramanian S:
MicroRNA miR-183 functions as an oncogene by targeting the
transcription factor EGR1 and promoting tumor cell migration.
Cancer Res. 70:9570–9580. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Diao Y, Guo X, Jiang L, et al: miR-203, a
tumor suppressor frequently down-regulated by promoter
hypermethylation in rhabdomyosarcoma. J Biol Chem. 289:529–539.
2014. View Article : Google Scholar :
|
|
48
|
Armeanu-Ebinger S, Herrmann D, Bonin M, et
al: Differential expression of miRNAs in rhabdomyosarcoma and
malignant rhabdoid tumor. Exp Cell Res. 318:2567–2577. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sun MM, Li JF, Guo LL, et al: TGF-beta1
suppression of microRNA-450b-5p expression: a novel mechanism for
blocking myogenic differentiation of rhabdomyosarcoma. Oncogene.
33:2075–2086. 2014. View Article : Google Scholar
|
|
50
|
Goljanek-Whysall K, Sweetman D and
Münsterberg AE: microRNAs in skeletal muscle differentiation and
disease. Clin Sci (Lond). 123:611–625. 2012. View Article : Google Scholar
|
|
51
|
Muthiah M, Park IK and Cho CS:
Nanoparticle-mediated delivery of therapeutic genes: focus on miRNA
therapeutics. Expert Opin Drug Deliv. 10:1259–1273. 2013.
View Article : Google Scholar : PubMed/NCBI
|