Introduction
Non-melanoma skin carcinoma is one of the most
frequent types of tumor worldwide and includes two histological
types: Basal cell carcinoma and squamous cell carcinoma (SCC)
(1–3). Cutaneous SCC is the second most
common cutaneous carcinoma and accounts for 20% of all types of
cutaneous tumor (4–6). Previous studies have made significant
progress in understanding the mechanisms underlying the
tumorigenesis and development of cutaneous SCC (7–12).
However, the incidence rates of patients with cutaneous SCC have
been increasing rapidly each year, due to the lack of molecular
targeted therapies and reliable molecular markers in early
diagnosis. Therefore, novel molecular biomarkers and targeted
therapeutic approaches for the diagnosis and treatment of patients
with cutaneous SCC are urgently required.
Hath1, also termed Math1, Atoh1 and Cath1 in mouse,
drosophila and chicken, respectively, is a basic helix-loop-helix
(bHLH) transcription factor (13).
Hath1 was initially identified as a tumor suppressor gene in colon
cancer (14). However, increasing
evidence has revealed that Hath1 is not only a tumor suppressor in
colon cancer (14–24), but is also downregulated and
involved in the tumorigenesis of various other types of human
cancer, including medulloblastoma (25,26),
gastric carcinoma (27,28), intestinal neuroendocrine tumors
(29), lung cancer (30,31)
and Merkel cell carcinoma (32).
Inherited or acquired defective mutants of Atoh were initially
identified in patients with Merkel cell carcinoma by Leonard in
2002 (32). Deletion of Hath1
using retrovirus vectors led to the disruption of Sonic hedgehog
signaling in the developing cerebellum and finally resulted in
medulloblastoma formation (26).
Bossuyt et al confirmed that the incidence of colonic polyps
in mice lacking the Hath1 gene was 100% compared with that in wild
type mice (16). Zhu et al
found that Hath1 expression is down-regulated in non-mucinous
adenocarcinomas (19–21) and that Hath1 inhibits the
proliferation of colon cancer cells possibly through upregulating
the expression of Muc2 and p27 and downregulating the expression of
cyclin D1 (24). Collectively,
these studies highlight the important role of Hath1 as a tumor
suppressor gene in these types of tumor and further investigation
of the functions of Hath1 may provide potential molecular targets
for the treatment of these tumors.
However, to the best of our knowledge, no studies on
the biological role of Hath1 in cutaneous SCC have been reported to
date. Therefore, the current study predominantly focused on the
biological function of the Hath1 gene. In order to examine the
effects of the Hath1 gene on the growth, proliferation and
apoptosis of cutaneous SCC, Hath1 was alternately silenced with
short hairpin RNA (shRNA), or overexpressed using a recombinant
eukaryotic expression vector carrying the Hath1 gene. KUMA5 human
cutaneous squamous carcinoma cells were infected with
pcDNA3.1-Hath1 or pGenesil-1-Hath1. Subsequently, cell
proliferation and apoptosis were examined by MTT assay and flow
cytometry to provide further insight into the potential use of
Hath1 for the targeted therapy of cutaneous SCC. Collectively,
these data suggest that Hath1 may be a novel molecular target for
the treatment of cutaneous SCC.
Materials and methods
Construction of the pcDNA3.1-Hath1
vector
The present study was approved by an ethical review
committee of the Shanghai First People's Hospital (Shanghai,
China), and written informed consent was obtained from all
participants at the Shanghai First People's Hospital Affiliated to
Shanghai Jiao Tong University School of Medicine (Shanghai, China).
Total RNA was extracted from normal human cutaneous tissue using 1
ml of TRIzol reagent (cat. no. 15596-018; Invitrogen Life
Technologies, Carlsbad, CA, USA) according to the manufacturer's
instructions. cDNA was then synthesized using an ABI TaqMan
one-step RT-PCR Master Mix Reagents kit (cat. no. 4309109; Applied
Biosystems, Foster City, CA, USA). Based on the GenBank sequence
(NC_000004.12), upstream and downstream primers were synthesized
for Hath1 gene amplification. The restriction enzyme sites for
BamHI and HindIII were added to the 5′ end of each
primer. Subsequently, the full-length Hath1 gene was amplified by
quantitative polymerase chain reaction (qPCR) from the cDNA
template with the primers containing the BamHI and
HindIII restriction sites. The PCR product was then ligated
into the pMD18-T vector (cat. no. 6011; Takara Bio., Inc., Dalian,
China), transformed and screened for positive clones. Following
sequence verification, the correct sequence was cloned into the
pcDNA3.1 expression vector (cat. no. V790-20; Invitrogen Life
Technologies) to construct the pcDNA3.1-Hath1 recombinant
expression vector. The Hath1 primer was synthesized by Sangon
Biotech (Shanghai) Co., Ltd. (Shanghai, China).
shRNA design and expression plasmid
vector construction
The following sequence was used to design the shRNA:
Hath1,
AAACGACAAGAAGCTGTCCAAATAGTGAAGCCACAGATGTATTTGGACAGCTTCTTGTCGTTG.
The DNA sequence targeting the Hath1 gene was cloned into the
pGenesil-1 (cat. no. VRG0358; Wuhan Genesil Biotechnology Co.,
Ltd., Wuhan, China), a vector expressing shRNA and enhanced green
fluorescent protein in mammalian cells. Recombinant plasmid
pGenesil-1-Hath1 and pGenesil-1 scramble control were constructed
and verified by Wuhan Genesil Biotechnology Co., Ltd.
Cell culture and transfection
The KUMA5 human cutaneous squamous carcinoma cell
line was purchased from the Cell Bank of the Chinese Academy of
Sciences (Shanghai, China) and cultured in Dulbecco's modified
Eagle's medium (DMEM)/F12 medium (Gibco-BRL, Carlsbad, CA, USA:
nutrient mixture F12; cat. no. 11039-021) supplemented with 10%
fetal bovine serum (Gibco-BRL; cat. no. 12483-020) in a humidified
incubator containing 5% CO2 at 37° C. When cultured to
exponential phase, KUMA5 cells were replated at 5×104
cells/well in six-well plates. Transient transfection was performed
using Lipofectamine 2000 transfection reagent (cat. no. 11668-019;
Invitrogen Life Technologies) according to the manufacturer's
instructions. Screening of stably transfected cells was performed
as follows: The cells were transfected with either the
pcDNA3.1-Hath1 or pGenesil-1-Hath1 eukaryotic expression vectors
and then cultured in DMEM/F12 containing 800 µg/ml G418
(cat. no. sc-29065; Santa Cruz Biotechnology, Inc., Santa Cruz, CA,
USA) for 2 weeks. Positive clones were screened and verified by
reverse transcription (RT)-qPCR and western blot analysis.
RNA isolation, RT-qPCR and western blot
analysis
mRNA transcript expression was quantified via
RT-qPCR and normalized to the expression of β-actin. Total
RNA from the KUMA5 cells that had been transfected with either the
pcDNA3.1-Hath1, pGenesil-1-Hath1, pGenesil-1 scramble control or
blank control was isolated using TRIzol reagent according to the
manufacturer's instructions and the quality of the RNA was
confirmed (i.e., A260/A280>1.8). qPCR was
performed using an ABI 7900HT Fast Real-Time PCR System (cat. no.
4329003; Applied Biosystems). Detailed sequences of the primers are
as follows: β-actin, forward 5′-ATGATATCGCCGCGCTCGTCGTC-3′
and reverse 5′-CGCTCGGCCGTGGTGGTGAA-3′; Hath1, forward
5′-CGCCGCCCAGTATTTGCTA-3′ and reverse 5′-ATTCACCTGTTTGCTGGAAGG-3′.
Total protein from cell lines was extracted using the ReadyPrep
Protein Extraction kit (cat. no. 163-2087; Bio-Rad Laboratories,
Hercules, CA, USA). The protein concentration was determined using
a BCA Protein Assay kit (cat. no. 23227; Pierce Biotechnology,
Inc., Rockford, IL, USA). Protein lysates were loaded onto a 10%
sodium dodecyl sulfate polyacryl-amide gel. Separated protein bands
were electrotransferred onto polyvinylidene fluoride (PVDF)
membranes. The PVDF membranes were then blocked in Tris-buffered
saline (TBS; 10 mM Tris-HCl, pH 7.5 and 150 mM NaCl) containing 5%
non-fat dry milk at room temperature for 1 h and incubated at 4° C
overnight with anti-Hath1 monoclonal antibody (1:400; cat. no.
sc-136173; Santa Cruz Biotechnology, Inc.) and anti-β-actin
monoclonal antibody (1:1,000; cat. no. sc-47778; Santa Cruz
Biotechnology, Inc.). Following primary antibody incubation, the
membranes were washed in TBS with Tween 20 (10 mM Tris-HCl, pH 7.5,
150 mM NaCl and 0.05% Tween-20) and a rabbit anti-mouse
IgG-horseradish peroxidase antibody (1:2,000; cat. no. sc-358920;
Santa Cruz Biotechnology, Inc.) was added. The membrane was then
incubated at 37° C for 2 h. ECL Plus Western Blotting Substrate
(cat. no. 32132; Pierce Biotechnology, Inc.) was used to visualize
the immunoreactive bands. Relative protein level was normalized to
β-actin concentration. Three separate experiments were performed in
duplicate for each treatment.
Examination of cell viability
Cell viability was determined using an MTT assay
(cat. no. M2128; Sigma-Aldrich, St. Louis, MO, USA). Cells were
cultured in 24-well plates at a concentration of 5×104
and allowed to adhere. Following treatment for different time
intervals, 100 µl MTT (0.5 mg/ml) was added into the cells
and the mixture was incubated for 4 h at 37° C. Subsequently, the
supernatant was removed and dimethyl sulfoxide (cat. no. D2650;
Sigma-Aldrich) was used to dissolve the resultant formazan
crystals. The absorbance was read at 570 nm (EL309 Automated
Microplate Reader; Bio-Tek Instruments, lnc., Winooski, VT, USA).
Six wells were measured for each group and the experiment was
repeated three times.
Flow cytometric analysis of cell
apoptosis
To detect cell apoptosis, the proliferating phase
KUMA5 cells (5×104 clones) were trypsinized, washed with
cold phosphate-buffered saline and resuspended in binding buffer
according to the manufacturer's instructions of the Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) Apoptosis
Detection kit (cat. no. KGA106; Nanjing KeyGen Biotech Co., Ltd.,
Nanjing, China). FITC-Annexin V and PI were added to the fixed
cells for 20 min in darkness at room temperature. Subsequently,
Annexin V binding buffer was added to the mixture prior to
measuring fluorescence using a FACSort flow cytometer (BD
Biosciences, Mountain View, CA, USA). Cell apoptosis was analyzed
using Cell Quest 3.0 software (Becton Dickinson, Franklin Lakes,
NJ, USA).
Statistical analysis
SPSS 13.0 (SPSS, Inc., Chicago, IL, USA) was used
for data analysis. All experiments were repeated three times and
data are presented as the mean ± standard deviation. Differences
between groups were analyzed using one-way analysis of variance.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Expression of Hath1 in KUMA5 cells
Total cellular RNA and protein were extracted from
KUMA5 cells that had been transfected with the pcDNA3.1-Hath1,
pGenesil-1-Hath1, pGenesil-1 scramble control or a blank control.
RT-qPCR and western blot analysis were performed to determine the
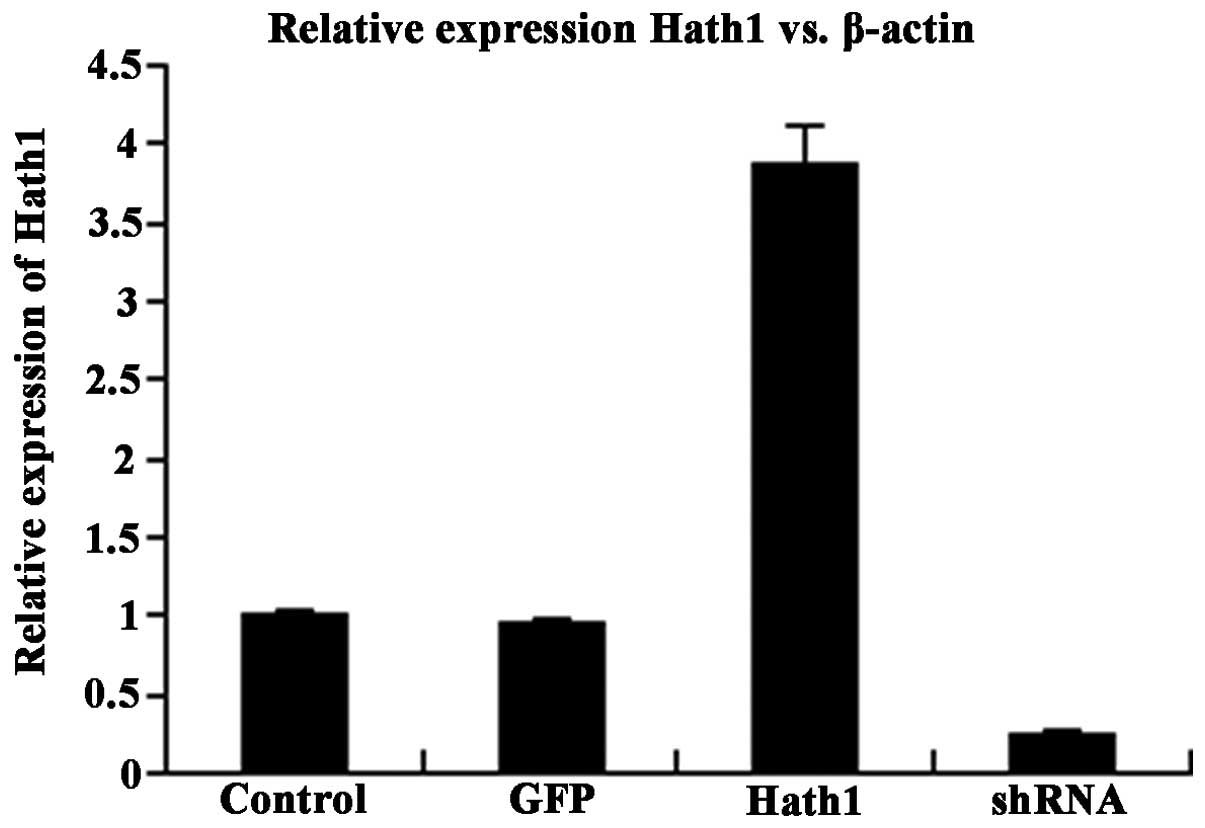
mRNA and protein expression of Hath1, respectively. The results
demonstrated that the expression of Hath1 in KUMA5 cells
transfected with pcDNA3.1-Hath1 was nearly four times higher than
in the control groups (Fig. 1).
The efficiency of gene silencing through pGenesil-1-Hath1
transfection was 78% (Fig. 1).
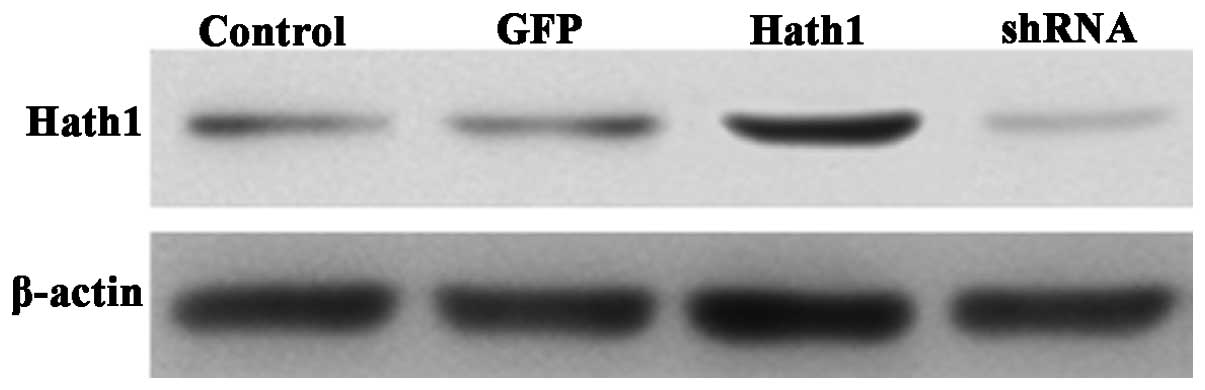
The protein expression of Hath1 was high in the
pcDNA3.1-Hath1-transfected group, low in the control group, and
barely detectable in the pGenesil-1-Hath1-transfected group
(Fig. 2). Therefore, it was
concluded that KUMA5 strains with stable overexpression or
depletion of the Hath1 gene had been successfully created.
Effect of Hath1 on KUMA5 cell
proliferation
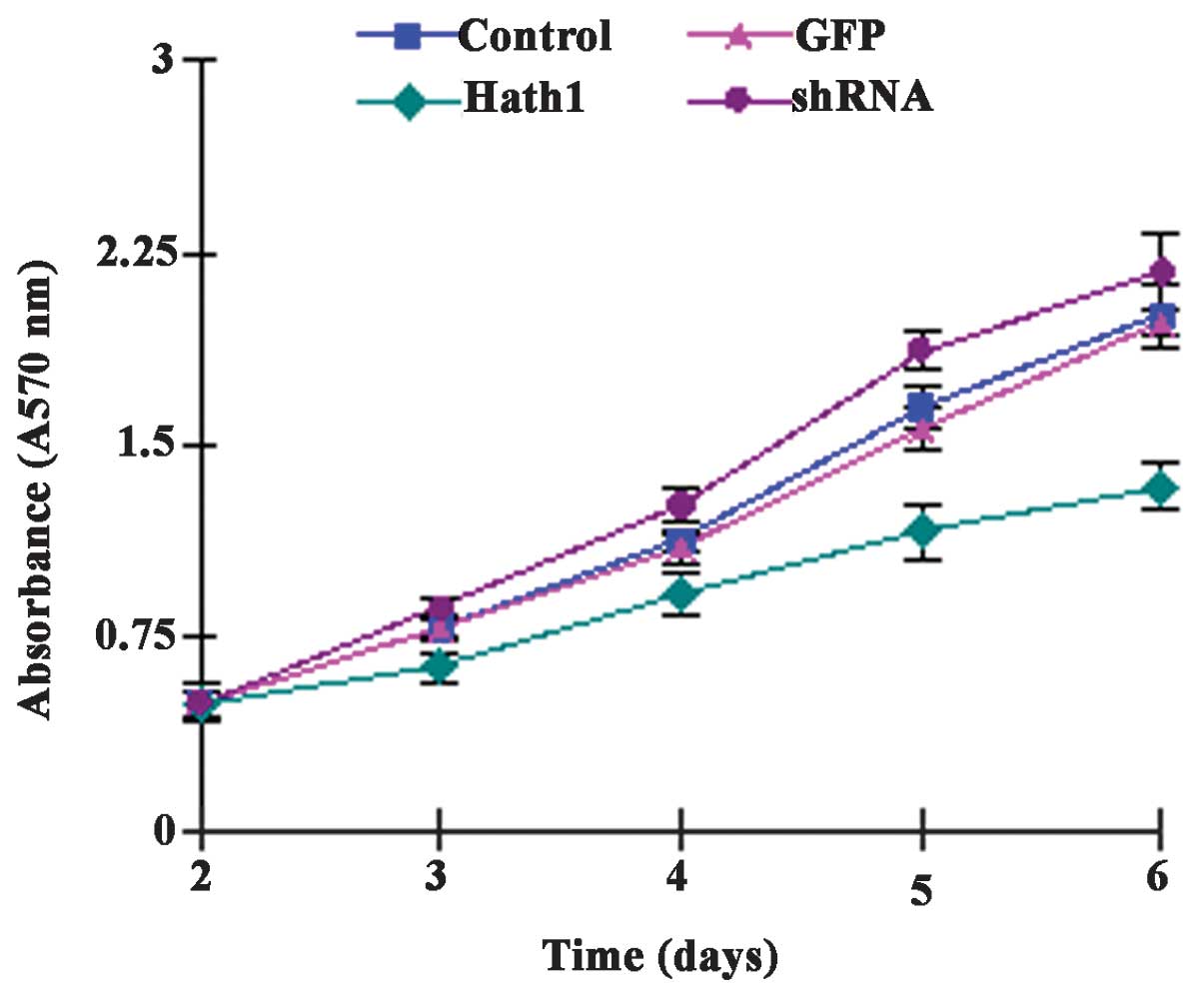
It was hypothesized that cell proliferation may be
affected by the level of Hath1 in cutaneous SCC cells. To examine
the effect of Hath1 on cell proliferation, an equal number of KUMA5
cells transfected with either the pcDNA3.1-Hath1, pGenesil-1-Hath1,
pGenesil-1 scramble control or a blank control were seeded and
analyzed using an MTT assay. The present study found that Hath1
overexpression attenuated the proliferation of KUMA5 cells and
Hath1 gene RNA interference enhanced the proliferation of KUMA5
cells (Fig. 3). Taken together,
Hath1 clearly inhibited the proliferation and growth of SCC cells,
indicating that Hath1 may act as a potential tumor suppressor in
cutaneous SCC and that Hath1 may represent a novel target for the
treatment of SCC.
Effect of Hath1 on the apoptosis of KUMA5
cells
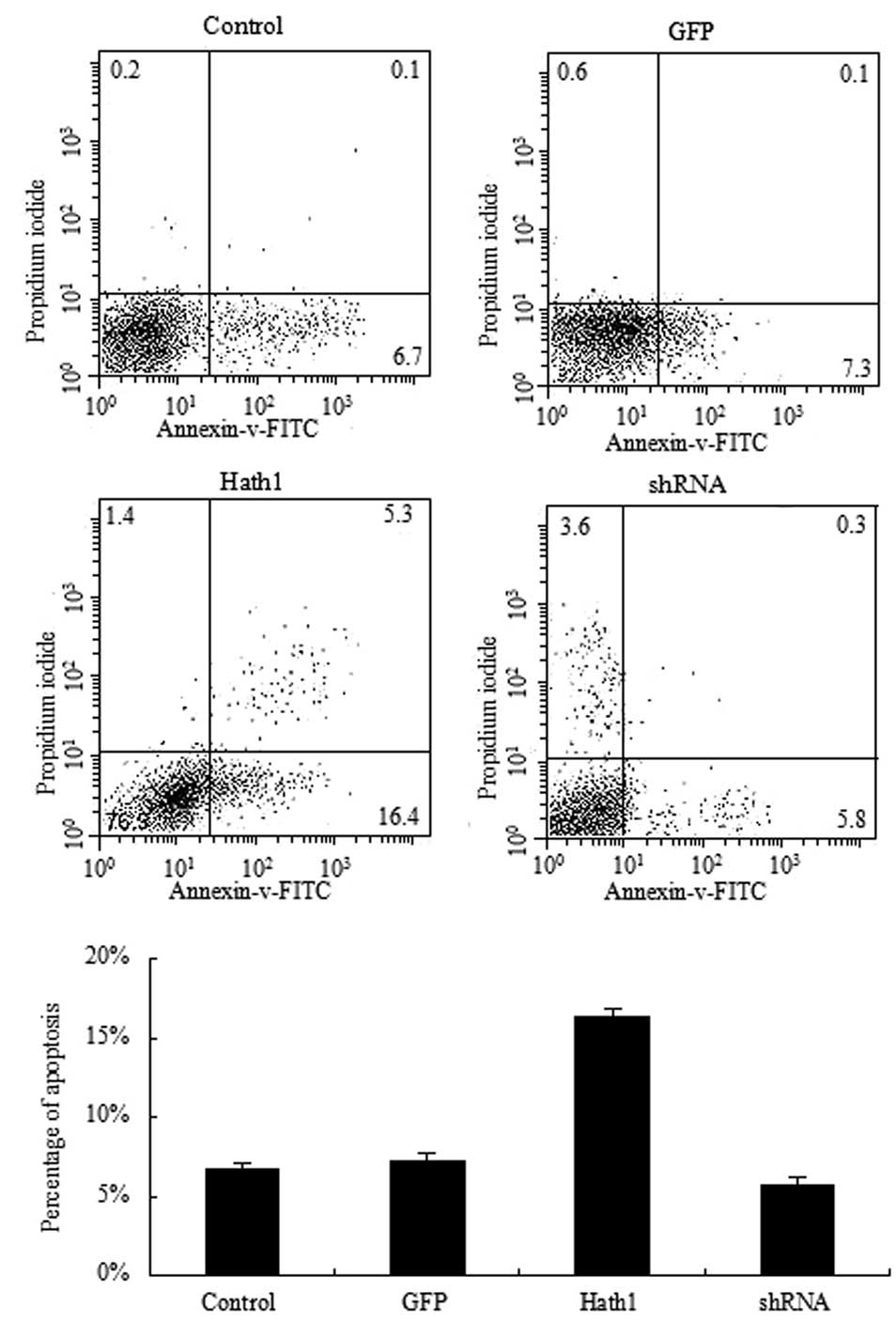
To further confirm the inhibitory effect of Hath1 on
KUMA5 cells, the rate of apoptosis was examined by Annexin V and PI
staining of KUMA5 cells transfected with the pcDNA3.1-Hath1,
pGenesil-1-Hath1, pGenesil-1 scramble control or a blank control.
FITC-Annexin V and PI were added to the fixed cells for 20 min in
darkness at room temperature. Subsequently, Annexin V binding
buffer was added to the mixture prior to measuring the fluorescence
using a FACSort flow cytometer (BD Biosciences). Cell apoptosis was
analyzed using Cell Quest 3.0 software (Becton Dickinson). The
results demonstrated that a significantly higher rate of apoptosis
was present in the KUMA5 cells overexpressing Hath1 compared with
that of the control groups (Fig.
4). In addition, a significantly lower rate of apoptosis was
observed in the KUMA5 cells transfected with pGenesil-1-Hath1
compared with that of the control groups (Fig. 4). Therefore, these results suggest
that overexpression of Hath1 may promote apoptosis in KUMA5
cells.
Discussion
Hath1, located on the chromosomal region 4q22, is
the mammalian homologue of Drosophila atonal, a bHLH transcription
factor. It is known to be important in the differentiation of
various cells, including cerebellar granule cells (33), inner hair cells in the auditory
system (34), epidermal Merkel
cells (32), intestinal secretory
cells (35,36) and hindbrain neurons (37). Previous studies have demonstrated
that Hath1 acts as a tumor suppressor gene in various types of
cancer, including colon cancer (14-24),
medulloblastoma (25,26), gastric carcinoma (27,28),
intestinal neuroendocrine tumors (29), lung cancer (30,31)
and Merkel cell carcinoma (32).
However, to date, no studies on the biological role
of Hath1 in cutaneous SCC have been reported. The present study
demonstrated for the first time, to the best of our knowledge, the
anti-proliferative and pro-apoptotic activity of Hath1 in cutaneous
SCC, using a variety of techniques, including overexpression of
Hath1 and shRNA-mediated Hath1 silencing. As shown in Figs. 1 and 2, Hath1 was successfully silenced or
overexpressed. The expression level of Hath1 in
pGenesil-1-Hath1-transfected cells was significantly decreased to
22% of the normal levels (P<0.05). Cells transfected with
pcDNA3.1-Hath1 had significantly higher levels of Hath1 expression,
which was ~3.9 times higher than the control groups (P<0.05).
MTT assays demonstrated that the proliferation of
KUMA5/pGenesil-1-Hath1-transfected cells was promoted in a
time-dependent manner, however, proliferation was inhibited in
KUMA5/pcDNA3.1-Hath1-infected cells. Furthermore, the apoptotic
rate of the pGenesil-1-Hath1-trans-fected group significantly
decreased. By contrast, the apoptotic rate of the
KUMA5/pcDNA3.1-Hath1-infected group significantly increased. These
experimental results demonstrated that overexpression of Hath1
could repress the proliferation of KUMA5 cells and promote
apoptosis. However, shRNA-mediated Hath1 silencing enhanced the
growth of KUMA5 cells and inhibited the apoptosis of KUMA5
cells.
The expression level of the Hath1 gene was
frequently found to significantly affect cell proliferation and
apoptosis (14,16,20–22,24).
Zhu et al (20,21,24)
and Leow et al (14)
identified that overexpression of Hath1 inhibits proliferation of
HT29 colon cancer cells through downregulation of cyclin D1 and
upregulation of p27 and MUC2, a goblet cell differentiation marker.
In a previous study, knockout of Hath1 was demonstrated to promote
tumorigenesis in colorectal mouse models with intestine-specific
deletion of Hath1 (18). The
anti-tumor effects of Hath1, including the inhibition of colon
cancer cell proliferation and enhancement of apoptosis in
vitro and in vivo, may be achieved by activation of the
Jun N-terminal kinase signaling pathway (16). According to Zhao et al
(22), Hath1 can effectively
inhibit the proliferation of four colon cancer cell lines, namely
HT-29, LS17T, SW480 and SW620 with weak or absent expression of
Hath1. In addition, the inhibition of cell proliferation by Hath1
was inversely proportional to the expression of Hath1 in these four
colon cancer cell lines. However, instead of inducing apoptosis, an
increase in tumor cell sensitivity to the chemotherapeutic drug
5-fluorouracil was observed in one of the four colon cancer cell
lines. By contrast, shRNA-mediated Hath1 gene silencing using
retroviral vectors could significantly accelerate the growth rate
of the normal rat intestinal epithelial cell line IEC6 with
suppressed growth by high expression of the Hath1 gene, which also
confirmed the tumor suppressor activity of Hath1 (22). A previous study (38) demonstrated that retroviral-mediated
Hath1 expression can significantly reduce human esophageal
keratinocyte cell proliferation and induce marked expression of
Barrett's esophagus markers Mucin-2 and Keratin-20.
The results of the present study, which build on earlier studies
(14,16,20–22,24),
confirm the anti-proliferative and pro-apoptotic activities of
Hath1.
In conclusion, the present results verify the
anti-tumor effects of Hath1 on the proliferation and apoptosis of
cutaneous SCC. Hath1 may be a potential therapeutic target for the
inhibition of growth and progression of cutaneous SCC. These
studies provide insights into the tumorigenesis and development of
cutaneous SCC and provide new strategies and targets for the
treatment of cutaneous SCC. However, further studies are required
to clarify the associated molecular mechanisms and signal
transduction of Hath1.
References
|
1
|
Veness MJ, Morgan GJ, Palme CE and Gebski
V: Surgery and adjuvant radiotherapy in patients with cutaneous
head and neck squamous cell carcinoma metastatic to lymph nodes:
Combined treatment should be considered best practice.
Laryngoscope. 115:870–875. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Preston DS and Stern RS: Nonmelanoma
cancers of the skin. N Engl J Med. 327:1649–1662. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Euvrard S, Kanitakis J and Claudy A: Skin
cancers after organ transplantation. N Engl J Med. 348:1681–1691.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
McGuire JF, Ge NN and Dyson S: Nonmelanoma
skin cancer of the head and neck I: Histopathology and clinical
behavior. Am J Otolaryngol. 30:121–133. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Martinez JC and Cook JL: High-risk
cutaneous squamous cell carcinoma without palpable lymphadenopathy:
Is there a therapeutic role for elective neck dissection? Dermatol
Surg. 33:410–420. 2007.PubMed/NCBI
|
|
6
|
Weinberg AS, Ogle CA and Shim EK:
Metastatic cutaneous squamous cell carcinoma: An update. Dermatol
Surg. 33:885–899. 2007.PubMed/NCBI
|
|
7
|
Bertero T, Bourget-Ponzio I, Puissant A,
et al: Tumor suppressor function of miR-483-3p on squamous cell
carcinomas due to its pro-apoptotic properties. Cell Cycle.
12:2183–2193. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu D, Feng X, Wu X, et al: Tumor
suppressor in lung cancer 1 (TSLC1), a novel tumor suppressor gene,
is implicated in the regulation of proliferation, invasion, cell
cycle, apoptosis and tumorigenicity in cutaneous squamous cell
carcinoma. Tumour Biol. 34:3773–3783. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang C, Wu D, Jia J, et al: DLC1 as a
regulator of proliferation, invasion, cell cycle and apoptosis in
cutaneous squamous cell carcinoma. Tumour Biol. 34:2633–2643. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Scortegagna M, Martin RJ, Kladney RD,
Neumann RG and Arbeit JM: Hypoxia-inducible factor-1alpha
suppresses squamous carcinogenic progression and
epithelial-mesenchymal transition. Cancer Res. 69:2638–2646. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sharma A, Ramanjaneyulu A, Ray R and
Rajeswari MR: Involvement of high mobility group B proteins in
cisplatin-induced cytotoxicity in squamous cell carcinoma of skin.
DNA Cell Biol. 28:311–318. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bito T, Sumita N, Ashida M, et al:
Inhibition of epidermal growth factor receptor and PI3K/Akt
signaling suppresses cell proliferation and survival through
regulation of Stat3 activation in human cutaneous squamous cell
carcinoma. J Skin Cancer. 2011:8745712011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ben-Arie N, McCall AE, Berkman S, Eichele
G, Bellen HJ and Zoghbi HY: Evolutionary conservation of sequence
and expression of the bHLH protein Atonal suggests a conserved role
in neurogenesis. Hum Mol Genet. 5:1207–1216. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Leow CC, Romero MS, Ross S, Polakis P and
Gao WQ: Hath1, down-regulated in colon adenocarcinomas, inhibits
proliferation and tumorigenesis of colon cancer cells. Cancer Res.
64:6050–6057. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Leow CC, Polakis P and Gao WQ: A role for
Hath1, a bHLH transcription factor, in colon adenocarcinoma. Ann NY
Acad Sci. 1059:174–183. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bossuyt W, Kazanjian A, De Geest N, et al:
Atonal homolog 1 is a tumor suppressor gene. PLoS Biol. 7:e392009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sikandar SS, Pate KT, Anderson S, et al:
NOTCH signaling is required for formation and self-renewal of
tumor-initiating cells and for repression of secretory cell
differentiation in colon cancer. Cancer Res. 70:1469–1478. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kazanjian A, Noah T, Brown D, Burkart J
and Shroyer NF: Atonal homolog 1 is required for growth and
differentiation effects of notch/gamma-secretase inhibitors on
normal and cancerous intestinal epithelial cells. Gastroenterology.
139:918–928. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu DH, Gong J, Ren K, et al: Hath1
expression is down-regulated in colon non-mucinous adenocarcinomas
compared with the expression in normal colon mucosa. Chong Qing Yi
Ke Da Xue Xue Bao. 35:666–669. 2010.In Chinese.
|
|
20
|
Zhu DH, Gong JP, Ren K, Sun JM and Wei SD:
Hath1 gene transfer inhibits the proliferation of colonic cancer
cells in vitro. Nan Fang Yi Ke Da Xue Xue Bao. 30:1005–1008.
2010.In Chinese. PubMed/NCBI
|
|
21
|
Zhu DH, Wen YW, Yang H and Liang G:
Anti-cancer effect of Hath1 in pathogenesis of non-mucinous colon
adenocarcinoma. Ji Lin Da Xue Xue Bao (Yi Xue Ban). 37:700–706.
2011.In Chinese.
|
|
22
|
Zhao LL, Liu Z and Zhang YJ: Hath1 gene
inhibits colorectal cancer cell growth. Zhong Guo Yi Yao Sheng Wu
Ji Shu. 6:204–208. 2011.In Chinese.
|
|
23
|
Peignon G, Durand A, Cacheux W, et al:
Complex interplay between β-catenin signalling and Notch effectors
in intestinal tumorigenesis. Gut. 60:166–176. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu DH, Niu BL, Du HM, Ren K, Sun JM and
Gong JP: Hath1 inhibits proliferation of colon cancer cells
probably through up-regulating expression of Muc2 and p27 and
down-regulating expression of cyclin D1. Asian Pac J Cancer Prev.
13:6349–6355. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao H, Ayrault O, Zindy F, Kim JH and
Roussel MF: Post-transcriptional down-regulation of Atoh1/Math1 by
bone morphogenic proteins suppresses medulloblastoma development.
Genes Dev. 22:722–727. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Flora A, Klisch TJ, Schuster G and Zoghbi
HY: Deletion of Atoh1 disrupts Sonic Hedgehog signaling in the
developing cerebellum and prevents medulloblastoma. Science.
326:1424–1427. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sekine A, Akiyama Y, Yanagihara K and
Yuasa Y: Hath1 up-regulates gastric mucin gene expression in
gastric cells. Biochem Biophys Res Commun. 344:1166–1171. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang X, Yu H, Yang Y, et al: SOX2 in
gastric carcinoma, but not Hath1, is related to patients'
clinicopathological features and prognosis. J Gastrointest Surg.
14:1220–1226. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Heiskala K, Arola J, Heiskala M and
Andersson LC: Expression of Reg IV and Hath1 in neuroendocrine
neoplasms. Histol Histopathol. 25:63–72. 2010.
|
|
30
|
Chen JB, Liu JH, Liu YF, et al: Expression
and clinical significance of Atoh1 in lung cancer. Lin Chuang Fei
Ke Za Zhi. 11:1744–1746. 2011.In Chinese.
|
|
31
|
Xu HT, Xie XM, Li QC, et al: Atonal
homolog 1 expression in lung cancer correlates with inhibitors of
the Wnt pathway as well as the differentiation and primary tumor
stage. APMIS. 121:111–119. 2013. View Article : Google Scholar
|
|
32
|
Leonard JH, Cook AL, Van Gele M, et al:
Proneural and proneuroendocrine transcription factor expression in
cutaneous mechanoreceptor (Merkel) cells and Merkel cell carcinoma.
Int J Cancer. 101:103–110. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ben-Arie N, Bellen HJ, Armstrong DL, et
al: Math1 is essential for genesis of cerebellar granule neurons.
Nature. 390:169–172. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bermingham NA, Hassan BA, Price SD, et al:
Math1: An essential gene for the generation of inner ear hair
cells. Science. 284:1837–1841. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang Q, Bermingham NA, Finegold MJ and
Zoghbi HY: Requirement of Math1 for secretory cell lineage
commitment in the mouse intestine. Science. 294:2155–2158. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shroyer NF, Helmrath MA, Wang VY, Antalffy
B, Henning SJ and Zoghbi HY: Intestine-specific ablation of mouse
atonal homolog 1 (Math1) reveals a role in cellular homeostasis.
Gastroenterology. 132:2478–2488. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rose MF, Ren J, Ahmad KA, et al: Math1 is
essential for the development of hindbrain neurons critical for
perinatal breathing. Neuron. 64:341–354. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kong J, Crissey MA, Sepulveda AR and Lynch
JP: Math1/Atoh1 contributes to intestinalization of esophageal
keratinocytes by inducing the expression of Muc2 and Keratin-20.
Dig Dis Sci. 57:845–857. 2012. View Article : Google Scholar :
|