Introduction
Visceral adipose tissue-derived serine protease
inhibitor (vaspin) was initially detected in the visceral adipose
tissue of Otsuka Long-Evans Tokushima Fatty rats (1). Vaspin is a compensatory adipokine
with anti-inflammatory properties that is able to improve insulin
sensitivity, as demonstrated by the administration of recombinant
human vaspin protein to diet-induced obese mice (1,2). In
addition, elevated serum vaspin concentrations have been detected
in humans with obesity, type 2 diabetes and polycystic ovary
syndrome, and a correlation has been identified between vaspin and
C-reactive protein levels (3–7).
In addition to its ability to ameliorate diabetes,
vaspin exerts protective effects on heart disease. For example, low
vaspin concentrations correlate with coronary artery disease (CAD)
severity and unstable angina pectoris (8). Previous studies have also
demonstrated that vaspin may inhibit endothelial cell apoptosis and
attenuate high glucose-stimulated vascular smooth muscle cell
(VSMC) proliferation and chemokinesis (9,10).
Furthermore, vaspin has been reported to stimulate the 78-kDa
glucose-regulated protein (GRP78)/murine tumor cell DnaJ-like
protein 1 (MTJ-1) complex and subsequent signals, exerting
beneficial effects on endoplasmic reticulum (ER) stress-induced
metabolic dysfunction (11). In
the cardiovascular system, vaspin acts as a ligand for the
cell-surface GRP78/voltage-dependent anion channel (VDAC) complex
in endothelial cells, promotes proliferation, inhibits apoptosis,
and protects against diabetes mellitus-associated vascular injury
(12).
Macrophage apoptosis is an important characteristic
of atherosclerotic plaque development. Particularly in advanced
lesions, macrophage apoptosis is a key factor that aggravates
advanced plaque necrosis (13). ER
stress has emerged as a general mediator of vascular inflammation
and endothelial dysfunction in atherosclerosis, and has been
reported to contribute to plaque vulnerability via the induction of
macrophage and VSMC apoptosis (14). ER is the major cellular site for
protein folding and trafficking, and is central to numerous
cellular functions. Failure of the adaptive capacity of ER results
in activation of the unfolded protein response, which can lead to
cell pathology and subsequent tissue dysfunction (15). In pathological settings, prolonged
ER stress may trigger apoptosis, either through the functions of
inositol-requiring enzyme-1 at the ER or through downstream
effectors, including C/EBP-homologous protein (CHOP). Activation of
ER stress contributes to macrophage death and subsequent plaque
necrosis in advanced atheroma, a process that has widely been
accepted as an important factor in atherosclerosis (16).
It has previously been demonstrated that low vaspin
serum concentrations are correlated with recent ischemic events in
patients with carotid stenosis (17). In addition, decreased vaspin serum
levels have been detected in asymptomatic patients with CAD
(18). Furthermore, Zhang et
al also demonstrated that low vaspin concentrations are
correlated with the severity of CAD and acute coronary diseases
(19).
The present study aimed to determine whether the
effects of vaspin on plaque progression and morphology in
apolipoprotein E (apoE)−/− mice were mediated by
inhibition of ER stress-induced apoptotic pathways. In vitro
experiments were also performed to examine the role of vaspin in
THP-1 macrophages.
Materials and methods
Ethics statement
The experiments outlined in the present study
conform to the Guide for the Care and Use of Laboratory Animals
published by the National Institutes of Health (20). All animal procedures were approved
by the Animal Care and Use Committees of the Shanghai Tenth
People's Hospital (Shanghai, China).
Lentivirus preparation
The vaspin-encoding lenti-virus (LV5-mus-vaspin) and
control lentivirus (LV5NC) were purchased from Shanghai GenePharma
Co., Ltd. (Shanghai, China). The lentiviruses also expressed green
fluorescent protein (GFP). Final titers were 1×108 TU/ml
for LV5-mus-vaspin and 1×109 TU/ml for LV5NC. All
experiments described in the present study were performed using the
same lentiviruses.
Experimental animals
Male apoE−/− mice (age, 6 weeks; n=16)
and male C57BL/6 mice (age, 6 weeks; n=8) were obtained from
Beijing Vital River Laboratory Animal Technology Co., Ltd.
(Beijing, China) and fed a high cholesterol diet containing 16.6%
fat, 10.6% sucrose and 1.3% cholesterol (Shanghai SLAC Laboratory
Animal Co., Ltd., Shanghai, China) for 12 weeks. The mice were
housed in cages with a 12-h light/dark cycle. The temperature of
the animal room was maintained at 23±2°C and the relative humidity
was maintained at 55±15%. All of the animals were provided free
access to drinking water and food. The mice were divided into two
treatment groups, LV5NC (n=8) and LV-mus-vaspin (n=8). Each mouse
was injected with 200 µl lentivirus through the tail vein.
Body weight and food intake were monitored every 5 days in both
groups.
Plasma glucose and lipid panel
analysis
Following a 12-h fast and the induction of
anesthesia with 3% pentobarbital (30 mg/kg; China National
Medicines Corporation, Ltd., Shanghai, China), plasma samples (250
µl) were obtained from all apoE−/− mice. Fasting
glucose, total cholesterol and triglyceride, low-density
lipoprotein cholesterol (LDL-C), and high-density lipoprotein
cholesterol (HDL-C) levels were measured using a colorimetric
enzymatic assay system (Roche Modular P-800; Roche Diagnostics,
Basel, Switzerland).
Histological analysis
Following 12 weeks of treatment, at the age of 18
weeks, the animals were sacrificed by euthanasia with an
intraperitoneal injection of 3% pentobarbital at a concentration of
70 mg/kg. The heart and aorta were removed following in situ
perfusion with phosphate-buffered saline (PBS). The tissue samples
were fixed in 4% paraformaldehyde for 1 day, and 4-µm
paraffin-embedded cross-sections of the aortic root were prepared
for the subsequent histological and immunohistochemical analyses.
The aortic arch and abdominal aorta were stained en face with Oil
Red O (60% saturated Oil Red O in 40% deionized water;
Sigma-Aldrich, St. Louis, MO, USA), in order to measure lesion
development. The cross-sections were stained with hematoxylin and
eosin kit (1.5% hematoxylin and 1% eosin; Beyotime Institute of
Biotechnology, Haimen, China) and Masson's trichrome
(Sigma-Aldrich) containing Biebrich scarlet-acid fuchsin solution
(0.9% biebrich scarlet, 0.1% acid fuchsin and 1% acetic acid), 10%
phosphotungstic acid solution, 10% phosphomolybdic acid solution
and aniline blue solution (2.4% aniline blue and 2% acetic acid),
in order to evaluate atherosclerotic lesion complexity and the
percentage of plaque size and necrotic core size. All the
experiments were performed according to the manufacturer's
protocols. Images of staining were captured using a Leica DMI6000
microscope (Leica Microsystems, Wetzlar, Germany). The percentages
were quantified using Image-Pro Plus 6.0 (Media Cybernetics, Inc.,
Rockville, MD, USA).
Immunohistochemical analysis
The tissue cross-sections were labeled with mouse
monoclonal anti-CHOP (1:300; Cell Signaling Technology, Inc.,
Danvers, MA, USA; cat. no. 2895), mouse monoclonal anti-GFP (1:300;
Cell Signaling Technology; cat. no. 2955), and goat monoclonal
anti-vaspin (1:200; Santa Cruz Biotechnology, Inc., Dallas, TX,
USA; cat. no. sc-79815) at 4°C overnight, following microwave
antigen retrieval in citrate buffer. After washing with PBS with
Tween 20 (PBST), the tissue samples were incubated with rabbit
anti-mouse (1:200; Gene Tech Biotechnology Co., Ltd., Shanghai,
China; cat. no. GK500705) and donkey anti-goat secondary antibodies
(1:200; Santa Cruz Biotechnology, Inc.; cat. no. sc-2042) at 37°C
for 1 h, and then incubated with diaminobenzidine substrate (0.05%;
Sigma-Aldrich) for 1–3 min. Specimens were counter-stained with
hematoxylin and images were captured under a Leica DMI6000
microscope. Three sections from each animal were stained for lesion
quantification, which was expressed as a percentage of total lesion
area. Data are presented as the mean ± standard error of the mean
(SEM).
Cell culture
Human monocytic THP-1 cells were purchased from the
Institute of Biochemistry and Cell Biology (Shanghai, China). The
cells were cultured in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 2 mM
L-glutamine (Gibco; Thermo Fisher Scientific, Inc.) and 10% fetal
bovine serum (BSA; Gibco; Thermo Fisher Scientific, Inc.) at 37°C
with 5% CO2 in a humidified atmosphere (21). The THP-1 cells were incubated with
100 nM phorbol-12-myristate 13-acetate (PMA; Sigma-Aldrich) for 48
h and allowed to differentiate into macrophages. Prior to
incubation with oxidized (ox)-LDL (50 µg/ml; Guangzhou
Yiyuan Biotech., Co., Ltd., Guangzhou, China) for 24 or 48 h, the
PMA-induced macrophages were treated with vaspin (100 ng/ml;
Sigma-Aldrich) for 24 h and harvested for subsequent
experiments.
Flow cytometry (FCM) analysis
Apoptosis was evaluated in three independent
experiments using an Annexin V Apoptosis Detection kit (Roche
Diagnostics Operations, Indianapolis, IN, USA). Macrophages
(1×105 cells/well) were pre-treated with vaspin (100
ng/ml) for 24 h and then incubated with ox-LDL (50 µg/ml)
for 24 or 48 h. The cells were then harvested and resuspended in
binding buffer, followed by double staining with fluorescein
isothiocyanate-conjugated Annexin V (20 µg/ml; 5 µl)
and propidium iodide (50 µg/ml; 5 µl) for 15 min at
20°C, in Ca2+-enriched binding buffer. Fluorescence was
detected by FACScan flow cytometer (BD Biosciences, Franklin Lakes,
NJ, USA).
Terminal deoxyribonucleotide
transferase-mediated deoxyuridine triphosphate nick-end labeling
(TUNEL) assay
The TUNEL assay was performed using an In
Situ Cell Death Detection kit (Roche Diagnostics, Mannheim,
Germany). Macrophages were pretreated with vaspin (100 ng/ml) for
24 h and then stimulated with ox-LDL (50 µg/ml) for 48 h.
The macrophages were subsequently fixed with 4% paraformaldehyde
for 25 min, followed by permeabilization with 0.2% Triton X-100 for
5 min. Finally, the macrophages were incubated with TUNEL reaction
mixture for 1 h at 37°C in the dark, and stained with Hoechst 33342
(Invitrogen; Thermo Fisher Scientific, Inc.) for 20 min. The
fluorescence images were obtained using a fluorescence microscope
(Leica DMI6000; magnification, ×50). The number of TUNEL-positive
cells was expressed as a percentage of positive cells
double-stained with TUNEL and Hoechst 33342.
Protein extraction and western blot
analysis
Protein extraction and western blot analysis were
performed as described in our previous study (21). Cells were lysed with ice-cold
radio-immunoprecipitation assay buffer (Santa Cruz Biotechnology,
Inc.) and centrifuged at 10,000 x g for 5 min at 4°C. Protein
concentrations in the supernatants were measured using a
Bicinchoninc Acid Protein Assay kit (Pierce Biotechnology, Inc.,
Rockford, IL, USA). Briefly, protein samples (20 µg) were
separated by 10 or 12% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred to nitrocellulose membranes (CW
Biotech, Beijing, China). The membranes were blocked with 5% BSA
for 1 h and were then incubated with the following antibodies at
4°C overnight: Anti-activating transcription factor (ATF)6
(1:1,000; ProteinTech Group, Inc., Chicago, IL, USA; cat. no.
24169-AP), anti-CHOP (1:1,000), anti-total (t)-c-Jun N-terminal
kinases (JNK)1/2 (1:800; Cell Signaling Technology, Inc., cat. no.
9258, anti-phosphorylated (p)-JNK1/2 (1:800; Cell Signaling
Technology, Inc.; cat. no. 4671), anti-cleaved-caspase 12 (1:800;
BioVision, Inc., Milpitas, CA, USA; cat. no. 3282-100),
anti-cleaved-caspase 9 (1:800; Abcam, Cambridge, MA, USA; cat. no.
ab2325) and anti-cleaved-caspase 3 (1:800; Abcam; cat. no. ab2302).
Anti-b-actin served as the control antibody (1:5,000; Shanghai
Yeasen Biotechnology Co., Ltd., Shanghai, China; cat. no.
30101ES50) The membranes were then washed with PBST and incubated
with horseradish peroxidase-conjugated anti-rabbit immunoglobulin G
(IgG) secondary antibody (1:2,000; GE Healthcare Life Sciences,
Chalfont, UK; cat. no. NA9310) or anti-mouse IgG secondary
antibodies (1:2,000; GE Healthcare Life Sciences; NA931) for 1 h,
and were visualized using an Odyssey Imaging system (LI-COR
Biosciences, Lincoln, NE, USA).
Statistical analysis
All statistical analyses were performed using SPSS
14.0 (SPSS, Inc., Chicago, IL, USA). Data are presented as the mean
± SEM. Differences between two groups were examined using Student's
t-test. One-way analysis of variance was used to compare multiple
groups, if appropriate, with Bonferroni correction for the post-hoc
analysis. P<0.05 was considered to indicate a statistically
significant difference, and P≤0.016 was considered to indicate a
statistically significant difference for post-hoc analysis. All
experiments were performed at least three times.
Results
Vaspin inhibits the progression of
atherosclerosis in apoE−/− mice
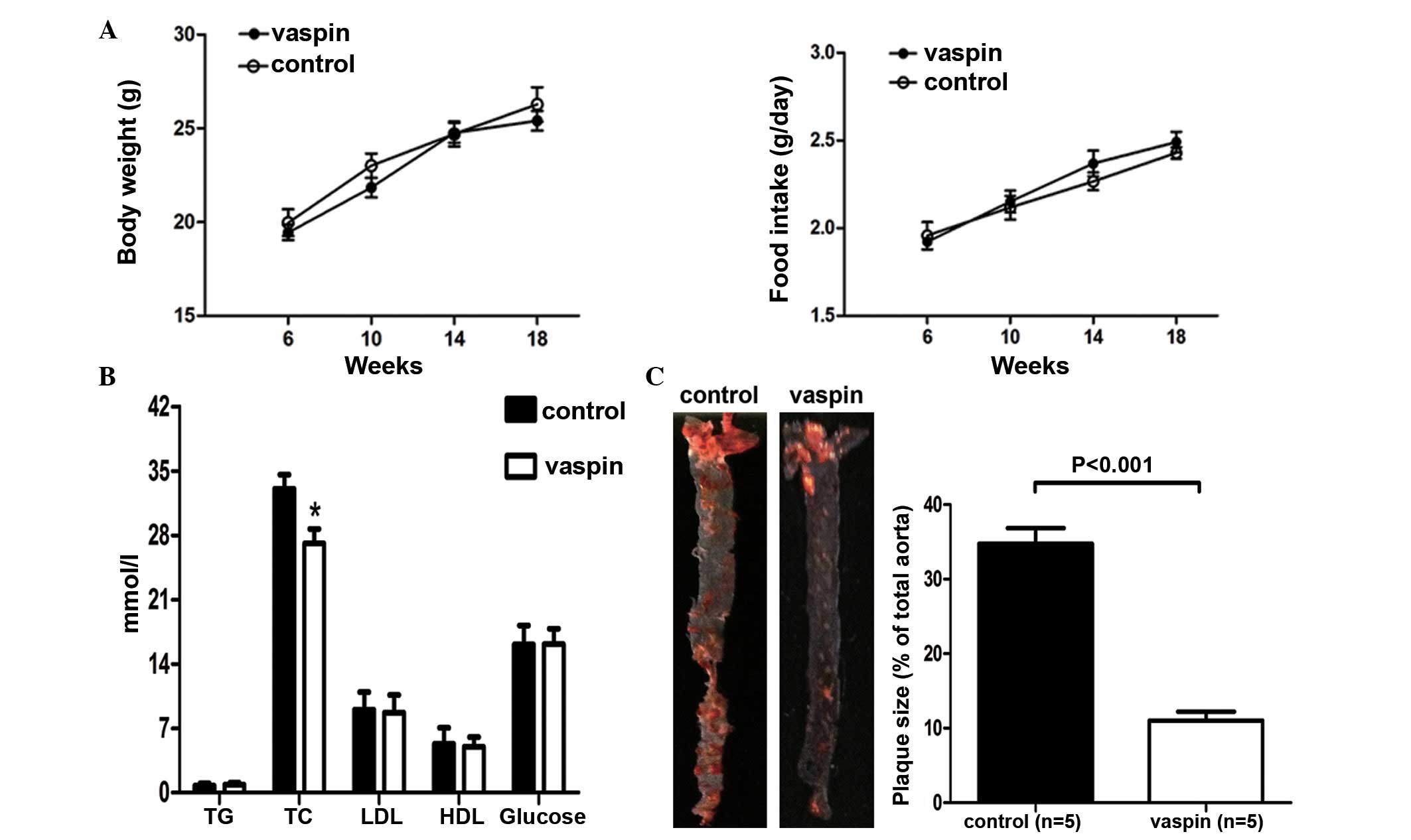
Body weight and food intake of the
apoE−/− mice were not influenced by LV5-mus-vaspin
transfection during the 12-week feeding regimen (Fig. 1A). There was no difference between
the LV5NC and LV5-mus-vaspin-transfected groups in circulating
levels of triglyceride, LDL-C, HDL-C and glucose; however, the
total cholesterol level was markedly decreased in the
LV5-mus-vaspin-transfected mice (27.12±1.39 vs. 33.04±1.70 mmol/l;
P<0.001; n=8; Fig. 1B). Whole
aortas, from the ascending aorta to the abdominal aorta, were cut
open, periaortic fat and blood clots were removed, and the vessels
were longitudinally sectioned and stained with Oil Red O, in order
to detect lipid deposition. The lesions were then measured as a
percentage of plaque area. As shown in Fig. 1C, LV5-mus-vaspin-transfected
apoE−/− mice exhibited significantly reduced lesion
development, as compared with the LV5NC-transfected group (plaque
area 11.03±1.34 vs. 34.74±1.98%; P<0.001; n=5).
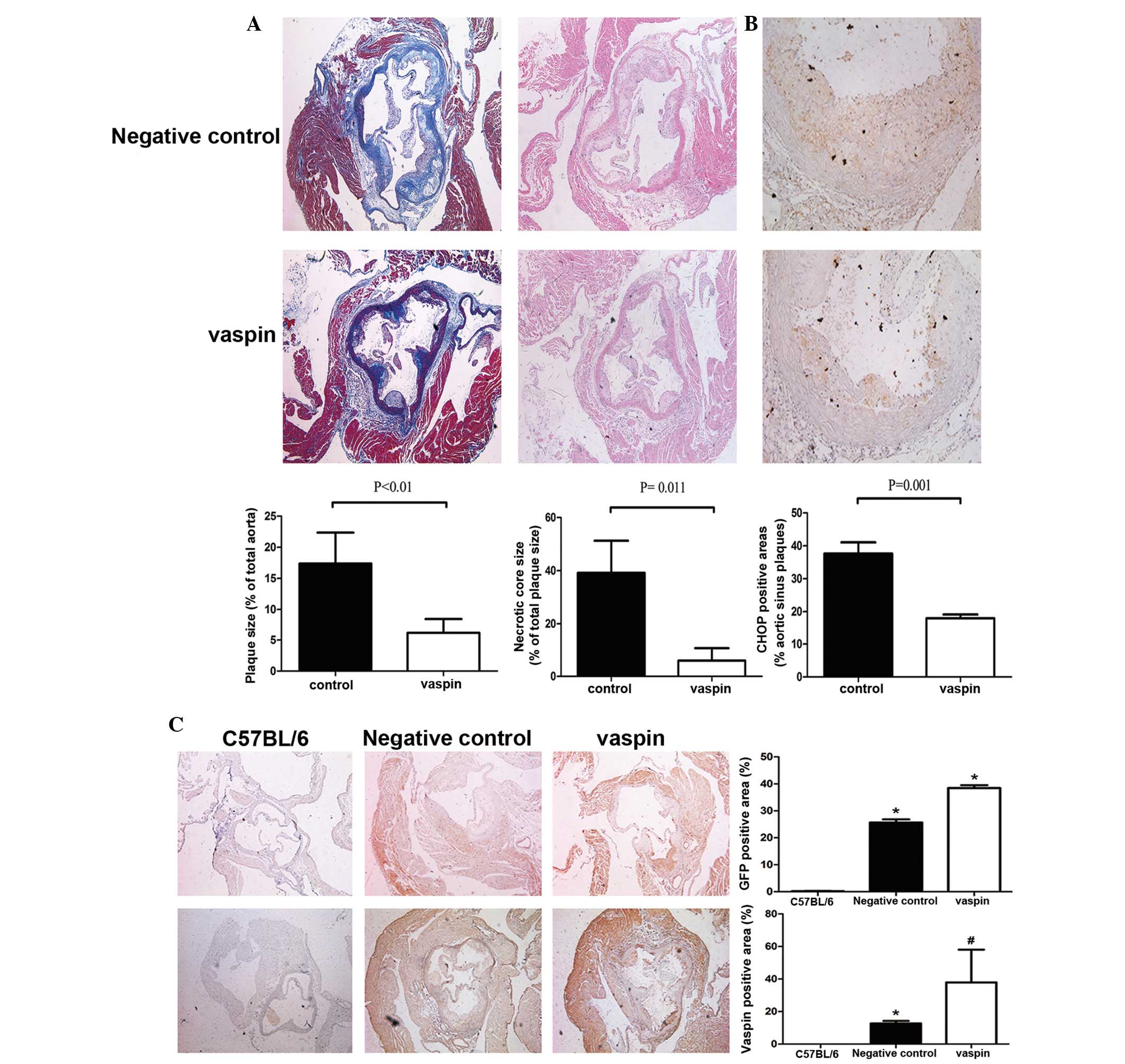
To confirm that the apoE−/− mice were
successfully transfected with the lentiviruses, immunohistochemical
staining of GFP and vaspin was conducted in the aortic sinus of the
C57BL/6 mice, LV5NC-transfected apoE−/− mice (negative
control), and LV5-mus-vaspin-transfected mice. As shown in Fig. 2C, the expression of GFP was higher
in the LV5-mus-vaspin-transfected apoE−/− mice, as
compared with the C57BL/6 mice. In addition, the expression of
vaspin was increased in the LV5-mus-vaspin-transfected
apoE−/− mice, as compared with the LV5NC-transfected
group and C57BL/6 mice.
Vaspin decreases atherosclerotic plaque
size and attenuates ER stress in apoE−/− mice
Masson's trichrome staining was used to evaluate
plaque stabilization. Vaspin markedly enhanced collagen content,
whereas the lipid-rich necrotic core size was significantly
decreased following vaspin transfection (P<0.01; Fig. 2A). Considering other aspects of
plaque composition, an immunohistochemical analysis detected
reduced CHOP expression in the aortic sinus of
LV5-mus-vaspin-transfected apoE−/− mice (P<0.001;
Fig. 2B).
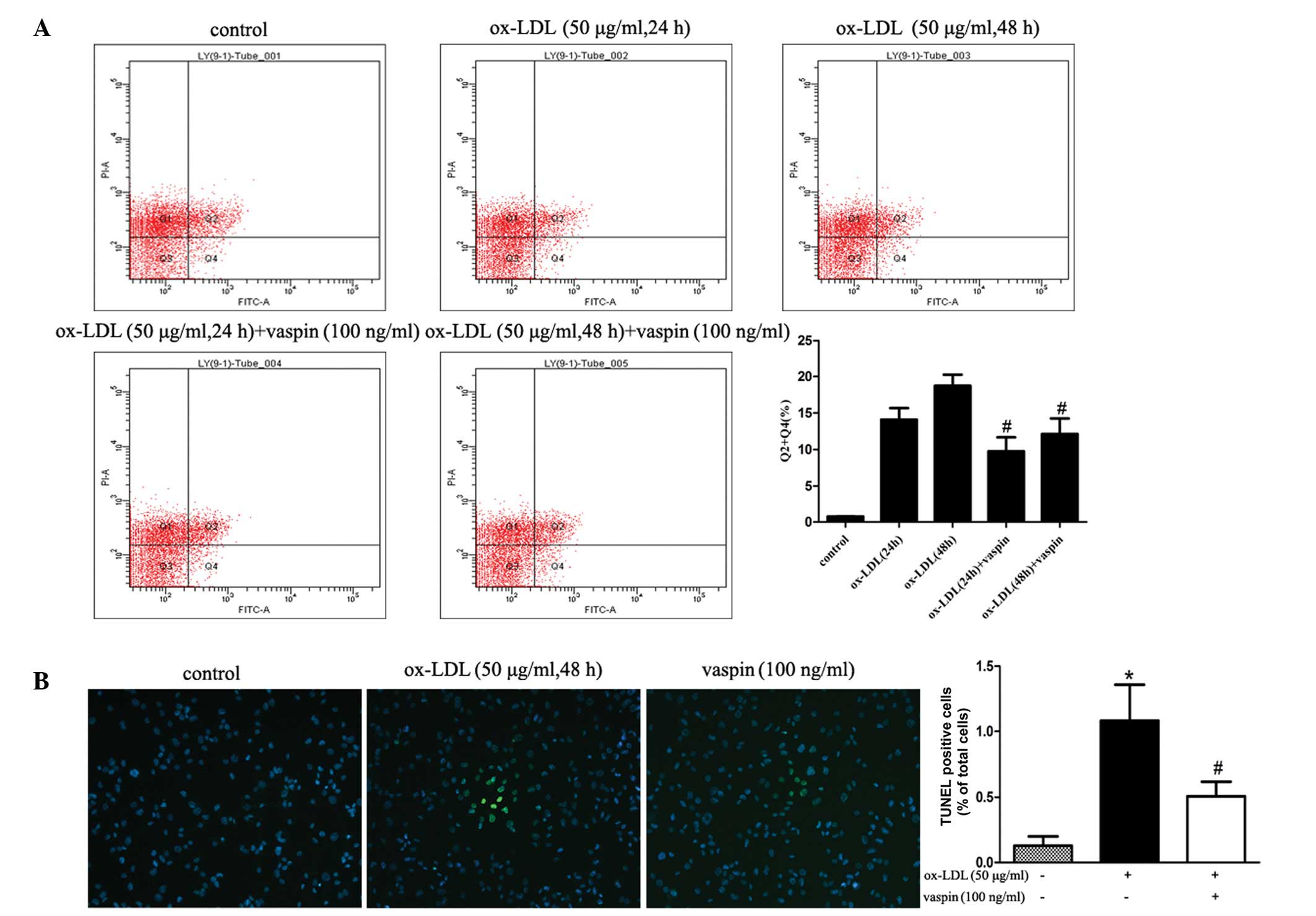
Vaspin inhibits ER stress-induced
apoptosis in macrophages
To confirm the inhibitory effects of vaspin on ER
stress-induced apoptosis, Annexin V/propidium iodide dual staining
and FCM, and TUNEL analyses were performed. Representative images
from the FCM assays and summarized data are shown in Fig. 3A. As compared with the control
group, ox-LDL over 24 and 48 h significantly increased rates of
apoptosis (0.73±0.06 vs. 14.10±1.57%; P<0.001; 0.73±0.06 vs.
18.73±1.53%; P<0.001, respectively). Pre-treatment with vaspin
(100 ng/ml) significantly reduced the percentage of late and early
apoptotic macrophages (14.10±1.57 vs. 9.77±1.90% and 18.73±1.53 vs.
12.13±2.12%, respectively; P=0.038 and P=0.012). In addition,
pretreatment with vaspin decreased the percentage of TUNEL-positive
cells (Fig. 3B).
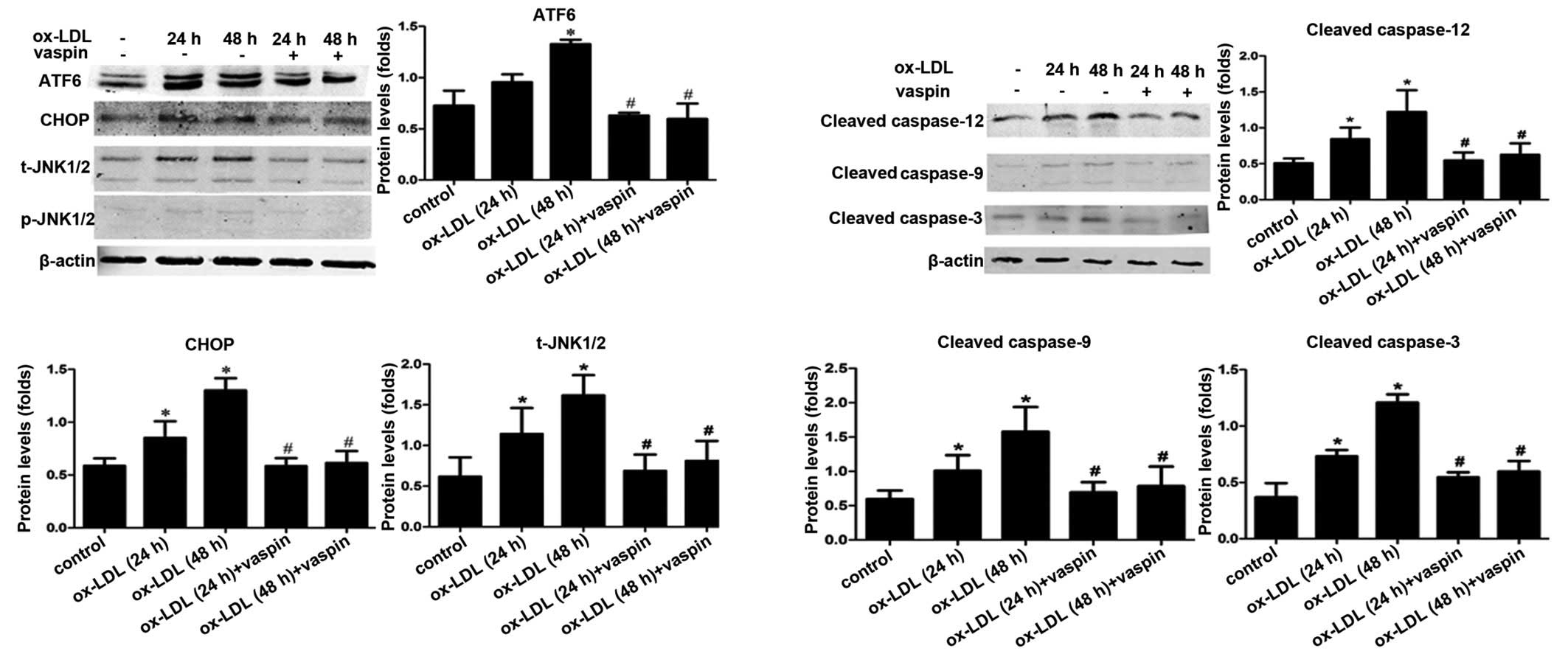
Vaspin downregulates the expression of
proteins associated with ER stress-induced apoptosis in
macrophages
As shown in Fig. 4A
and B, ATF6, CHOP, t-JNK1/2, cleaved-caspase 12, cleaved-caspase 9
and cleaved-caspase 3 were overexpressed in response to ox-LDL (50
µg/ml) incubation but were markedly suppressed by vaspin
pretreatment (100 ng/ml). Western blotting was performed to confirm
the that vaspin inhibited the progression of atherosclerotic
plaques via downregulation of proteins associated with ER
stress-induced apoptosis.
Discussion
The results of the present study demonstrated that
vaspin was able to inhibit the progression of atherosclerotic
plaques in apoE−/− mice. The underlying protective
mechanism was partly associated with the inhibition of ER
stress-induced macrophage apoptosis. Vaspin is well known for its
ability to improve insulin sensitivity (1,2).
Numerous studies regarding vaspin, including our previous study,
have demonstrated that vaspin exerts anti-inflammatory and
beneficial effects in cardiovascular disease, in addition to its
protective role in metabolic disease (10,12,22).
As well as our previous study (10), Phalitakul et al (23) reported that vaspin has an
inhibitory role on VSMC inflammation by affecting inflammatory
signaling pathways. In addition, Jung et al demonstrated
that vaspin may protect vascular endothelial cells against free
fatty acid-induced apoptosis via upregulation of the
phosphoinositide 3-kinase/Akt signaling pathway (9).
The results of the present study demonstrated that
vaspin was able to inhibit ER stress-induced macrophage apoptosis.
In addition, vaspin inhibited ER stress and apoptosis in
vivo, as detected by reductions in the expression levels of
CHOP in the aortic sinus following injection with a vaspin-encoding
lenti-virus. Notably, total cholesterol, but not serum glucose or
lipid levels, was significantly altered by vaspin, thus suggesting
that the inhibitory effects of vaspin on lesion growth may be
largely independent of lipid profile changes. However, the
mechanisms underlying the antiapoptotic effects of vaspin on
macrophages remain unclear. Nakatsuka et al (11,12)
reported that vaspin is a ligand for the cell surface-associated
GRP78/MTJ-1 complex in the liver, and is a novel ligand for the
cell surface GRP78/VDAC complex in endothelial cells, promoting
proliferation, inhibiting apoptosis, and protecting against
diabetes-induced vascular injury. The function of GRP78 is largely
associated with the ER of cells. Nakatsuka et al was first
to clarify the mechanisms underlying the antiapoptotic action of
vaspin in vascular endothelial cells, and implied that vaspin may
have a positive role against ER stress.
It is well known that prolonged ER stress leads to
cell death via apoptosis, and numerous apoptotic pathways have been
identified. One apoptotic pathway is associated with the
transcriptional induction of CHOP, which is a member of the C/EBP
family of transcription factors (24) that can be induced by ATF6 (16). Under normal conditions, CHOP is
either not expressed or expressed at low levels; however, it is
markedly activated in response to ER stress (25). CHOP overexpression can induce
apoptosis, and a previous study demonstrated that CHOP expression
within atherosclerotic plaques exhibited a strong correlation with
the stage of coronary artery lesion, with both parameters markedly
increasing in plaques with vulnerable morphology (26). Another pathway is associated with
the activation of JNK, which comprises a family of signal
transduction proteins that regulate gene expression and participate
in decisions regarding apoptosis and survival in response to stress
(24). The results of the present
study demonstrated that the expression levels of ATF6, CHOP and
JNK1/2 were significantly inhibited by vaspin, and CHOP expression
and necrotic area were decreased in the atherosclerotic plaques of
vaspin-transfected apoE−/− mice. These results confirmed
that vaspin was able to inhibit the progression of atherosclerosis
by suppressing ER stress-induced macrophage apoptosis in
apoE−/− mice.
Zhou et al demonstrated that macrophages
accumulated in atherosclerotic lesions undergoing ER stress, and
CHOP expression was increased during the progression of
atherosclerosis in chow-fed or Western diet-fed apoE−/−
mice (27). These data indicated
that CHOP expression increases as lesions progress, a concept that
has been expanded to human atherosclerosis by Myoishi et al
(26). In addition, a previous
study reported that CHOP deficiency suppressed atherosclerotic
progression in apoE−/− mice, which is consistent with
the results of the present study, and confirmed that the mechanism
underlying atherosclerotic inhibition via CHOP deficiency is
associated with decreased atherosclerotic plaque necrosis and
apoptosis of macrophages (28).
CHOP-mediated macrophage apoptosis also contributes to the
instability of atherosclerotic plaques, thus suggesting the
importance of CHOP in macrophages (29).
ER stress may also lead to apoptosis via activation
of caspase 12 (24). Caspase 12 is
specifically activated in cells undergoing ER stress. Furthermore,
caspase 12-deficient cells are resistant to ER stress inducers,
suggesting that caspase-12 is significant in ER stress-induced
apoptosis (30). Upon ER stress,
procaspase 12 is cleaved and activated, which in turn activates
caspase 9/3, thereby leading to cell death (31). The present study demonstrated that
the expression levels of caspase 12, caspase 9 and caspase 3 were
decreased by vaspin pretreatment. Furthermore, vaspin suppressed ER
stress-induced apoptosis, as determined by FCM analysis and TUNEL
assay.
In conclusion, the results of the present study
indicated that vaspin was able to inhibit the progression of
atherosclerotic plaques in apoE−/− mice. The potential
underlying mechanism is partly associated with the inhibition of ER
stress-induced macrophage apoptosis. These findings confirmed that
vaspin may have a beneficial role in ameliorating the progression
of atherosclerosis, and provide novel insights into the protective
function of vaspin in atherosclerosis, thus suggesting that vaspin
may be potentially useful for preventing vascular, as well as
metabolic, diseases.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant no. 81270256) to
Professor Yawei Xu.
References
|
1
|
Hida K, Wada J, Eguchi J, Zhang H, Baba M,
Seida A, Hashimoto I, Okada T, Yasuhara A, Nakatsuka A, et al:
Visceral adipose tissue-derived serine protease inhibitor: A unique
insulin-sensitizing adipocytokine in obesity. Proc Natl Acad Sci
USA. 102:10610–10615. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wada J: Vaspin: A novel serpin with
insulin-sensitizing effects. Expert Opin Investig Drugs.
17:327–333. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chang HM, Lee HJ, Park HS, Kang JH, Kim
KS, Song YS and Jang YJ: Effects of weight reduction on serum
vaspin concentrations in obese subjects: Modification by insulin
resistance. Obesity (Silver Spring). 18:2105–2110. 2010. View Article : Google Scholar
|
|
4
|
Hida K, Poulsen P, Teshigawara S, Nilsson
E, Friedrichsen M, Ribel-Madsen R, Grunnet L, Lund SS, Wada J and
Vaag A: Impact of circulating vaspin levels on metabolic variables
in elderly twins. Diabetologia. 55:530–532. 2012. View Article : Google Scholar
|
|
5
|
Youn BS, Klöting N, Kratzsch J, Lee N,
Park JW, Song ES, Ruschke K, Oberbach A, Fasshauer M, Stumvoll M
and Blüher M: Serum vaspin concentrations in human obesity and type
2 diabetes. Diabetes. 57:372–377. 2008. View Article : Google Scholar
|
|
6
|
Tan BK, Heutling D, Chen J, Farhatullah S,
Adya R, Keay SD, Kennedy CR, Lehnert H and Randeva HS: Metformin
decreases the adipokine vaspin in overweight women with polycystic
ovary syndrome concomitant with improvement in insulin sensitivity
and a decrease in insulin resistance. Diabetes. 57:1501–1507. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Seeger J, Ziegelmeier M, Bachmann A,
Lössner U, Kratzsch J, Blüher M, Stumvoll M and Fasshauer M: Serum
levels of the adipokine vaspin in relation to metabolic and renal
parameters. J Clin Endocrinol Metab. 93:247–251. 2008. View Article : Google Scholar
|
|
8
|
Li HL, Peng WH, Cui ST, Lei H, Wei YD, Li
WM and Xu YW: Vaspin plasma concentrations and mRNA expressions in
patients with stable and unstable angina pectoris. Clin Chem Lab
Med. 49:1547–1554. 2011.PubMed/NCBI
|
|
9
|
Jung CH, Lee WJ, Hwang JY, Seol SM, Kim
YM, Lee YL and Park JY: Vaspin protects vascular endothelial cells
against free fatty acid-induced apoptosis through a
phosphatidylinositol 3-kinase/Akt pathway. Biochem Biophys Res
Commun. 413:264–269. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li H, Peng W, Zhuang J, Lu Y, Jian W, Wei
Y, Li W and Xu Y: Vaspin attenuates high glucose-induced vascular
smooth muscle cells proliferation and chemokinesis by inhibiting
the MAPK, PI3K/Akt and NF-κB signaling pathways. Atherosclerosis.
228:61–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nakatsuka A, Wada J, Iseda I, Teshigawara
S, Higashio K, Murakami K, Kanzaki M, Inoue K, Terami T, Katayama
A, et al: Vaspin is an adipokine ameliorating ER stress in obesity
as a ligand for cell-surface GRP78/MTJ-1 complex. Diabetes.
61:2823–2832. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nakatsuka A, Wada J, Iseda I, Teshigawara
S, Higashio K, Murakami K, Kanzaki M, Inoue K, Terami T, Katayama
A, et al: Visceral adipose tissue-derived serine proteinase
inhibitor inhibits apoptosis of endothelial cells as a ligand for
the cell-surface GRP78/voltage-dependent anion channel complex.
Circ Res. 112:771–780. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Seimon T and Tabas I: Mechanisms and
consequences of macrophage apoptosis in atherosclerosis. J Lipid
Res. 50(Suppl): S382–S387. 2009. View Article : Google Scholar :
|
|
14
|
Andrés V, Pello OM and Silvestre-Roig C:
Macrophage proliferation and apoptosis in atherosclerosis. Curr
Opin Lipidol. 23:429–438. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hotamisligil GS: Endoplasmic reticulum
stress and the inflammatory basis of metabolic disease. Cell.
140:900–917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Scull CM and Tabas I: Mechanisms of ER
stress-induced apoptosis in atherosclerosis. Arterioscler Thromb
Vasc Biol. 31:2792–2797. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Aust G, Richter O, Rohm S, Kerner C, Hauss
J, Klöting N, Ruschke K, Kovacs P, Youn BS and Blüher M: Vaspin
serum concentrations in patients with carotid stenosis.
Atherosclerosis. 204:262–266. 2009. View Article : Google Scholar
|
|
18
|
Kadoglou NP, Gkontopoulos A, Kapelouzou A,
Fotiadis G, Theofilogiannakos EK, Kottas G and Lampropoulos S:
Serum levels of vaspin and visfatin in patients with coronary
artery disease-Kozani study. Clin Chim Acta. 412:48–52. 2011.
View Article : Google Scholar
|
|
19
|
Zhang B, Peng W, Li H, Lu Y, Zhuang J,
Wang K, Su Y and Xu Y: Plasma vaspin concentrations are decreased
in acute coronary syndrome, but unchanged in patients without
coronary lesions. Clin Biochem. 46:1520–1525. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals. 8th
edition. National Institutes of Health, The National Academies
Press; Washington (DC): 2011
|
|
21
|
Zhuang J, Peng W, Li H, Lu Y, Wang K, Fan
F, Li S and Xu Y: Inhibitory effects of vinpocetine on the
progression of atherosclerosis are mediated by Akt/NF-kB dependent
mechanisms in apoE−/− mice. PloS One. 8:e825092013.
View Article : Google Scholar
|
|
22
|
Phalitakul S, Okada M, Hara Y and Yamawaki
H: A novel adipocytokine, vaspin inhibits platelet-derived growth
factor-BB-induced migration of vascular smooth muscle cells.
Biochem Biophys Res Commun. 423:844–849. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Phalitakul S, Okada M, Hara Y and Yamawaki
H: Vaspin prevents TNF-α-induced intracellular adhesion molecule-1
via inhibiting reactive oxygen species-dependent NF-κB and PKCθ
activation in cultured rat vascular smooth muscle cells. Pharmacol
Res. 64:493–500. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Oyadomari S, Araki E and Mori M:
Endoplasmic reticulum stress-mediated apoptosis in pancreatic
beta-cells. Apoptosis. 7:335–345. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang XZ, Lawson B, Brewer JW, Zinszner H,
Sanjay A, Mi LJ, Boorstein R, Kreibich G, Hendershot LM and Ron D:
Signals from the stressed endoplasmic reticulum induce
C/EBP-homologous protein (CHOP/GADD153). Mol Cell Biol.
16:4273–4280. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Myoishi M, Hao H, Minamino T, Watanabe K,
Nishihira K, Hatakeyama K, Asada Y, Okada K, Ishibashi-Ueda H,
Gabbiani G, et al: Increased endoplasmic reticulum stress in
atherosclerotic plaques associated with acute coronary syndrome.
Circulation. 116:1226–1233. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou J, Lhoták S, Hilditch BA and Austin
RC: Activation of the unfolded protein response occurs at all
stages of athero-sclerotic lesion development in apolipoprotein
E-deficient mice. Circulation. 111:1814–1821. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Thorp E, Li G, Seimon TA, Kuriakose G, Ron
D and Tabas I: Reduced apoptosis and plaque necrosis in advanced
athero-sclerotic lesions of Apoe−/− and
Ldlr−/− mice lacking CHOP. Cell Metab. 9:474–481. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tsukano H, Gotoh T, Endo M, Miyata K,
Tazume H, Kadomatsu T, Yano M, Iwawaki T, Kohno K, Araki K, et al:
The endoplasmic reticulum stress-C/EBP homologous protein
pathway-mediated apoptosis in macrophages contributes to the
instability of atherosclerotic plaques. Arterioscler Thromb Vasc
Biol. 30:1925–1932. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Morishima N, Nakanishi K, Takenouchi H,
Shibata T and Yasuhiko Y: An endoplasmic reticulum stress-specific
caspase cascade in apoptosis. Cytochrome c-independent activation
of caspase-9 by caspase-12. J Biol Chem. 277:34287–34294. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Minamino T, Komuro I and Kitakaze M:
Endoplasmic reticulum stress as a therapeutic target in
cardiovascular disease. Circ Res. 107:1071–1082. 2010. View Article : Google Scholar : PubMed/NCBI
|