Introduction
Prostate cancer is one of the most common
malignancies among men and is the second leading cause of
cancer-associated mortality in males (1). In 2014, prostate cancer accounted for
an estimate of 27% of newly diagnosed cancer cases and 10% of
cancer-associated mortalities in men in the USA (2). Since prostate cancer is
organ-confined in most cases at the time-point of diagnosis,
radical prostatectomy and definitive radiotherapy are the current
standard treatment for the vast majority of prostate cancer cases
(3); however, a sub-set of
patients present with disease progression, including
metastasization to other organs or tissues in the body, such as
bones or bladder (4). A widely
accepted treatment option for such patients is androgen-deprivation
therapy, following which, however, prostate cancer recurrence in
its androgen-independent or hormone-refractory type is common
(5). While the accurate etiology
of prostate cancer is complex, several studies have suggested that
it may be associated with factors including smoking, environment,
dietary habits, endocrine system, age and ethnicity (6–9). In
the clinic, the prevention and treatment of prostate cancer and
associated metastasis have remained challenging, as the underlying
molecular mechanisms have remained to be fully elucidated (1). Therefore, it is urgent to discover
novel therapeutic targets and approaches to suppress prostate
cancer metastasis.
Certain microRNAs (miRs) have been found to be
correlated with the clinical outcome of specific cancer types
(10). miRs are a class of
naturally occurring, endogenous small non-coding RNAs, with a size
of 19–22 nt (11). They
specifically bind the 3′-untranslated regions (3′UTR) of their
target messenger RNAs (mRNAs) to induce their degradation or
inhibit their translation, causing partial or total silencing of
the respective protein-coding gene (12). miRs are thought to regulate
two-thirds of the entire protein-encoding genes (13). The expression of miRs themselves
can also be regulated similarly to that of protein-coding genes,
either through genetic or epigenetic shifts. miR expression is
frequently aberrant in various cancer types (14). These alterations have been shown to
be accountable for almost all facets of cancer development and
progression (15). Upregulated
miRs in cancer may function as oncogenes by negatively regulating
tumor suppressors, while, downregulated miRs may function as tumor
suppressor genes in healthy cells and inhibit cancer by regulating
oncogenes (16,17).
miR-497 was found to be significantly downregulated
in prostate cancer, and its restoration was shown to have
anti-proliferative and apoptotic effects (18). Based on these findings, the present
study assessed the effects of miR-497 restoration on the migration
and invasion of prostate cancer cells as processes associated with
metastasis, and identified the underlying molecular mechanism.
Hepatoma-derived growth factor (HDGF), which was indicated to be a
putative target of miR-497, was confirmed using a luciferase
reporter assay. The present study found that miR-497 represses the
migratory and invasive capacity of the DU145 and PC-3 prostate
cancer cells by downregulating HDGF.
Materials and methods
Cell culture and transfection
The human prostate cancer cell lines DU145 and PC-3
were purchased from the Shanghai Institute of Biochemistry and Cell
Biology (Shanghai, China) and cultured in RPMI-1640 medium
supplemented with 10% heat-inactivated fetal bovine serum (FBS;
Gibco, Thermo Fisher Scientific, Waltham, MA, USA) in a humidified
atmosphere containing 5% CO2 at 37°C.
Mature miR-497 mimics (5′-CAG CAG CAC ACU GUG GUU
UGU-3′) and negative control (NC) miRNA mimics (5′-UUC UCC GAA CGU
GUC ACG UTT-3′) were designed and synthesized by GenePharma
(Shanghai, China). When the density of DU145 and PC-3 cells reached
30–40%, cells were transfected using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific), according to the
manufacturer's instructions.
Cell migration and invasion assays
The cell migration and invasion assays were
performed using Transwell chambers (8 µm; Costar; Thermo
Fisher Scientific). For the migration assay, 1×105
transfected cells (miR-497 mimics and NC) were placed into the
upper chamber cultured in medium with 2% FBS, while 500 µl
medium containing 20% FBS was added to the lower chamber. For the
invasion assays, 1×105 transfected cells (miR-497 mimics
and NC) cultured in medium with 2% FBS were placed into the upper
chamber pre-coated with Matrigel (BD Biosciences, Franklin Lakes,
NJ, USA), while 500 µl medium containing 20% FBS was added
to the lower chamber. Cells were incubated for 12 h for the
migration assay and 24 h for the invasion assay. At the end of the
experiments, the cells that had not migrated or invaded through the
pores were carefully removed. The filters were then fixed with 100%
methanol (Beyotime Institute of Biotechnology, Inc., Haimen, China)
for 2 min, stained in 0.5% crystal violet (Beyotime Institute of
Biotechnology, Inc.) for 2 min, rinsed in phosphate-buffered saline
and then subjected to microscopic inspection (magnification, ×200).
Five random fields were counted per chamber using an inverted
microscope (CKX41; Olympus Corp., Tokyo, Japan). Each condition was
assayed in triplicate and each experiment was repeated at least
three times.
Western blot analysis
Primary antibodies used in the present study,
including HDGF and β-actin were products of Cell Signaling
Technology, Inc. (Danvers, MA, USA). DU145 or PC-3 cells were
seeded in six-well plates, and 72 h after transfection with miR-497
mimics or NC, cells were harvested and homogenized with
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Inc.). The protein concentration was measured using
the bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology, Inc.). Equal amounts of protein were separated using
10% sodium dodecyl sulfate polyacrylamide electrophoresis (Beyotime
Institute of Biotechnology, Inc.) gel and then transferred onto
polyvinylidene difluoride membranes (Beyotime Institute of
Biotechnology, Inc.), which were blocked with 5% non-fat dry milk
for 2 h and incubated with primary antibodies, including mouse
anti-human monoclonal HDGF (cat. no. sc-271344; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) and mouse anti-human
monoclonal β-actin (cat. no. 3700; Cell Signaling Technology,
Inc.). The membrane was washed and then incubated for 1 h with the
corresponding horseradish peroxidase-conjugated secondary antibody
(sc-2005; Santa Cruz Biotechnology, Inc., CA, USA) in Tris-buffered
saline with Tween 20 (Beyotime Institute of Biotechnology, Inc.).
Finally, blots were visualized using enhanced chemiluminescence
solution (Pierce, Rockford, IL, USA) and images were captured using
the FluorChem imaging system (Alpha Innotech, San Leandro, CA,
USA).
Luciferase reporter assay
A search with TargetScan (http://www.targetscan.org) predicted HDGF as a direct
target gene of miR-497. A seven-nucleotide seed match of position
37-43 of the HDGF 3′-UTR with miR-497 was revealed. In order to
verify this interaction, a luciferase reporter assay was performed.
Cells were plated in a 12-well plate at ~90% confluence and
transfected with miR-497 mimics or NC using Lipofectamine 2000
(Invitrogen). For the reporter assays, stably
miR-497-overexpressing cells were transiently transfected with
reporter plasmids driven by a wild-type or mutated fragment from
the 3′UTR of HDGF. The luciferase reporter vectors, including
HDGF-3′UTR Wt and HDGF-′UTR Mut, were obtained from GenePharma Co.,
Ltd. (Shanghai, China). Each sample was also co-transfected with
0.05 µg pRL-CMV plasmid expressing Renilla Luciferase
(Promega Corp., Madison, WI, USA) as an internal control for
transfection efficiency. After 48 h of incubation, the cells were
harvested and lysed, and luciferase activity was measured using the
Dual-Luciferase Reporter Assay System (Promega Corp.) according to
the manufacturer's instructions. Firefly luciferase activity was
normalized to Renilla luciferase activity for each
transfected well. Each assay was replicated three times.
Statistical analysis
Values are expressed as the mean ± standard
deviation. Comparison between groups was performed using Student's
t-test. All statistical analyses were performed using Stata
10.0 (College Station, Texas, USA). A double-tailed P-value of
<0.05 was considered to indicate a statistically significant
difference between values.
Results
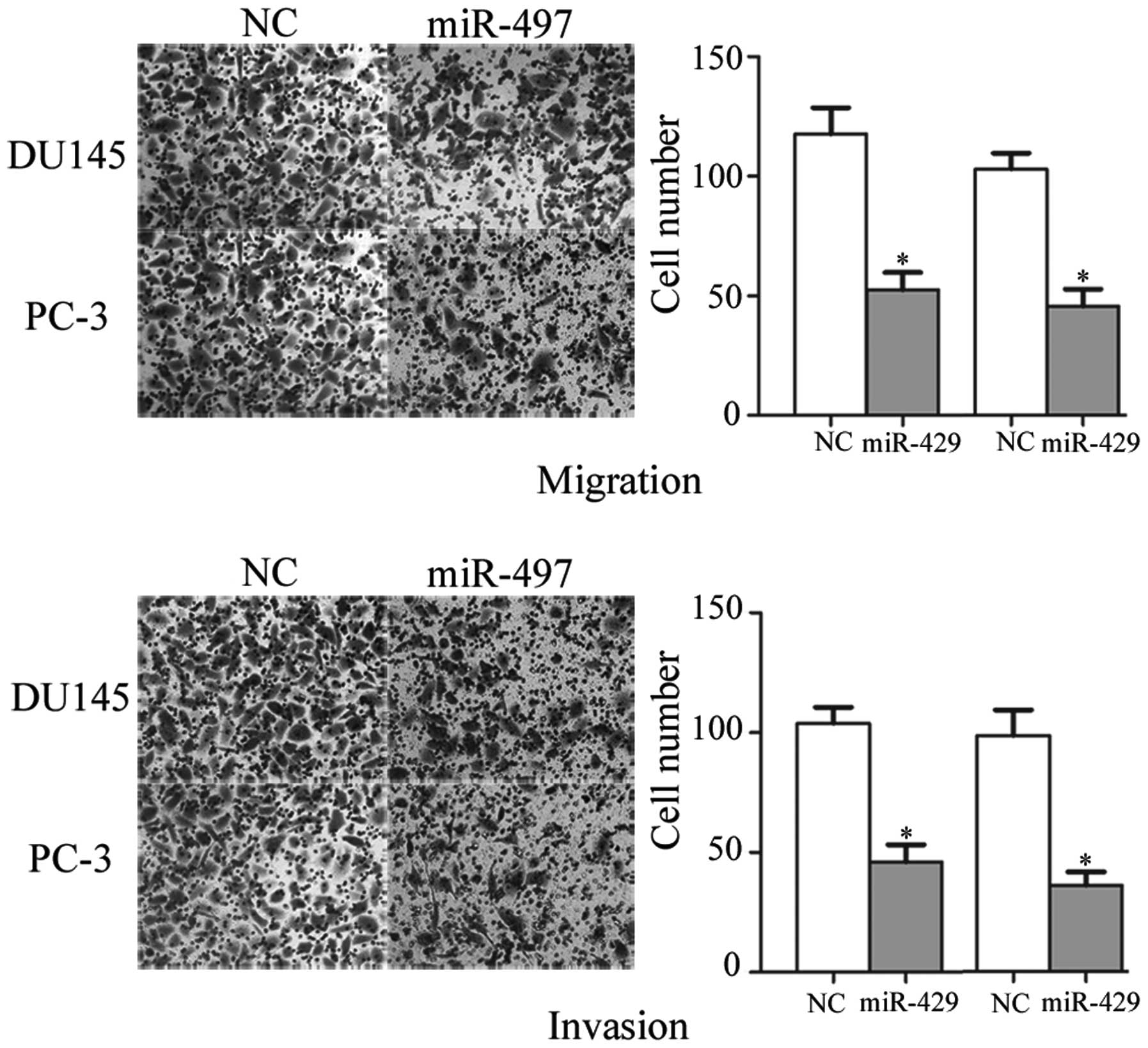
miR-497 inhibits cell migration and
invasion of prostate cancer cell lines
To determine whether miR-497 regulates human
prostate cancer cell migration and invasion, the DU145 and PC-3
cells were transfected with miR-497 and NC mimics and subjected to
Transwell assays. As expected, overexpression of miR-497
significantly decreased the migratory and invasive capacity of
DU145 and PC-3 cells (Fig. 1).
These observations indicated that miR-497 was a negative regulator
of prostate cancer cell migration and invasion.
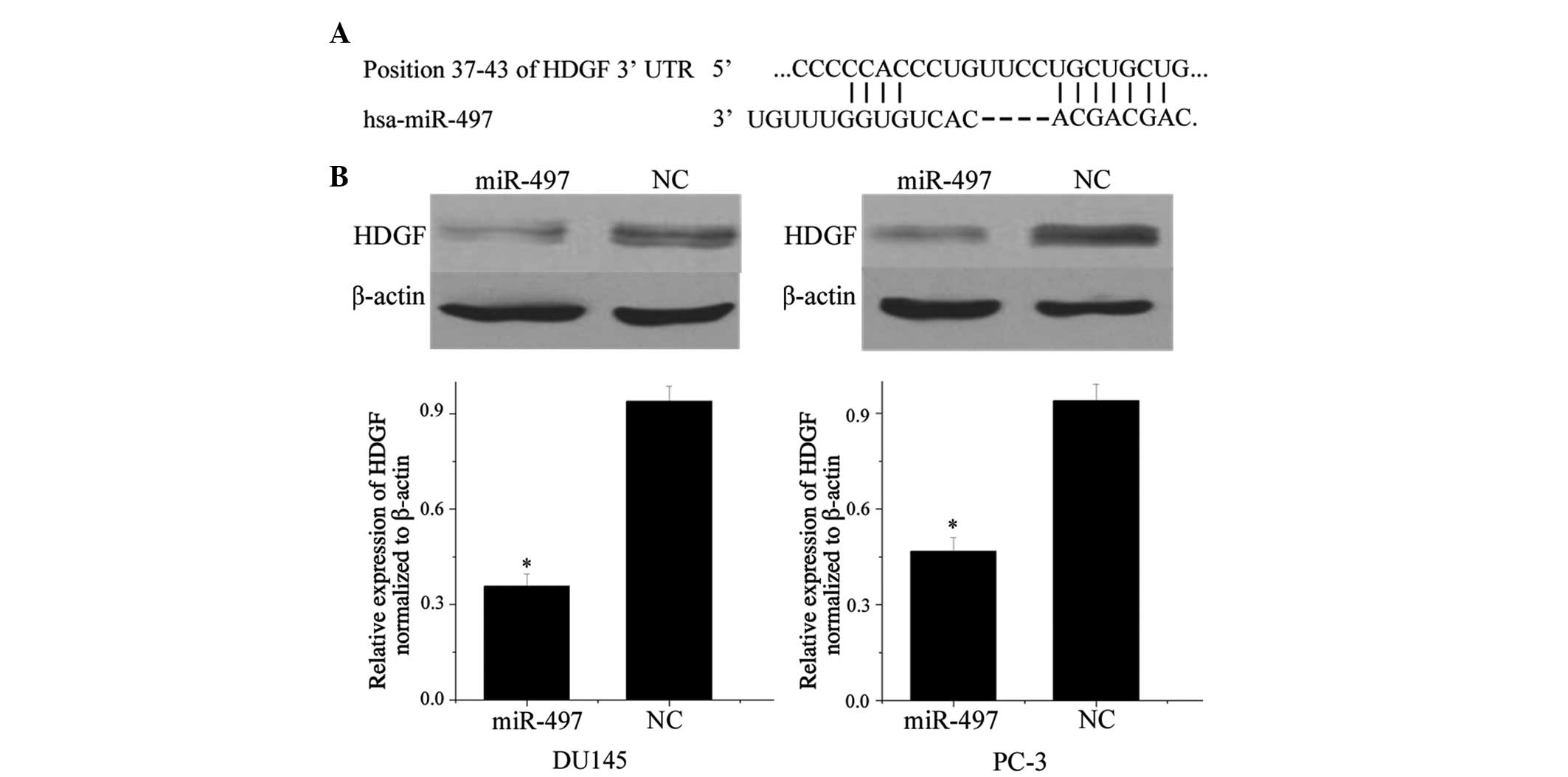
miR-497 suppresses the expression of HDGF
in prostate cancer cell lines
A search with TargetScan predicted that HDGF is a
direct target gene of miR-497. As illustrated in Fig. 2A, position 37-43 of the HDGF 3′-UTR
represents a seven-nucleotide seed match with miR-497.
Western blot analysis was then performed to explore
whether miR-497 affected the expression of HDGF in prostate cancer
cell lines. As shown in Fig. 2B,
HDGF was significantly down-regulated in the DU145 and PC-3
prostate cancer cell lines after transfection with miR-497
(P<0.05). These results revealed that miR-497 suppressed the
protein expression of HDGF in prostate cancer cell lines.
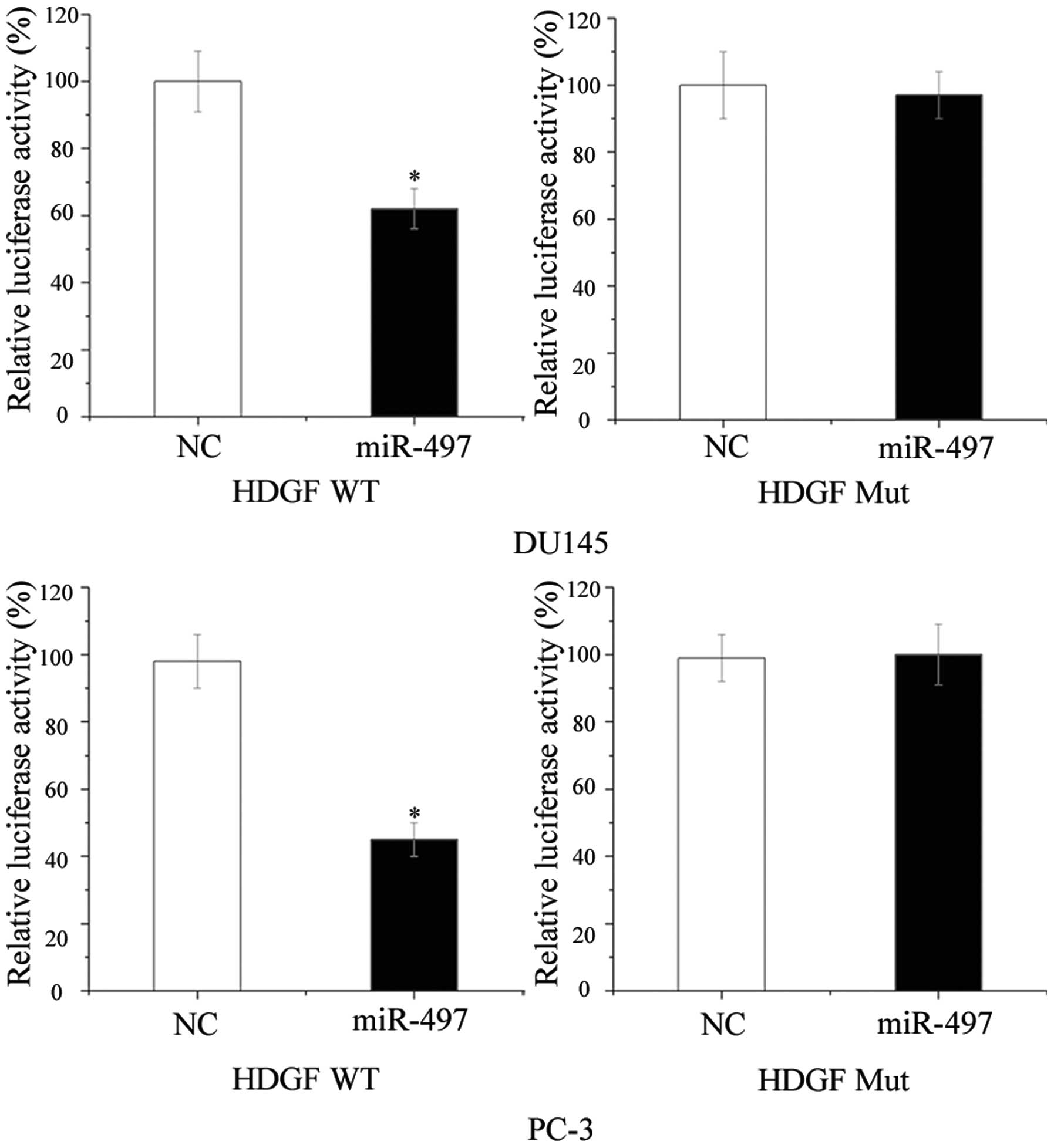
HDGF is a direct target gene of miR-497
in prostate cancer
To further confirm that HDGF is a direct target of
miR-497, DU145 and PC-3 cells were subjected to luciferase reporter
assays. As illustrated in Fig. 3,
transfection with miR-497 significantly decreased the activity of
the HDGF 3′-UTR luciferase reporter compared with that in the
NC-transfected group (P<0.05), while the activity of the
reporter containing the mutated sequence was not affected. These
results confirmed in vitro that HDGF was a target gene of
miR-497 in prostate cancer cells.
Discussion
Metastasis is a major event associated with tumor
recurrence and poor patient outcome (19). miRs are a group of small,
non-coding RNAs that are commonly dysregulated in human malignant
cancers (20). The involvement of
miRs in the development of tumor metastasis has provided a novel
perspective on the tumorigenic process (21). The present study revealed that
miR-497 may function as a tumor suppressor through repression of
HDGF in prostate cancer. Restoration of miR-497 suppressed cell
migration and invasion in prostate cancer cells.
miR-195 and miR-497 are encoded by the same gene
cluster located on chromosomal band 17p13.1, and due to their
sequence AGCAGC starting at either the first nucleotide or the
second nucleotide from the 5′ end of the mature (~22 nt, single
stranded) miRNA, they are members of the miR-15/107 gene group
(22). Expression of miR-497 has
been proved to be frequently reduced in various tumors, including
breast cancer (23),
hepatocellular carcinoma (24),
gastric cancer (25),
nasopharyngeal carcinoma (26),
endometrial cancer (27),
colorectal cancer (28) and
prostate cancer (18). In breast
cancer, it has been demonstrated that miR-497 markedly inhibits
cellular growth, induces apoptosis and disrupts the cell cycle by
targeting Bcl-w; furthermore, miR-497 expression was negatively
correlated with the pathological stage, lymphatic metastasis, tumor
size and HER-2 (29). In
hepatocellular carcinoma, it was demonstrated that CHEK1 is a
target gene of miR-497 (24).
Identification of the target genes of miR-497 is
crucial for understanding its role in tumorigenesis and for the
discovery of novel therapeutic targets. Studies have certified that
in human cells, miR-497 regulates the expression of certain
oncogenes, including Bcl-w, ERK1, MEK1, IGF-1R, CHEK1 and MAPK
(24,28–31).
Therefore, upregulation of miR-497 or administration of
pharmaceutical compounds with analogous effects is expected to
represent an effective therapeutic strategy for tumors resulting
from activation or overexpression of these oncogenes. The present
study demonstrated the tumor suppressive effects of miR-497 in
DU145 and PC-3 cell lines, which included the inhibition of cell
migration and invasion by downregulation of HDGF expression. The
findings of the present study suggested that miR-497 may represent
a novel molecular marker and therapeutic agent to inhibit
metastasis of prostate cancer.
HDGF is a novel growth factor with acidic
heparin-binding ability and was first isolated from the cultured
media of the HuH-7 human hepatoma cell line (32). It consists of 240 amino acids with
functional domains, including a conserved N-terminal 100-amino-acid
domain responsible for heparin and DNA binding, and two bipartite
nuclear localization sequences (33). Studies have revealed that HDGF is
expressed in most organs (i.e. liver, brain, lung and intestine)
during embryonic development as well as in mature organisms
(34). Besides its mitogenic
activity, HDGF is implicated in organ development and tissue
differentiation (35). HDGF has
been found to be highly expressed in non-small cellular lung cancer
(36), hepatocellular carcinoma
(37), colorectal carcinoma
(38), oesophagal carcinoma
(39), pancreatic carcinoma
(40) and melanoma (41). In addition, high levels of HDGF are
associated with poor prognosis in several types of cancer,
including non-small cell lung cancer, hepatocellular carcinoma,
esophageal cancer, gastric cancer and gastrointestinal stromal
tumors (33). Therefore, HDGF
represents a potential prognostic factor for these cancers
types.
The effects of HDGF overexpression in carcinogenesis
have become a main focus in cancer biology and have been studied in
various types of cancer. Previous studies have indicated that HDGF
participates in numerous cellular processes, including astrocyte
proliferation, renal development, vascular lesion formation
(42), sensitization to
irradiation (43), cardiovascular
differentiation as well as cell migration and invasion (44–48).
In prostate cancer, HDGF was shown to be overexpressed and
knockdown of HDGF consistently inhibited proliferation. Moreover,
knockdown of HDGF inhibited the migration and invasion of the DU145
prostate cancer cell line (48).
The results of the present study suggested that miR-497 suppressed
prostate cancer cell migration and invasion through down-regulation
of HDGF. miR-497 may therefore be utilized as a predictive
parameter for early detection of tumor recurrence and as a target
of drugs to block prostate cancer metastasis.
In conclusion, the present study was the first to
show that miR-497 regulates HDGF and regulates cell migration and
invasion of prostate cancer cells. The identification of candidate
target genes of miR-497 may provide an understanding of potential
carcinogenic mechanisms in prostate cancer. miRNA-based therapy is
expected to be more efficient than the traditional single target
therapy, since miRNAs regulate multiple target genes
simultaneously. Therefore, miR-497 may be utilized for the
treatment of prostate cancer.
References
|
1
|
Wu D, Huang P, Wang L, Zhou Y, Pan H and
Qu P: MicroRNA-143 inhibits cell migration and invasion by
targeting matrix metalloproteinase 13 in prostate cancer. Mol Med
Rep. 8:626–630. 2013.PubMed/NCBI
|
|
2
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mangoni M, Desideri I, Detti B, Bonomo P,
Greto D, Paiar F, Simontacchi G, Meattini I, Scoccianti S, Masoni
T, et al: Hypofractionation in prostate cancer: Radiobiological
basis and clinical appliance. Biomed Res Int. 2014:7813402014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lafeuille MH, Gravel J, Grittner A,
Lefebvre P, Ellis L and McKenzie RS: Real-World corticosteroid
utilization patterns in patients with metastatic
castration-resistant prostate cancer in 2 large US administrative
claims databases. Am Health Drug Benefits. 6:307–316. 2013.
|
|
5
|
Nelson CJ, Lee JS, Gamboa MC and Roth AJ:
Cognitive effects of hormone therapy in men with prostate cancer: A
review. Cancer. 113:1097–1106. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Discacciati A and Wolk A: Lifestyle and
dietary factors in prostate cancer prevention. Recent Results
Cancer Res. 202:27–37. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lai GY, Giovannucci EL, Pollak MN, Peskoe
SB, Stampfer MJ, Willett WC and Platz EA: Association of C-peptide
and leptin with prostate cancer incidence in the health
professionals follow-up study. Cancer Causes Control. 25:625–632.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu J and Yu E: Insulin-like growth factor
receptor-1 (IGF-IR) as a target for prostate cancer therapy. Cancer
Metastasis Rev. 33:607–617. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Barba M, McCann SE, Schünemann HJ,
Stranges S, Fuhrman B, De Placido S, Carruba G, Freudenheim JL,
Trevisan M, Russell M, et al: Lifetime total and beverage
specific-alcohol intake and prostate cancer risk: A case-control
study. Nutr J. 3:232004. View Article : Google Scholar
|
|
10
|
Joung JG, Kim D, Lee SY, Kang HJ and Kim
JH: Integrated analysis of microRNA-target interactions with
clinical outcomes for cancers. BMC Med Genomics. 7(Suppl 1):
S102014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Garzon R, Pichiorri F, Palumbo T,
Visentini M, Aqeilan R, Cimmino A, Wang H, Sun H, Volinia S, Alder
H, et al: MicroRNA gene expression during retinoic acid-induced
differentiation of human acute promyelocytic leukemia. Oncogene.
26:4148–4157. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Carthew RW and Sontheimer EJ: Origins and
Mechanisms of miRNAs and siRNAs. Cell. 136:642–655. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar :
|
|
14
|
Kwak PB, Iwasaki S and Tomari Y: The
microRNA pathway and cancer. Cancer Sci. 101:2309–2315. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Oom AL, Humphries BA and Yang C:
MicroRNAs: Novel players in cancer diagnosis and therapies. Biomed
Res Int. 2014:9594612014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang L, Li B, Li L and Wang T:
MicroRNA-497 suppresses proliferation and induces apoptosis in
prostate cancer cells. Asian Pac J Cancer Prev. 14:3499–3502. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Amate P, Huchon C, Dessapt AL, Bensaid C,
Medioni J, Le Frère Belda MA, Bats AS and Lécuru FR: Ovarian
cancer: SItes of recurrence. Int J Gynecol Cancer. 23:1590–1596.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Martignani E, Miretti S, Accornero P and
Baratta M: MiRNAs highlights in stem and cancer cells. Mini Rev Med
Chem. 11:1165–1182. 2011. View Article : Google Scholar
|
|
21
|
Hummel R, Hussey DJ and Haier J:
MicroRNAs: Predictors and modifiers of chemo- and radiotherapy in
different tumour types. Eur J Cancer. 46:298–311. 2010. View Article : Google Scholar
|
|
22
|
Finnerty JR, Wang WX, Hébert SS, Wilfred
BR, Mao G and Nelson PT: The miR-15/107 group of microRNA genes:
Evolutionary biology, cellular functions and roles in human
diseases. J Mol Biol. 402:491–509. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li D, Zhao Y, Liu C, Chen X, Qi Y, Jiang
Y, Zou C, Zhang X, Liu S, Wang X, et al: Analysis of MiR-195 and
MiR-497 expression, regulation and role in breast cancer. Clin
Cancer Res. 17:1722–1730. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xie Y, Wei RR, Huang GL, Zhang MY, Yuan YF
and Wang HY: Checkpoint kinase 1 is negatively regulated by miR-497
in hepa-tocellular carcinoma. Med Oncol. 31:8442014. View Article : Google Scholar
|
|
25
|
Guo J, Miao Y, Xiao B, Huan R, Jiang Z,
Meng D and Wang Y: Differential expression of microRNA species in
human gastric cancer versus non-tumorous tissues. J Gastroenterol
Hepatol. 24:652–657. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen HC, Chen GH, Chen YH, Liao WL, Liu
CY, Chang KP, Chang YS and Chen SJ: MicroRNA deregulation and
pathway alterations in nasopharyngeal carcinoma. Br J Cancer.
100:1002–1011. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hiroki E, Akahira J, Suzuki F, Nagase S,
Ito K, Suzuki T, Sasano H and Yaegashi N: Changes in microRNA
expression levels correlate with clinicopathological features and
prognoses in endometrial serous adenocarcinomas. Cancer Sci.
101:241–249. 2010. View Article : Google Scholar
|
|
28
|
Guo ST, Jiang CC, Wang GP, Li YP, Wang CY,
Guo XY, Yang RH, Feng Y, Wang FH, Tseng HY, et al: MicroRNA-497
targets insulin-like growth factor 1 receptor and has a tumour
suppressive role in human colorectal cancer. Oncogene.
32:1910–1920. 2013. View Article : Google Scholar :
|
|
29
|
Shen L, Li J, Xu L, Ma J, Li H, Xiao X,
Zhao J and Fang L: MiR-497 induces apoptosis of breast cancer cells
by targeting Bcl-w. Exp Ther Med. 3:475–480. 2012.PubMed/NCBI
|
|
30
|
Zheng D, Radziszewska A and Woo P:
MicroRNA 497 modulates interleukin 1 signalling via the MAPK/ERK
pathway. FEBS Lett. 586:4165–4172. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Luo M, Shen D, Zhou X, Chen X and Wang W:
MicroRNA-497 is a potential prognostic marker in human cervical
cancer and functions as a tumor suppressor by targeting the
insulin-like growth factor 1 receptor. Surgery. 153:836–847. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen X, Yun J, Fei F, Yi J, Tian R, Li S
and Gan X: Prognostic value of nuclear hepatoma-derived growth
factor (HDGF) localization in patients with breast cancer. Pathol
Res Pract. 208:437–443. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lin YW, Li CF, Chen HY, Yen CY, Lin LC,
Huang CC, Huang HY, Wu PC, Chen CH, Chen SC and Tai MH: The
expression and prognostic significance of hepatoma-derived growth
factor in oral cancer. Oral Oncol. 48:629–635. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Abouzied MM, Baader SL, Dietz F, Kappler
J, Gieselmann V and Franken S: Expression patterns and different
subcellular localization of the growth factors HDGF
(hepatoma-derived growth factor) and HRP-3 (HDGF-related protein-3)
suggest functions in addition to their mitogenic activity. Biochem
J. 378:169–176. 2004. View Article : Google Scholar
|
|
35
|
Sedlmaier A, Wernert N, Gallitzendörfer R,
Abouzied MM, Gieselmann V and Franken S: Overexpression of
hepatoma-derived growth factor in melanocytes does not lead to
oncogenic transformation. BMC Cancer. 11:4572011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ren H, Tang X, Lee JJ, Feng L, Everett AD,
Hong WK, Khuri FR and Mao L: Expression of hepatoma-derived growth
factor is a strong prognostic predictor for patients with
early-stage non-small-cell lung cancer. J Clin Oncol. 22:3230–3237.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hu TH, Huang CC, Liu LF, Lin PR, Liu SY,
Chang HW, Changchien CS, Lee CM, Chuang JH and Tai MH: Expression
of hepatoma-derived growth factor in hepatocellular carcinoma.
Cancer. 98:1444–1456. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lepourcelet M, Tou L, Cai L, Sawada J,
Lazar AJ, Glickman JN, Williamson JA, Everett AD, Redston M, Fox
EA, et al: Insights into developmental mechanisms and cancers in
the mammalian intestine derived from serial analysis of gene
expression and study of the hepatoma-derived growth factor (HDGF).
Development. 132:415–427. 2005. View Article : Google Scholar
|
|
39
|
Yamamoto S, Makuuchi H, Shimada H, Chino
O, Nishi T, Kise Y, Kenmochi T and Hara T: Clinical analysis of
reflux esophagitis following esophagectomy with gastric tube
reconstruction. J Gastroenterol. 42:342–345. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Uyama H, Tomita Y, Nakamura H, Nakamori S,
Zhang B, Hoshida Y, Enomoto H, Okuda Y, Sakon M, Aozasa K, et al:
Hepatoma-derived growth factor is a novel prognostic factor for
patients with pancreatic cancer. Clin Cancer Res. 12:6043–6048.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bernard K, Litman E, Fitzpatrick JL,
Shellman YG, Argast G, Polvinen K, Everett AD, Fukasawa K, Norris
DA, Ahn NG and Resing KA: Functional proteomic analysis of melanoma
progression. Cancer Res. 63:6716–6725. 2003.PubMed/NCBI
|
|
42
|
Okuda Y, Nakamura H, Yoshida K, Enomoto H,
Uyama H, Hirotani T, Funamoto M, Ito H, Everett AD and Hada T:
Hepatoma-derived growth factor induces tumorigenesis in vivo
through both direct angiogenic activity and induction of vascular
endothelial growth factor. Cancer Sci. 94:1034–1041. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Matsuyama A, Inoue H, Shibuta K, Tanaka Y,
Barnard GF, Sugimachi K and Mori M: Hepatoma-derived growth factor
is associated with reduced sensitivity to irradiation in esophageal
cancer. Cancer Res. 61:5714–5717. 2001.PubMed/NCBI
|
|
44
|
Everett AD: Identification, cloning and
developmental expression of hepatoma-derived growth factor in the
developing rat heart. Dev Dyn. 222:450–458. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Everett AD, Lobe DR, Matsumura ME,
Nakamura H and McNamara CA: Hepatoma-derived growth factor
stimulates smooth muscle cell growth and is expressed in vascular
development. J Clin Invest. 105:567–575. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Oliver JA and Al-Awqati Q: An endothelial
growth factor involved in rat renal development. J Clin Invest.
102:1208–1219. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
47
|
Crossin KL, Tai MH, Krushel LA, Mauro VP
and Edelman GM: Glucocorticoid receptor pathways are involved in
the inhibition of astrocyte proliferation. Proc Natl Acad Sci USA.
94:2687–2692. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Guo Z, He Y, Wang S, Zhang A, Zhao P, Gao
C and Cao B: Various effects of hepatoma-derived growth factor on
cell growth, migration and invasion of breast cancer and prostate
cancer cells. Oncol Rep. 26:511–517. 2011.PubMed/NCBI
|