Introduction
Liver fibrosis is the final pathological consequence
of chronic liver diseases, and is characterized by the formation
and accumulation of extracellular matrix, which leads to remodeling
of the liver architecture. Following acute and chronic liver
injury, inflammation is a hallmark pathological feature of chronic
liver disease. The persistent inflammatory responses contribute to
liver fibrosis and finally lead to cirrhosis, portal hypertension
and hepatocellular carcinoma (HCC). During the process of chronic
liver damage, adaptive immune cells are crucially involved in the
pathogenesis of hepatic inflammation. The infiltrate contains
CD4+ T cells, which are important in liver injury,
antiviral defenses to hepatitis viruses and autoimmunity (1,2).
Previous studies have suggested that Th17 cells and
associated cytokines are critical factors in the pathogenesis of
liver disease (3,4). Th17 cells expressing
retinoic-acid-related orphan receptor (ROR-γt), have been
identified as a novel T cell subset, involved in the
pathophysiology of inflammatory disease. Serial investigations have
demonstrated that an increase in the number of Th17 cells
contributes to disease progression in patients with liver fibrosis
(5). By contrast, regulatory T
cells (Tregs), also known as
CD4+CD25+FoxP3+ T cells, are
involved in the maintenance of immune tolerance and homeostasis via
contact-dependent suppression or the release of anti-inflammatory
cytokines (6,7). Of note, several studies have
indicated that the imbalance in the development and function of
Th17 cells and Tregs are of critical importance in liver diseases,
including autoimmune hepatitis, primary biliary cirrhosis (PBC) and
liver fibrosis (8–10). In our previous study, Th17/Treg
cell imbalance was found to exist in mice with liver fibrosis, and
hepatic stellate cell (HSC) activation was promoted by Th17 cells,
but inhibited by Tregs, in vitro (11). Th17 cells and Tregs can be
interconverted, and are reciprocally regulated during
differentiation depending on the cytokine environment. Therefore,
correcting Th17/Treg cell imbalance to suppress Th17 and enhance
Treg cell numbers may be an attractive target for the treatment of
liver fibrosis.
It has been demonstrated that the mammalian target
of rapamycin (mTOR) inhibitor, rapamycin, is an immunosuppressive
compound, which has been used in allograft rejection (12). It has been reported that rapamycin
suppresses the transforming growth factor (TGF)-β and interleukin
(IL)-6-induced generation of IL-17-producing cells, and promotes
the TGF-β-mediated generation of Tregs (13). In addition, several studies have
indicated that rapamycin attenuates inflammatory responses through
promoting the differentiation of Tregs and inhibiting the
generation of Th17 cells (14,15).
Rapamycin markedly decreases numbers of lymphocytes, including
CD4+ T cell subsets, but selectively expands the Treg
cell population, whilst maintaining cell function (16). Additional investigations have
indicated that these expanded Tregs prevent allograft rejection
in vivo and suppress the proliferation of T cells in
vitro (17). However, whether
rapamycin exerts immunoregulatory effects in liver fibrosis remain
to be fully elucidated. Therefore, the present study aimed to
further investigate the protective effects of rapamycin, and its
function in regulating the Th17/Treg cell balance, in a carbon
tetrachloride (CCl4)-induced murine liver fibrosis
model.
Materials and methods
Animals
Male C57BL⁄6 mice (aged 6–8 weeks) were obtained
from the Shanghai SLAC Experimental Animal Centre (Shanghai, China)
and maintained in specific pathogen-free conditions at 24°C with a
12 h light/dark cycle and unlimited access to food and water. The
animal experiments were approved by the Research Ethics Committee
of Renji Hospital (Shanghai, China; no. SYXY (hu) 2011–0121). The
animals were cared for in accordance with protocols approved by the
Animal Care and Use Committee of Renji Hospital.
Induction of liver fibrosis and rapamycin
treatment
A total of 30 mice were randomly divided into three
groups (n=10). To induce liver fibrosis, the mice were injected
intraperitoneally, twice each week for 8 weeks, with 5 µl/g
of 20% CCl4 (Shanghai Jiahe Biotechnology, Shanghai,
China) dissolved in olive oil. Negative control mice were
administered with the same volume of olive oil only, for for 8
weeks. Beginning on the day of the CCl4 injection, the
mice in the treatment group were treated intraperitoneally with
rapamycin (1.25 mg/kg/day; Gene Operation, Ann Arbor, MI, USA) for
the duration of the investigation. In the remaining group of mice,
rather than rapamycin, the mice received equivalent volumes of
phosphate-buffered saline (PBS) during the course of the
experiment. All the mice were sacrificed 72 h following the final
CCl4 injection at 8 weeks, using CO2
asphyxia.
Blood biochemistry
At the time of sacrifice, blood samples were
collected from the mouse eyes (~1 ml) and immediately centrifuged
for 10 min. Liver function tests were performed at Renji Hospital
on serum samples to evaluate the levels of alanine
aminotransferase, aspartate aminotransferase and total
bilirubin.
Histological examination
At the time of sacrifice, the liver was dissected
from 6–8 mice. The liver sections were fixed in 4% formaldehyde,
embedded in paraffin, and sectioned (5 mm). The liver sections were
then stained with hematoxylin and eosin (H&E) for
histopathological examination. Masson's trichrome staining was used
for collagen determination. The content of collagen deposition was
quantified using an image analyzer (Image-Pro Plus; Media
Cybernetics, Inc., Rockville, MD, USA). Immunohistochemical
examinations were performed to detect the expression of α-smooth
muscle actin (SMA). In brief, the paraffin sections were
deparaffinized and rehydrated. The sections were exposed to fresh
3% hydrogen peroxide for 20 min, and then washed with PBS. Antigens
were retrieved in 0.01 M citric acid. The samples were incubated
for 30 min at room temperature in 5% normal blocking serum, and
incubated with a 1:100 dilution of polyclonal rabbit anti-α-SMA
(ab5694) or a 1:100 dilution of monoclonal mouse anti-TGF-β
(ab64715; Abcam, Cambridge, MA, USA) overnight at 4°C. The slides
were then incubated with HRP-conjugated goat anti-rabbit secondary
antibody (1:2,000; ab7090; Abcam) or HRP-conjugated goat anti-mouse
(1:2,000; ab6789; Abcam) for 60 min at room temperature, and with
3,3′-diaminobenzidine as a substrate. Negative controls were
obtained by omitting the primary antibodies. Finally, the sections
were counterstained with haematoxylin, and mounted.
Western blot analysis
Frozen liver tissue (50 mg) was rapidly thawed and
homogenized at 4°C in 0.5 ml lysis buffer (Beyotime Institute of
Biotechnology) for 30 min. Homogenates were centrifuged at 12,000 ×
g for 10 min at 4°C to yield supernatants. A bicinchoninic acid
protein assay kit was used to detect the protein concentrations.
The samples were then heated at 95°C for 5 min. The proteins (30
µg) were separated by 12% SDS-PAGE (Beyotime Institute of
Biotechnology, Shanghai, China) and then transferred onto a
polyvinylidene difluoride membrane (Beyotime Institute of
Biotechnology). The membrane was blocked with 5% non-fat milk for 2
h at room temperature, and incubated with indicated antibodies,
including anti-α-SMA (1:500 dilution), monoclonal mouse
anti-forkhead/winged helix transcription factor P3 (FoxP3; 1:500
dilution; ab20034; Abcam) and polyclonal rabbit anti-ROR-γt
(1:1,000 dilution; ab78007; Abcam) in Tris-buffered saline with
0.05% Tween overnight at 4°C. All antibodies were purchased from
Abcam. The membranes were then rinsed and incubated at room
temperature with HRP-conjugated secondary antibody (1:10,000
dilution; LI-COR Biosciences, Lincoln, NE, USA). Signals were
detected using an Odyssey Infrared image system (LI-COR, Inc.,
Lincoln, NE, USA).
Intracellular cytokine staining
Intracellular cytokine staining was performed, as
previously described (18). To
investigate the frequency of Tregs, lymphocytes from the spleen and
liver were prepared. To obtain single-cell suspensions from the
spleen and liver, tissue was dissected into small pieces, ground
and filtered through stainless steel meshes. Splenocytes were
isolated from the cell suspensions by gradient centrifugation at
1,400 × g for 25 min at 4°C with Lymphoprep (Dakewei Biotechnology,
Shenzhen, China). Following surface staining with
anti-CD4-fluorescein isothiocyanate (FITC; 11-0041; eBioscience,
San Diego, CA, USA) and anti-CD25-peridinin-chlorophyll-protein
(45-0251; eBioscience), the cells were fixed, permeabilized and
stained with anti-FoxP3-phycoerythrin (PE; 12-5773; eBioscience).
For Th17 cell detection, the lymphocytes from the spleen and liver
were stimulated for 5 h with phorbol-12-myristate-13-acetate (50
ng/ml), ionomycin (1 µg/ml) and 5 µg/ml brefeldin A
(Sigma-Aldrich, St. Louis, MO, USA) in RPMI-1640 (GE Healthcare
Life Sciences, Logan, UT, USA) containing 10% fetal bovine serum
(FBS; GE Healthcare Life Sciences). The cells were harvested and
stained with anti-CD4-FITC. The cells were fixed and permeabilized
with Cytofix/Cytoperm (BD Pharmingen, San Diego, CA, USA) and
stained intracellularly with anti-IL-17A-PE (12-7177; eBioscience).
Cells stained with IgG isotype control were used as controls. All
antibodies were purchased from eBioscience. Data were obtained
using a FACSCalibur flow cytometer and analyzed using CXP analysis
software, version 2.1 (Beckman Coulter, Inc., Brea, CA, USA).
Cell isolation and culture
To obtain single-cell suspensions from the spleen,
tissue was dissected into small pieces, ground and filtered through
stainless steel meshes. Splenocytes were isolated from the cell
suspensions by gradient centrifugation at 1,400 × g for 25 min at
4°C with Lymphoprep (Dakewei Biotechnology). The splenocytes were
separated into CD4+CD25+ T cells and
CD4+CD25− T cells using a
CD4+CD25+ regulatory T cell isolation kit
(Miltenyi Biotec, Bergisch Gladbach, Germany), according to the
manufacturer's protocol. These cells were cultured in RPMI-1640
medium supplemented with 10% FBS and penicillin/streptomycin. All
cells were cultured at 37°C in a humidified 5% CO2
incubator.
Suppression assay
The splenocytes from the rapamycin- or PBS-treated
mice were separated into CD4+CD25+ T cells
and CD4+CD25− T cells, as described above.
Subsequently, 2×105 CD4+CD25− T
cells (responder cells) with titrated
CD4+CD25+ T cells (effector cells) were
suspended in 200 µl RPMI-1640, seeded into 96-well plates,
and stimulated with a combination of soluble anti-CD3 (1
µl/ml; 557306; BD Pharmingen) and soluble anti-CD28 (1
µl/ml; 553295; BD Pharmingen). In co-culture experiments,
purified CD4+CD25+ T cells and
CD4+CD25− T cells were simultaneously added
into the wells, and cultured at 37°C for 72 h. Following an 18 h
pulse with [3H] thymidine (1 µCi/well; Shanghai
Institute of Nuclear Research, Shanghai, China), proliferation was
analyzed using a scintillation counter (Beckman LE 5000CE, Beckman
Coulter, Inc.).
Preparation of HSCs
Mouse HSCs were isolated from the livers of the
C57BL⁄6, as described previously (19). In brief, collagenase perfusion
in situ and density gradient centrifugation were used for
isolating the HSCs. The liver was perfused with collagenase at 37°C
at a flow rate of 18, until the hepatic parenchyma beneath the
capsule appeared liquefied. This was then subjected to gradient
centrifugation using OptiPrep (Sigma-Aldrich), at 2,000 × g for 20
min at 4°C. The isolated HSCs were cultured in RPMI-1640 (GE
Healthcare Life Sciences) containing 10% FBS and
penicillin/streptomycin. The HSCs were plated at 3×105
cells/well into 6-well plates, and were cultured for 6 days,
following which they were harvested for subsequent use. Cell
viability was >90%, determined using a trypan blue exclusion
assay. The purity of the HSCs was >92%, as determined by the
typical appearance of the lipid droplets under a light microscope
(IX50; Olympus Corporation, Tokyo, Japan) (20). All cells were cultured in a
humidified incubator with 5% CO2 at 37°C.
Co-culture experiment
A co-culture experiment was performed using the HSCs
and splenic Tregs isolated from the rapamycin- and PBS-treated
mice. The HSCs were plated at a density of 1×105
cells/well in a 24-well plate for 24 h, following which Treg cells
(1×105 or 2×105 cells/well) were added to the
culture system. The cells were co-cultured for 3 days and the
expression levels of α-SMA were detected using immunofluorescence
staining and western blot analysis.
Immunofluorescence staining
The cells were fixed in 4% formaldehyde and
permeabilized in methanol. Following blocking, the cells were
incubated overnight with rabbit anti-mouse α-SMA monoclonal
antibody (1:100 dilution; Abcam) followed by CY3-conjugated goat
anti-rabbit antibody (1:400 dilution; 112-165-143, Jackson
ImmunoResearch, West Grove, PA, USA) at room temperature. The
nuclei were stained with DAPI. The expression of α-SMA was observed
under a fluorescent microscope (Nikon Eclipse Ti-S, Nikon
Corporation, Tokyo, Japan) and fluorescence intensity was analyzed
using Image-Pro Plus software, version 6.0.
Statistical analysis
Data are presented as the mean ± standard error of
the mean. Statistical analyses were performed using SPSS version
17.0 software (SPSS, Inc., Chicago, IL, USA). Data were analyzed
using Student's t-test or one way analysis of variance.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Rapamycin protects mice against
CCl4-induced liver fibrosis
The present study aimed to assess the antifibrogenic
effects of rapamycin via the expansion of Tregs and inhibition of
Th17 cells in CCl4-induced hepatic fibrosis. To examine
the effect of rapamycin on liver function, total bilirubin and
serum aminotransferases were detected. The liver function tests
showed that rapamycin significantly reduced the concentrations of
total bilirubin and aminotransferases in the
CCl4-induced liver fibrosis model, compared with the
PBS-treated control group (Table
I).
 | Table IEffects of rapamycin on serum levels
of AST, ALT and TBIL in mice with liver fibrosis. |
Table I
Effects of rapamycin on serum levels
of AST, ALT and TBIL in mice with liver fibrosis.
| Group | ALT | AST | TBIL |
|---|
| Vehicle | 43.32±1.98 | 105.78±6.53 | 1.42±0.30 |
|
CCl4+PBS | 232.17±89.75 | 407.13±134.45 | 19.80±8.41 |
|
CCl4+rapamycin | 91.30±8.45a | 136.92±6.87a | 9.08±0.89a |
The liver fibrosis model was established by chronic
CCl4 injection, and the degree of hepatic fibrosis was
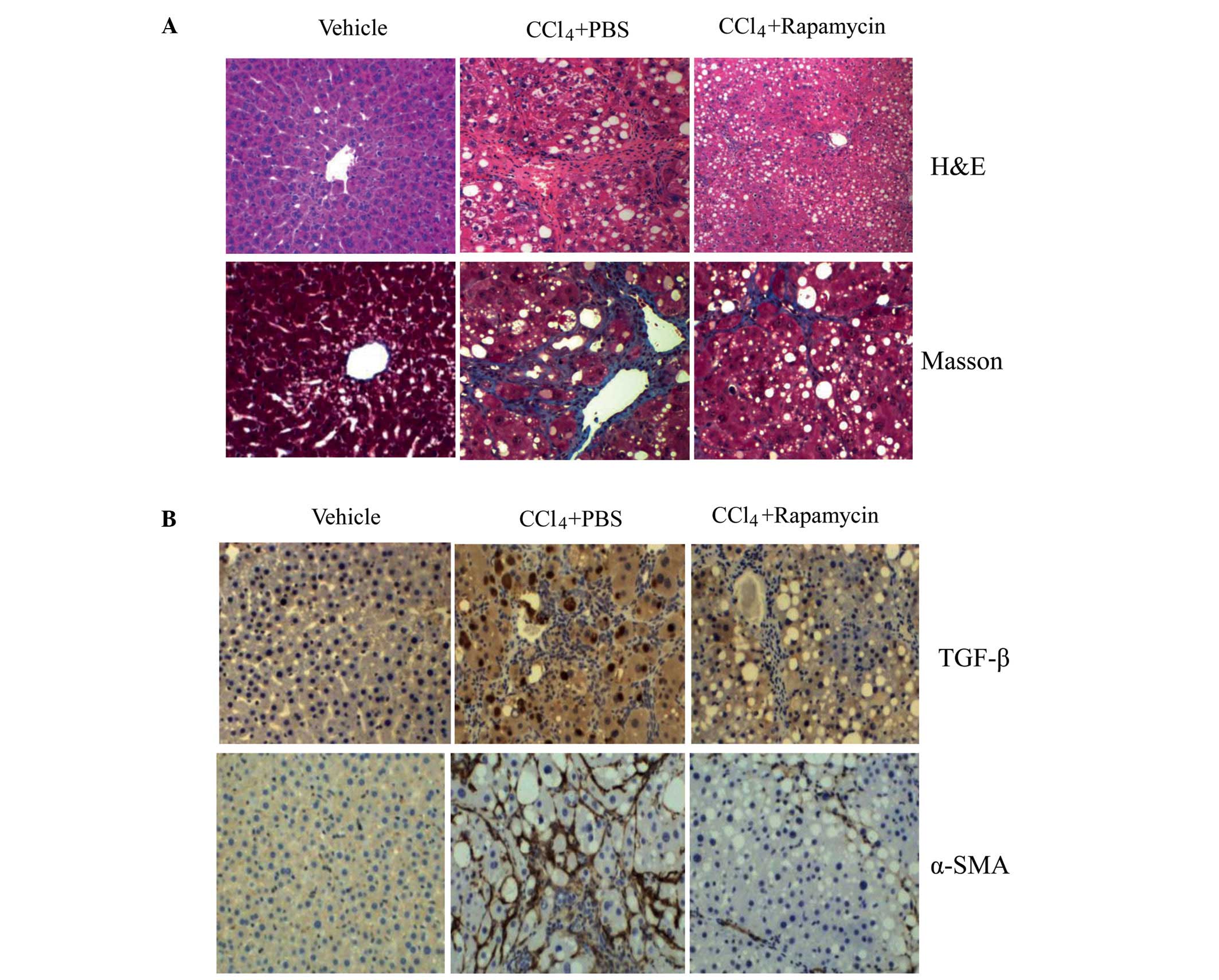
detected using H&E and Masson's trichrome staining (Fig. 1A). Following CCl4
administration, the liver tissues of the mice treated with PBS
exhibited a distorted architecture, with extensive fibrosis
combined with the development of micronodules throughout the liver
parenchyma shown in the H&E staining. Liver injury was
attenuated in the rapamycin-treated mice. The deposition of
collagen fibers as an indicator of liver fibrosis was determined
using Masson's trichrome staining. The pathological progression in
liver fibrosis was attenuated by rapamycin, with fewer and smaller
fibrotic nodules observed. As a marker of HSC activation, α-SMA is
one of the sensitive indices of the rate of fibrogenesis (21). As shown in Fig. 1B, the expression of α-SMA was
significantly elevated in the mice with CCl4-induced
liver fibrosis, and was reduced following rapamycin administration.
As a key factor of fibrogenesis, TGF-β in the rapamycin-treated
mice showed reduced expression, compared with that in the
PBS-treated mice.
Rapamycin inhibits the generation of Th17
cells
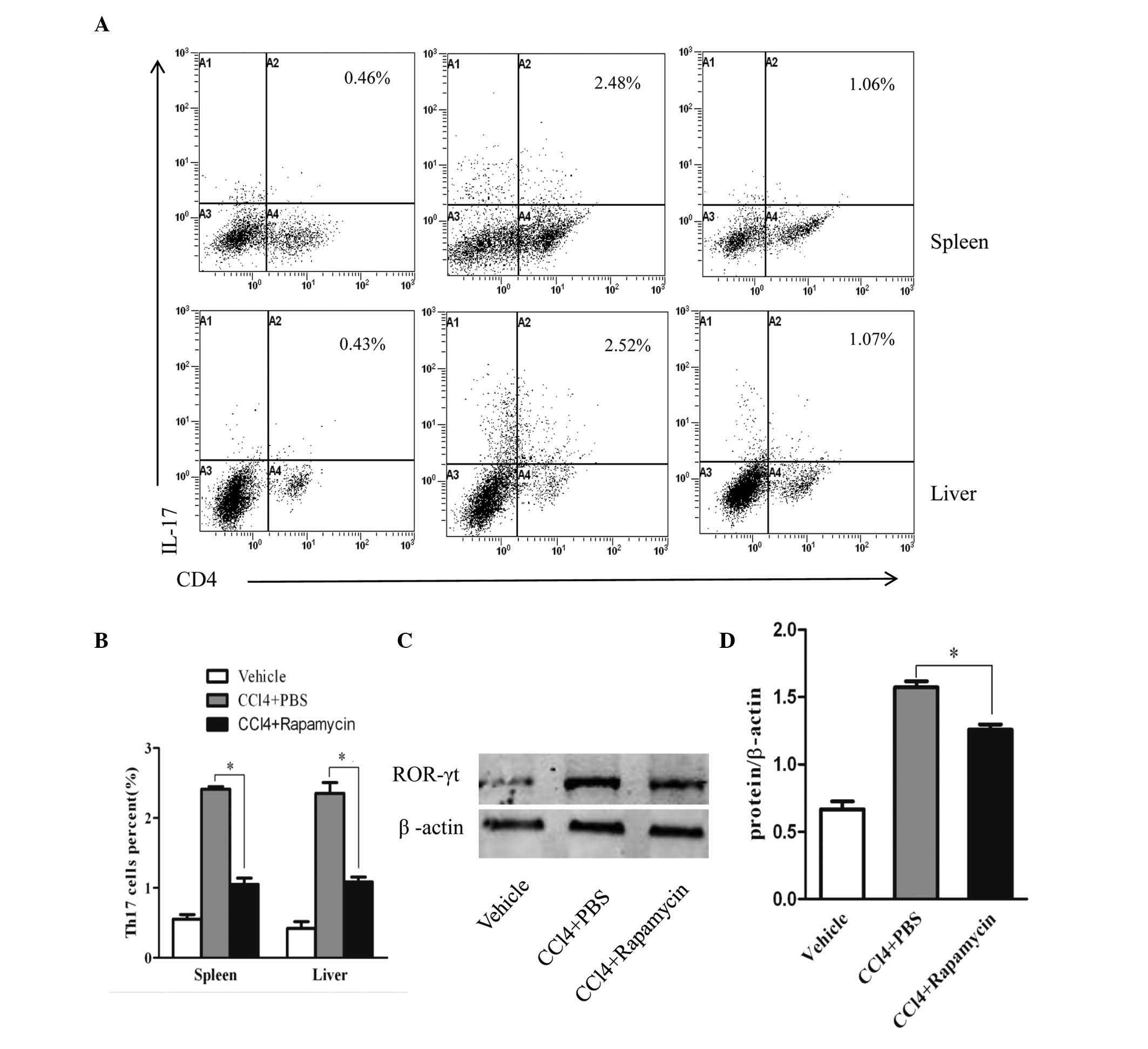
To analyze the immunoregulatory effect of rapamycin
on Th17 cells in CCl4-induced liver fibrosis, the
percentages of Th17 cells in the mouse spleen and liver were
measured using flow cytometry (Fig.
2A). As shown in Fig. 2B, the
cells from the PBS-treated mice showed markedly higher percentages
of Th17 cells, compared with the negative control group (oil
without CCl4). Following rapamycin administration, the
percentages of Th17 cells were significantly lower, compared with
those in the PBS-treated mice, but were higher, compared with the
negative control group (Fig. 2B).
In addition, as a crucial transcription factor of Th17 cell
differentiation, the expression of ROR-γt in the rapamycin-treated
mice was decreased, compared with that in the PBS-treated mice
(Fig. 2C). The relative protein
levels are shown in Fig. 2D.
Rapamycin expands the Treg cell
population, with a suppressive effect in hepatic fibrosis
The present study subsequently investigated whether
the effect of rapamycin was associated with modulation of Treg cell
function in CCl4-induced hepatic fibrosis. The
percentages of Tregs in the spleen and liver were analyzed using
flow cytometry (Fig. 3A). The
frequencies of Tregs in the spleen and liver from the
rapamycin-treated mice were significantly higher, compared with
those of the PBS-treated mice (Fig.
3B). The hepatic protein expression of FoxP3 was then examined
using western blot analysis. Consistent with the results of the
flow cytometric analysis, the expression of FoxP3 in the liver from
the rapamycin-treated mice was markedly enhanced, compared with
that from the PBS-treated mice (Fig.
3C). The relative protein levels are shown in Fig. 3D.
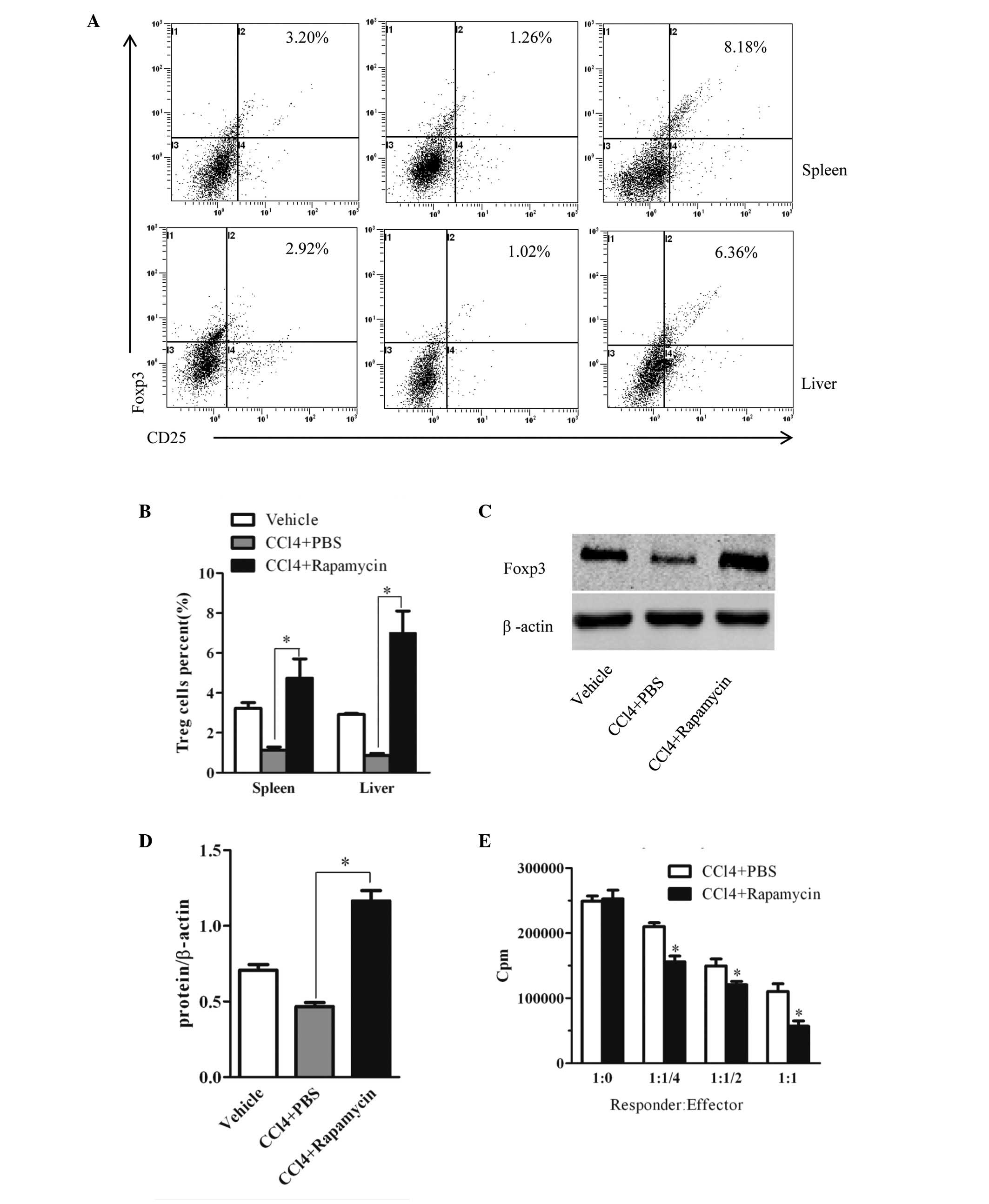 | Figure 3Rapamycin treatment upregulates the
suppressive efficacy of Tregs in mice with liver fibrosis. (A)
Cells were isolated from the spleen and liver tissues of mice in
each group and subjected to intracellular FoxP3 staining. Treg cell
frequency was analyzed using flow cytometry and (B) quantified.
Data represent one experiment of three with similar results and
values are expressed as the mean ± standard error of the mean
(n=5). *P<0.01, vs. PBS-treated group. (C) Protein
levels of FoxP3 in the liver were determined using western blot
analysis and (D) expressed as relative change, compared with the
control animals. Data are presented as the mean ± standard error of
the mean (n=6). *P<0.01, vs. PBS-treated group. (E)
CD4+CD25+ T cells were purified from the
spleen tissues of the rapamycin- or PBS-treated mice. The titrated
CD4+CD25+ T cells were co-cultured with
2×105 CD4+CD25- T cells, as responder cells, which were
obtained from PBS-treated mice in the presence of anti-CD3/CD28
antibody. The co-cultured cells were maintained for 72 h, and 1
µCi [3H]-thymidine was added to the culture 18 h prior to
harvesting. The ratios of CD4+CD25− T cells:
CD4+CD25+ T cells are shown. Data are
presented as the mean ± standard error of the mean (n=3 in each
group). *P<0.05, vs. PBS-treated group. Tregs,
regulatory T cells; CCl4, carbon tetrachloride; PBS,
phosphate-buffered saline; FoxP3, forkhead/winged helix
transcription factor P3. |
Furthermore, to clarify the effect of the mTOR
inhibitor on the suppressive efficacy of Tregs, splenic
CD4+CD25− responder T cells from the
PBS-treated mice were stimulated with anti-CD3/CD28 monoclonal
antibody and co-cultured with CD4+CD25+ Tregs
from the rapamycin- or PBS-treated mice. As shown in Fig. 3E, the suppressive activities of the
Tregs in the splenic tissues of the rapamycin-treated mice were
markedly higher, compared with those of the PBS-treated mice.
Effect of rapamycin-induced Treg cell
expansion on HSCs
It is known that regulatory immune cells, including
CD4+CD25+ T cells, are important in the
pathogenesis of liver fibrosis and the activation of HSCs (9,11).
To examine the effect of the Treg cell expansion by rapamycin on
HSCs, HSCs were isolated from the mouse liver, and then cultured
with splenic Tregs from the rapamycin- or PBS-treated mice.
Immunofluorescence staining and western blot analysis were
performed to evaluate changes in the expression of α-SMA. As shown
in Fig. 4A–C, immunofluorescence
staining showed that the expression of α-SMA was significantly
reduced following exposure to the Tregs from the rapamycin-treated
mice, compared with those from the PBS-treated mice. Fig. 4D shows the fluorescence intensity
of each group, which shows the trends described above. The results
of the western blot analysis showed that, compared with the control
group, the expression of α-SMA was reduced following exposure to
the Tregs, however, the difference was not significant. The protein
expression of α-SMA was also markedly decreased following exposure
to Tregs from the rapamycin-treated mice, compared with the
PBS-treated mice (Fig. 4E). The
expression levels of α-SMA are shown in Fig. 4F. These data showed that the
expansion of the Treg cell population by rapamycin increased the
capacity to inhibit the activation of HSCs.
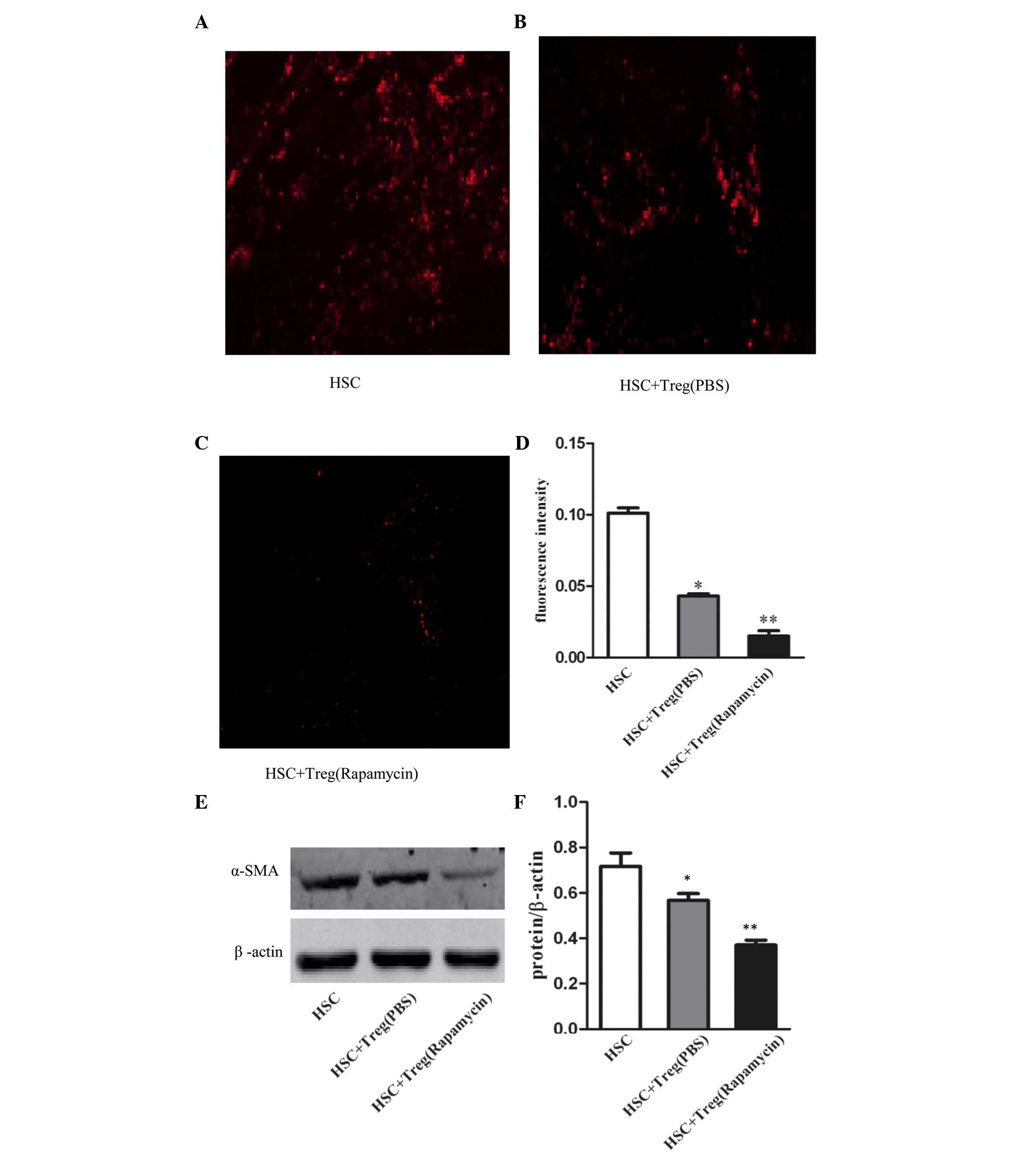 | Figure 4Treg expansion by rapamycin
suppresses the activation of HSCs. According to previous methods,
Tregs were isolated from rapamycin- or PBS-treated mice, and
co-cultured with HSCs. (A-C) Expression levels of α-SMA were
detected using immunofluorescence staining (magnification, ×200)
and western blot analysis. (D) Fluorescence intensity reflecting
the relative levels of intracellular α-SMA. Values are expressed as
the mean ± standard error of the mean of triplicate experiments.
*P<0.01, vs. control; **P<0.01, vs.
HSC+Treg (PBS). (E) Western blot analysis and (F) quantification
following treatment of HSCs with Tregs from different groups. Data
are presented as the mean ± standard error of the mean (n=3 in each
group). *P>0.05, vs. control; **P<0.01,
vs. HSC+Treg (PBS). HSCs, hepatocellular stellate cells; Treg,
regulatory T cell; CCl4, carbon tetrachloride; PBS,
phosphate-buffered saline; α-SMA, α-smooth muscle actin. |
Discussion
In the present study, prominent decreases in the
histological changes and the severity of liver fibrosis were
observed in the mice treated with rapamycin, compared with those
treated with PBS. The data from the present study showed that
rapamycin effectively protected the liver in
CCl4-induced hepatic fibrosis, which resulted from a
significant increase in the functional activity of
CD4+CD25+ Tregs. Rapamycin was involved in
the downregulation of the Th17 cell response in the development of
liver fibrosis. Furthermore, rapamycin enhanced the suppressive
capacities of the Tregs on the activation of HSCs.
Previous studies have demonstrated that Tregs
positively restrict the inflammatory response, prevent the
development of autoimmune diseases and inhibit a series of immune
responses (22,23). By contrast, Th17 cells are involved
in the induction of autoimmune diseases (24,25).
Of note, Rong et al (26)
found that the Th17 cell population and expression of ROR-γt were
markedly increased in the peripheral blood of patients with primary
biliary cirrhosis; whereas the Treg cell population and expression
of FoxP3 were markedly decreased. Consequently, a lower Treg/Th17
ratio is likely to indicate increased liver injury and progression
of fibrosis. Additional studies have reported that Tregs isolated
from patients with chronic hepatitis B inhibited the proliferation
and activation of HSCs, whereas recombinant IL-17 promoted the
proliferation and activation of HSCs (27). In accordance animal experiments
have supported findings that Treg/Th17 imbalance in mice with liver
fibrosis potentially promotes liver fibrosis via HSC activation
(11). In addition, naïve
CD4+ T cell differentiation is manipulated by distinct
expression patterns of transcription factors. For example, signal
transducer and activator of transcription (STAT)5 and FoxP3 direct
Treg cell differentiation and induce the production of regulatory
cytokines, including TGF-β and IL-10 (28), whereas STAT3 and RORγt dominate
Th17 cell polarization and IL-17 production (29). The mTOR inhibitor, rapamycin, has
been reported to regulate FoxP3 and ROR-γt genes directly, which
contribute to the induction of Treg cell differentiation and
suppresses the formation of Th17 cells (16,30).
However, the role of mTOR inhibition in regulating Th17 and Treg
cell differentiation has not been examined thoroughly in mice with
liver fibrosis.
The present study found that, in the progression of
CCl4-induced liver fibrosis, rapamycin treatment led to
a significant decrease in the percentages of Th17 cells in the
spleen and liver. Simultaneously, the numbers of Tregs in the
spleen and liver were markedly increased, compared with the
PBS-treated mice. In addition, the hepatic expression of ROR-γt was
markedly suppressed, whereas the expression of FoxP3 in the liver
was promoted following rapamycin treatment. These findings
indicated that, in liver fibrosis, rapamycin was able to correct
the imbalance between Th17 and Treg cells, thus inhibiting the
inflammatory response. The mechanism underlying the
rapamycin-induced downregulation of ROR-γt and upregulation of
Foxp3 in hepatic fibrosis remains to be fully elucidated,
indicating further in vitro experiments are required.
There has been increasing recognition that Tregs are
important in suppressing autoimmune diseases. In vitro,
Tregs with suppressive functions have been found in several
diseases, including multiple sclerosis (31), type 1 diabetes (32), and acute hepatitis C virus
infection (33). In addition,
there is increasing evidence showing that rapamycin is able to
promote the expansion of Tregs and suppress the proliferation of
responder T cells (34,35). To determine whether rapamycin can
expand the population of functional Tregs from mice with liver
fibrosis, the present study co-cultured
CD4+CD25+ T cells and
CD4+CD25− T cells to examine the suppressive
function. The results revealed an enhanced immunosuppressive
capacity of the Tregs in the rapamycin-treated group of mice,
compared with the PBS-treated mice. Therefore, the present study
also confirmed that the inhibitor of mTOR, rapamycin promoted the
differentiation of Tregs, with a suppressive function in the
development of liver fibrosis.
Saxena et al (36) reported that Tregs regulated the
cardiac fibroblast phenotype and reduced the expression of α-SMA
in vitro. According to our previous studies (11,15),
Tregs suppress immune responses, and inhibit the activation of
HSCs. The activation of HSCs is the dominant event occurring in
liver fibrogenesis, which is characterized by the transformation of
HSCs into myofibroblasts. The expression level of α-SMA is a
sensitive indicator of the fibrogenic myofibroblasts (37). To investigate whether the expansion
of Tregs by rapamycin in vitro increased the inhibitory
activity on HSC activation, the HSCs were co-cultured with Tregs
from rapamycin-treated mice or PBS-treated mice, respectively. It
was found that Treg expansion due to rapamycin reduced the
expression of α-SMA and effectively inhibited HSC activation. In
vivo, rapamycin treatment significantly reduced the expression
levels of α-SMA and TGF-β1 in the liver and reduced the progress of
fibrogenesis. These results indicated that rapamycin had an
inhibitory effect on HSC activation and hepatic fibrosis.
Acknowledgments
This study was supported by Grants from the
Scientific Research Innovation Project of Shanghai Education
Committee, China (grant no. 13yz040) and by the Shanghai Municipal
Commission of Health and Family Planning Fund (award no.
20154Y0207).
References
|
1
|
Racanelli V and Rehermann B: The liver as
an immunological organ. Hepatology. 43(2 Suppl 1): S54–S62. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Crispe IN: The liver as a lymphoid organ.
Annu Rev Immunol. 27:147–163. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lan RY, Salunga TL, Tsuneyama K, Lian ZX,
Yang GX, Hsu W, Moritoki Y, Ansari AA, Kemper C, Price J, et al:
Hepatic IL-17 responses in human and murine primary biliary
cirrhosis. J Autoimmun. 32:43–51. 2009. View Article : Google Scholar
|
|
4
|
Wang L, Chen S and Xu K: IL-17 expression
is correlated with hepatitis B-related liver diseases and fibrosis.
Int J Mol Med. 27:385–392. 2011.PubMed/NCBI
|
|
5
|
Sun HQ, Zhang JY, Zhang H, Zou ZS, Wang FS
and Jia JH: Increased Th17 cells contribute to disease progression
in patients with HBV-associated liver cirrhosis. J Viral Hepat.
19:396–403. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Workman CJ, Szymczak-Workman AL, Collison
LW, Pillai MR and Vignali DA: The development and function of
regulatory T cells. Cell Mol Life Sci. 66:2603–2622. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sakaguchi S, Yamaguchi T, Nomura T and Ono
M: Regulatory T cells and immune tolerance. Cell. 133:775–787.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fenoglio D, Bernuzzi F, Battaglia F,
Parodi A, Kalli F, Negrini S, De Palma R, Invernizzi P and Filaci
G: Th17 and regulatory T lymphocytes in primary biliary cirrhosis
and systemic sclerosis as models of autoimmune fibrotic diseases.
Autoimmun Rev. 12:300–304. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li J, Qiu SJ, She WM, Wang FP, Gao H, Li
L, Tu CT, Wang JY, Shen XZ and Jiang W: Significance of the balance
between regulatory T (Treg) and T helper 17 (Th17) cells during
hepatitis B virus related liver fibrosis. PLoS One. 7:e393072012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yasumi Y, Takikawa Y, Endo R and Suzuki K:
Interleukin-17 as a new marker of severity of acute hepatic injury.
Hepatol Res. 37:248–254. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun XF, Gu L, Deng WS and Xu Q: Impaired
balance of T helper 17/T regulatory cells in carbon
tetrachloride-induced liver fibrosis in mice. World J
Gastroenterol. 20:2062–2070. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shin HJ, Baker J, Leveson-Gower DB, Smith
AT, Sega EI and Negrin RS: Rapamycin and IL-2 reduce lethal acute
graft-versus-host disease associated with increased expansion of
donor type CD4+CD25+Foxp3+ regulatory T cells. Blood.
118:2342–2350. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kopf H, de la Rosa GM, Howard OM and Chen
X: Rapamycin inhibits differentiation of Th17 cells and promotes
generation of FoxP3+ T regulatory cells. Int Immunopharmacol.
7:1819–1824. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yin H, Li X, Zhang B, Liu T, Yuan B, Ni Q,
Hu S and Gu H: Sirolimus ameliorates inflammatory responses by
switching the Treg/Th17 profile in murine colitis. Immunology.
139:494–502. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hennig M, Bauer D, Wasmuth S, Busch M,
Walscheid K, Thanos S and Heiligenhaus A: Everolimus improves
experimental autoimmune uveoretinitis. Exp Eye Res. 105:43–52.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Battaglia M, Stabilini A and Roncarolo MG:
Rapamycin selectively expands CD4+CD25+FoxP3+ regulatory T cells.
Blood. 105:4743–4748. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qu Y, Zhang B, Zhao L, Liu G, Ma H, Rao E,
Zeng C and Zhao Y: The effect of immunosuppressive drug rapamycin
on regulatory CD4+CD25+Foxp3+T cells in mice. Transpl Immunol.
17:153–161. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ogino H, Nakamura K, Ihara E, Akiho H and
Takayanagi R: CD4+CD25+ regulatory T cells suppress Th17-responses
in an experimental colitis model. Dig Dis Sci. 56:376–386. 2011.
View Article : Google Scholar
|
|
19
|
Copple BL, Bai S, Burgoon LD and Moon JO:
Hypoxia-inducible factor-1α regulates the expression of genes in
hypoxic hepatic stellate cells important for collagen deposition
and angiogenesis. Liver Int. 31:230–244. 2011. View Article : Google Scholar
|
|
20
|
Chen CH, Kuo LM, Chang Y, Wu W, Goldbach
C, Ross MA, Stolz DB, Chen L, Fung JJ, Lu L and Qian S: In vivo
immune modulatory activity of hepatic stellate cells in mice.
Hepatology. 44:1171–1181. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Friedman SL: Evolving challenges in
hepatic fibrosis. Nat Rev Gastroenterol Hepatol. 7:425–436. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mack DG, Falta MT, McKee AS, Martin AK,
Simonian PL, Crawford F, Gordon T, Mercer RR, Hoover MD, Marrack P,
et al: Regulatory T cells modulate granulomatous inflammation in an
HLA-DP2 transgenic murine model of beryllium-induced disease. Proc
Natl Acad Sci USA. 111:8553–8558. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Eastaff-Leung N, Mabarrack N, Barbour A,
Cummins A and Barry S: Foxp3+ regulatory T cells, Th17 effector
cells and cytokine environment in inflammatory bowel disease. J
Clin Immunol. 30:80–89. 2010. View Article : Google Scholar
|
|
24
|
Shah K, Lee WW, Lee SH, Kim SH, Kang SW,
Craft J and Kang I: Dysregulated balance of Th17 and Th1 cells in
systemic lupus erythematosus. Arthritis Res Ther. 12:R532010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lohr J, Knoechel B, Wang JJ, Villarino AV
and Abbas AK: Role of IL-17 and regulatory T lymphocytes in a
systemic autoimmune disease. J Exp Med. 203:2785–2791. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rong G, Zhou Y, Xiong Y, Zhou L, Geng H,
Jiang T, Zhu Y, Lu H, Zhang S, Wang P, et al: Imbalance between T
helper type 17 and T regulatory cells in patients with primary
biliary cirrhosis: The serum cytokine profile and peripheral cell
population. Clin Exp Immunol. 156:217–225. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kong X, Feng D, Wang H, Hong F, Bertola A,
Wang FS and Gao B: Interleukin-22 induces hepatic stellate cell
senescence and restricts liver fibrosis in mice. Hepatology.
56:1150–1159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fontenot JD, Gavin MA and Rudensky AY:
Foxp3 programs the development and function of CD4+CD25+ regulatory
T cells. Nat Immunol. 4:330–336. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ivanov II, McKenzie BS, Zhou L, Tadokoro
CE, Lepelley A, Lafaille JJ, Cua DJ and Littman DR: The orphan
nuclear receptor RORgammat directs the differentiation program of
proinflammatory IL-17+ T helper cells. Cell. 126:1121–1133. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yin H, Li X, Zhang B, Liu T, Yuan B, Ni Q,
Hu S and Gu H: Sirolimus ameliorates inflammatory responses by
switching the regulatory T/T helper type 17 profile in murine
colitis. Immunology. 139:494–502. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Viglietta V, Baecher-Allan C, Weiner HL
and Hafler DA: Loss of functional suppression by CD4+CD25+
regulatory T cells in patients with multiple sclerosis. J Exp Med.
199:971–979. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lawson JM, Tremble J, Dayan C, Beyan H,
Leslie RD, Peakman M and Tree TI: Increased resistance to
CD4+CD25hi regulatory T cell-mediated suppression in patients with
type 1 diabetes. Clin Exp Immunol. 154:353–359. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Smyk-Pearson S, Golden-Mason L, Klarquist
J, Burton JR Jr, Tester IA, Wang CC, Culbertson N, Vandenbark AA
and Rosen HR: Functional suppression by FoxP3+CD4+CD25 (high)
regulatory T cells during acute hepatitis C virus infection. J
Infect Dis. 197:46–57. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Battaglia M, Stabilini A, Migliavacca B,
Horejs-Hoeck J, Kaupper T and Roncarolo MG: Rapamycin promotes
expansion of functional CD4+CD25+FOXP3+ regulatory T cells of both
healthy subjects and type 1 diabetic patients. J Immunol.
177:8338–8347. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tresoldi E, Dell'Albani I, Stabilini A,
Jofra T, Valle A, Gagliani N, Bondanza A, Roncarolo MG and
Battaglia M: Stability of human rapamycin-expanded CD4+CD25+ T
regulatory cells. Haematologica. 96:1357–1365. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Saxena A, Dobaczewski M, Rai V, Haque Z,
Chen W, Li N and Frangogiannis NG: Regulatory T cells are recruited
in the infarcted mouse myocardium and may modulate fibroblast
phenotype and function. Am J Physiol Heart Circ Physiol.
307:H1233–H1242. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Huang GC, Zhang JS and Tang QQ:
Involvement of C/EBP-alpha gene in in vitro activation of rat
hepatic stellate cells. Biochem Biophys Res Commun. 324:1309–1318.
2004. View Article : Google Scholar : PubMed/NCBI
|