Introduction
Despite advances in pharmacological and mechanical
therapies, acute decompensated heart failure (ADHF) remains one of
the most frequently encountered cardiovascular diseases and its
post-discharge mortality rates can reach 8.1 and 20.5% within 3
months and 1 year, respectively (1–3).
Over the past decades, it has become evident that acute heart
failure (HF) and renal failure (RF) are associated with systemic
inflammatory activation (4). It
has become increasingly recognized that venous congestion and
volume overload are involved in the cardiorenal syndrome (4,5).
Certain animal and human studies indicate that congestion may lead
to venous endothelial activation with peripheral synthesis and
release of proinflammatory mediators, including cyclooxygenase-2,
inducible nitric oxide synthase, interleukin-6 (IL-6) and tumor
necrosis factor-α (6–8). These suggest that the venous
endothelium is in a proinflammatory state in times of venous
congestion (4). To investigate
global mRNA changes during peripheral venous congestion, Colombo
et al (5) simulated
peripheral venous congestion and analyzed mRNA by using the
Affymetrix HG-U133 Plus 2.0 microarray.
Long noncoding RNAs (lncRNAs) have become a research
hotspot for numerous diseases. A number of lncRNAs have been
demonstrated to have important and diverse functions (9,10).
LncRNA-associated dysfunction has been demonstrated to be important
in cancer (11), cardiovascular
diseases (12), and
neurodegeneration diseases (11).
Particularly, it is becoming evident that lncRNA may be involved in
cardiovascular diseases. For example, the myocardial
infarction-associated transcript lncRNA is associated with
myocardial infarction (13).
Another study identified 15 lncRNAs modulated in the heart of mice
subjected to aortic constriction (14). However, global analysis of lncRNA
associated with peripheral venous congestion is required and the
potential underlying regulatory mechanisms remain unclear, due to
the limited RNA sequencing (RNA-Seq) data. Thus, the present study
re-annotated an Affymetrix microarray associated with peripheral
venous congestion, then constructed a dynamic lncRNA-mRNA
co-expression network (5).
Following functional analysis of this network, it was demonstrated,
although the genes (mRNAs) and lncRNAs were different, certain
venous congestion-associated GO terms, including ion channel
activity, were identified. The current study also identified
certain lncRNA dynamically regulated pathways, including dilated
cardiomyopathy and the calcium signaling pathway, in which the
involvement of lncRNAs persistently occurred from normal and
peripheral venous congestion conditions. To the best of our
knowledge, the present study was the first to analyze the dynamic
lncRNA-associated mechanism of peripheral venous congestion and
provide insights into the understanding of the functional mechanism
of peripheral venous congestion and lncRNAs.
Materials and methods
Microarray data
The microarray data set GSE38783 was accessed using
the Gene Expression Omnibus database (ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE38783). This
data set was obtained from 12 healthy subjects using the Affymetrix
HG-U133 Plus 2.0 microarray (Affymetrix, Inc., Santa Clara, CA,
USA) (5). Venous arm pressure was
increased to 30 mmHg above the baseline level by inflating a
tourniquet cuff around the dominant arm (test arm). Then
endothelial cells were obtained from blood samples from the test
and control arm (lacking an inflated cuff) before and after 75 min
of venous congestion (5).
Functional re-annotation of lncRNAs
To re-annotate micro-array data, a non-coding RNA
function annotation server (ncFANs) was used to re-annotate the
probes of the HG-U133 Plus 2.0 array as described on the ncFANs
website (15). Then each probe was
converted into gene Ensembl Gene IDs. There were 3495 lncRNAs
re-annotated. When a gene matched more than one probe, the
expression value of this mRNA or lncRNA was computed by taking the
average expression value of all the corresponding probes.
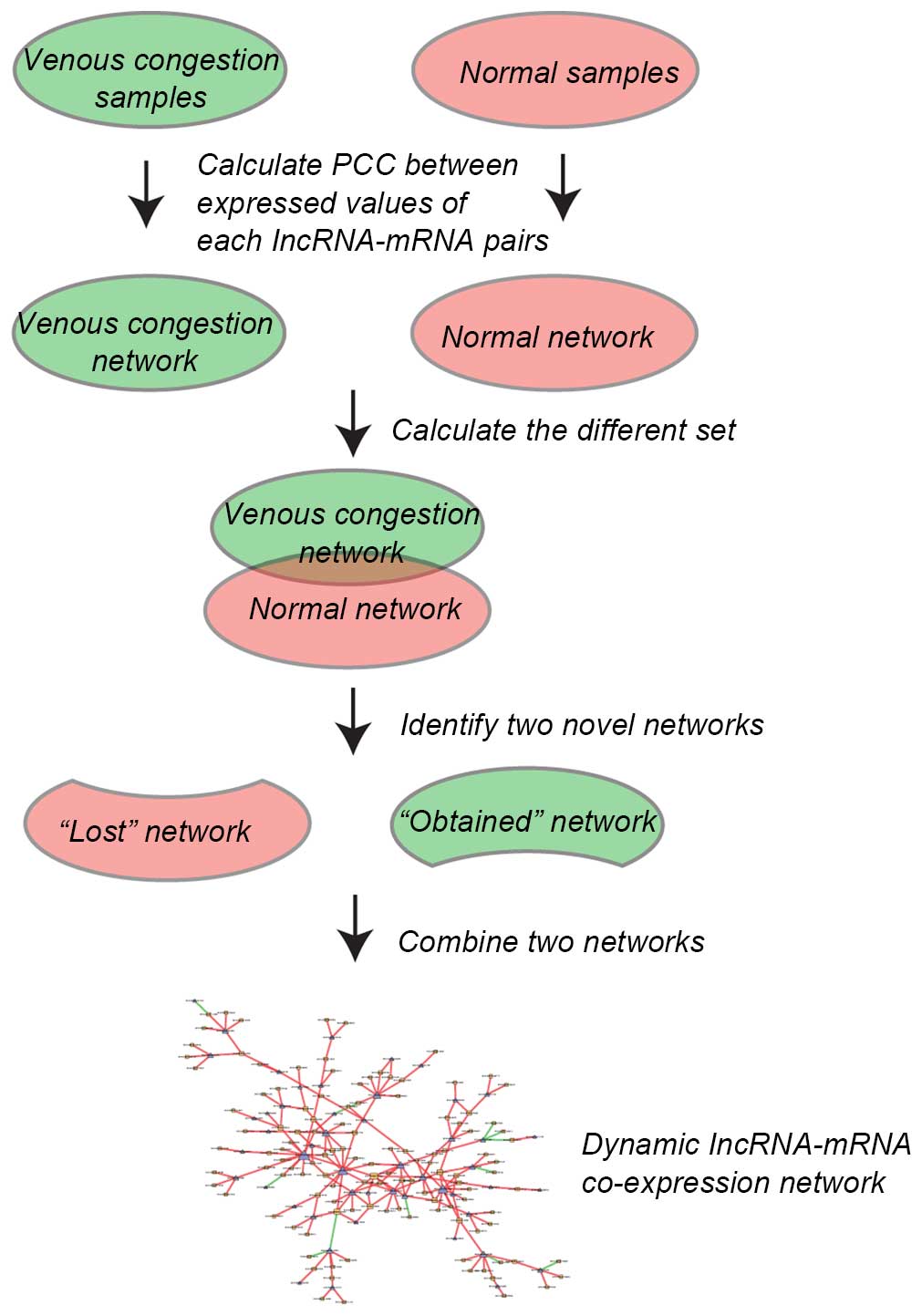
Construction of dynamic lncRNA-mRNA
co-expression network
Pearson's correlation coefficient (PCC) was
calculated between expressed values of each lncRNA-mRNA pair in
normal samples and venous congestion samples. The co-expressed
lncRNA-mRNA pairs with PCC>0.99 or <−0.99 and P<0.01 were
selected. To construct the dynamic lncRNA-mRNA co-expression
network, two lncRNA-mRNA co-expression networks were initially
constructed based on lncRNA-mRNA co-expression associations in
normal samples and venous congestion samples. Then, the different
sets of the above two networks were calculated and the following
two novel networks identified: i) 'Lost' network, in which
lncRNA-mRNA co-expression pairs only appeared in normal samples and
not in the venous congestion samples; and ii) 'obtained' network in
which co-expression pairs only appeared in venous congestion
samples and not in normal samples. Finally, the 'lost' and
'obtained' networks were combined together to obtain the final
dynamic lncRNA-mRNA co-expression network (the edges of 'lost' and
'obtained' network were added). The procedure is illustrated in
Fig. 1.
Functional enrichment
To determine potential functions of lncRNAs,
co-expressed genes for the lncRNAs were obtained then GO enrichment
was implemented to identify the affected GO terms of lncRNA
co-expressed genes using the Database for Annotation, Visualization
and Integrated Discovery 6.7 (16). P<0.01 was considered to indicate
a statistically significant difference. Pathway annotation used the
Kyoto Encyclopedia of Genes and Genomes (KEGG) search and color
tools.
Results
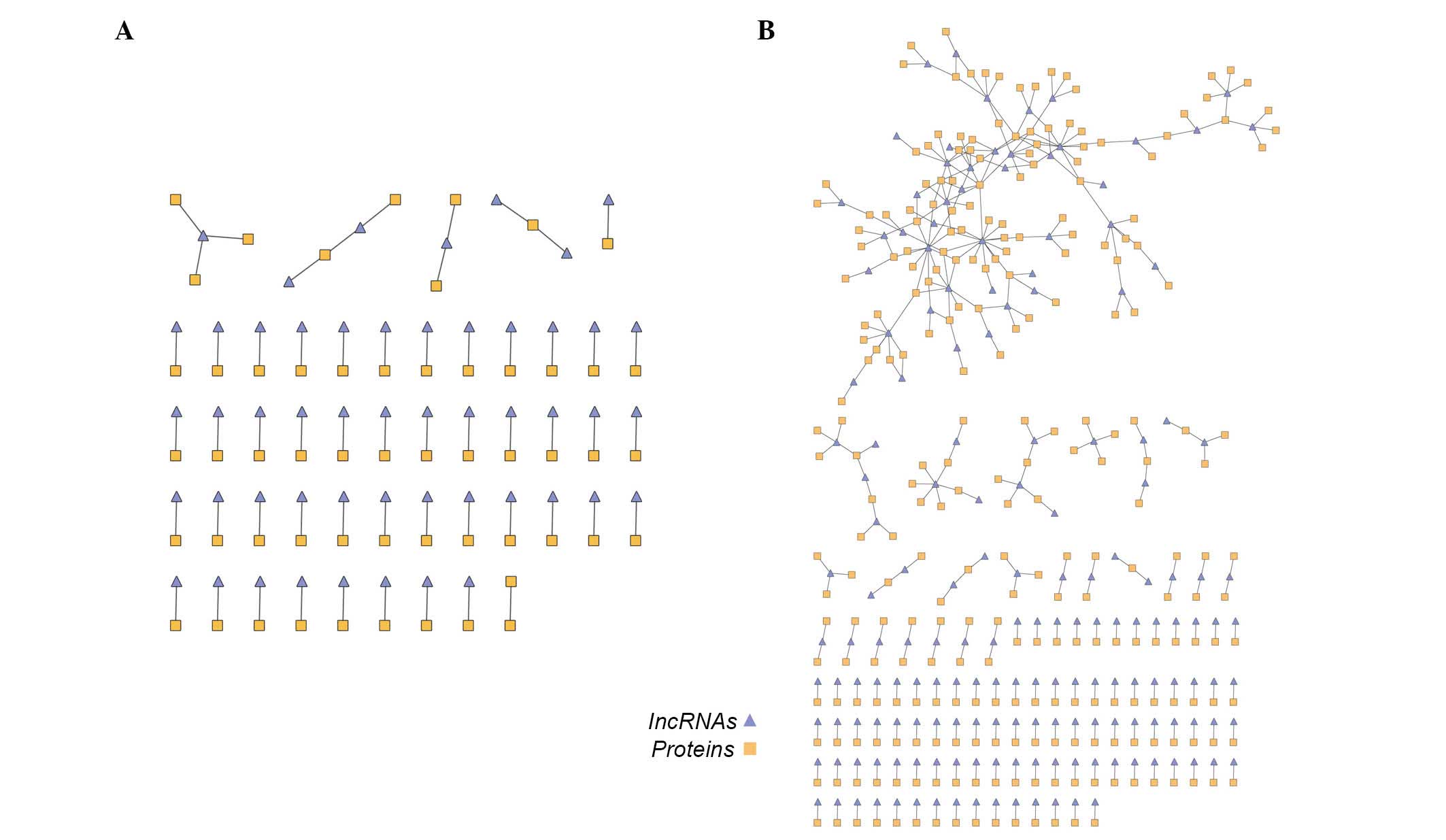
Co-expression networks based on normal
and venous congestion samples
To construct the dynamic lncRNA-mRNA co-expression
network, two lncRNA-mRNA co-expression networks based on
lncRNA-mRNA co-expression associations in normal samples and venous
congestion samples were initially constructed. In the 'venous
congestion' co-expression network, there were 56 lncRNA nodes, 61
mRNA nodes and 64 edges, whereas the 'normal' network contained 364
lncRNA-gene co-expressing associations composed of 175 lncRNAs and
275 mRNAs (Fig. 2A and B).
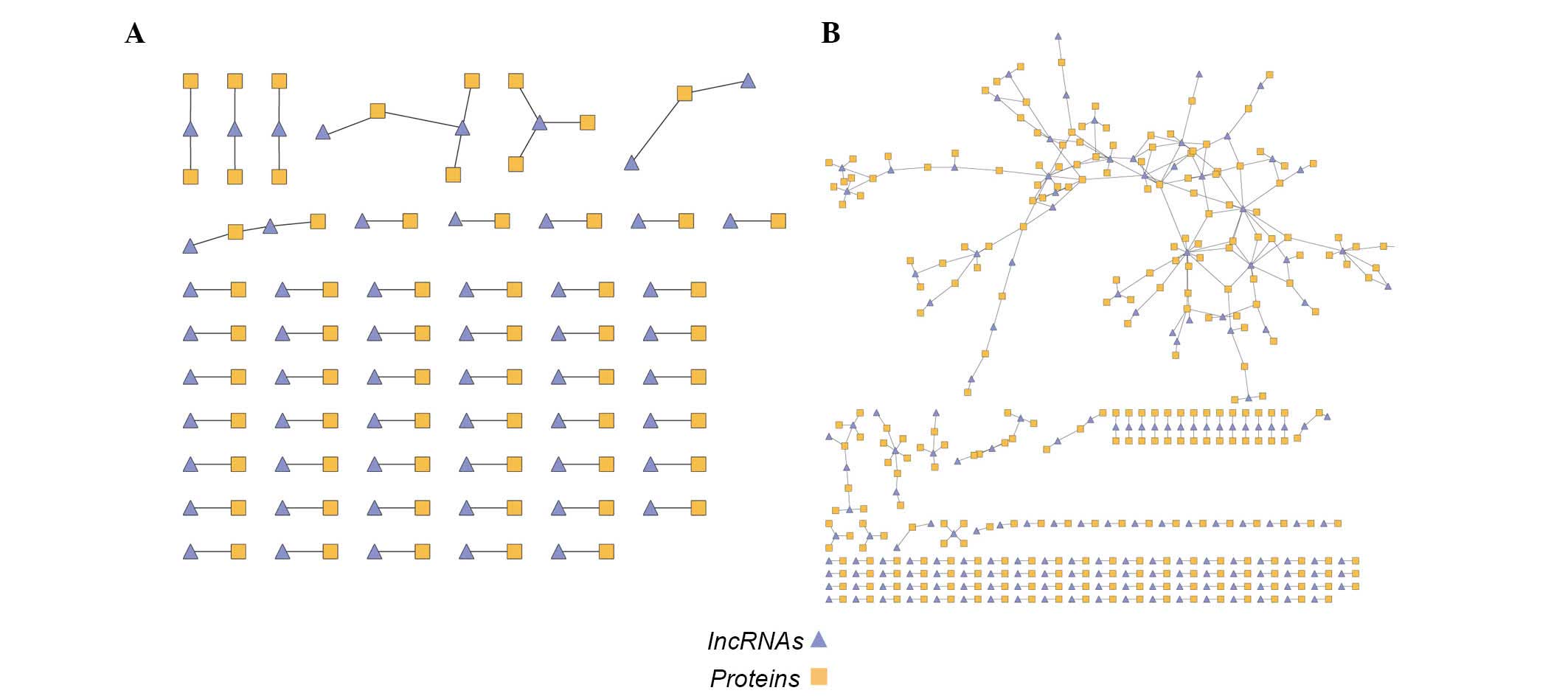
'Lost' and 'obtained' network
The different sets of the above two networks were
then calculated and the following two novel networks were created:
i) The 'lost' network contained 350 edges composed by 172 lncRNA
and 266 mRNA nodes and ii) the 'obtained' network contained 56
edges composed of 52 lncRNA and 54 mRNA nodes (Fig. 3A and B). To investigate the 'lost'
and 'obtained' functions, GO enrichment was performed on the two
networks (Tables I and II). Certain terms (including ion channel
activity) were significantly enriched (P<0.01), although the
mRNAs in the two networks differed.
 | Table IGO terms enriched by 'obtained'
genes. |
Table I
GO terms enriched by 'obtained'
genes.
| GO term | P-value |
|---|
| GO:0005216 ion
channel activity | 0.002033 |
| GO:0022838 substrate
specific channel activity | 0.002323 |
| GO:0015267 channel
activity | 0.002700 |
| GO:0022803 passive
transmembrane transporter activity | 0.002728 |
| GO:0005261 cation
channel activity | 0.003950 |
| GO:0022836 gated
channel activity | 0.006033 |
| GO:0046873 metal ion
transmembrane transporter activity | 0.007346 |
| Table IIGO terms enriched by 'lost' genes. |
Table II
GO terms enriched by 'lost' genes.
| GO terms | P-value |
|---|
| GO:0015267 channel
activity | 0.001265 |
| GO:0022803 passive
transmembrane transporter activity | 0.001296 |
| GO:0043565
sequence-specific DNA binding | 0.001758 |
| GO:0003700
transcription factor activity | 0.002589 |
| GO:0005261 cation
channel activity | 0.002731 |
| GO:0046873 metal ion
transmembrane transporter activity | 0.003682 |
| GO:0005216 ion
channel activity | 0.005109 |
| GO:0043176 amine
binding | 0.005961 |
| GO:0022838 substrate
specific channel activity | 0.006561 |
| GO:0000287 magnesium
ion binding | 0.007641 |
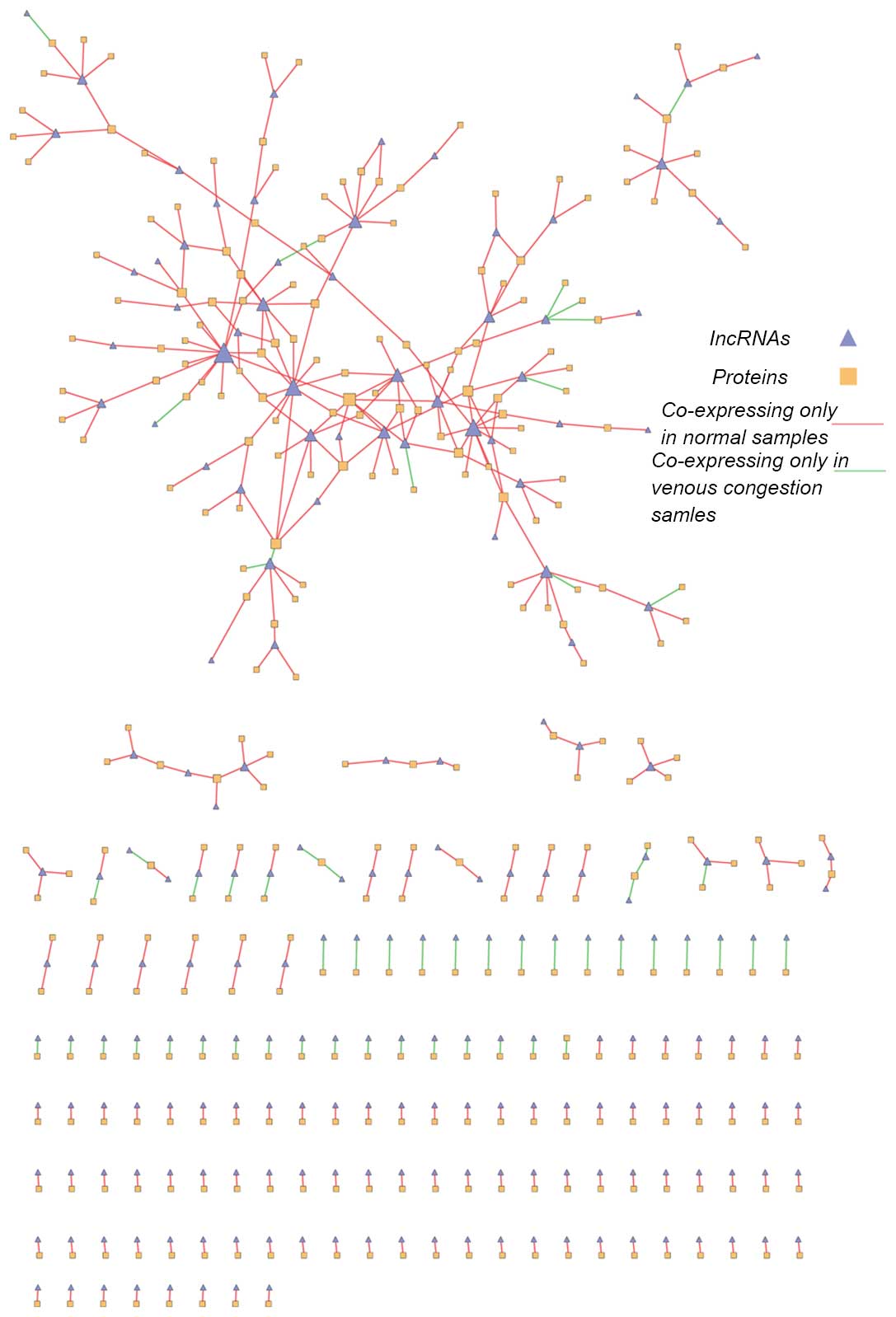
Dynamic lncRNA-mRNA co-expression
network
The 'lost' and 'obtained' networks were combined to
obtain the final dynamic lncRNA-mRNA co-expression network
(Fig. 4). This network contained
524 nodes (211 lncRNA nodes and 313 mRNA nodes) and 406 edges. The
giant component was formed of 172 nodes. The top 10 highest degree
nodes are demonstrated in Tables
III and IV. The current study
demonstrated that certain lncRNAs exhibited co-expressed mRNAs in
the normal and venous congestion conditions, indicating that these
lncRNAs may dynamically regulate certain biological process during
the development of venous congestion. Certain mRNAs also had
co-expressed lncRNA partners in both conditions, indicating that
they were dynamically regulated by different lncRNAs during the
development of venous congestion.
| Table IIITop 10 long non-coding RNA nodes with
the highest degree. |
Table III
Top 10 long non-coding RNA nodes with
the highest degree.
| Ensembl ID | Name | Degree |
|---|
| ENSG00000196273 | LINC00523 | 14 |
| ENSG00000225946 | RP11-395B7.2 | 11 |
| ENSG00000267649 | CTD-2587H24.10 | 11 |
| ENSG00000233101 | HOXB-AS3 | 8 |
| ENSG00000260394 | LA16c-313D11.9 | 8 |
| ENSG00000224184 | AC096559.1 | 8 |
| ENSG00000270168 | LA16c-380H5.4 | 7 |
| ENSG00000272933 | RP11-47A8.5 | 7 |
| ENSG00000228008 | CTD-2330K9.3 | 7 |
| ENSG00000259905 | PWRN1 | 7 |
| Table IVTop 10 mRNA nodes with the highest
degree. |
Table IV
Top 10 mRNA nodes with the highest
degree.
| Ensembl ID | Name | Degree |
|---|
| ENSG00000176884 | GRIN1 | 7 |
| ENSG00000183250 | C21orf67 | 5 |
| ENSG00000167011 | NAT16 | 5 |
| ENSG00000108342 | CSF3 | 4 |
| ENSG00000086205 | FOLH1 | 4 |
| ENSG00000186868 | MAPT | 4 |
|
ENSG00000123989 | CHPF | 4 |
|
ENSG00000178498 | DTX3 | 3 |
|
ENSG00000141150 | RASL10B | 3 |
|
ENSG00000154252 | GAL3ST2 | 3 |
Detection of dynamically alternative
pathways potentially regulated by lncRNAs
To investigate the dynamic alternative biological
functions of lncRNAs during the development of venous congestion,
lncRNA co-expressing mRNAs in the 'lost' and 'obtained' networks
were mapped into KEGG pathways. A dynamic alternative pathway
network was constructed (Fig. 5A).
The present study demonstrated that i) There were certain pathways
directly associated with venous congestion (hsa05414, dilated
cardiomyopathy; these pathways are illustrated with red labels in
Fig. 5A); and ii) certain pathways
(hsa05414, dilated cardiomyopathy; hsa04020, calcium signaling
pathway), were annotated by the 'lost' and 'obtained' mRNAs. For
detailed investigation of the dynamic changes, the 'lost' and
'obtained' mRNAs were mapped into proteins in the calcium signaling
pathway (Fig. 5B; pink rectangle
and blue pentagram represent proteins which were potentially
regulated by lncRNAs in normal and venous congestion samples).
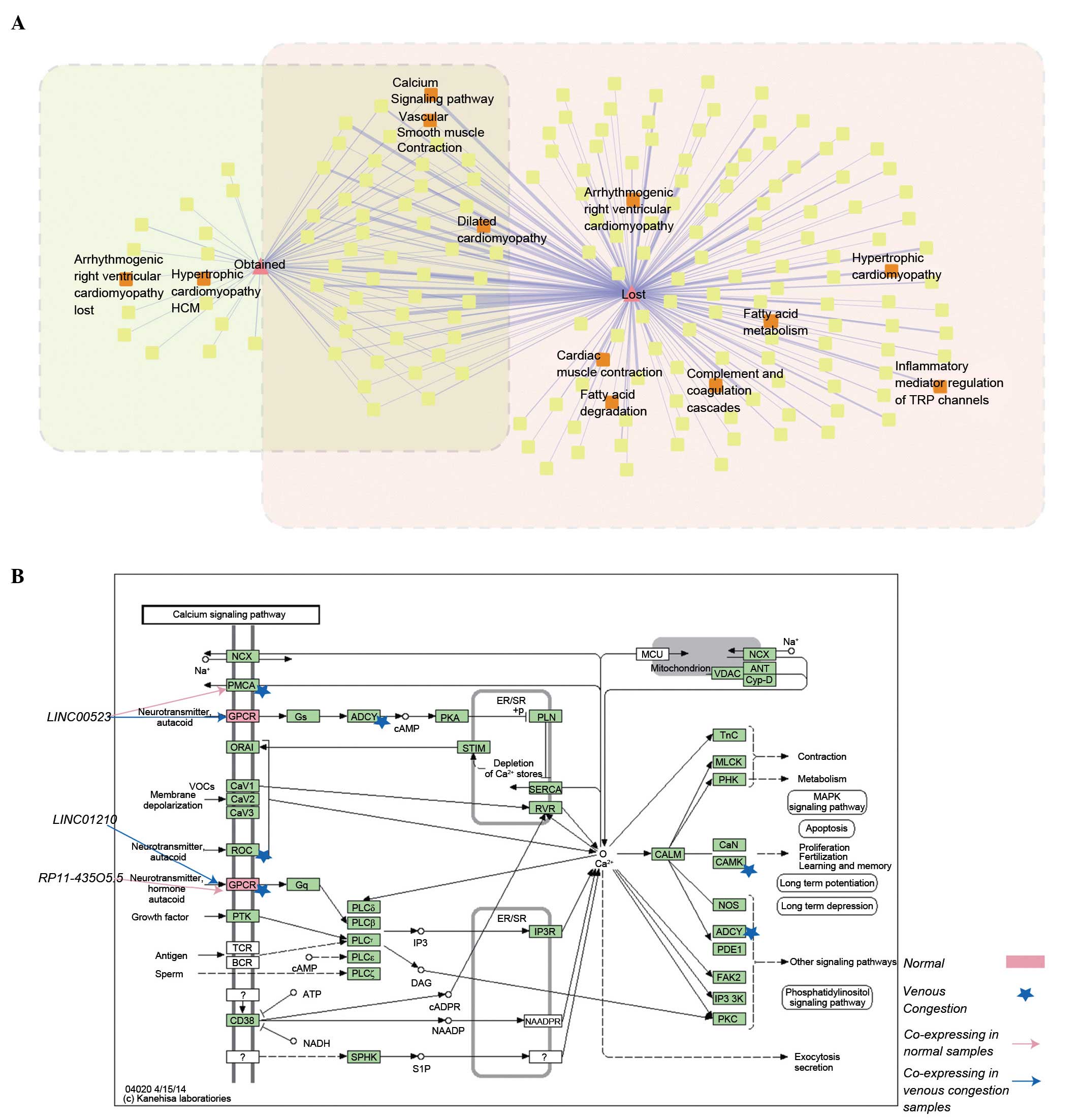 | Figure 5(A) Dynamically alternative pathway
network. (B) Calcium signaling pathway; the pink rectangle and blue
pentagram denoted proteins which were annotated by 'lost' and
'obtained' mRNAs, meaning that they were dynamically regulated
lncRNAs in normal and venous congestion condition, respectively.
lncRNA, long non-coding RNA; NCX: Na+/Ca2+
exchanger-3; PMCA, plasma membrane calcium ATPase; GPCR, G
protein-coupled receptor; ORAI, calcium release-activated calcium
modulator; CAV, certain voltage-activated Ca2+ channels;
ROC, nicotinic acetylcholine receptor α-7; Gq, guanine
nucleotide-binding protein G(q) subunit α; PTK, receptor
tyrosine-protein kinase erbB-2; TCR, T cell receptor α-chain V
region; BCR, immunoglobulin heavy chain; CD38, ADP-ribosyl cyclase
1; Gs, guanine nucleotide-binding protein G(s) subunit α; ADCY,
adenylate cyclase 1; PLC, phosphatidylinositol phospholipase C;
SPHK, sphingosine kinase; PKA, protein kinase A; STIM, stromal
interaction molecule; PLN, phospholamban; SERCA, Ca2+
transporting ATPase, sarcoplasmic/endoplasmic reticulum; RYR,
ryanodine receptor; IP3R, inositol 1,4,5-triphosphate receptor
type; CALM, calmodulin; VDAC, voltage-dependent anion channel
protein; TnC, troponin C, slow skeletal and cardiac muscles; MLCK,
myosin-light-chain kinase; PHK, phosphorylase kinase; CaN,
serine/threonine-protein phosphatase 2B; CAMK,
calcium/calmodulin-dependent protein kinase; NOS, nitric-oxide
synthase; PDE1, phosphodiesterase 1; FAK2, focal adhesion kinase 2;
IP33K, 1D-myo-inositol-triphosphate 3-kinase; PKC, protein kinase
C. |
Discussion
In recent years, the significant functional
molecular mechanisms of lncRNAs have been recognized, particularly
in cardiovascular diseases. RNA-Seq is currently a prevalent
technique to profile lncRNAs. However, due to relatively high cost
of RNA-Seq techniques, publicly available RNA-Seq data are limited
compared with microarray expression profile data. Furthermore,
RNA-Seq data lack high sample numbers, in contrast with microarray
expression profile data that often contains dozens to hundreds of
pair-matched samples. Thus, the current study adopted a
re-annotation method to identify lncRNAs associated with venous
congestion (15).
Furthermore, increasing evidence has demonstrated
that lncRNAs may be important for regulating gene expression. The
functions of lncRNAs are predominantly performed by their secondary
structures, however, this is difficult to decipher (17). Due to considerable challenges in
investigating the lncRNA functions, a co-expression-based method
was developed, in which lncRNA functions were predicted based on
the functions of their co-expressed protein-coding genes (17), as genes with similar expression
patterns under multiple conditions tend to be involved in the same
pathways (18). The co-expressed
protein-coding genes are potentially regulated by the corresponding
lncRNAs.
Thus, in the current study, an Affymetrix microarray
associated with peripheral venous congestion was re-annotated and
normal and venous congestion co-expression networks were
constructed (Fig. 2A and B). Based
on these networks, another two networks were created; the 'lost'
and 'obtained' networks. The 'lost' network represented the
lncRNA-mRNA co-expression associations that occurred in the normal
condition but disappeared in the venous congestion condition. By
contrast, the 'obtained' network represented the novel lncRNA-mRNA
co-expression associations that occurred in the venous congestion
conditions, but were not present in the normal conditions. Then, GO
enrichment was performed on the protein-coding mRNAs of the two
networks. It was assumed that the GO terms of 'obtained' network
represented 'novel' biological processes that were potentially
influenced by lncRNAs, whereas the GO terms of the 'lost' network
represented dysregulated biological processes that maintained the
normal functions of the human body. Notably, numerous terms closely
associated with venous congestion, including ion channel activity
(19), were significantly
enriched, suggesting that lncRNAs may be involved in similar
biological processes by regulating the same mRNAs or different
mRNAs in the same functional term (Tables I and II).
To decipher the dynamically regulated process during
the development of venous congestion, the 'lost' and 'obtained'
networks were combined to form the dynamic lncRNA-mRNA
co-expression network (Fig. 4).
The nodes with the highest degree are demonstrated in Tables III and IV. The most-connected mRNAs nodes were
glutamate ionotropic receptor N-methyl D-aspartate 1 (GRIN1). It
has been previously reported that the release of proinflammatory
cytokine IL-1β in venous congestion condition enhanced GRIN1
responses through IL-1 receptor activation (20).
The current study also demonstrated that certain
lncRNAs were 'dynamically' co-expressed with different mRNAs, and
certain mRNAs were 'dynamically' co-expressed with different
lncRNAs. To further investigate the dynamic biological pathways, a
dynamic alternative pathway network was constructed (Fig. 5A). This demonstrated that there
were certain pathways (for example, the pathway of 'dilated
cardiomyopathy') directly associated with venous congestion or
cardiovascular diseases (21).
Furthermore, certain pathways, for example, hsa04020: calcium
signaling pathway, were annotated by the 'lost' and 'obtained'
mRNAs, suggesting that lncRNA dynamically regulate this pathway.
The 'lost' and 'obtained' mRNAs and their corresponding lncRNAs
were annotated to the calcium signaling pathway (Fig. 5B). G protein-coupled receptors
(GPCRs) were potentially regulated in the normal and venous
congestion conditions, but by different lncRNAs (LINC01210 in
venous congestion and RP11-435O5.5 in normal samples; Fig. 5B). A previous novel study
demonstrated the importance of GPCR signaling partners in
endothelial cell nitric-oxide synthase (eNOS) function and the
fundamental implications for vascular disorders involving
dysregulated eNOS (22). In the
normal samples, lncRNA-LINC00523 were co-expressed with PMCA,
indicating the potential regulated association of LINC00523 with
PMCA. However, in the venous congestion, this association was
changed. LINC00523 were co-expressed with GPCRs. PMCA is a calcium
pump, with a pivotal function in maintaining intracellular calcium
homeostasis (23,24). Therefore, this observation
indicated that, although further experimental validation is
required, the dynamic regulated switch of LINC00523 from PMCA to
GPCR may be involved in the damage to steady state intracellular
calcium levels.
In the current study, the underlying role of lncRNAs
in the development of venous congestion was investigated by the
construction of a dynamic lncRNA-mRNA co-expression network.
Functional analysis identified several dynamic alternative
pathways, including the calcium signaling pathway. Notably, it was
suggested that the dynamically regulated switch of LINC00523 from
PMCA to GPCR may be involved in damage to the intracellular calcium
homeostasis, leading to the development of venous congestion. In
brief, the current study demonstrated a potential novel mechanism
of lncRNA function during venous congestion.
Acknowledgments
This work was supported in part by the Health and
Family Planning Commission of Heilongjiang Province [Grant Nos.
2012-357] and Health and Family Planning Commission of Heilongjiang
Province [Grant Nos. 2013-110].
References
|
1
|
Gheorghiade M and Braunwald E:
Reconsidering the role for digoxin in the management of acute heart
failure syndromes. JAMA. 302:2146–2147. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gheorghiade M and Pang PS: Acute heart
failure syndromes. J Am Coll Cardiol. 53:557–573. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Inohara T, Kohsaka S, Sato N, Kajimoto K,
Keida T, Mizuno M and Takano T; ATTEND Investigators: Prognostic
impact of renal dysfunction does not differ according to the
clinical profiles of patients: Insight from the acute decompensated
heart failure syndromes (ATTEND) registry. PloS One. 9:e1055962014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Colombo PC, Ganda A, Lin J, Onat D, Harxhi
A, Iyasere JE, Uriel N and Cotter G: Inflammatory activation:
Cardiac, renal and cardio-renal interactions in patients with the
cardiorenal syndrome. Heart Fail Rev. 17:177–190. 2012. View Article : Google Scholar
|
|
5
|
Colombo PC, Onat D, Harxhi A, Demmer RT,
Hayashi Y, Jelic S, LeJemtel TH, Bucciarelli L, Kebschull M,
Papapanou P, et al: Peripheral venous congestion causes
inflammation, neurohormonal and endothelial cell activation. Eur
Heart J. 35:448–454. 2014. View Article : Google Scholar
|
|
6
|
Wang BW, Chang H, Lin S, Kuan P and Shyu
KG: Induction of matrix metalloproteinases-14 and -2 by cyclical
mechanical stretch is mediated by tumor necrosis factor-alpha in
cultured human umbilical vein endothelial cells. Cardiovasc Res.
59:460–469. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Onat D, Jelic S, Schmidt AM, Pile-Spellman
J, Homma S, Padeletti M, Jin Z, Le Jemtel TH, Colombo PC and Feng
L: Vascular endothelial sampling and analysis of gene transcripts:
A new quantitative approach to monitor vascular inflammation. J
Appl Physiol (1985). 103:1873–1878. 2007. View Article : Google Scholar
|
|
8
|
Colombo PC, Banchs JE, Celaj S, Talreja A,
Lachmann J, Malla S, DuBois NB, Ashton AW, Latif F, Jorde UP, et
al: Endothelial cell activation in patients with decompensated
heart failure. Circulation. 111:58–62. 2005. View Article : Google Scholar
|
|
9
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Spizzo R, Almeida MI, Colombatti A and
Calin GA: Long non-coding RNAs and cancer: A new frontier of
translational research? Oncogene. 31:4577–4587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schonrock N, Harvey RP and Mattick JS:
Long noncoding RNAs in cardiac development and pathophysiology.
Circ Res. 111:1349–1362. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ishii N, Ozaki K, Sato H, Mizuno H, Saito
S, Takahashi A, Miyamoto Y, Ikegawa S, Kamatani N, Hori M, et al:
Identification of a novel non-coding RNA, MIAT, that confers risk
of myocardial infarction. J Hum Genet. 51:1087–1099. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee JH, Gao C, Peng G, Greer C, Ren S,
Wang Y and Xiao X: Analysis of transcriptome complexity through RNA
sequencing in normal and failing murine hearts. Circ Res.
109:1332–1341. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liao Q, Xiao H, Bu D, Xie C, Miao R, Luo
H, Zhao G, Yu K, Zhao H, Skogerbø G, et al: ncFANs: A web server
for functional annotation of long non-coding RNAs. Nucleic Acids
Res. 39(Web Server issue): W118–W124. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liao Q, Liu C, Yuan X, Kang S, Miao R,
Xiao H, Zhao G, Luo H, Bu D, Zhao H, et al: Large-scale prediction
of long non-coding RNA functions in a coding-non-coding gene
co-expression network. Nucleic Acids Res. 39:3864–3878. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Eisen MB, Spellman PT, Brown PO and
Botstein D: Cluster analysis and display of genome-wide expression
patterns. Proc Natl Acad Sci USA. 95:14863–14868. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Witthoft A, Filosa JA and Karniadakis GE:
Potassium buffering in the neurovascular unit: Models and
sensitivity analysis. Biophysical J. 105:2046–2054. 2013.
View Article : Google Scholar
|
|
20
|
Liu T, Jiang CY, Fujita T, Luo SW and
Kumamoto E: Enhancement by interleukin-1b of AMPA and NMDA
receptor-mediated currents in adult rat spinal superficial dorsal
horn neurons. Mol Pain. 9:162013. View Article : Google Scholar
|
|
21
|
Young R and Worthley LI: Current concepts
in the management of heart failure. Crit Care Resusc. 6:31–53.
2004.
|
|
22
|
Liu S, Premont RT and Rockey DC:
G-protein-coupled receptor kinase interactor-1 (GIT1) is a new
endothelial nitric-oxide synthase (eNOS) interactor with functional
effects on vascular homeostasis. J Biol Chem. 287:12309–12320.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cicek FA, Ozgur EO, Ozgur E and Ugur M:
The interplay between plasma membrane and endoplasmic reticulum
Ca(2+) ATPases in agonist-induced temporal Ca(2+) dynamics. J
Bioenerg Biomembr. 46:503–510. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Roy N, Chakraborty S, Paul Chowdhury B,
Banerjee S, Halder K, Majumder S, Majumdar S and Sen PC: Regulation
of PKC mediated signaling by calcium during visceral leishmaniasis.
PloS One. 9:e1108432014. View Article : Google Scholar : PubMed/NCBI
|