Introduction
The heart is one of the first organs to form during
development from the lateral plate mesoderm (1–3). The
primitive heart tube consists of two layers: The endocardium and
the myocardium (4). Progenitor
cells in the lateral plate mesoderm migrate toward the midline,
where they form the two-layered heart tube. Subsequent
morphological changes are necessary to shape this primitive heart
tube into the chambered heart organ and for the atrioventricular
(AV) boundary to develop between chambers that provide directional
flow. In addition to the endocardium and myocardium, a third
cellular layer, the epicardium, develops from an extracardiac
population of cells called the proepicardial organ (PE) (5–12).
The mesodermally-derived epicardium covers the
myocardial layer and is known to provide signaling required for
proper development of the heart (13–15).
However, the molecular regulation of epicardial function is just
beginning to be investigated. Wilm's tumor protein (wt1),
vascular cell adhesion protein 1 and α4 integrin genes
are essential for epicardium formation. Targeted mutagenesis of
these genes in mice provided genetic evidence supporting a role in
cardiac development (16–19). Additional evidence suggested that
the PE contributes to the development of the coronary vessels, and
is required for continued cardiac development and function
(5–11,20,21).
Previous studies indicated that in zebrafish,
similar to other vertebrates, the PE can be distinguished
morphologically at 48 h post fertilization (hpf) as a group of
cells located in close proximity to the ventral wall of the heart
(22,23). The PE is characterized by the
expression of wt1a, transcription factor 21 (tcf21), and
T box 18 (tbx18) at 48 hpf (22,23).
Specification of the PE requires a bone morphogenic protein (BMP)
ligand, which is not, as previously assumed, derived from the liver
bud (23). Instead,
cardiac-specific bmp4 signaling from the myocardium induces
PE specification. Independent from Bmp4 signaling, tbx5
expression in the lateral mesoderm during the early somite stages
is also required for PE specification (23). Previous studies in adult zebrafish
hearts indicate that the epicardium forms a smooth surface covering
the entire heart wall. Additionally, it was discovered that partial
amputation of the adult heart elevated the expression of
tbx18 and retinaldehyde dehydrogenase 2, which
suggests that epicardial genes serve a critical role in the
response to injury (24–26). The activated epicardial cells
proliferate and undergo epithelial-to-mesenchymal transition, which
requires platelet-derived growth factor (27) and β-catenin and retinoic acid
signaling (21). The
developmentally activated epicardial cells quickly invade the
proliferating myocardium at the site of injury and create a dense
vascular network that is likely to encourage regeneration (24–27).
In addition to its role in response to heart injury,
the epicardium has important regulatory functions in the normal
development of the myocardium (17,18,28–31).
Despite the central role of epicardial signaling during heart
development and repair, the molecular signals driving specification
of the PE remain poorly understood.
In chicks, the PE first forms in a bilaterally
symmetrical fashion, and then develops asymmetrically. Disruption
of genes required for specifying right-sidedness in the body, such
as fibroblast growth factor 8 (fgf8) or snail homolog 1
(snail1), prevented PE specification (32). In mice, the dependence of PE
specification on the left-right (LR) body axis is unclear. In this
organism, both PE precursors develop equally on both sides, and
later fuse at the midline to generate the PE (33). Establishment of the LR axis in the
zebrafish heart depends on global LR cues that are generated by
cilia during early gastrulation stages (34). Abnormal LR development can affect
cardiac looping during vertebrate morphogenesis. Therefore, some
cardiac defects that are associated with the abnormal positioning
of the cardiac chambers or with vessel malformation are secondary
to altered LR development (35).
However, some patients demonstrate heterotaxy of the heart without
other obvious LR defects, suggesting that ciliary function may also
have an important role in cardiovascular morphogenesis itself
(36–38). Slough et al (39) reported the presence of mono-cilia
in several areas of the mouse embryonic heart, suggesting a role
for cilia in cardiac morphogenesis. Furthermore, hearts in
kinesin-3a (Kif3a) mutants developed abnormal
endocardial cushions (ECCs) and thinner compact myocardium, and
completely lacked cardiac cilia (39). Additionally, the compact myocardium
was thinner in embryos mutant for polycystic kidney disease 2, a
protein that functions as a mechanosensor in the kidney and node
(39).
The WT1-interacting protein (Wtip) was originally
identified as an interacting partner of WT-1 in a yeast two-hybrid
screen (40). Recent studies and
previous work by our group revealed the following roles for Wtip:
i) Wtip interacts with the C-terminus of receptor tyrosine
kinase-like orphan receptor 2 in yeast and mammalian cells
(41); ii) Ajuba LIM proteins
[Ajuba, LIM domain containing 1 (LIMD1), and Wtip] interact with
Snail to remodel epithelial dynamics (42,43);
iii) Wtip is a LIM domain protein of the Ajuba/Zyxin family, which
is enriched in the basal bodies of cells in the pronephros and
Kupffer's vesicle in zebrafish (44). In zebrafish, wtip deletion
leads to pronephric cysts, hydrocephalus, body axis curvature and
pericardial edema (44); iv)
cryptic deletion in the human wtip gene was reported to
cause hypospadias, accompanied by congenital heart disease
(45), but the roles for Wtip in
the heart are unknown. Since Wtip plays a role in ciliopathy
(44), interacts with wt1
(40), which is expressed in the
PE, and Wtip depletion leads to pericardial edema (44), we hypothesized that Wtip function
in the heart may be associated with PE specification and may be
involved in LR patterning.
In the present study, we show that cardiac
expression of wtip mirrors wt1a, tcf21, and
tbx18 expression, confirming that wtip is expressed
in the PE. This is the first report, to the best of our knowledge,
to describe how Wtip, a basal body protein, is important for PE
specification, heart looping, AV formation and LR patterning.
Materials and methods
Zebrafish maintenance
All animal experiments were performed in strict
accordance with the recommendations in the Guide for the Care and
Use of Laboratory Animals of the National Institutes of Health and
were approved by the Institutional Animal Care and Use Committee of
the University of Oklahoma Health Sciences Center. Danio
rerio (AB strain) were maintained and raised at 28.5°C under a
14-h light/10-h dark cycle. Zebrafish embryos were kept in 0.5X E2
egg medium (7.5 mM NaCl, 0.25 mM KCl, 0.5 mM CaCl2, 0.5
mM MgSO4, 0.075 mM KH2PO4, 0.025
mM Na2HPO4, 0.35 mM NaHCO3, 0.01%
methylene blue). To suppress pigmentation of zebrafish embryos,
0.0045% 1-Phenyl-2-thiourea (Sigma-Aldrich, St. Louis, MO, USA) was
added to egg medium as needed. Embryos and larvae were staged
according to h post-fertilization (hpf) or days post-fertilization
(dpf) (46).
Morpholino and mRNA injections
A translational blocking morpholino oligonucleotide
(MO) targeted against the 5′UTR of wtip (wtipMO;
5′-GATCCTCGTCGTATTCATCCATGTC-3′; 44), wt1a (wt1aMO:
5′-GAGCAAGAGATACTGACCTGAAGGC-3′; 22), and randomized control
MO (conMO: 5′-CCTCTTACCTCAGTTACAATTTATA-3′) were
obtained from Gene Tools, LLC (Philomath, OR, USA). A volume of 4.6
nl with a concentration of 0.225 mM wtipMO, 0.056 mM
wtipMO, 0.25 mM wt1aMO, 0.0625 mM wt1aMO, and
92 pg wtip mRNA was injected at the one-cell stage using a
nanoliter 2000 microinjector (World Precision Instruments, Inc.,
Sarasota, FL, USA). Zebrafish wtip mRNA was synthesized with
T7 RNA polymerase (Thermo Fisher Scientific, Inc., Waltham, MA,
USA), after linearization of the
pCR®-BluntII-TOPO®-wtip construct with
HindIII (Thermo Fisher Scientific, Inc.). All mRNAs were
purified using the RNeasy Mini Kit (Qiagen, Inc., Valencia, CA).
Microinjections into 1-cell embryos were performed as described by
Feng et al (47). To
knockdown wtip specifically in dorsal forerunner (DFCs),
wtipMO was injected into the yolk cell of the ~1,000-cell
stage embryos as previously described (48).
In situ hybridization
Whole-mount in situ hybridization was performed as
previously described (49,50), using the following probes: wtip,
wt1a, tcf21, tbx18, bmp4, LR determination factor 1 (lefty1),
lefty2, southpaw (spaw), cardiac myosin light chain 2 (cmlc2),
α-cardiac myosin heavy chain (amhc), ventricular MHC (vmhc) and
natriuretic peptide A (nppa) (34,44,51–54).
Polymerase chain reaction (PCR) DNA templates were used for wt1a,
tcf21, bmp4, lefty1, lefty2, spaw, cmlc2, amhc, vmhc and nppa. DNA
templates were used for wtip and tbx18. We used the following
primers to prepare the PCR DNA template for reverse transcription
(RT)-PCR and 2nd PCR for wt1a (765 bp): wt1a-51F1
5′-CCGGTGGAAACGGTAACTGTA-3′; wt1a-1161R1
5′-TCTGCAGTTGAAGGGCTTCTC-3′; wt1a-240F2
5′-GCACTTCTCCGGACAGTTCAC-3′; and wt1a-1004R2T7
5′-GGTAATACGACTCACTATAGGGAGAACCTGCGACCACAGTCT-3′. For tcf21 (682
bp), we used the following primers: tcf21-68F1
5′-TCATCTCCACGTCCAGTCAGA-3′; tcf21-876R1
5′-CACACTGTTGCCTTGAACCAG-3′; tcf21-124F2 5′-TCTCCACTCCACCCTTGT
CTC-3′; and tcf21-805R2T7
5′-GGTAATACGACTCACTATAGGATGCGAGTGAGGATGTTGTCC-3′. The primers used
for left1 (875 bp) were: lefty1-83F1 5′-ACGCTCTGCTGAAGAAACTGG-3′;
lefty1-1065R2 5′-AATATTGTCCATTGCGCATCC-3′; lefty1-153F2
5′-GATCCCAACGCACGTAAAGAA-3′; and lefty1-1027R2T7
5′-GGTAATACGACTCACTATAGGTGTTTGGGAATTCAGCCACTT-3′. Primers for
lefty2 (807 bp) were: lefty2-31F1 5′-ACCACAGCGATCTCACTCACA-3;
lefty2-1029R1 5′-TTCTGCCACCTCGATTTCAGT-3′; lefty2-35F2
5′-CAGCGATCTCACTCACACAGG-3′; and lefty2-841R2T7
5′-GGTAATACGACTCACTATAGGTGAGCTCACGGAAGTTGATGA-3′. For spaw (656
bp), we used the following primers: spaw-210F1
5′-TAGTGTTGACAACCCGGCTCT-3′; spaw-1200R1
5′-CTCCTCCACGATCATGTCCTC-3′; spaw-461F2
5′-AACTGTTTCTGGGCAGCGTTA-3′; and spaw-1116R2T7
5′-GGTAATACGACTCACTATAGGCACCACTCCATCCCCTTCATA-3′. The primers for
bmp4 (851 bp) were: bmp4-76F1 5′-CTGATACCCGAGGAAGGGAAG-3′;
bmp4-1068R1 5′-CACCGAGTTCACCAGTGTCTG-3′; bmp4-84F2
5′-CGAGGAAGGGAAGAAGAAAGC-3′; and bmp4-934R2T7
5′-GGTAATACGACTCACTATAGGCGTCGCTGAAATCCACATACA-3′. For myl7 (440
bp), we used: myl7-25F1 5′-AAGAGGGGGGAAACTGCTCAA-3′; myl7-518R1
5′-CAAGATTCCTCTTTTTCATCACCA-3′; myl7-27F2 GAGGGGGAAAACTGCTCAAAG-3′;
and myl7-466R2T7 GGTAATACGACTCACTATAGGAATCAATATTTCCAGCCACGTC. The
following primers were used for amhc (939 bp): amhc-4486F1
5′-AGGCTCGCAGTTTAAGCACTG-3′; amhc-5651R1
5′-CAAATTCTTGCGATCCTCGTC-3′; amhc-4614F2
5′-GTAAGCGAAGGGCGAAAGAGT-3′; and amhc-5552R2T7
5′-GGTAATACGACTCACTATAGGAGCGTCCAGCTCATTCTCAAG-3′. Primers for vmhc
(903 bp) were: vmhc-3141F1 5′-GGCAAAAGCAAAGCTAGAGCA-3′; vmhc-4276R1
5′-CGGTTTCCTGTAAGCGTTGAG-3′; vmhc-3258F2
5′-AACCCAGGAAAGCCTAATGGA-3′; and vmhc-4160R2T7
5′-GGTAATACGACTCACTATAGGGACATGCCTCGCTGTAGCTCT-3′.
All digoxigenin-UTP-labeled anti-sense RNA probes
were synthesized using DIG-RNA (Sigma-Aldrich) labeling mix and T7
RNA polymerase (PCR DNA templates for wt1a, tcf21,
bmp4, lefty1, lefty2, spaw, cmlc2,
amhc, vmhc and nppa) or SP6 RNA polymerase (plasmid
templates for wtip and tbx18; New England BioLabs,
Inc., Ipswich, MA, USA), according to the manufacturer's
instructions. Alkaline phosphatase-conjugated anti-digoxigenin
(Sigma-Aldrich) was used to localize the probes (50). NBT/BCIP (Sigma-Aldrich) was used as
the chromogenic substrate to produce blue precipitate. The
zebrafish tbx18 DNA template was subcloned to
pCR-BluntII-TOPO vector (Thermo Fisher Scientific, Inc.). Primers
used to prepare DNA template for RT-PCR and 2nd PCR for
tbx18 (828 bp) were as follows: tbx18-14F1
5′-GACGGTCCCCGTGTACTATGA-3′; tbx18-997R1
5′-TCCCAAGGATGTCCTCAAATG-3′; tbx18-91F2
5′-AAATTGGAGGAGGAGGACAGC-3′; and tbx18-918R2
5′-CATTCTGTTTCGTCCCGAGTC-3′. Finally, tbx18 plasmid was
linearized by XhoI and SP6 RNA polymerase, and wtip
plasmid was linearized by NotI and SP6 RNA polymerase
(44).
Cardiac looping assessments
The extent of cardiac looping was assessed in
wtip morphants and buffer-injected control embryos using the
Tg(myl7:EGFP) line. At 48 hpf, embryos were fixed in 4%
formaldehyde, mounted in 1% agarose and were optimally positioned
for cardiac imaging. The looping angle (LA) was defined as the
angle between the anterior/posterior axis of the embryo and the
cross-sectional plane of the AV junction (55). Student's t-test using an α of 0.05
was used to evaluate differences in looping angle means (n=10
hearts/treatment). Statistical analysis was performed using
Microsoft Excel 2016 (Microsoft Corporation, Redmond, WA, USA).
Antibodies and whole-mount
immunohistochemistry
A previously published protocol was followed
(44,56). Primary antibodies were: Custom-made
anti-Wtip (1:100 dilution; Covance, Inc., Denver, PA, USA),
anti-γ-tubulin (GTU-88; 1:800 dilution; Sigma-Aldrich),
anti-acetylated α-tubulin (6-11B-1; 1:800 dilution; Sigma-Aldrich),
and anti-centrin (20H5; 1:1,000 dilution; EMD Millipore, Billerica,
MA, USA). The following secondary antibodies were obtained from
Thermo Fisher Scientific, Inc.: Goat anti-rabbit Alexa
Fluor®546 (IgG [H+L]), goat anti-mouse Alexa
Fluor®488 (IgG2b), goat anti-mouse Alexa
Fluor®488 [IgG1 (γ1)], goat anti-mouse Alexa
Fluor®488 [IgG2a (γ2a)] and goat anti-mouse Alexa
Fluor®546 [IgG1 (γ1)]. Whole-mount immunohistochemistry
samples were dehydrated with a graded series of methanol, embedded
in JB4 resin (Polysciences, Inc., Warrington, PA, USA), and were
cut into 5–7 µm sections using an RN2255 microtome (Leica
Technology, Exton, PA, USA). The sections were stained with DAPI
(Kirkegaard & Perry Laboratories, Inc., Gaithersburg, MD, USA),
mounted in Fluorescent Mounting Media (Kirkegaard & Perry
Laboratories, Inc.), and were imaged with a FV-1000 confocal
laser-scanning microscope (Olympus America, Inc., Center Valley,
PA, USA).
Histological analysis
Embryos were fixed with histology fixative [1.5%
glutaraldehyde, 4% formaldehyde, 3% sucrose in 0.1 mM phosphate
buffer (PB, pH 7.3)] overnight at 4°C. Fixed embryos were then
dehydrated with a graded series of methanol and embedded in JB4
resin (Polysciences, Inc., Warrington, PA, USA). Sections (4 µm)
were cut with an RN2255 microtome (Leica Technology) and were
stained with Harris hematoxylin and special eosin II (BBC
Biochemical, Mount Vernon, WA, USA). Once the sections were mounted
in Polymount (Polysciences, Inc.), the stained sections were imaged
with a Provis AX-70 microscope (Olympus America, Inc.) equipped
with a RETIGA EXi digital camera (QImaging, Surrey, Canada).
Results
Wtip expression in the cardiac region is
identical to PE marker genes
WT1 is expressed in the PE and glomerulus
podocytes in mammals, birds, zebrafish and medaka (22,57–61).
In addition to wt1a gene expression in zebrafish PE
(22), the expression patterns of
other PE markers, including tcf21 and tbx18, are well
documented at 48 hpf, 57 hpf, and 4 days post-fertilization (dpf)
(22,23). It was previously demonstrated that
zebrafish embryos depleted for wtip via anti-sense
morpholino (wtipMO) developed pericardial edema (44).
To investigate possible roles for Wtip in PE
specification and development, whole-mount in situ
hybridization was used (Fig. 1).
Wtip mRNA was detected at 48 hpf (Fig. 1A) and 4 dpf (Fig. 1E) in zebrafish embryonic hearts. At
48 hpf, wtip-positive cells at the pericardial surface of
the yolk were observed to be broadly dispersed at the AV-to-sinus
venous region (Fig. 1A, yellow
arrowhead marks the AV junction). By 4 dpf, wtip-positive
cells appeared to have spread over the heart to cover the
myocardium (Fig. 1E, yellow
arrowhead). To determine the identity of the cells expressing
wtip, the common PE marker genes wt1a (Fig. 1B and F, yellow arrowheads),
tcf21 (Fig. 1C and G,
yellow arrowheads) and tbx18 (Fig. 1D and H, yellow arrowheads) were
evaluated at 48 hpf (Fig. 1A–D)
and 4 dpf (Fig. 1E–H). At 48 hpf,
wt1a, tcf21 and tbx18 expression was detected at the
level of the sinus venous and adjacent to the AV junction (Fig. 1B–D, yellow arrowheads). At this
stage, cardiac expression typically appears punctate, as would be
expected for the isolated clumps of PE, which have not yet formed a
uniform layer. In addition, tcf21 expression occurred in the
pharyngeal arch (Fig. 1C, black
arrows), and tbx18 expression occurred in the pectoral fin
(Fig. 1D, asterisk). By 4 dpf, the
wt1a-, tcf21- and tbx18-positive cells spread
over the heart to cover the myocardium (Fig. 1F–H, yellow arrowheads). Therefore,
the wtip-positive cells exhibited an expression pattern
similar to the known PE marker genes wt1a, tcf21 and
tbx18, and corresponded in physical appearance to
extracardiac cell populations, indicating their PE identity
(22,23).
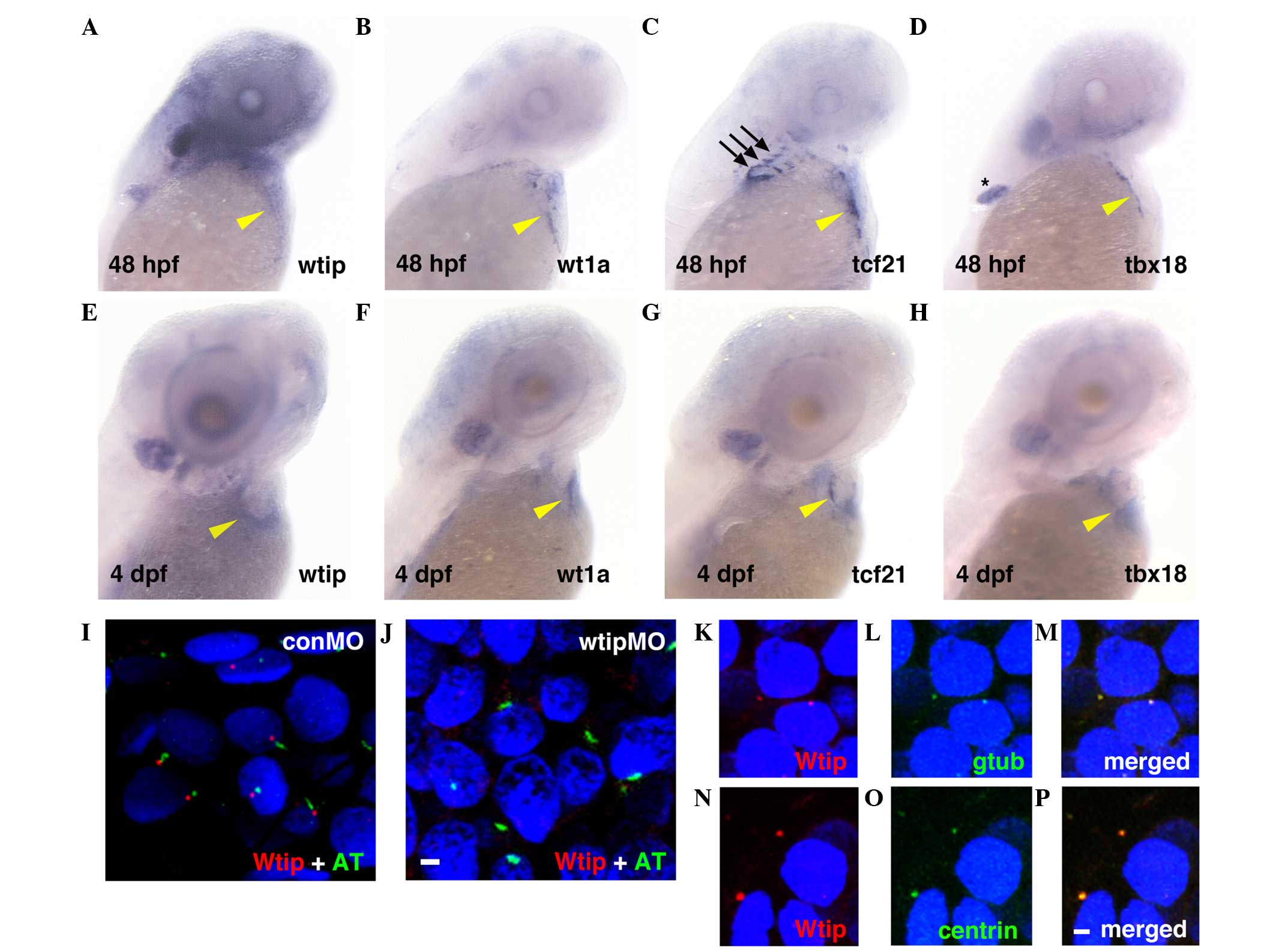 | Figure 1wtip expression is identical
to PE-specific markers during zebrafish heart development. Lateral
view of (A–D) 48 hpf and (E–H) 4 dpf whole-mount in situ
hybridization of (A,E) wtip, (B,F) wt1a, (C,G)
tcf21 and (D,H) tbx18. At 48 hpf, (A) wtip,
and PE markers (B) wt1a-, (C) tcf21- and (D)
tbx18- positive cells were in the clusters near the sinus
venosus, between the atrium and the yolk and near the AV junction,
in contact with the ventral surface of the heart (yellow arrowhead
in A–D). At 48 hpf, tcf21 expression occurred in the
pharyngeal arch (black arrows in C) and tbx18 expression
occurred in the pectoral fin (black asterisk in D). By 4 dpf, the
wt1a-, tcf21- and tbx18-positive cells spread
over the heart to cover the myocardium (yellow arrowhead in F–H).
By 4 dpf, wtip-, tcf21- and tbx18-positive
cells appeared to spread over the heart to cover the myocardium
along the dorsal surface of the heart (yellow arrowhead in E–H).
(I,J) Double immunofluorescence for the cilia marker acetylated
α-tubulin (green) and Wtip (red) in confocal projections of the
zebrafish PE at 48 hpf. (J) No Wtip signal was detectable in
wtipMO knockdown embryos, confirming antibody specificity.
(K–P, red) Localization of Wtip in the basal bodies of cilia was
confirmed by double immunostaining with Wtip antibody and either
(L,M, green) anti-γ-tubulin or (O,P, green) anti-centrin, which are
basal body markers in heart. 4′,6-diamidino-2-phenylindole was used
to counterstain nuclei (blue). Scale bar=10 µm. wtip, Wilms tumor 1
interacting protein; PE, proepicardial organ; hpf, hours
post-fertilization; dpf, days post fertilization; wt1a,
Wilms tumor 1a; tcf21, transcription factor 21;
tbx18, Tbox 18; wtipMO, wtip morpholino oligonucleotide;
conMO, control MO; AT, acetylated α-tubulin. |
To further assess a possible cell autonomous role
for Wtip in the PE, the subcellular localization of this protein in
the PE was examined using a zebrafish-specific anti-Wtip antibody
(44). In a previous study, Wtip
protein was observed to be enriched in the basal body, a structure
that resides at the base of cilia in the pronephros, Kupffer's
vesicle (KV) and other ciliated tissues (44). While cilia have not been reported
in the zebrafish embryonic heart, they have been discovered in
mouse (39) and chick hearts
(62). Accordingly, the
possibility that Wtip may localize to the basal body at the base of
cilia in the zebrafish embryonic heart was considered. Therefore,
double immunostaining was performed with antibodies against
zebrafish Wtip and the cilia marker acetylated α-tubulin (Fig. 1I and J), or the basal body marker
γ-tubulin (Fig. 1K–M) and basal
body marker centrin (Fig. 1N–P),
in 48 hpf PE. Sagittal section staining suggested that Wtip was
localized to the basal body of cilia located on PE in the 48 hpf
embryos (Fig. 1I–P). The
specificity of the localization pattern was verified by knockdown
of endogenous wtip expression with wtipMO (44), which abolished Wtip immunostaining
(Fig. 1J). Together, these data
provide the first evidence, to the best of our knowledge, that the
zebrafish embryonic heart develops cilia and basal bodies (Fig. 1I–P). In addition, these data
indicate that Wtip is expressed in the basal bodies of cells in the
PE of the zebrafish embryonic heart.
Wtip signaling is required for PE
specification
wt1 null mutant mice develop cardiac
abnormalities (16), and the
epicardium fails to form properly (19). The wt1 gene is expressed in
both the epicardial lineage and in kidney podocytes, and this
pattern is well conserved in mammals, zebrafish and medaka
(22,59,61,63).
To test whether Wtip is required for PE formation and specification
during zebrafish embryonic heart development, a wtip
morpholino antisense oligonucleotide (MO) was used to inhibit
translation of wtip transcripts (44). First, the PE formation in 48 hpf
wtip knockdown embryos was examined (Fig. 2) using the PE-specific markers
tcf21 (Fig. 2B, C and G)
and tbx18 (Fig. 2E, F and
H). Injection of wtipMO produced embryos with body axis
curvature, pronephric cysts, hydrocephalus and pericardial edema.
Approximately 99% of injected embryos (n=64) developed
pericardial edema. In wtip knockdown embryos, the expression
of tcf21 and tbx18 in the heart was found to be
significantly reduced (25/36, 69.4% for tcf21 and 14/27,
51.9% for tbx18; Fig. 2B, E, G
and H, yellow arrowheads) or absent (11/36, 30.6% for
tcf21 and 13/27, 48.1% for tbx18; Fig. 2C, F, G and H, yellow arrowheads).
By contrast, despite modest developmental delay (2 h) in these
embryos, tcf21 expression in the pharyngeal arch (Fig. 2B and C, black arrows) and
tbx18 expression in the pectoral fin were unaffected
(Fig. 2E and F, asterisk). These
data indicate that Wtip signaling is essential for early epicardial
development and is required to specify the PE.
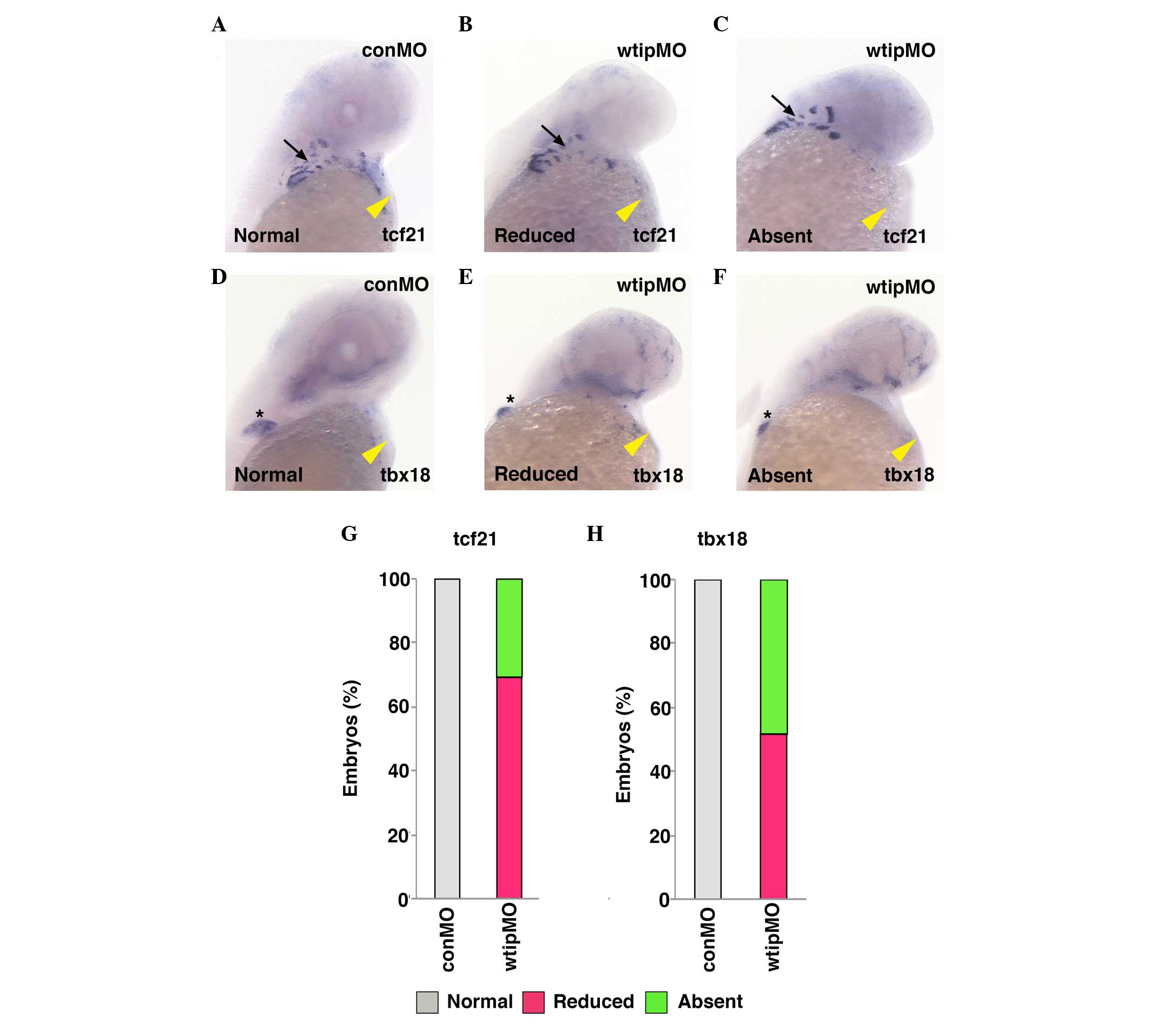 | Figure 2Wtip is required for PE
specification. Ventral view of 48 hpf whole-mount in situ
hybridization of (A–C) tcf21 and (D–F) tbx18 in
(A,D,G,H)control (1.035 pmol/embryo) and (B,C,E–H) wtip
morphant embryos (1.035 pmol/embryo). Both tcf21 and
tbx18 express specifically in the PE in the control embryos
(yellow arrowhead in A, D), but is reduced or absent in wtip
morphant embryos. tcf21 expression in the pharyngeal arch
(black arrow in A–C) and tbx18 expression in the pectoral
fin (black asterisk in D–F) in wtip morphants are
unaffected. wtip, Wilms tumor 1 interacting protein; PE,
proepicardial organ; hpf, hours post fertilization; tcf21,
transcription factor 21; tbx18, Tbox 18; conMO, control
morpholino oligonucleotide. |
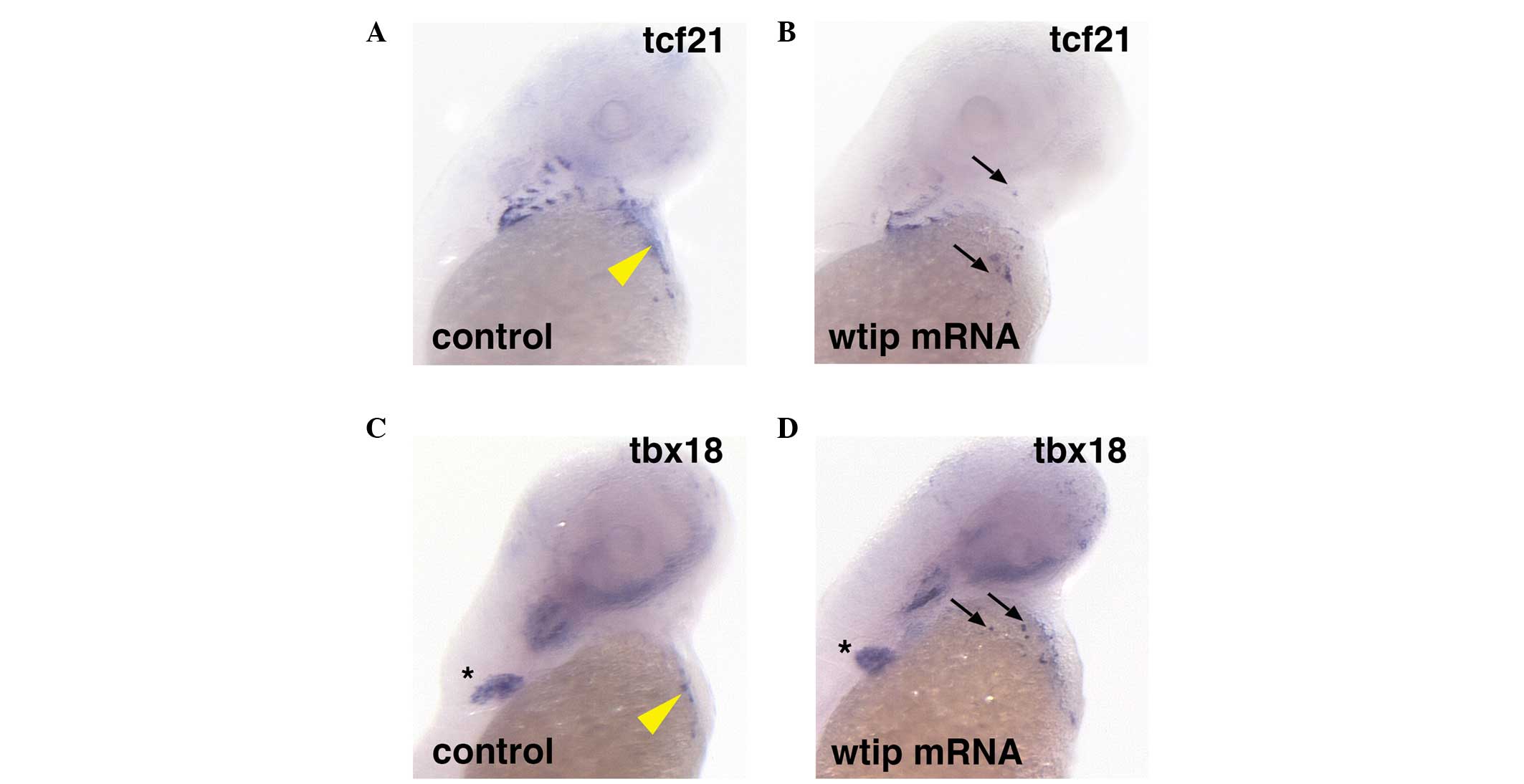
To determine whether Wtip signaling is sufficient
to induce PE marker gene expression, wtip mRNA was
overexpressed (Fig. 3). In embryos
with ubiquitous wtip mRNA expression, ectopic tcf21
(34/34, 100%; Fig. 3B, black
arrows) and tbx18 (31/31, 100%) expression was observed in
the heart region (Fig. 3D, black
arrows) compared with control tcf21 (n=26; Fig. 3A) and tbx18 (n=26;
Fig. 3C) expression in the PE
region of the heart (yellow arrowheads indicate sinus venous). In
all of the wtip mRNA-overexpressing embryos, ectopic
clusters of the tcf21- and tbx18-positive cells were
observed, which appeared more widespread and numerous than the
clusters seen in wild-type embryos. Occasional ectopic clusters
were located in or near the craniofacial area (Fig. 3B and D, black arrows), and were
consistently observed in the cardiac region. Overexpression of
wtip mRNA in embryos did not affect tbx18 expression
in the pectoral fin (Fig. 3C and
D, black asterisk).
Wtip and wt1a functionally interact
during PE development
Mouse wtip was originally identified as an
interacting partner of wt1 in a yeast two-hybrid screen
(40). However, the implications
of the potential interation between Wtip and WT1 in PE formation
have not been reported. The co-expression of wt1a (Fig. 1B and F, yellow arrowhead; 22) and
wtip (Fig. 1A and E, yellow
arrowhead) in PE control embryos, along with the similar cardiac
phenotypes generated by knockdown of either gene, support the
notion that these two genes may function together in zebrafish PE
formation. In wt1a morphants (Fig. 4), the expression of PE markers was
reduced (7/47, 14.9% for tcf21, Fig. 4M; and 3/33, 9.1% for tbx18;
Fig. 4N) or absent (40/47, 85.1%
for tcf21, Fig. 4G and M,
black arrows; and 30/33, 90.9% for tbx18; Fig. 4M and N, black arrows). As described
(Fig. 2B, C, E–H), in wtip
morphants, PE markers were similarly reduced (21/30, 70% for
tcf21, Fig. 4M; and 13/25,
52% for tbx18, Fig. 4N) or
absent (9/30, 30% for tcf21, Fig. 4C and M, black arrow; and 12/25, 48%
for tbx18, Fig. 4D, and N,
black arrow). To further investigate the potential for Wtip and WT1
interaction, a combined knockdown of wtip and wt1a
was performed using sub-threshold doses of both morpholinos to
assess whether a genetic interaction between these proteins would
alter the expression of the PE markers tcf21 and
tbx18. Prior to this, the appropriate sub-threshold doses
for each morpholino was determined: i) wt1aMO, n=28
for tcf21 (Fig. 4I and M,
yellow arrowhead) and n=27 for tbx18 (Fig. 4J and M, yellow arrowhead) and ii)
wtipMO, n=36 for tcf21 (Fig. 4E and M, yellow arrowhead) and
n=35 for tbx18 (Fig. 4F
and M, yellow arrowhead). Sub-threshold doses for either
morpholino alone had no effect on tcf21 (Fig. 4E, I and M, yellow arrowhead) or
tbx18 (Fig. 4F, J and N,
yellow arrowhead) expression in the PE: wt1aMO, 26/28, 92.9%
for tcf21 (Fig. 4I and M,
yellow arrowhead) and 25/27, 92.6% for tbx18 (Fig. 4J and N, yellow arrowhead);
wtipMO, 34/36, 92.9% for tcf21 and 33/35, 94.3% for
tbx18 (Fig. 4E, F, M and N,
yellow arrowhead). However, in wtip and wt1a double
morphants, tcf21 and tbx18 expression was severely
reduced (3/27, 11.1% for tcf21, Fig. 4M; and 3/32, 9.4% for tbx18,
Fig. 4N) or absent (24/32, 88.9%
for tcf21, Fig. 4K and M,
black arrow; and 29/32, 90.5% for tbx18, Fig. 4L and N, black arrow) in the cardiac
region. These data indicate that Wtip is genetically associated
with Wt1a, and that together they influence PE specification.
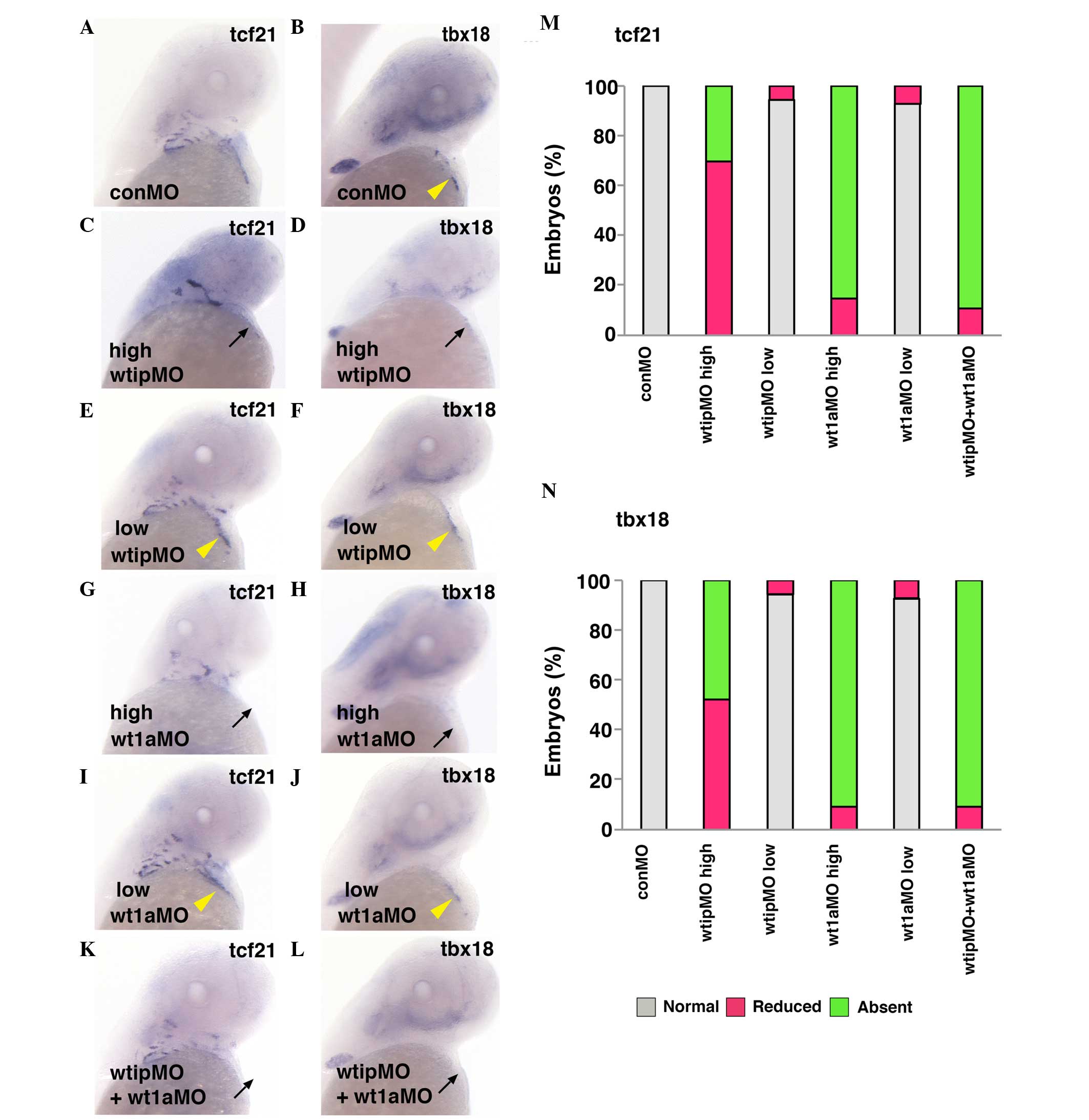 | Figure 4Wt1a and Wtip cooperate for PE
formation. Ventral view of 48 hpf whole-mount in situ
hybridization of (A,C,E,G,I,K) tcf21 and (B,D,F,H,J,L)
tbx18 expression in control (yellow arrowhead, A,B), high
concentration of wtipMO (black arrow, C,D; 1.035
pmol/embryo), sub-threshold concentration of wtipMO (yellow
arrowhead, E,F; 0.259 pmol/embryo), high concentration of
wt1aMO (black arrow, G,H; 1.15 pmol/embryo), sub-threshold
concentration of wt1aMO (yellow arrowhead, I,J;
wt1aMO, 0.288 pmol/embryo), sub-threshold concentrations of
wt1aMO (1.15 pmol/embryo)+wtipMO (0.259 pmol/embryo;
black arrow, K,L) in PE at 48 hpf. With high concentrations of
wt1aMO, PE marker expressions were reduced or absent (black
arrow, for tcf21; G,M, and for tbx18; H,N). With
wtipMO, PE marker expressions were reduced (for
tcf21, M and for tbx18, N) or absent (black arrow,
for tcf21; C,M, and for tbx18; D,N). Sub-threshold
doses for each MO had no effect on tcf21 (yellow arrowhead,
E,I,M) or tbx18 (yellow arrowhead, F,J,N) expression in PE.
In the wtip and wt1a double morphants, tcf21
and tbx18 were severely reduced (black arrow, for
tcf21; K,M and for tbx18; L,N) or no longer expressed
(for tcf21 and for tbx18) in the cardiac region.
These data indicate that Wtip is associated with Wt1a and
influences PE specification. Wt1a, Wilm's tumor 1a; Wtip, Wilm's
tumor 1 interacting protein; PR, proepicardial organ; hpf, hours
post fertilization; tcf21, transcription factor 21;
tbx18, Tbox 18; wtipMO, wtip morpholino oligonucleotide. |
Wtip is required for cardiac looping and
valve formation, but not for chamber patterning
Cardiac looping is a morphogenic process in
vertebrates. This process re-shapes the linear heart tube by
bending the cardiac chambers into close juxtaposition. In
zebrafish, cardiac looping begins around 30 hpf as the heart tube
folds, gradually bringing the atrium and ventricle into a
side-by-side position by 48 hpf (15,37).
The continued bending of the heart tube, accompanied by chamber
ballooning (~48–58 hpf) (64), and
concentric growth within the chambers (65), substantially remodels the shape of
the heart by 72 hpf. Functionally, the organ changes rapidly over
this period, as heart rate and cardiac output increase steadily
(3,66,67).
Blood flow, which initially incorporates some back-flow through the
AV junction, becomes consistently unidirectional as endocardial
cushions and AV valves develop (68). Transgenic Tg(myl7:EGFP)
embryos, which express GFP in the embryonic heart, provided
easy visualization of the heart (Fig.
5A and B). As a simple rubric to describe the progression of
cardiac looping in wtip morphants, the cardiac 'looping
angle' was measured, defined as the degree of difference in the
anterior/posterior (A/P) body axis and the plane of the AV junction
(Fig. 5C) (55). At 48 hpf, control hearts exhibited
an average looping angle of 16°, whereas looping angles in embryos
injected with wtipMO were significantly larger, indicating
that they were less looped (Fig.
5A–D). These data indicate that cardiac looping was impaired by
Wtip depletion.
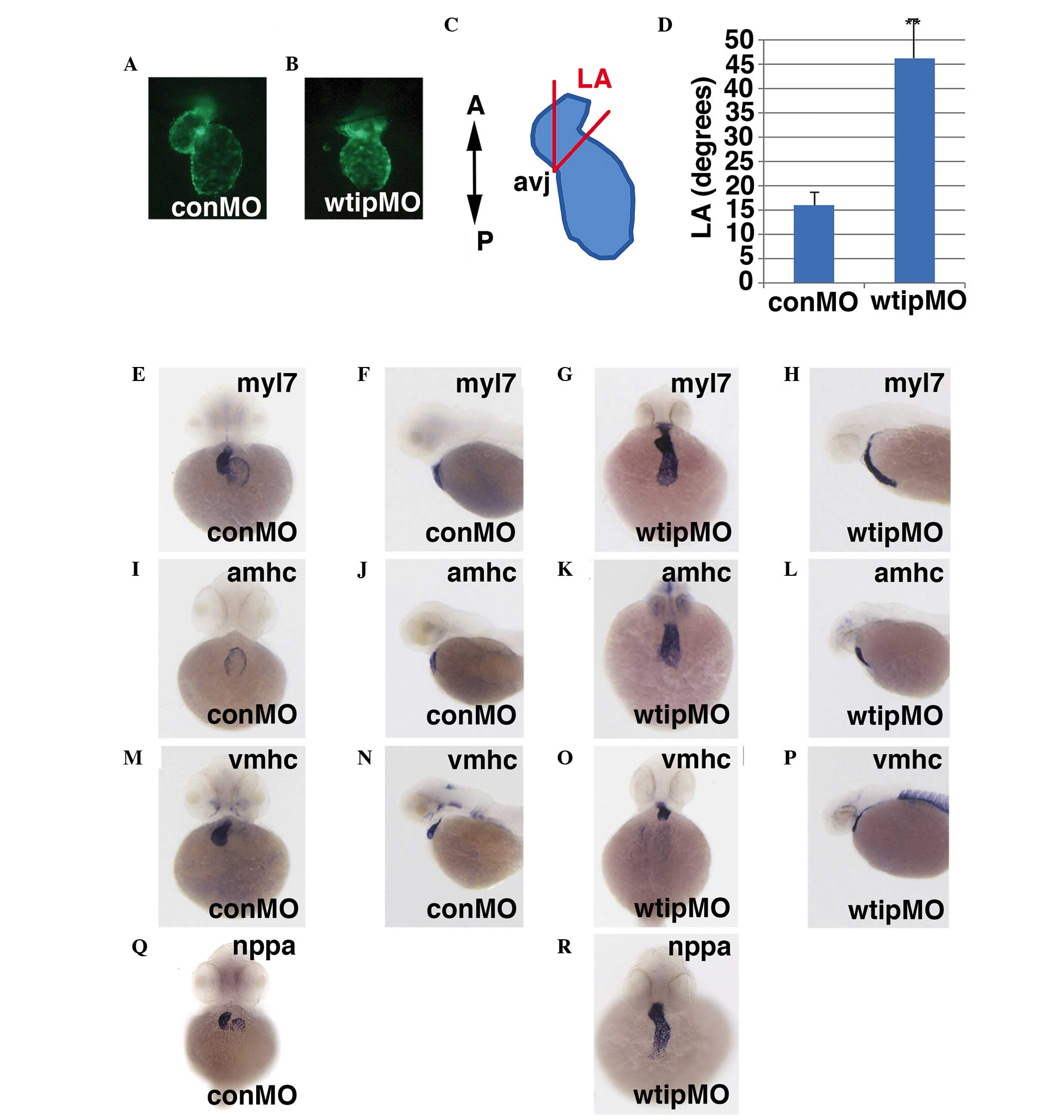 | Figure 5Wtip is required for AV boundary
formation, but does not affect chamber patterning. Hearts of
transgenic Tg(myl7:EGFP) embryos were visualized in 48 hpf
(A) control or (B) wtip morphant embryos (1.035 pmol/embryo)
to assess the extent of cardiac looping. (C) Diagram depicting the
cardiac LA, defined as the degree of difference in the
anterior/posterior body axis and the plane of the AV junction. (D)
Wtip depletion decreases the mean LA by 48 hpf. The mean ± standard
error is shown. *P<0.000359. n=10
embryos/treatment. 48 hpf whole-mount in situ hybridization
in (E,F,I,J,M,N,Q) control and (G,H,K,L,O,P,L) wtip morphant
embryos (1.035 pmol/embryo) of (E–H) myl7, (I–L)
amhc, (M–P) vmhc and (Q,R) nppa.
(E,G,I,K,M,O,Q,R) Ventral view and (F,H,J,L,N,P) lateral view. Wtip
does not regulate chamber patterning, but regulates nppa
expression to maintain chamber myocardium expression and exclusion
from the myocardium of the AV boundary. Wtip, Wilm's tumor 1
interacting protein; AV, atrioventricular; myl7, myosin
light chain 7; EGFP, enhanced green fluorescent protein; hpf, hours
post fertilization; LA, looping angle; amhc, α myosin heavy
chain; vmhc, ventricular myosin heavy chain; nppa,
natriuretic peptide A; conMO, control morpholino oligonucleotide;
A, anterior; P, posterios; avj, AV junction. |
Next, it was examined whether the cardiac looping
defect of wtip morphants was due to improper establishment
of cardiac patterning. At 48 hpf, wtip morphants showed
normal expression of the cardiac chamber-specific markers
myl7 (cmlc2) (Fig.
5E–H), amhc (Fig. 5I–L)
and vmhc (Fig. 5M–P). By 48
hpf, hearts in wild-type embryos have initiated cardiac looping,
which shifts the ventricle to the right of the atrium (Fig. 5E, I and M). In the process of
checking the heart patterning, a role for Wtip in differentiating
the AV boundary was identified. At 52 hpf, nppa was detected
in both the ventricle and atrium but was absent from the AV
boundary in wild-type embryos (Fig.
5Q, black arrows). Conversely, nppa was detected
throughout the heart, with no distinguishable exclusion from the AV
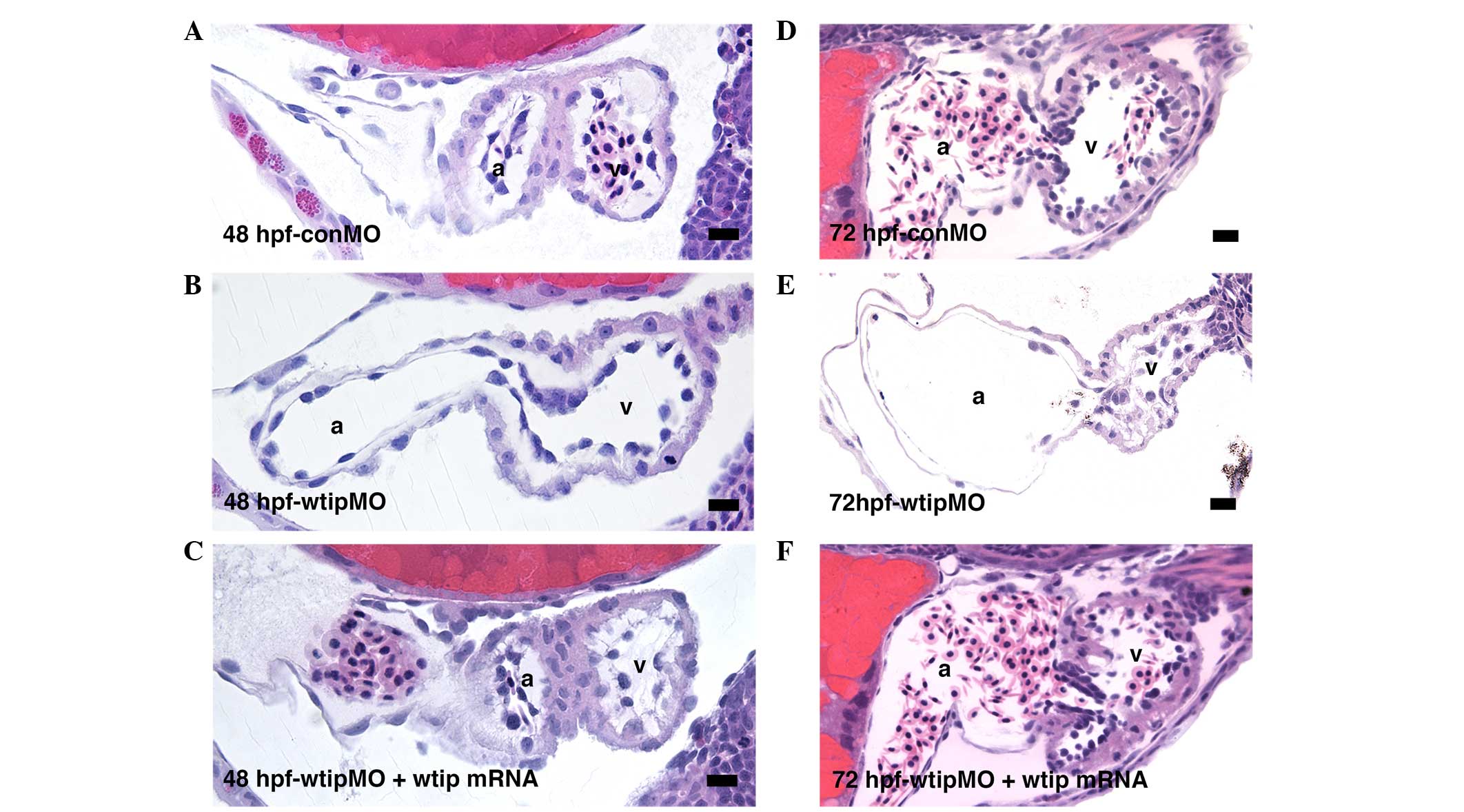
boundary in wtip knockdown embryos (Fig. 5R). Analysis of sections (Fig. 6) revealed a thinned myocardial
layer in the atrium, and increased separation of the myocardium and
endocardium in the ventricle (Fig. 6B
and E). Rather than transitioning to the expected cuboidal
shape, AV endocardial cells in wtip knockdown embryos
retained a squamous appearance at 72 hpf, and clear leaflets did
not form (Fig. 6E). Importantly,
the lack of AV boundary, thinned myocardial layer in the atrium,
and increased separation of the myocardium and endocardium in the
ventricle phenotype of wtip morphants could all be rescued
by the co-injection of wtip mRNA, demonstrating that the
phenotypes specifically result from wtip knockdown (Fig. 6C and F).
Knockdown of wtip results LR asymmetry
defects
Ajuba LIM proteins (LIMD1, WTIP, AJUBA) are
categorized as a family of LIM domain proteins, however, their
functional similarities remain unclear. Embryos depleted for the
Ajuba homolog in medaka (69) and
zebrafish (70) exhibited
developmental abnormalities, including LR patterning defects. It
has been previously shown by in situ hybridization that
wtip is expressed in a broad range of tissues during the
gastrula period and early somitogenesis (44), including KV, which is equivalent to
the mammalian node in teleosts (71–73).
Furthermore, Wtip protein expression was investigated by
immunostaining using a zebrafish-specific Wtip antibody, which
localized to the basal bodies of pronephros, KV and ciliated
tissues (44). The wild-type heart
typically 'jogs' to the left and subsequently loops to the right
(34). To investigate the
establishment of the LR axis within the forming zebrafish heart
tube, bmp4 expression was examined (Fig. 7), which is normally expressed
predominantly on the left side of the cardiac cone (118/118, 100%;
Fig. 7I and M) (34). It was observed that wtip
depletion caused embryos to exhibit bilateral bmp4
expression (85/118, 72%; Fig. 7J and
M). The heart tube of wtip morphants never underwent
looping as occurred in control embryos at 48 hpf, which may reflect
altered left-right patterning of the body (Fig. 5A–D, G, K, O and R). Next, it was
checked whether Wtip regulates not only bmp4, but also other
early LR asymmetry markers in zebrafish. The left1/2
and southpaw (spaw) genes encode TGF-β superfamily proteins
that are expressed in the left lateral plate mesoderm in wild-type
embryos. These early laterality markers are essential for the
establishment of LR asymmetry in zebrafish (53). To investigate whether Wtip
regulates zebrafish LR patterning by modulating the formation
and/or function of KV, wtipMO was injected into the yolk at
the early blastula stage (128-cell stage) to target the precursors
of KV (DFCs) and left1/2 (Fig. 7A–D, K) and spaw (Fig. 7E–H, L) expression patterns were
examined. Injection of wtipMO led to randomization of the LR
axis, as shown by left1/2 (Fig. 7A–D, K) and spaw (Fig. 7E–H, L) gene expression in the
lateral plate mesoderm at 20 hpf. All control embryos displayed a
left-sided left1/2 (118/118, 100%) and spaw
(180/180, 100%) expression (Fig. 7A,
E, K and L). By contrast, wtip morphants exhibited the
full range of possible expression patterns: Left-sided expression
(left1/2, 111/159, 69.8%; spaw, 120/185,
64.9%), bilateral expression (left1/2, 6/159, 3.8%;
spaw, 37/185, 20%), right-sided expression
(left1/2, 4/159, 2.5%; spaw, 14/185, 7.5%),
and no expression (left1/2, 38/159, 23.9%;
spaw, 14/185, 7.5%; Fig. 7A–H,
K and L). These data suggest that Wtip signaling is required
for early LR asymmetry.
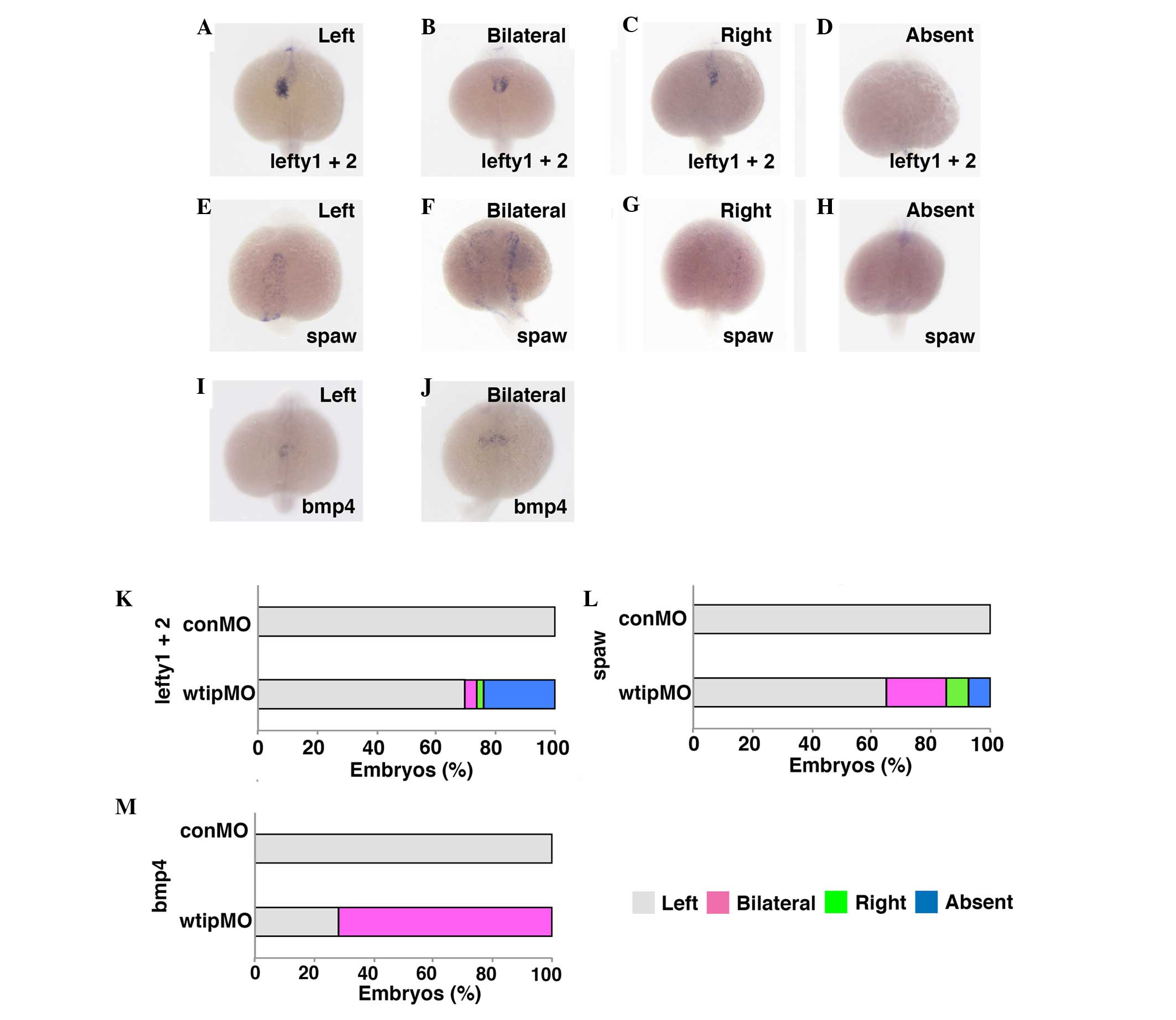 | Figure 7Wtip is required for early left-right
patterning. Dorsal views, anterior to the bottom of the panels.
(A–D,K) lefty1 and lefty2, (E–H,L) spaw and
(I,J,M) bmp4 expressions in (A–J) 22-somite stage embryos.
Normally, (A,E,I) left1+2, spaw and bmp4
transcripts expressed in the left cardiac lateral plate mesoderm,
(B,F,J) have bilateral, (C,G) right and (D,H) absent expression in
wtip morphants (1.035 pmol/embryo). Bar chart details the
left-right patterning for control and wtip morphants. Wtip,
Wilm's tumor 1 interacting protein; lefty1, left right
determination factor 1; spaw, southpaw; bmp4, bone
morphogenic factor 4; conMO, control morpholino
oligonucleotide. |
Discussion
In humans, a wide range of cardiovascular defects
is associated with kidney cystic disease, although the etiology of
these abnormalities remains unclear. Similar to humans, pericardial
edema in zebrafish is often observed in kidney cystic mutants or
MO-mediated knockdown embryos (74,75).
However, the specific roles of cilia and basal bodies in PE, heart
morphogenesis and function during normal development, and
congenital heart disease (CHD) remain unknown (37,76).
Compared to myocardium and endocardium, relatively little is known
about how PE develops and differentiates. The PE is an
extra-cardiac source for several cardiac cell types, and increasing
evidence suggests that the PE plays a role in heart development,
repair of cardiac injury and ingrowth of coronary vessels (5–12).
Despite the potentially central role of the epicardium in all of
these processes, the molecular signals driving the specification
and morphogenesis of PE remain poorly understood. In the present
study, the role of Wtip in PE development was investigated and
genetic evidence presented that WT1, in association with Wtip, is
required for PE specification. We further revealed a critical role
for Wtip in heart looping and the establishment of the AV boundary.
Based on the expression pattern of wtip mRNA and protein,
blocked Wtip function, and wtip over-expression data, it is
proposed that Wtip is required at early somite stages to provide LR
asymmetry, and independently regulates later cardiac development,
including heart-looping, PE specification and differentiation of
the AV boundary.
Wtip was originally identified as an interacting
partner of WT1 in a yeast two-hybrid screen (40). WT1 is expressed in the PE and
glomerulus podocytes in mammals, birds, zebrafish and medaka
(22,57–61).
It was previously reported that depletion of the Wtip basal body
was associated with the development of pronephric cysts, with it
noted that these embryos also developed pericardial edema (44). Previously, cryptic deletion in the
human wtip gene was reported to cause hypospadias (urethra
malformation in males) that can be associated with congenital heart
disease (45). Therefore, we
predicted that pericardial edema in wtip knockdown embryos
may be due to PE defects.
In the present study, a critical role of Wtip in PE
specification during heart development was identified. It was
demonstrated that wtip shows cardiac expression in PE at 48
hpf and 4 dpf, in a pattern that specifically matches that of known
PE-specific genes wt1a, tcf21, and tbx18
(22,23). Additionally, it was confirmed that
the PE contains ciliated cells, and that Wtip localizes to the
basal bodies within these cells in 48 hpf embryos, as was the case
for ciliated cells in the pronephros and Kupffer's vesicle
(44). This is the first evidence
that the zebrafish embryonic heart develops cilia and basal bodies
during embryonic development. Furthermore, it was observed that
depletion of wtip leads to the severe reduction or absence
of PE formation and that Wtip and wt1a cooperate for PE
formation. By overexpressing wtip mRNA, ectopic expression
of PE-specific markers was observed in the cardiac region and in
the pharyngeal arch area. Based on wtip gene and protein
expression patterns and on phenotypes generated by blocking Wtip
function, it is proposed that Wtip is required initially to
establish early LR asymmetry, and later to form the PE, modulate
heart looping and promote AV differentiation.
Recently, the PE has been investigated for its role
in normal heart morphogenesis (13,15,77)
and additional capacity to form coronary vascular endothelial cells
in mice (20). Zebrafish are now
well established as a key model system for embryonic heart
development and function, and possess a natural capacity for adult
myocardial regeneration (78). To
directly examine the possible critical roles for Wtip or the PE in
response to injury and myocardial regeneration in adults, we would
need to further consider using wt1a, tcf21 or
tbx18 transgenic lines (24–26).
A previous study reported that cells with primary
9+0 cilia were found in both the embryonic and the adult human
heart (79). Previous studies
revealed that monocilia are found in the mouse embryonic heart at
embryonic day (e) e9.5–e12.5, by which time blood flow is present
(39,76). The data suggest that in zebrafish
embryos, cilia occur in the PE by 48 hpf, a similar stage to what
has been observed in the chick embryonic heart (62). Previous reports suggest that mice
lacking cardiac cilia developed abnormal ECCs and compact
myocardium (CM) at e9.5 (39,76).
Notably, our observations in zebrafish suggest that wtip
knockdown embryos lack a differentiated AV boundary, which is
similar to the findings in mice, which lack ECCs (39,76).
In addition, Wtip and cilia markers are co-expressed in the
endocardium and myocardium prior to when the PE is formed in the
zebrafish heart (data not shown), which may provide a connection to
the AV valves that are simultaneously developing from the
endocardium and myocardium. Previous studies have reported the
timing and distribution of cilia in the embryonic mouse heart
(39,76). At e9.5, cilia on the endocardial
cells are predominantly located on the luminal surface, while cilia
in the mesenchymal cells of the developing cushions are randomly
oriented. By e12.5, atrial and ventricular septation is underway;
only few cilia are found on the atrial endocardial layer. In the
ECCs, however, cilia are found on the endothelial surface and are
visible in the mesenchymal cells. The present study is the first,
to the best of our knowledge, to report cilia on PE cells. Cilia in
the endothelium may sense the flow during embryonic heart
development. PE cells may be sensing information about their
environment that relates to pericardial fluid or heartbeats, or
potentially PE cilia may be involved in sensing biochemical
information, such as the movement of fluid.
In mice, cilia are required for cardiac
development, independently from their function in the development
of LR asymmetry (39,76). Thus, it is possible that cardiac
cilia function as mechanosensors, integrating information regarding
flow, cardiac function and morphogenesis. If so, the prediction is
that mutations in gene products that localize to cilia and/or basal
bodies may affect heart morphogenesis and function. In zebrafish,
many cystic mutant phenotypes accompany LR asymmetry defects
originating in early somitogenesis stages, visible as altered
spaw and left1/2 gene expression in the
lateral plate mesoderm and by altered bmp4 expression in the
zebrafish heart.
In control embryos, the progression of cardiac
looping is a morphogenic process. The cardiac looping angle is
defined as the degree of difference in the anterior/posterior (A/P)
body axis and the plane of the AV boundary (55). The present study was unable to
detect alterations in the expression of early patterning and
chamber identity genes, suggesting that additional causes must be
responsible for the malformation in the heart. Of note,
chamber-specific markers amhc and vmhc were not
altered, however, nppa was detected throughout the heart,
with no distinguishable exclusion from the AV boundary in
wtip knockdown embryos. Cardiac morphogenesis was
significantly morphologically and molecularly affected at 48 and 72
hpf in wtip knockdown embryos, and gene restriction at the
AV boundary was lost.
A number of the cardiac morphology mutants
described in the original zebrafish large-scale mutagenesis screens
have been well characterized, and the mutated genes have been
identified. However, whether any of these mutants exhibit defects
in cardiac cilia formation or function remains unknown, since the
presence of cilia in the heart was not formerly described. Cardiac
cilia are beginning to be characterized in other animal model
systems, including chickens (62)
and mice (39). As it was unknown
whether basal bodies could affect heart morphogenesis and function,
it was unclear whether heart defects could also be associated with
early embryonic defects, such as LR patterning, which are thought
to involve ciliary function.
The zebrafish system offers unique advantages for
studying cell biology during vertebrate organogenesis, as zebrafish
embryos develop externally and are practically transparent
throughout early development, thereby allowing non-invasive
observation. Previous studies in zebrafish indicate that
endocardial cells in the AV differentiate before the onset of
epithelial-to-mesenchymal transformation, thereby defining a
previously unappreciated step during AV valve formation (80,81).
New zebrafish mutants exhibiting AV valve development will provide
a unique set of tools with which to further understand the genetic
basis of PE cell behavior and its effect on heart development
(80,81). Importantly, AV-associated
phenotypes have been observed in mutant mouse models (81). The combination of genetics and
pharmacological studies in animal models and the cell biology
should provide novel insights into the developmental biology of
cardiac organogenesis and provide relevant information to predict
and understand human valvular and septal malformations.
The current study provides the first genetic
evidence, to the best of our knowledge, for a novel role for the
Wtip basal body protein in regulating the PE cell fate during
development. In addition, our tools for wtip knockdown will
be valuable for future studies to determine how Wtip signaling,
possibly mediated via cilia, could indirectly modulate
cardiac looping or AV development. The present study may
additionally provide an avenue for understanding how the PE
stimulates a response to cardiac injury, leading to its eventual
repair.
Acknowledgments
The authors would like to thank all members of the
Obara laboratory for the helpful discussion, and Kathy Kyler and
Hiroyuki Matsumoto for critical reading of the manuscript. The
authors would like to acknowledge the Zebrafish Information
Resource Center for providing fish. Professor Tomoko Obara
acknowledges financial support from the University of Oklahoma
Health Sciences Center (OUHSC). Professor Tomoko Obara was
supported by NIH grants R21-DK069604 and R01-DK078209, and the
Oklahoma Center for the Advancement of Science and Technology
(OCAST) grant HR14-082. The present study is supported in part by
the Diabetes Histology and Image Acquisition and Analysis Core
Facility at OUHSC (NIH: COBRE-1P20RR024215).
References
|
1
|
DeHaan RL: Morphogenesis of the vertebrate
heart. Organogenesis. DeHaan RL and Ursprung H: Holt, Rinehart and
Winston; New York: pp. 377–419. 1965
|
|
2
|
Serbedzija GN, Chen JN and Fishman MC:
Regulation in the heart field of zebrafish. Development.
125:1095–1101. 1998.PubMed/NCBI
|
|
3
|
Bagatto B, Francl J, Liu B and Liu Q:
Cadherin2 (N-cadherin) plays an essential role in zebrafish
cardiovascular development. BMC Dev Biol. 6:232006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fishman MC and Chien KR: Fashioning the
vertebrate heart: Earliest embryonic decisions. Development.
124:2099–2117. 1997.PubMed/NCBI
|
|
5
|
Mikawa T and Fishman DA: Retroviral
analysis of cardiac morphogenesis: Discontinuous formation of
coronary vessels. Proc Natl Acad Sci USA. 89:9504–9508. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mikawa T and Gourdie RG: Pericardial
mesoderm generates a populations of coronary smooth muscle cells
migrating into the heart along with ingrowth of the epicardial
organ. Dev Biol. 174:221–232. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dettman RW, Denetclaw W Jr, Ordahl CP and
Bristow J: Common epicardial origin of coronary vascular smooth
muscle, perivascular fibroblasts, and intermyocardial fibroblasts
in the avian heart. Dev Biol. 193:169–181. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pérez-Pomares JM, Macías D, García-Garrido
L and Munõz-Chápuli R: Immunolocalization of the vascular
endothelial growth factor receptor-2 in the subepicardial
mesenchyme of hamster embryos: Identification of the coronary
vessel precursors. HIstochem J. 30:627–634. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vrancken Peeters MP, Gittenberger-de Groot
AC, Mentink MM and Poelmann RE: Smooth muscle cells and fibroblasts
of the coronary arteries derive from epithelial-mesenchymal
transformation of the epicardium. Anat Embryol (Berl). 199:367–378.
1999. View Article : Google Scholar
|
|
10
|
Gittenberger-de Groot AC, Vrancken Petters
MP, Bergweff M, Mentink MM and Poelmann RE: Epicardial outgrowth
inhibition leads to compensatory mesothelial outflow tract collar
and abnormal cardiac septation and coronary formation. Cir Res.
87:969–971. 2000. View Article : Google Scholar
|
|
11
|
Reese DE, Mikawa T and Bader DM:
Development of the coronary vessel system. Circ Res. 91:761–768.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
von Gise A and Pu WT: Endocardial and
epicardial to mesenchymal transitions in heart development and
disease. Circ Res. 110:1628–1645. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Manner J: Experimental study on the
formation of the epicardium in chick embryos. Anat Embryol (Berl).
187:281–289. 1993. View Article : Google Scholar
|
|
14
|
Svensson EC: Deciphering the signals
specifying the proepiardium. Cir Res. 106:1789–1790. 2010.
View Article : Google Scholar
|
|
15
|
Bakkers J: Zebrafish as a model to study
cardiac development and human cardiac disease. Cardiovasc Res.
91:279–288. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kreidberg JA, Sariola H, Loring JM, Maeda
M, Pelletier J, Housman D and Jaenisch R: WT-1 is reqiored for
early kidney development. Cell. 74:679–691. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kwee L, Baldwin HS, Shen HM, Stewart CL,
Buck C, Buck CA and Laobow MA: Defective development of the
embryonic and extraembryonic circulatory systems in vascular cell
adhesion molecule (VCAM-1) deficient mice. Development.
121:489–503. 1995.PubMed/NCBI
|
|
18
|
Yang JT, Raybum H and Hynes RO: Cell
adhesion events mediated by alpha 4 integrins are essential in
placental and cardiac development. Development. 121:549–560.
1995.PubMed/NCBI
|
|
19
|
Moore AW, Mclnnes L, Kreidberg J, Hastie
ND and Schedl A: YAC complementation shows a requirement for Wt1 in
the development of epicardium, adrenal gland and throughout
nephrogenesis. Development. 126:1845–1857. 1999.PubMed/NCBI
|
|
20
|
Red-Horse K, Ueno H, Weissman IL and
Krasnow MA: Coronary arteries from by developmental reprogramming
of venous cells. Nature. 464:549–553. 2010. View Article : Google Scholar :
|
|
21
|
von Gise A, Zhou B, Honor LB, Ma Q, Petryk
A and Pu WT: WT1 regulates epicardial epithelial to mesenchymal
transition through β-catenin and retinoic acid signaling pathways.
Dev Biol. 356:421–431. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Serluca FC: Development of the
proepicardial organ in the zebrafish. Dev Biol. 315:18–27. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu J and Stainier DY: Tbx5 and Bmp
signaling are essential for proepicardium specification in
zebrafish. Circ Res. 106:1818–1828. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lepilina A, Coon AN, Kikuchi K, Holdway
JE, Roberts RW, Burns CG and Poss KD: A dynamic epicardial injury
response supports progenitor cell activity during zebrafish heart
regeneration. Cell. 127:607–619. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kikuchi K, Gupta V, Wang J, Holdway JE,
Wills AA, Fang Y and Poss KD: tcf21+ epicardial cells adopt
non-myocardial fates during zebrafish heart development and
regeneration. Development. 138:2895–2902. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kikuchi K, Holdway JE, Major RJ, Blum N,
Dahn RD, Begemann G and Poss KD: Retinoic acid production by
endocardium and epicardium is an injury response essential for
zebrafish heart regeneration. Dec Cell. 20:397–404. 2011.
View Article : Google Scholar
|
|
27
|
Kim J, Wu Q, Zhang Y, Wiens KM, Huan Y,
Rubin N, Shimada H, Handin RI, Chao MY, Tuan TL, et al: PDGF
signaling is required for epicardial function and blood vessel
formation in regenerating zebrafish hearts. Proc Natl Acad Sci USA.
107:17206–17210. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu H, Lee SH, Gao J, Liu X and
Iruela-Arispe ML: Inactivation of erythropoietin leads to defects
in cardiac morphogenesis. Development. 126:3597–3605.
1999.PubMed/NCBI
|
|
29
|
Chen TH, Chang TC, Kang JO, Choudhary B,
Makita T, Tran CM, Burch JB, Eid H and Sucov HM: Epicardial
induction of fetal cardiomyocyte proliferation via a retinoic
acid-inducible trophic factor. Dev Biol. 250:198–207. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sengbusch JK, He W, Pinco KA and Yang JT:
Dual functions of [alpha]4[beta]1 integrin in epicardial
development: Initial migration and long-term attachment. J Cell
Biol. 157:873–882. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hatcher CJ, Diman NY, Kim MS, Pennisi D,
Song Y, Goldstein MM, Mikawa T and Basson CT: A role for Tbx5 in
proepicardial cell migration during cardiogenesis. Physiol
Genomics. 18:129–140. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Schlueter J and Brand T: A right-sided
pathway involving FGF8/Snai1 controls asymmetric development of the
proepicardium in the chick embryo. Proc Natl Acad Sci USA.
106:7485–7490. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schulte I, Schlueter J, Abu-Issa R, Brand
T and Männer J: Morphological and molecular left-right asymmetries
in the development of the proepicardium: A comparative analysis on
mouse and chick embryos. Dev Dyn. 236:684–695. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen JN, van Eeden FJ, Warren KS, Chin A,
Nüsslein-Volhard C, Haffter P and Fishman MC: Left-right pattern of
cardiac BMP4 may drive asymmetry of the heart in zebrafish.
Development. 124:4373–4382. 1997.PubMed/NCBI
|
|
35
|
Shu X, Huang J, Dong Y, Choi J,
Langenbacher A and Chen JN: Na,K-ATPase alpha2 and Ncx4a regulate
zebrafish left-right patterning. Development. 134:1921–1930. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fakhro KA, Choi M, Ware SM, Belmont JW,
Towbin JA, Lifton RP, Khokha MK and Brueckner M: Rare copy number
variations in congenital heart disease patients identify unique
genes in left-right patterning. Proc Natl Acad Sc USA.
108:2915–2920. 2011. View Article : Google Scholar
|
|
37
|
Chin AJ, Saint-Jeannet JP and Lo CW: How
insights from cardiovascular developmental biology have impacted
the care of infants and children with congenital heart disease.
Mech Dev. 129:75–97. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Francis RJ, Christopher A, Devine WA,
Ostrowski L and Lo C: Congenital heart disease and the
specification of left-right asymmetry. Am J Physiol Heart Circ
Physiol. 302:H2102–H2111. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Slough J, Cooney L and Brueckner M:
Monocilia in the embryonic mouse heart suggest a direct role for
cilia in cardiac morphogenesis. Dev Dyn. 237:2304–2314. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Srichai MB, Konieczkowski M, Padiyar A,
Konieczkowski DJ, Mukherjee A, Hayden PS, Kamat S, El-Meanawy MA,
Khan S, Mundel P, et al: A WT1 co-regulator controls podocyte
phenotype by shuttling between adhesion structures and nucleus. J
Biol Chem. 279:14398–14408. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
van Wijk NV, Witte F, Feike AC, Schambony
A, Birchmeier W, Mundlos S and Stricker S: The LIM domain protein
Wtip interacts with the receptor tyrosine kinase Ror2 and inhibits
canonical Wnt signaling. Biochem Biophys Res Commun. 390:211–216.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Langer EM, Feng Y, Zhaoyuan H, Rauscher FJ
III, Kroll KL and Longmore GD: Ajuba LIM proteins are snail/slug
corepressors required for neural crest development in Xenopus. Dev
Cell. 14:424–436. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Das Thakur M, Feng Y, Jagannathan R, Seppa
MJ, Skeath JB and Longmore GD: Ajuba LIM proteins are negative
regulators of the Hippo signaling pathway. Curr Biol. 20:657–662.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bubenshchikova E, Ichimura K, Fukuyo Y,
Powell R, Hsu C, Morrical SO, Sedor JR, Sakai T and Obara T: Wtip
and Vangl2 are required for mitotic spindle orientation and cloaca
morphogenesis. Biol Open. 1:588–596. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gana S, Veggiotti P, Sciacca G, Fedeli C,
Bersano A, Micieli G, Maghnie M, Ciccone R, Rossi E, Plunkett K, et
al: 19q13.11 cryptic deletion: Description of two new cases and
indication for a role of WTIP haploinsufficiency in hypospadias.
Eur J Hum Genet. 20:852–856. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Westerfield M: The Zebrafish Book A Guide
For The Laboratory Use Of Zebrafish Danio (Brachydanio) Rerio. 4th
edition. University of Oregon Pressm; Eugene, OR: 2000
|
|
47
|
Feng J, Jia N, Han LN, Huang FS, Xie YF,
Liu J and Tang JS: Microinjection of morphine into thalamic nucleus
submedius depresses bee venom-induced inflammatory pain in the rat.
J Pharm Pharmacol. 60:1355–1363. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Amack JD and Yost HJ: The T box
transcription factor no tail in ciliated controls zebrafish
left-right asymmetry. Curr Biol. 14:685–690. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hauptmann G and Gerster T: Multicolor
whole-mount in situ hybridization. Methods Mol Biol. 137:139–148.
2000.PubMed/NCBI
|
|
50
|
Thisse C and Thisse B: High-resolution in
situ hybridization to whole-mount zebrafish embryos. Nat Protoc.
3:59–69. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yelon D, Home SA and Stainier DY:
Restricted expression of cardiac myosin genes reveals regulated
aspects of heart tube assembly in zebrafish. Dev Biol. 214:23–37.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Berdougo E, Coleman H, Lee DH, Stainier DY
and Yelon D: Mutation of weak atrium/atrial myosin heavy chain
disrupts atrial function and influences ventricular morphogenesis
in zebrafish. Development. 130:6121–6129. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Long S, Ahmad N and Rebagliati M: The
zebrafish nodal-related gene southpaw is required for visceral and
diencephalic left-right asymmetry. Development. 130:2303–2316.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ahmad I, Pacheco M and Santos MA:
Exzymatic and nonenzymatic antioxidants as an adaptaion to
phagocyte-induced damage in Anguilla Anguilla L. following in situ
harbor water exposure. Exotoxicol Environ Saf. 57:290–302. 2004.
View Article : Google Scholar
|
|
55
|
Chernyavskaya Y, Ebert AM, Milligan E and
Garrity DM: Voltage-gated calcium channel CACNB2 (β2.1) protein is
required in the heart for control of cell proliferation and heart
tube integrity. Dev Dyn. 241:648–662. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kim S, Zaghloul NA, Bubenshchikova E, Oh
EC, Rankin S, Katsanis N, Obara T and Tsiokas L: Nde1-mediated
inhibition of ciliogenesis affects cell cycle re-entry. Nat Cell
Biol. 13:351–360. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Pritchard-Jones K, Fleming S, Davison D,
Bickmore W, Porteous D, Gosden C, Bard J, Buckler A, Pelletier J,
Housman D, et al: The candidate Wilms' tumour gene is involved in
genitourinary development. Nature. 346:194–197. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Armstrong JF, Pritchard-Jones K, Bickmore
WA, Hastie ND and Bard JB: The expression of the Wilms' tumour
gene, WT1, in the developing mammalian embryo. Mech Dev. 40:85–97.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Drummond IA, Majumdar A, Hentschel H,
Elger M, Solnica-Krezel L, Schier AF, Neuhauss SC, Stemple DL,
Zwartkruis F, Rangini Z, et al: Early development of the zebrafish
pronephros and analysis of mutations affecting pronephric function.
Development. 125:4655–4667. 1998.PubMed/NCBI
|
|
60
|
Carmona R, González-Iriarte M,
Pérez-Pomares JM and Muñoz-Chápuli R: Localization of the Wilm's
tumour protein WT1 in avian embryos. Cell Tissue Res. 303:173–186.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ichimura K, Bubenshchikova E, Powell R,
Fukuyo Y, Nakamura T, Tran U, Oda S, Tanaka M, Wessely O, Kurihara
H, et al: A comparative analysis of glomerulus development in the
pronephros of medaka and zebrafish. PLoS One. 7:e452862012.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Van der Heiden K, Groenendijk BC, Hierck
BP, Hogers B, Koerten HK, Mommaas AM, Gittenberger-de Groot AC and
Poelmann RE: Monocilia on chicken embryonic endocardium in low
shear stress areas. Dev Dyn. 235:19–28. 2006. View Article : Google Scholar
|
|
63
|
Perner B, Englert C and Bollig F: The
Wilms tumor genes wt1a and wt1b control different steps during
formation of the zebrafish pronephros. Dev Biol. 309:87–96. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Auman HJ, Coleman H, Riley HE, Olale F,
Tsai HJ and Yelon D: Functional modulation of cardiac form through
regionally confined cell shape changes. PLoS Biol. 5:e532007.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Mably JD, Modhideen MA, Burns CG, Chen JN
and Fishman MC: Heart of glass regulates the concentric growth of
the heart in zebrafish. Curr Biol. 13:2138–2147. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Baker K, Warren KS, Yellen G and Fishman
MC: Defective 'pacemaker' current (Ih) in a zebrafish mutant with a
slow heart reate. Proc Natl Acad Sci USA. 94:4554–4559. 1997.
View Article : Google Scholar
|
|
67
|
Jacob E, Drexel M, Schwerte T and Pelster
B: Influence of hypoxia and of hypoxemia on the development of
cardiac activity in zebrafish larvae. Am J Physiol Regul Integr
Comp Physiol. 283:R911–R917. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Vermot J, Forouhar AS, Liebling M, Wu D,
Plummer D, Gharib M and Fraser SE: Reversing blood flows act
through klf2a to ensure normal valvulogenesis in the developing
heart. PLoS Biol. 7:e10002462009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Nagai Y, Asaoka Y, Namae M, Saito K,
Momose H, Mitani H, Furutani-Seiki M, Katada T and Nishina H: The
LIM protein Ajuba is required for ciliogenesis and left-right axis
determination in medaka. Biochem Biophys Res Commun. 396:887–893.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Witzel HR, Jungblut B, Choe CP, Crump JG,
Braun T and Dobreva G: The LIM protein Ajuba restricts the second
heart field progenitor pool by regulating Isl1 activity. Dev Cell.
23:58–57. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Essner JJ, Vogan KJ, Wagner MK, Tabin CJ,
Yost HJ and Brueckner M: Conserved function for embryonic nodal
cilia. Nature. 418:37–38. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Essner JJ, Amack JD, Nyholm MK, Harris EB
and Yost HJ: Kupffer's vesicle is a ciliated organ of asymmetry in
the zebrafish embryo that initiates left-right development of the
brain, heart and gut. Development. 132:1247–1260. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Kramer-Zucker AG, Olale F, Haycraft CJ,
Yoder BK, Schier AF and Drummond IA: Cilia-driven fluid flow in the
zebrafish pronephros, brain and Kupffer's vesicle is required for
normal organogenesis. Development. 132:1907–1921. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Wessely O and Obara T: Fish and frogs:
Models for vertebrate cilia signaling. Front Biosci. 13:1866–1880.
2008. View Article : Google Scholar
|
|
75
|
Swanhart LM, Cosentino CC, Diep CQ,
Davidson AJ, de Caestecker M and Hukriede NA: Zebrafish kidney
development: basic science to translational research. Birth Defects
Res C Embryo Today. 93:141–156. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Brueckner M: Impact of genetic diagnosis
on clinical management of patients with congenital heart disease:
Cilia point the way. Circulation. 125:2178–2180. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Svensson LG: Percutaneous aortic valves:
Effective in inoperable patients, what price in high-ris patients?
J Thorac Cardiovasc Surg. 140(6 Suppl): S10–S13; discussion
S86–S91. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Poss KD, Wilson LG and Keating MT: Heart
regeneration in zebrafish. Science. 298:2188–2190. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Myklebust R, Engedal H, Saetersdai TS and
Ulstein M: Primary 9 + 0 cilia in the embryonic and the adult human
heart. Anat Embryol (Berl). 151:127–139. 1977. View Article : Google Scholar
|
|
80
|
Beis D, Bartman T, Jin SW, Scott IC,
D'Amico LA, Ober EA, Verkade H, Frantsve J, Field HA, Wehman A, et
al: Genetic and cellular analyses of zebrafish atrioventricular
cushion and valve development. Development. 132:4193–4204. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Smith KA, Langendijk AK, Courtney AD, Chen
H, Paterson D, Hogan BM, Wicking C and Bakkers J: Transmembrane
protein 2 (Tmem2) is required to regionally restrict
atrioventricular canal boundary and endocardial cushion
development. Development. 138:4193–4198. 2011. View Article : Google Scholar : PubMed/NCBI
|