Introduction
Ginseng, derived from the root of Panax
ginseng C.A. Meyer (Araliaceae), has been used as a
traditional medicine in Asian countries for centuries. Today it is
used as a dietary supplement worldwide. Due to widespread use,
documenting pharmacological efficacies has become an important
goal. Ginsenosides are believed to be responsible for a broad range
of pharmacological properties ascribed to ginseng, including
antitumor, anti-inflammatory, antinociceptive, antiallergic and
antidiabetic activities (1–7).
Ginsenosides, which belong to a class of steroidal glycosides, are
composed of an aglycone (sapogenin), including protopanaxadiol,
protopanaxatriol (PPT) or oleanolic acid, and sugar moieties
attached to the C-3 and C-20 positions. Protopanaxadiol and PPT
exist in S- and R-stereoisomeric forms, based upon
the configuration of C-20. Of the two stereoisomeric forms,
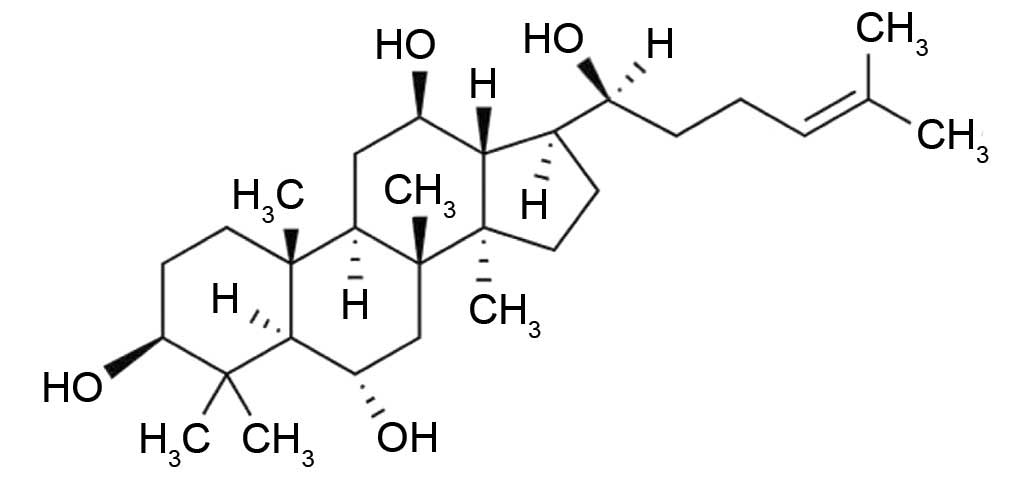
20(S)-PPT (Fig. 1) is the
major metabolic product of PPT-type ginsenosides, including Re, Rf,
Rg1, Rg2 and Rh1, which are deglycosylated by human intestinal
bacteria following oral consumption (8).
Purification of PPT and its stereoisomers has
enabled the pharmacological assessment of activities. PPT
suppresses the increase in lipopolysaccharide-induced inducible
nitric oxide synthase and cyclooxygenase-2 expression, which occur
via inactivation of nuclear factor-κB by preventing I-κBα
phosphorylation and degradation (9). This result is not consistent with
previous findings indicating that Panax ginseng extract rich
in PPT promotes nitric oxide production and relaxation of the
vascular endothelium via multiple signaling pathways (10). Peroxisome proliferator-activated
receptor (PPAR)γ, a member of the nuclear receptors of
ligand-activated transcription factor family, regulates the
expression of genes crucial for lipid and glucose metabolism, and
adipose cell differentiation. Accordingly, ligands of PPARγ are
potent insulin sensitizers used in the treatment of type 2 diabetes
(11). PPT enhances
PPARγ-transactivation activity and adipogenesis by increasing the
expression of PPARγ target genes, including adipocyte fatty
acid-binding protein, lipoprotein lipase, phosphoenolpyruvate
carboxykinase and glucose transporter 4, in 3T3-L1 adipocytes
(11). This finding suggested that
PPT may be a PPARγ agonist that can improve insulin resistance
associated with diabetes (11).
PPT inhibits corticosteroid production in adrenal
fasciculate cells in vivo and inhibits the conversion of
cholesterol to pregnenolone (12).
Pretreatment of human umbilical vein endothelial cells with PPT
produces a myriad of effects, including protecting cells against
hydrogen peroxide-induced cell injury and death, diminishing
hydrogen peroxide-induced DNA damage, and enhancing activation of
poly-ADP ribose polymerase and depleting its intracellular
substrate NAD+. PPT also prevents decreases in the ratio
of glutathione (GSH) to glutathione disulfide caused by hydrogen
peroxide, implying that PPT can protect human umbilical vein
endothelial cells against hydrogen peroxide-induced cell death by
protecting against oxidative stress (13). The 20(S)-PPT stereoisomer
also provides protection against myocardial ischemic injury by
enhancing the anti-free radical actions of heart tissues (14).
Findings indicated that 20(S)-PPT has
antioxidant activities in several cell types. However,
photoprotective properties in dermal keratinocytes remain to be
elucidated. Reactive oxygen species (ROS), produced by abnormal
metabolic reactions and exogenous stress-inducers, including
ultraviolet (UV) radiation, causes damage to macromolecules, which
can result in physiological dysfunction, genetic mutation, cell
aging and cell death (15). UV-B
radiation, with wavelengths between 280 and 315 nm, is one
well-known cause of skin photoaging, including wrinkle formation,
laxity, mottled pigmentation, coarseness, degradation of matrix
macromolecules, epidermal thickening, vascularization and
immunosuppression (16,17). Keratinocytes, the major cell
population in the basal layer of the skin, are the primary targets
of UV-B radiation. The present study was performed to determine
whether 20(S)-PPT possesses photoprotective properties that
combat the effects of UV-B irradiation in skin.
Materials and methods
Cell culture
Immortalized human HaCaT dermal keratinocytes
(American Type Culture Collection, Manassas, VA, USA) were grown in
Dulbecco's modified Eagle's medium (DMEM; HyClone Laboratories,
Logan, UT, USA) containing 10% heat-inactivated fetal bovine serum
(FBS; HyClone Laboratories), 100 U/ml penicillin and 100
µg/ml streptomycin (HyClone Laboratories) in a humidified
atmosphere with 5% CO2 at 37°C. Prior to treatments,
1×105 HaCaT cells were seeded into 24-well plates,
cultured overnight, washed twice with 1 ml phosphate-buffered
saline (PBS) and replaced with 1 ml FBS-free medium.
UV-B irradiation
An ultraviolet lamp (peak, 312 nm; VL-6 M; Vilber
Lourmat, Marne La Vallée, France) was used as a UV-B source. A
radiometer (VLX-3 W; Vilber Lourmat) with a sensor (bandwidth,
280–320 nm; CX-312; Vilber Lourmat) was used to monitor radiation
intensity. Mammalian cells were irradiated with UV-B radiation at
70 mJ/cm2.
Preparation of cellular lysates
Adherent cells were washed twice with PBS and stored
on ice for 5 min. The cells were harvested using a cell scraper and
centrifuged at 4°C for 10 min at 3,000 x g. The cell pellets were
resuspended in cell lysis buffer [50 mM HEPES (pH 7.5), 10% sucrose
and 0.1% Triton X-100] and were incubated for 30 min on ice.
Cellular lysates were obtained following centrifugation at 3,000 ×
g for 15 min and protein concentrations were determined using the
Bradford procedure (18). Bovine
serum albumin (BSA; Sigma-Aldrich, St Louis, MO, USA) was used as a
standard.
Determination of intracellular ROS
To determine intracellular ROS concentrations in
cultured keratinocytes, a redox-sensitive fluorescent probe,
2′,7′-dichlorofluorescein diacetate (DCFH-DA), which produces
2′,7′-dichlorofluorescein (DCF; λexcitation=485 nm,
λemission=530 nm) was used as previously described
(19). Following treatment with
20(S)-PPT (0, 5, 12 and 30 µM; Ambo Institute, Seoul,
Korea) and 20 µM DCFH-DA (Sigma-Aldrich) for 30 min at 37°C,
the cells were washed twice with 1 ml FBS-free medium. The cells
were resuspended in 1 ml FBS-free medium and irradiated with 70
mJ/cm2 UV-B. Intracellular ROS levels were immediately
quantified using a Multi-Mode Microplate Reader (Synergy Mx; BioTek
Instruments, Winooski, VT, USA). For imaging analyses, the cells
were treated with 20(S)-PPT and 20 µM
dihydrorhodamine-123 (DHR-123; Sigma-Aldrich) for 30 min at 37°C,
irradiated with 70 mJ/cm2 UV-B, and immediately analyzed
using confocal laser scanning microscopy (Fluoview-FV300; Olympus,
Tokyo, Japan). The assays were repeated at least three times.
Quantitation of nitrite
Accumulated nitrite (NO2−) in
culture supernatants was quantitated using a spectrophotometric
assay, based upon the Griess reaction (20). Briefly, an equal volume of Griess
reagent (1% sulfanilamide-0.1% N-1-naphthyl-ethylenediamine
dihydrochloride in 2.5% phosphoric acid; Sigma-Aldrich) was
incubated with culture supernatants for 10 min at room temperature
and the absorbance at 550 nm was measured using an enzyme-linked
immunosorbent assay reader (Molecular Devices, Sunnyvale, CA, USA).
A calibration curve was constructed using known concentrations
(0–160 µM) of sodium nitrite (Sigma-Aldrich).
Cell viability assay
To determine the survival of keratinocytes in the
presence of 20(S)-PPT, an MTT assay, which detects metabolic
activity (21), was used. The
cells were treated with 20(S)-PPT (0, 5, 12 and 30
µM) for 30 min. After removing the medium, the cells were
treated with 5 µg/ml
3-(4,5-dimeth-ylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT;
Sigma-Aldrich) in the medium. After incubating the cells at 37°C
for 4 h, the MTT formazan crystals were dissolved in dimethyl
sulfoxide (150 µl; Sigma-Aldrich). Formazan, generated from
reduction of MTT in the mitochondria of living cells, was
quantified using the absorbance at 540 nm.
Gelatin zymography
Gelatinase activities of matrix metalloproteinase
(MMP)-2 and -9 were determined using zymographic analyses, as
previously described (22). The
cells were incubated for 24 h at 37°C and were washed twice with 1
ml PBS. Cells, in 1 ml FBS-free medium, were treated with
20(S)-PPT for 30 min and irradiated with 70
mJ/cm2 UV-B. Culture supernatants, obtained from the
irradiated culture incubated for 24 h at 37°C, were fractionated on
10% (w/v) sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) gels impregnated with 1 mg/ml gelatin (Sigma-Aldrich)
under non-reducing conditions. The proteins in the gel were
renatured by agitating twice in 2.5% Triton X-100 at room
temperature for 30 min, and incubating in buffer [50 mM Tris buffer
(pH 7.8), 5 mM CaCl2, 0.15 M NaCl and 1% Triton X-100]
for 24 h. Following this, the gel was stained with 0.1% Coomassie
Brilliant Blue R-250 and gelatin-degrading enzyme activities were
shown as clear zones against the blue background. MMP-2 and MMP-9
activity bands were identified in accordance with molecular mass,
which was estimated using molecular mass markers (Quantity One
Software, version 4.6.9, Bio-Rad Laboratories, Richmond, CA,
USA).
Quantitation of total GSH
As previously described (23), the total GSH content was determined
using an enzymatic recycling assay based on glutathione reductase
(Sigma-Aldrich). The reaction mixture (200 µl), containing
175 mM KH2PO4, 6.3 mM EDTA, 0.21 mM NADPH
(Sigma-Aldrich), 0.6 mM DTNB (Sigma-Aldrich) and 0.5 units/ml
glutathione reductase, and the cellular lysate was incubated at
25°C for 5 min. The changes in absorbance at 412 nm were monitored
using a microplate reader. The total GSH was reported as
µg/mg protein.
Western blotting analysis
To detect the protein expression levels of MMP-2 and
MMP-9 in conditioned media and cellular lysates, western blotting
was performed. Conditioned media and cellular lysates were
separated on 10% SDS-PAGE gels and electrotransferred onto
polyvinylidene membranes. The membranes were blocked at room
temperature for 2 h with blocking buffer (2% BSA in 1X
Tris-buffered saline with 0.1% Tween-20). Following blocking, the
membranes were probed with primary antibody overnight at 4°C. The
following antibodies, after 1,000-fold dilution, were used:
Anti-MMP-2 (cat. no. ALX-210-753; Enzo Life Sciences, Farmingdale,
NY, USA), anti-MMP-9 (cat. no. #3852S; Cell Signaling Technology,
Inc., Danvers, MA, USA) and anti-glyceraldehyde 3-phosphate
dehydrogenase (GAPDH; cat. no. LF-PA0212; Young In Frontier Co.,
Ltd., Seoul, Korea). After the membranes were washed with PBS, they
were subsequently incubated with goat anti-rabbit immunoglobulin
G-pAb-horseradish peroxidase-conjugated secondary antibody (cat.
no. ADI-SAB-300; Enzo Life Sciences) for 1 h at room temperature,
and were developed using enhanced West-save up ™ (AbFrontier,
Seoul, Korea). The band strength, expressed as a percentage of the
control, was determined by densitometry using ImageJ software
(version 1.48; National Institutes of Health, Bethesda, MA,
USA).
Statistical analyses
The data are presented as the mean ± standard
deviation. Comparisons between experimental groups were
statistically analyzed using an unpaired Student's t-test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
UV-B-induced ROS elevation
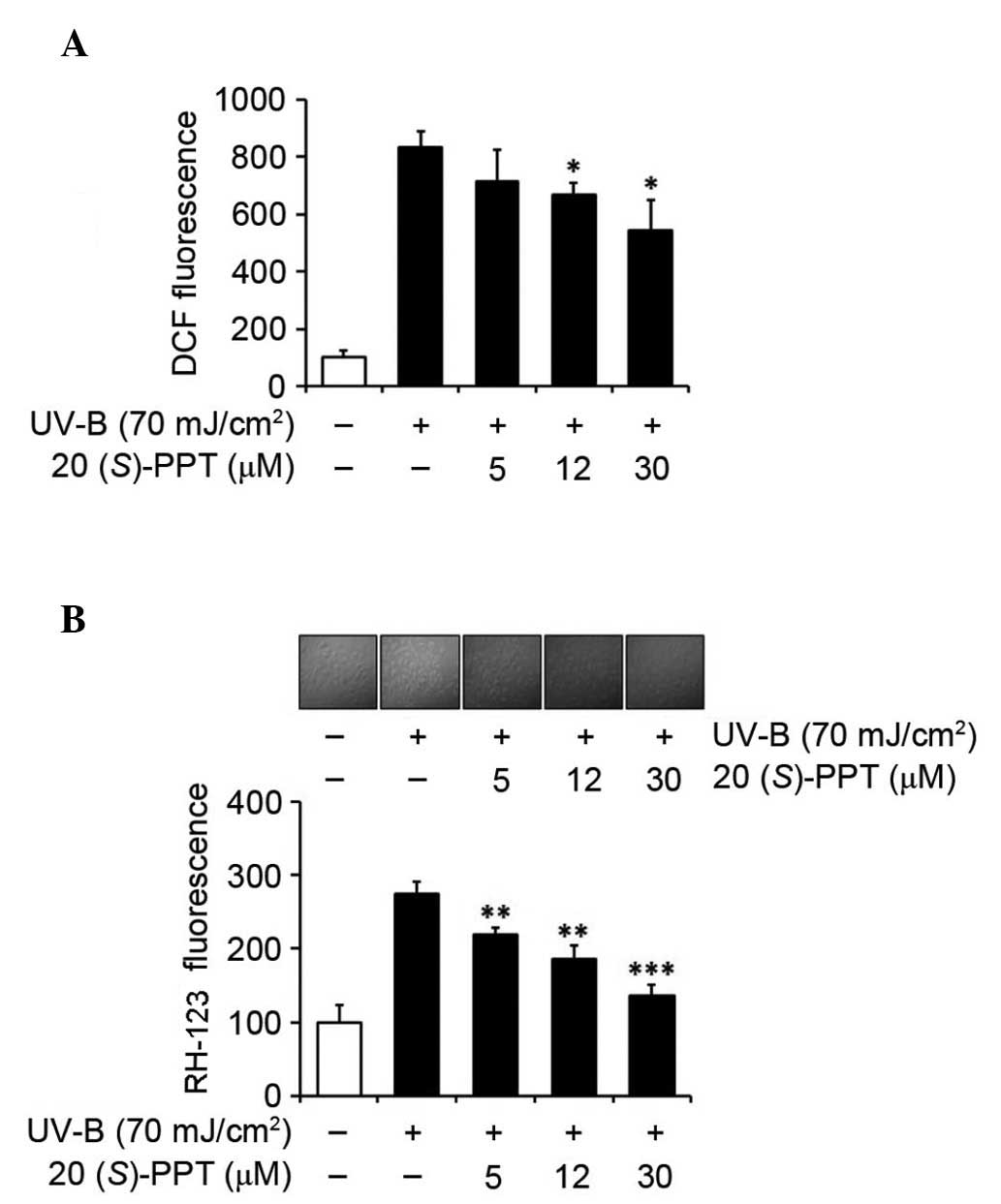
To determine the effects of UV-B radiation on the
production of ROS, cultured HaCaT cells were exposed to 70
mJ/cm2 UV-B radiation. Without pretreatment with
20(S)-PPT, intracellular ROS levels increased 8.3-fold
compared with the non-irradiated cells (Fig. 2A). Pretreatment of cells with
20(S)-PPT attenuated UV-B-induced ROS elevation in a
concentration-dependent manner (Fig.
2A). Treatment with 5, 12 and 30 µM 20(S)-PPT
reduced ROS elevation to 85.6, 80.2 and 65.4%, respectively, of
irradiated cells without 20(S)-PPT pretreatment (Fig. 2A). The activity of 20(S)-PPT
on the UV-B-induced ROS elevation in keratinocytes was also
determined by confocal microscopy analyses using
dihydrorhodamine-123 (Fig. 2B).
Treating cultured HaCaT cells with 5, 12 and 30 µM
20(S)-PPT prior to UV-B irradiation reduced ROS levels to
79.7, 67.6 and 50.0%, respectively, of the cultured HaCaT cells
without 20(S)-PPT pretreatment (Fig. 2B). These data indicated that
20(S)-PPT diminishes ROS elevation in keratinocytes
following UV-B irradiation.
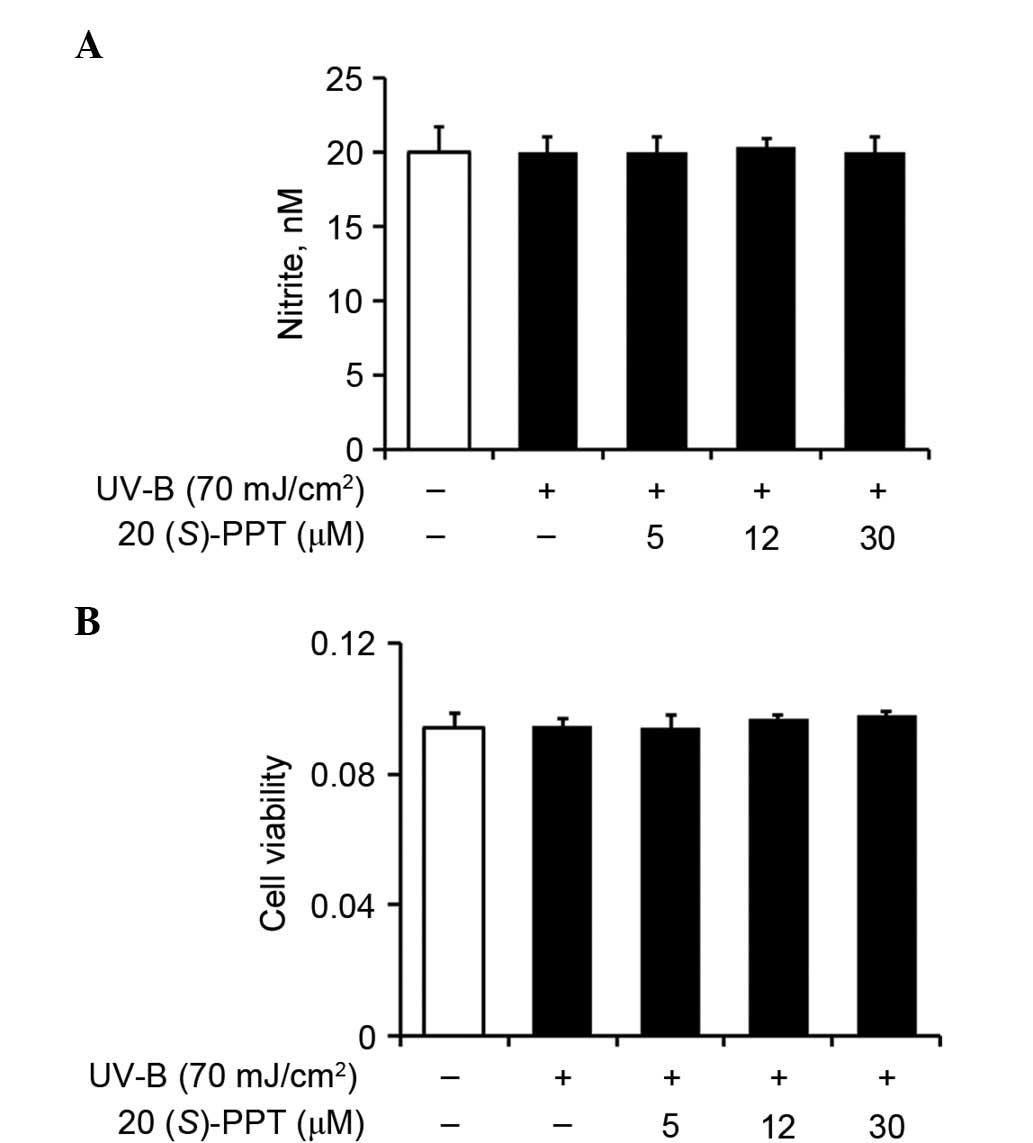
Nitric oxide production and cell
survival
Nitric oxide, synthesized in minute quantities by
constitutive nitric oxide synthases, is involved in normal
physiological processes. By contrast, it exhibits pathological
effects when synthesized in excessive quantities by inducible
nitric oxide synthases, which respond to proinflammatory agents.
Nitric oxide levels remained unchanged in UV-B-irradiated HaCaT
cells compared with the non-irradiated cells (Fig. 3A). These results were obtained
regardless of pretreatment with 20(S)-PPT (Fig. 3A), indicating that 20(S)-PPT
fails to modulate nitric oxide levels in UV-B-irradiated HaCaT
cells.
To determine whether UV-B radiation produced
cytotoxic affects in HaCaT cells, MTT assays were used to assess
cell viability. As shown in Fig.
3B, UV-B irradiation alone did not affect viability. Similarly,
pretreatment with 20(S)-PPT caused no alteration of the
effects of UV-B irradiation (Fig.
3B), indicating that 20(S)-PPT treatment had no affect
on the viability of irradiated keratinocytes.
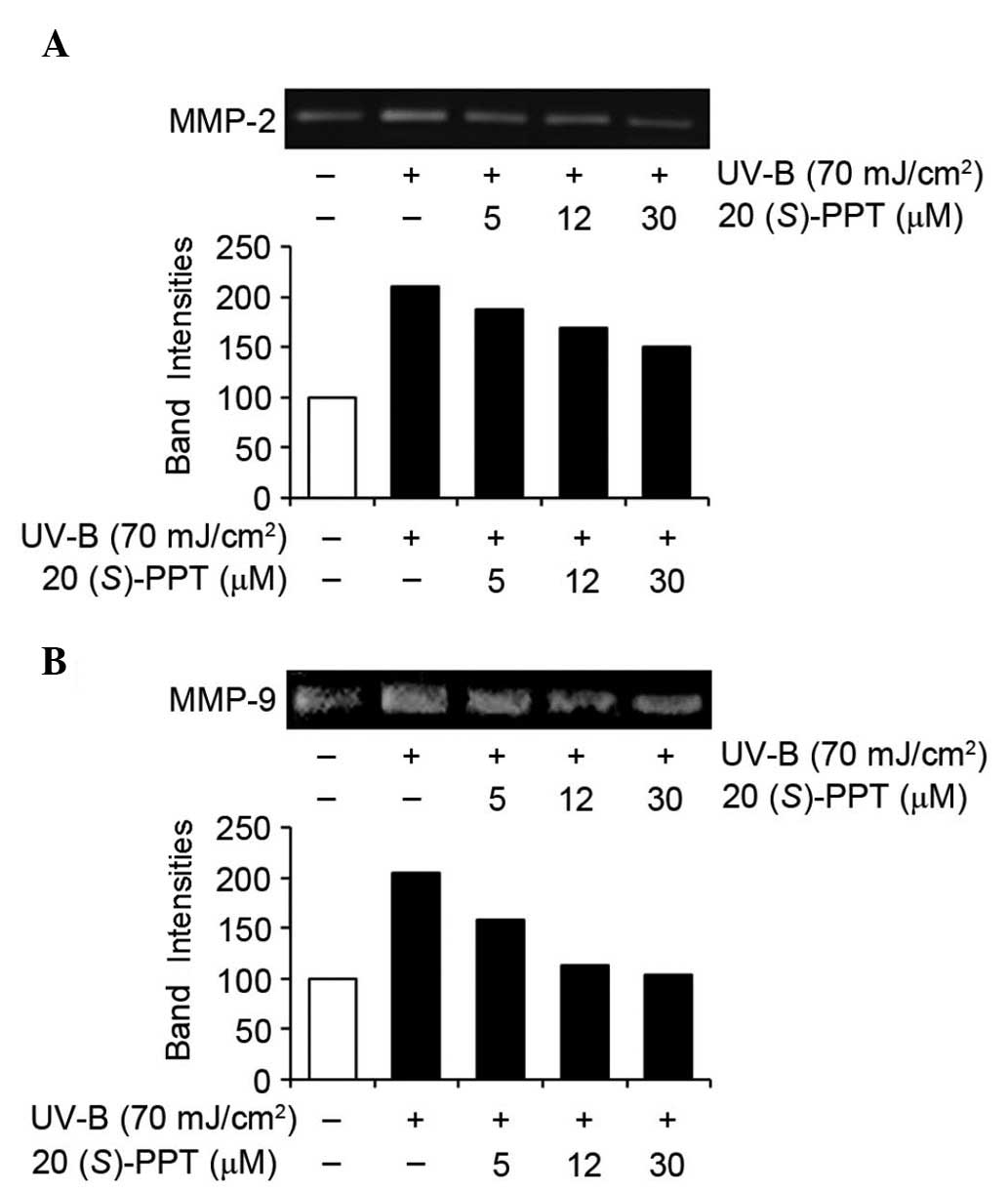
Activity and expression of MMP-2 and
MMP-9
UV irradiation is capable of inducing the expression
of certain MMP family members, including MMP-9, MMP-2 and MMP-1,
which degrade collagen and other extracellular matrix proteins of
the dermal connective tissue. Therefore, chronic exposure to UV
radiation alters the architecture of the skin, leading to
photoaging (24). ROS scavenging,
and suppression of MMP activity and expression attenuated premature
photoaging of skin. As shown in Fig.
4A, 20(S)-PPT attenuated the gelatinase activity of
MMP-2, which was significantly enhanced by UV-B irradiation in
HaCaT cells. In a similar manner, the gelatinase activity of MMP-9
was markedly enhanced by UV-B irradiation in HaCaT cells and
enhanced MMP-9 activity was diminished by 20(S)-PPT in a
concentration-dependent manner (Fig.
4B). These results indicated that 20(S)-PPT attenuates
the gelatinase activities of MMP-2 and MMP-9 in keratinocytes,
which were enhanced by UV-B irradiation.
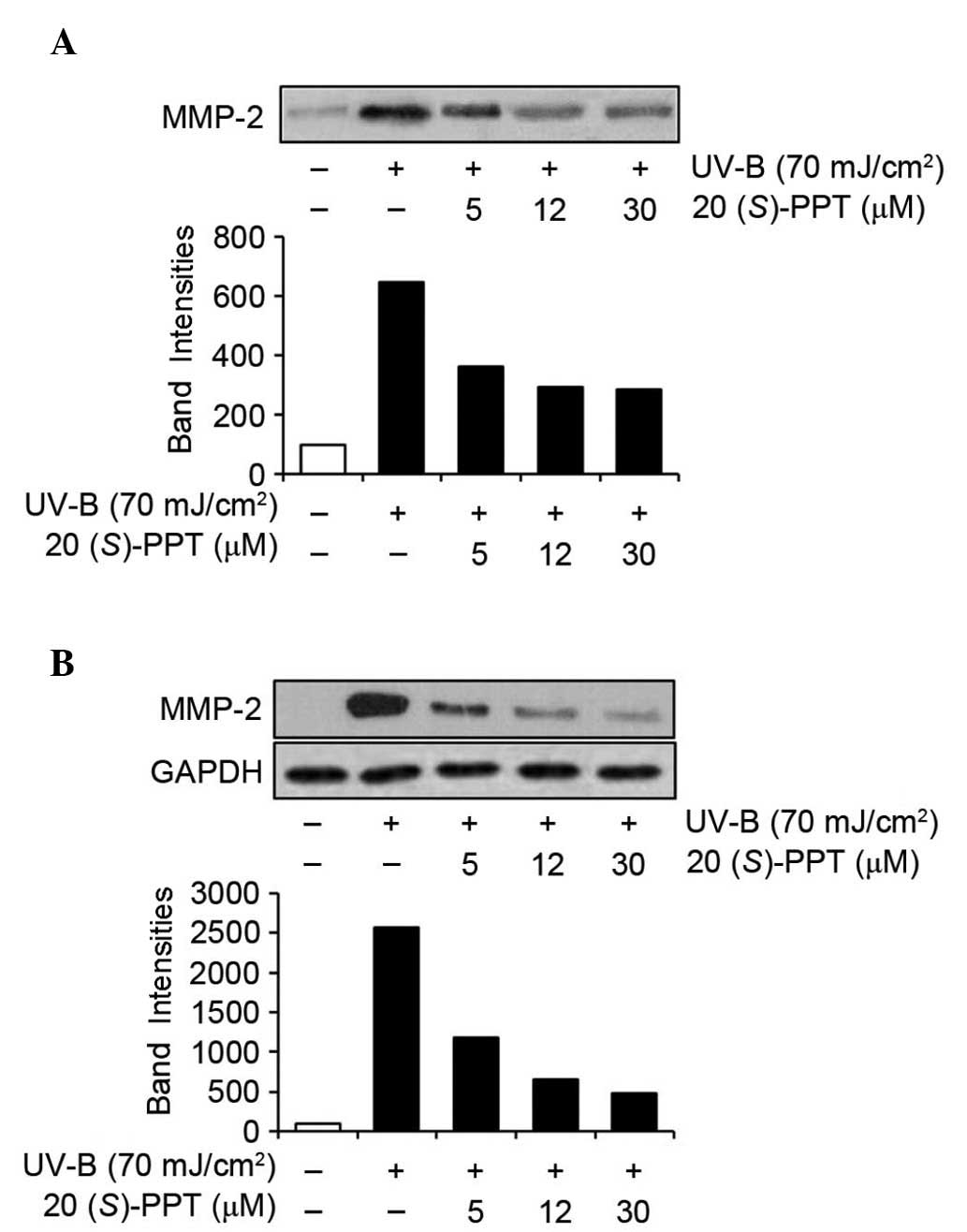
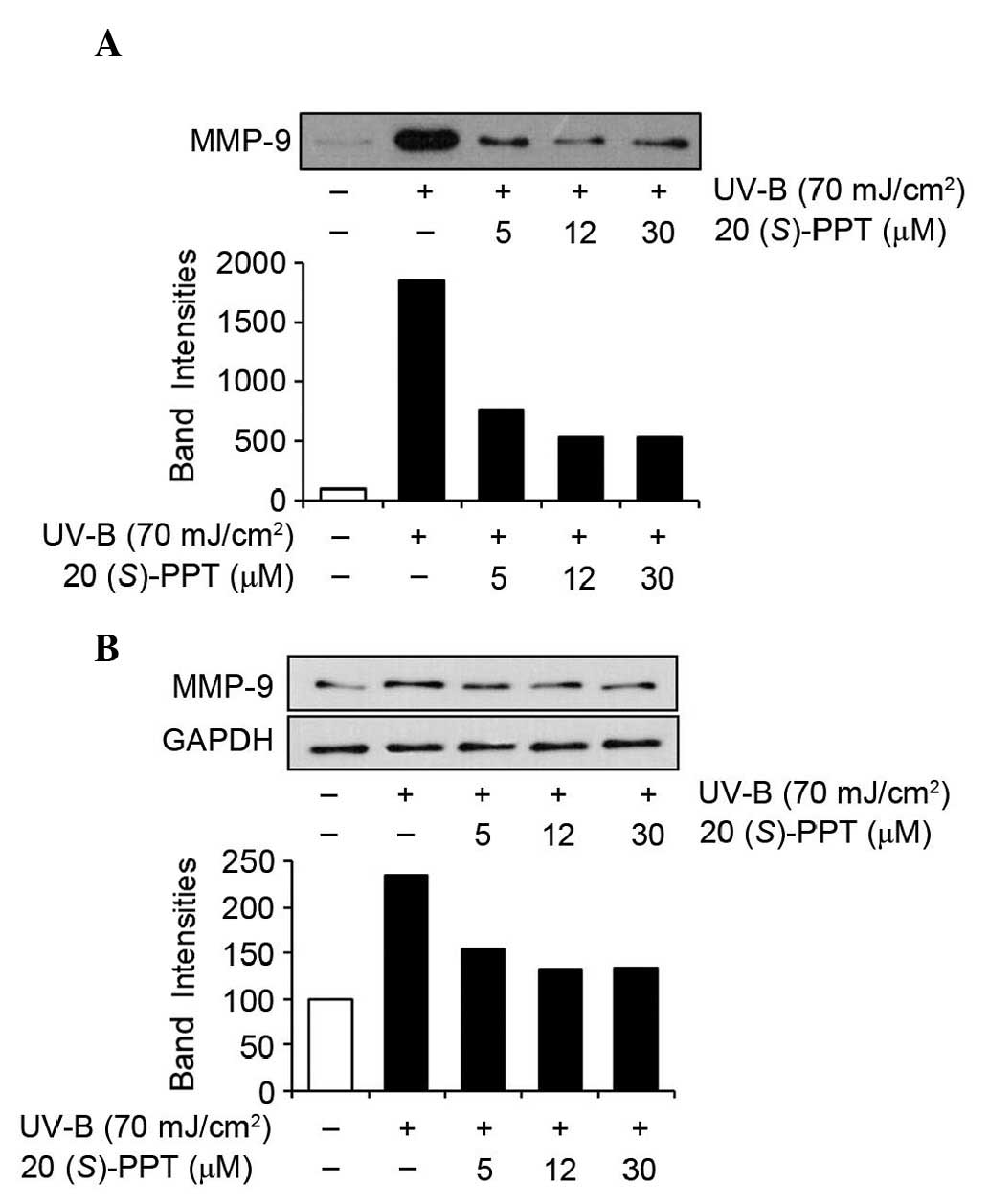
The effects of 20(S)-PPT on MMP-2 and MMP-9
were also determined by western blotting analyses of conditioned
media and cell lysates (Figs. 5
and 6). UV-B radiation enhanced
the protein expression of MMP-2 in conditioned media from HaCaT
cells (Fig. 5A), and
20(S)-PPT reduced UV-B-induced MMP-2 protein levels in
conditioned media in a concentration-dependent manner (Fig. 5A). Analyses of cell lysates
produced similar results. As shown in Fig. 5B, MMP-2 protein levels in cellular
lysates were enhanced by UV-B irradiation and the UV-B-enhanced
MMP-2 protein levels were reduced by 20(S)-PPT in a
concentration-dependent manner. MMP-9 protein levels in conditioned
media were significantly elevated by UV-B irradiation and
UV-B-induced MMP-9 protein levels were diminished by
20(S)-PPT (Fig. 6A).
Additionally, as shown in Fig. 6B,
MMP-9 protein levels in HaCaT cell lysates were induced by UV-B
irradiation and increased MMP-9 protein levels were attenuated by
20(S)-PPT. Collectively, these results indicated that
20(S)-PPT downregulates the UV-B-mediated induction of MMP-2
and MMP-9 in HaCaT cells.
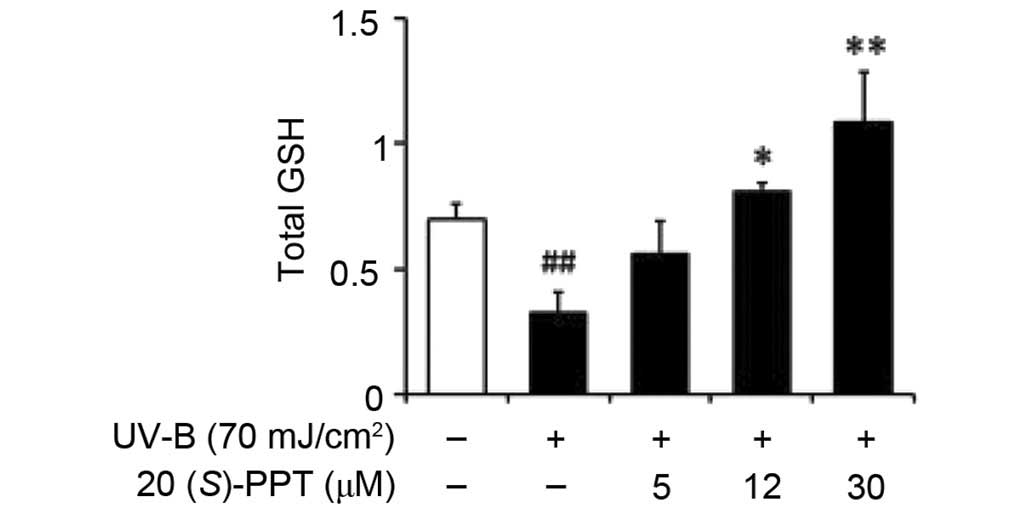
GSH content
The total GSH levels were significantly diminished
in HaCaT cells irradiated with 70 mJ/cm2 UV-B (Fig. 7). Pretreatment with 5, 12 and 30
µM 20(S)-PPT increased GSH levels by 1.7-, 2.5- and
3.3-fold, respectively, compared with irradiated keratinocytes that
did not receive pretreatment (Fig.
7). These data indicated that the GSH-enhancing effects of
20(S)-PPT assisted with combating the effects of UV-B
irradiation.
Discussion
The present study revealed that 20(S)-PPT, a
major metabolic intermediate of PPT-type ginsenosides, diminished
ROS levels in keratinocytes, suppressed the gelatinase activities
of MMP-2 and MMP-9, and decreased the protein expression levels of
MMP-2 and MMP-9 in cellular lysates and media. These data suggested
that 20(S)-PPT and associated PPT-type ginsenosides,
including Re, Rf, Rg1, Rg2 and Rh1, may have antiphotoaging
properties.
Oxidative stress, caused by imbalanced pro-oxidant
processes and antioxidant defense systems, occurs when cellular
redox homeostasis systems are disturbed. Chronic exposure to UV
light from the sun or artificial sources causes oxidative stress,
which results in the photoaging of skin. UV-A and UV-B radiation
from sunlight is hypothesized to be responsible for ~90% of skin
photoaging (25,26). UV-B radiation increases the
production of ROS, which cause oxidative damage in epidermal and
dermal cells, and severe oxidative stress may cause apoptotic or
necrotic death in dermal cells (16). UV radiation also induces the
expression and secretion of MMP-1, MMP-2, MMP-3, MMP-9 and MMP-13
in the epidermis and dermis, which contributes to skin damage and
photoaging (27,28).
Collagen destruction is closely associated with the
induction of MMPs, which are secreted by epidermal keratinocytes
and dermal fibroblasts (29).
Since skin photoaging is also a leading cause of skin cancer,
inhibiting the elevation of ROS and MMP activity may reduce skin
photoaging and cancer risk. Several crude plant extracts that
protect skin by downregulating ROS and/or MMPs have been previously
identified (30–33). A glutathione peroxidase mimic,
2-selenium-bridged β-cyclodextrin, reverses apoptosis and lipid
peroxidation in keratinocytes (34).
GSH is a well-known small-molecule antioxidant with
central roles in maintaining cellular redox homeostasis and
protecting against oxidative stress and injury. UV-B
irradiation-induced GSH depletion in cultured human keratinocytes
mimics the pathogenesis of several cutaneous disorders. Decreased
activity of γ-glutamylcysteine synthetase, a rate-limiting enzyme
in GSH biosynthesis, and diminished cysteine uptake through the
functional inhibition of the system Xc(−), a cysteine transporter
on the cell membrane (35), are
implicated in the pathogenesis. Indeed, inactivation of the
cysteine transporter system is a major contributor to the
UV-B-induced decrease of GSH levels in human keratinocytes
(35). GSH depletion has also been
shown in a commercially available reconstituted human epidermis
model of UV-B radiation (36).
When mouse lenses are exposed to UV-B radiation, simultaneous
decreases in ATP and GSH, which can be prevented by caffeine, were
observed (37). In the aquatic
organism Tubifex, UV-B radiation induced the production of singlet
oxygen, superoxide anions and hydroxyl radicals, and diminished the
levels of GSH, DNA, RNA and protein (38).
Several pure ginsenosides can restore GSH levels
after depletion caused by stress. Ginsenoside Rg1 decreased
malondialdehyde levels and intracellular ROS, and enhanced
superoxide dismutase activity and the total GSH in colistin-treated
rat pheochromocytoma cells, implying that neuroprotective effects
may be mediated via the inhibition of oxidative stress (39). Similarly, Rg1 enhanced the total
GSH in Schwann cells exposed to hydrogen peroxide (40), and Rb2, a protopanaxadiol-type
ginsenoside, decreased blood malondialdehyde and elevated total GSH
in ovariectomized mice. These data suggested that the
anti-osteoporosis effects of Rb2 may be linked to reduced oxidative
damage (41). Another ginsenoside,
Re, also restores GSH levels in the serum of diabetic rats and
attenuates diabetes-associated cognitive decline (42). Ginsenoside Rd was shown to increase
the total GSH and promote the antioxidant activities of catalase,
superoxide dismutase and glutathione peroxidase in cultured
hippocampal neurons exposed to oxygen-glucose deprivation (43); Rd also enhanced total GSH and
levels of γ-glutamylcysteine synthetase heavy chain in a rat
hepatocyte cell line (44).
Collectively, although GSH-restoring effects were not detected in
the previous experiments using UV-B irradiation, certain
ginsenosides were shown to enhance GSH levels that had declined
under other types of conditions (42–44).
The present results confirmed that 20(S)-PPT
suppresses the GSH depletion caused by UV-B exposure. Although the
mechanisms of these protective activities remain unknown,
restoration of GSH levels may be an initiating event that leads to
lowering of ROS in keratinocytes.
In conclusion, 20(S)-protopanaxatriol, a
major metabolic product generated by intestinal bacteria after oral
ingestion of PPT-type ginsenosides, has skin photoprotective
properties that may occur through restoration of GSH levels,
reduction of ROS production, and downregulation of MMP-2 and MMP-9.
Taken together, these results suggested that 20(S)-PPT and
other PPT-type ginsenosides must be investigated as natural
antiphotoaging treatments. As natural products, they may have
similar or enhanced activities with fewer side effects.
Acknowledgments
The authors would like to thank Ms. Hee Han for
technical assistance. This study was supported by a grant of the
Korean Health Technology R&D Project, Ministry of Health &
Welfare, Republic of Korea (grant no. HN12C0060).
References
|
1
|
Bu QT, Zhang WY, Chen QC, Zhang CZ, Gong
XJ, Liu WC, Li W and Zheng YN: Anti-diabetic effect of ginsenoside
Rb (3) in alloxan-induced diabetic mice. Med Chem. 8:934–941. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen J, Peng H, Ou-Yang X and He X:
Research on the antitumor effect of ginsenoside Rg3 in B16 melanoma
cells. Melanoma Res. 18:322–329. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu C, Zhang M, Hu MY, Guo HF, Li J, Yu
YL, Jin S, Wang XT, Liu L and Liu XD: Increased glucagon-like
peptide-1 secretion may be involved in antidiabetic effects of
ginsenosides. J Endocrinol. 217:185–196. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Park EK, Choo MK, Han MJ and Kim DH:
Ginsenoside Rh1 possesses antiallergic and anti-inflammatory
activities. Int Arch Allergy Immunol. 133:113–120. 2004. View Article : Google Scholar
|
|
5
|
Seo YJ, Kwon MS, Choi HW, Jang JE, Lee JK,
Sun Y, Jung JS, Park SH and Suh HW: Intracerebroventricular
ginsenosides are antinociceptive in proinflammatory
cytokine-induced pain behaviors of mice. Arch Pharm Res.
31:364–369. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tao T, Chen F, Bo L, Xie Q, Yi W, Zou Y,
Hu B, Li J and Deng X: Ginsenoside Rg1 protects mouse liver against
ischemia-reperfusion injury through anti-inflammatory and
anti-apoptosis properties. J Surg Res. 191:231–238. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang W, Zhang X, Qin JJ, Voruganti S, Nag
SA, Wang MH, Wang H and Zhang R: Natural product ginsenoside
25-OCH3-PPD inhibits breast cancer growth and metastasis through
down-regulating MDM2. PLoS One. 7:e415862012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hasegawa H, Suzuki R, Nagaoka T, Tezuka Y,
Kadota S and Saiki I: Prevention of growth and metastasis of murine
melanoma through enhanced natural-killer cytotoxicity by fatty
acid-conjugate of protopanaxatriol. Biol Pharm Bull. 25:861–866.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Oh GS, Pae HO, Choi BM, Seo EA, Kim DH,
Shin MK, Kim JD, Kim JB and Chung HT: 20 (S)-Protopanaxatriol, one
of ginsenoside metabolites, inhibits inducible nitric oxide
synthase and cyclooxygenase-2 expressions through inactivation of
nuclear factor-kappaB in RAW 264.7 macrophages stimulated with
lipopolysaccharide. Cancer Lett. 205:23–29. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ahn HY, Hong SY, Kim JY and Kwon O: Panax
ginseng extract rich in ginsenoside protopanaxatriol offers
combinatorial effects in nitric oxide production via multiple
signaling pathways. Springerplus. 2:962013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Han KL, Jung MH, Sohn JH and Hwang JK:
Ginsenoside 20S-protopanaxatriol (PPT) activates peroxisome
proliferator-activated receptor gamma (PPARgamma) in 3T3-L1
adipocytes. Biol Pharm Bull. 29:110–113. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hasegawa E, Nakagawa S, Miyate Y,
Takahashi K, Ohta S, Tachikawa E and Yamato S: Inhibitory effect of
protopanaxatriol ginseng metabolite M4 on the production of
corticosteroids in ACTH-stimulated bovine adrenal fasciculata
cells. Life Sci. 92:687–693. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kwok HH, Ng WY, Yang MS, Mak NK, Wong RN
and Yue PY: The ginsenoside protopanaxatriol protects endothelial
cells from hydrogen peroxide-induced cell injury and cell death by
modulating intracellular redox status. Free Radic Biol Med.
48:437–445. 2010. View Article : Google Scholar
|
|
14
|
Han B, Meng Q, Li Q, Zhang J, Bi Y and
Jiang N: Effect of 20(S)-protopanaxatriol and its epimeric
derivatives on myocardial injury induced by isoproterenol.
Arzneimittelforschung. 61:148–152. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nordberg J and Arnér ES: Reactive oxygen
species, antioxidants and the mammalian thioredoxin system. Free
Radic Biol Med. 31:1287–1312. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rabe JH, Mamelak AJ, McElgunn PJ, Morison
WL and Sauder DN: Photoaging: Mechanisms and repair. J Am Acad
Dermatol. 55:1–19. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Slominski A, Wortsman J and Tobin DJ: The
cutaneous serotoninergic/melatoninergic system: Securing a place
under the sun. FASEB J. 19:176–194. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Royall JA and Ischiropoulos H: Evaluation
of 2′, 7′-dichloro-fluorescin and dihydrorhodamine 123 as
fluorescent probes for intracellular H2O2 in
cultured endothelial cells. Arch Biochem Biophys. 302:348–355.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sherman MP, Aeberhard EE, Wong VZ,
Griscavage JM and Ignarro LJ: Pyrrolidine dithiocarbamate inhibits
induction of nitric oxide synthase activity in rat alveolar
macrophages. Biochem Biophys Res Commun. 191:1301–1308. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Freshney RI: Culture of animal cells: A
manual of basic technique. 4th Edition. Wiley-Liss Press; New York:
1994
|
|
22
|
Kleiner DE and Stetler-Stevenson WG:
Quantitative zymography: Detection of picogram quantities of
gelatinases. Anal Biochem. 218:325–329. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nakagawa K, Saijo N, Tsuchida S, Sakai M,
Tsunokawa Y, Yokota J, Muramatsu M, Sato K, Terada M and Tew KD:
Glutathione-S-transferase pi as a determinant of drug resistance in
transfectant cell lines. J Biol Chem. 265:4296–4301.
1990.PubMed/NCBI
|
|
24
|
Quan T, Qin Z, Xia W, Shao Y, Voorhees JJ
and Fisher GJ: Matrix-degrading metalloproteinases in photoaging. J
Investig Dermatol Symp Proc. 14:20–24. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jenkins G: Molecular mechanisms of skin
ageing. Mech Ageing Develop. 123:801–810. 2002. View Article : Google Scholar
|
|
26
|
Rittié L and Fisher GJ: UV-light-induced
signal cascades and skin aging. Ageing Res Rev. 1:705–720. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fisher GJ, Wang ZQ, Datta SC, Varani J,
Kang S and Voorhees JJ: Pathophysiology of premature skin aging
induced by ultraviolet light. N Engl J Med. 337:1419–1428. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee YM, Kang SM, Lee SR, Kong KH, Lee JY,
Kim EJ and Chung JH: Inhibitory effects of TRPV1 blocker on
UV-induced responses in the hairless mice. Arch Dermatol Res.
303:727–736. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee YM, Kim YK, Kim KH, Park SJ, Kim SJ
and Chung JH: A novel role for the TRPV1 channel in UV-induced
matrix metalloproteinase (MMP)-1 expression in HaCaT cells. J Cell
Physiol. 219:766–775. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Choi HK, Kim DH, Kim JW, Ngadiran S,
Sarmidi MR and Park CS: Labisia pumila extract protects skin cells
from photoaging caused by UVB irradiation. J Biosci Bioeng.
109:291–296. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim J, Lee CW, Kim EK, Lee SJ, Park NH,
Kim HS, Kim HK, Char K, Jang YP and Kim JW: Inhibition effect of
gynura procumbens extract on UV-B-induced matrix-metalloproteinase
expression in human dermal fibroblasts. J Ethnopharmacol.
137:427–433. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Krolikiewicz-Renimel I, Michel T,
Destandau E, Reddy M, André P, Elfakir C and Pichon C: Protective
effect of a Butea monosperma (Lam) Taub flowers extract against
skin inflammation: Antioxidant, anti-inflammatory and matrix
metalloproteinases inhibitory activities. J Ethnopharmacol.
148:537–543. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Piao MJ, Hyun YJ, Cho SJ, Kang HK, Yoo ES,
Koh YS, Lee NH, Ko MH and Hyun JW: An ethanol extract derived from
bonnemaisonia hamifera scavenges ultraviolet B (UVB)
radiation-induced reactive oxygen species and attenuates
UVB-induced cell damage in human keratinocytes. Mar Drugs.
10:2826–2845. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mu Y, Lv S, Ren X, Jin G, Liu J, Yan G, Li
W, Shen J and Luo G: UV-B induced keratinocyte apoptosis is blocked
by 2-selenium-bridged beta-cyclodextrin, a GPX mimic. J Photochem
Photobiol B. 69:7–12. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhu M and Bowden GT: Molecular mechanism
(s) for UV-B irradiation-induced glutathione depletion in cultured
human keratinocytes. Photochem Photobiol. 80:191–196. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Meloni M and Nicolay JF: Dynamic
monitoring of glutathione redox status in UV-B irradiated
reconstituted epidermis: Effect of antioxidant activity on skin
homeostasis. Toxicol In Vitro. 17:609–613. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Varma SD, Hegde KR and Kovtun S:
UV-B-induced damage to the lens in vitro: Prevention by caffeine. J
Ocul Pharmacol Ther. 24:439–444. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Misra RB, Babu GS, Ray RS and Hans RK:
Tubifex: A sensitive model for UV-B-induced phototoxicity.
Ecotoxicol Environ Saf. 52:288–295. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jiang GZ and Li JC: Protective effects of
ginsenoside Rg1 against colistin sulfate-induced neurotoxicity in
PC12 cells. Cell Mol Neurobiol. 34:167–172. 2014. View Article : Google Scholar
|
|
40
|
Ma J, Liu J, Wang Q, Yu H, Chen Y and
Xiang L: The beneficial effect of ginsenoside Rg1 on schwann cells
subjected to hydrogen peroxide induced oxidative injury. Int J Biol
Sci. 9:624–636. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Huang Q, Gao B, Jie Q, Wei BY, Fan J,
Zhang HY, Zhang JK, Li XJ, Shi J, Luo ZJ, et al: Ginsenoside-Rb2
displays anti-osteoporosis effects through reducing oxidative
damage and bone-resorbing cytokines during osteogenesis. Bone.
66:306–314. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu YW, Zhu X, Li W, Lu Q, Wang JY, Wei YQ
and Yin XX: Ginsenoside Re attenuates diabetes-associated cognitive
deficits in rats. Pharmacol Biochem Behav. 101:93–98. 2012.
View Article : Google Scholar
|
|
43
|
Ye R, Li N, Han J, Kong X, Cao R, Rao Z
and Zhao G: Neuroprotective effects of ginsenoside Rd against
oxygen-glucose deprivation in cultured hippocampal neurons.
Neurosci Res. 64:306–310. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kim ND, Pokharel YR and Kang KW:
Ginsenoside Rd enhances glutathione levels in H4IIE cells via
NF-kappaB-dependent gamma-glutamylcysteine ligase induction.
Pharmazie. 62:933–936. 2007.
|