Introduction
With high incidences of disability and other serious
consequences, traumatic spinal cord injury (TSCI) can result in
catastrophic damage to patients (1). Direct or indirect insult to the
backbone and spinal cord can lead to TSCI, which subsequently
affects the daily life of the patient and their family members
(2). TSCI morbidity is higher in
developed countries compared with developing countries. The USA
mortality rate is 20–45/100,000, and the rate of TSCI occurrence is
900/100,000 (3). Advancements to
the transportation, construction and mining industries have
increased the incidence of TSCI in China. The morbidity rate of
TSCI caused by earthquake disasters has reached 10%. Survey data
from the population of Beijing in 2005 indicated that the rate of
TSCI was 60.2/100,000 (4).
Due to serious complications, patients with TSCI may
require long periods of hospitalization. The TSCI clinical pathway,
a standardized method of patient care following TSCI, has achieved
great success (5).
Multidisciplinary investigation was used to develop a clinical
pathway for a standardized model of TSCI treatment. Following a
6-month trial period, the clinical pathway reduced the length of
patient hospitalization and the total costs (6). The TSCI clinical pathway has various
advantages and lasting influences on the overall quality of TSCI
treatment. The pathway reinforces to health professionals that
rehabilitation and nursing at the acute phase of TSCI are
inseparable (7). Additionally, the
diagnosis of TSCI is clear, the surgical invention is uniformed and
the complications are predictable. It is important to formulate a
similar standardized clinical pathway for the treatment of patients
with TSCI in China (8).
Allicin is a major component of garlic. Previous
studies have demonstrated that allicin has important health
benefits and medicinal effects (9). Allicin has been demonstrated to
inhibit the growth of pathogenic microorganisms, including certain
bacteria, viruses and fungi, and to inhibit tumor growth (10,11).
Additionally, it can prevent arteriosclerosis, stenocardia,
cerebral infarction, arrhythmia and hydrargyrism. Allicin can also
can reduce cholesterol and blood pressure, regulate blood sugar
levels, enhance the immune system and reduce oxidation (12). Therefore, the aim of the present
study was to investigate the use of allicin in TSCI, and to
elucidate whether the effects of allicin are mediated by the heat
shock protein 70 (HSP70), v-akt murine thymoma viral
oncogene homolog 1 (Akt) and inducible nitric oxide synthase (iNOS)
pathways.
Materials and methods
Animals, surgery and experimental
groups
Adult BALB/c mice (n=40; 30–40 g) were obtained from
The First Central Hospital of Baoding (Baoding, China). The mice
were housed at 24±1°C and maintained on a 12:12 h light/dark cycle.
The mice were provided with ad libitum access to food and
water. All experimental protocols were approved by the
Institutional Animal Care and Use Committee of The First Central
Hospital of Baoding and performed in accordance with the National
Institutes of Health Guide for the Care and Use of Laboratory
Animals.
All experimental mice were anesthetized by ketamine
(100 mg/kg; Sinopharm Chemical Reagent Co., Ltd., Shanghai, China)
and xylazine (10 mg/kg; Sinopharm Chemical Reagent Co., Ltd.)
injection via the caudal vein. The mice were randomly assigned into
five experimental groups (n=8 in each group), as follows: Sham
group; TSCI group; 1 mg/kg allicin group; 10 mg/kg allicin group;
and 50 mg/kg allicin group. Allicin (National Institute for the
Control of Pharmaceutical and Biological Products, Beijing, China)
and saline were delivered via intraperitoneal injection. The
anesthetized mice, excluding the sham group, received a laminectomy
at the T9 vertebral level using the Infinite Horizons impactor (75
kdyn; Precision Systems and Instrumentation, LLC, Fairfax Station,
VA, USA). Following TSCI, all mice were housed in warmed cages at
35–37°C. The mice received either 1, 5 or 10 mg/kg allicin. Sham
and TSCI mice received 2 ml sterile saline.
Locomotor recovery and the spinal cord
water content
Mice were relocated to the test environment (90×4
cm) where Basso, Beattie and Bresnahan (BBB) testing was performed
prior to and 1 week following TSCI. BBB scores range from 0 (no
observable hind-limb movements) to 21 (normal gait). Following
sacrifice by cervical dislocation under anesthesia, the spinal cord
samples were collected, weighed, recorded and dried at 80°C for
48–72 h. The spinal cord water content was calculated using the
following formula: [(Wet weight - dry weight)/wet weight] ×
100.
Enzyme-linked immunosorbent assay for
markers of oxidative stress and inflammation
Briefly, the spinal cord samples were collected and
homogenized, and the supernatant was collected by centrifugation at
12,000 × g and resuspended in lysis buffer containing protease
inhibitors (Beijing Biosynthesis Biotechnology Co., Ltd., Beijing,
China). The supernatant was collected and the protein concentration
was measured using a bicinchoninic acid (BCA) assay kit (Cell
Biolabs, Inc., San Diego, CA, USA). The activities of catalase
(CAT; #707002), superoxide dismutase (SOD; #706002), nuclear
factor-κB (NF-κB; #10007889) and tumor necrosis factor-α (TNF-α;
#500850) were measured using commercially available assay kits
(Cayman Chemical Company, Ann Arbor, MI, USA) following the
manufacturer's protocol.
Western blot analysis
Briefly, the spinal cord samples were homogenized,
and the supernatant was collected and harvested in lysis buffer
containing protease inhibitors. The protein concentration was
measured using the BCA protein assay kit. Protein samples (~60
μg) were separated by 10% SDS-PAGE and transferred onto a
polyvinylidene difluoride membrane (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The membrane was blocked with 5% non-fat milk
and incubated with the following primary antibodies, purchased from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA), at 4°C for 10–12
h: Rabbit polyclonal phosphatidylinositol-4,5-bisphosphate 3-kinase
(PI3K; 1:800; cat. no. sc-67306); rabbit polyclonal Akt (1:500;
cat. no. sc-8312); rabbit polyclonal phospho-Akt (1:500; cat. no.
sc-135650); rabbit polyclonal iNOS (1:500; cat. no. sc-8309); and
rabbit polyclonal β-actin (1:600; cat. no. sc-130656). Membranes
were then washed with Tris-buffered saline-Tween 20 and incubated
for 2 h at room temperature with goat anti-rabbit secondary
antibodies (Wuhan Boster Biological Technology, Co., Ltd., Wuhan,
China; cat. no. BA1054). The experiment was repeated in triplicate.
Membranes were exposed with enhanced chemiluminescence reagent
(Beyotime Institute of Biotechnology, Haimen, China) and quantified
with Image J version 3.0 analysis software (imagej.nih.gov/ij).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) of HSP70
Total RNA was isolated from spinal cord samples of
each group using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Total RNA (1 μl) was
used to synthesize cDNA using a TaqMan miRNA Reverse Transcription
kit (Applied Biosystems; Thermo Fisher Scientific, Inc.), following
treatment with DNase I for 10 min at 37°C. A SYBR Green PCR Master
mix (Applied Biosystems; Thermo Fisher Scientific, Inc.) and
Bio-Rad iQ5 Gradient Real-Time PCR system (Bio-Rad Laboratories,
Inc.) were used to perform qPCR and analyze the gene expression
levels of HSP70. The thermocycling conditions were as follows: 95°C
for 15 min; followed by 40 cycles of 94°C for 15 sec, 60°C for 30
sec and 72°C for 30 sec. The HSP70 primers used for all RT-qPCR
experiments were as follows: Forward, 5′-ACCAGGACACTGTTGAGTTC-3′;
and reverse, 5′-ACTCATCTCCGAGTTCACAC-3′. GAPDH was used as the
reference gene, with primers as follows: Forward,
5′-AAGGTGAAGGTCGGAGTCAA-3′; and reverse,
5′-AATGAAGGGGTCATTGATGG-3′. The qPCR data was analyzed using the
2−ΔΔCq method (13).
Mitochondrial reactive oxygen species
(ROS) and nicotinamide adenine dinucleotide (NADH) production
Briefly, the spinal cord samples were homogenized,
and the supernatant was collected, and ROS and NADH levels measured
using 10 μM dichlorofluorescein diacetate (Sigma-Aldrich,
St. Louis, MO, USA) for 1 h at 37°C in the dark. Mitochondrial ROS
levels were detected by fluorescence measurements at 480 nm
excitation and 530 nm emission wavelengths (Opsys MR; Dynex
Technologies, Chantilly, VA, USA). Mitochondrial NADH levels were
detected by measuring fluorescence at 360 nm excitation and 460 nm
emission wavelengths.
Statistical analysis
Statistical analysis was performed using SPSS
software, version 17.0 (SPSS Inc., Chicago, IL, USA). One-way
analysis of variance tests were performed to compare differences
between all groups, followed by least significant difference
post-hoc tests. Data are presented as the mean ± standard error.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Protective effects of allicin on
locomotor recovery in TSCI mice
The present study investigated the effect of allicin
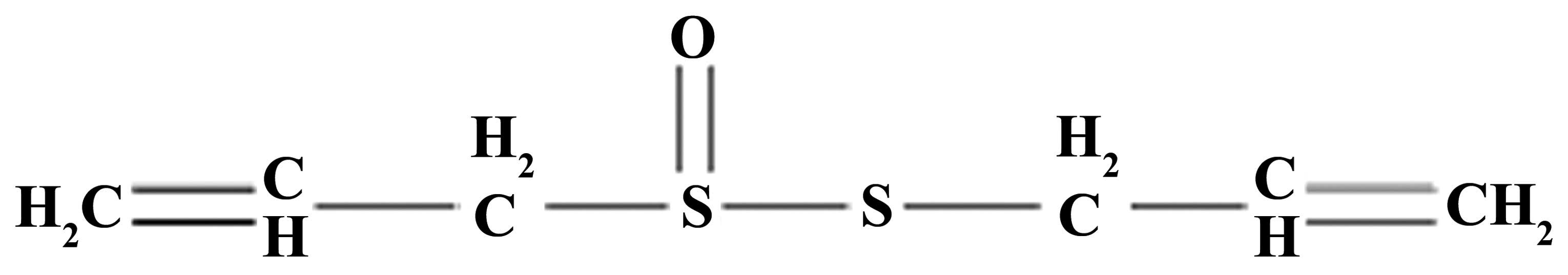
on locomotor recovery in TSCI mice. The chemical structure of
allicin is presented in Fig. 1. As
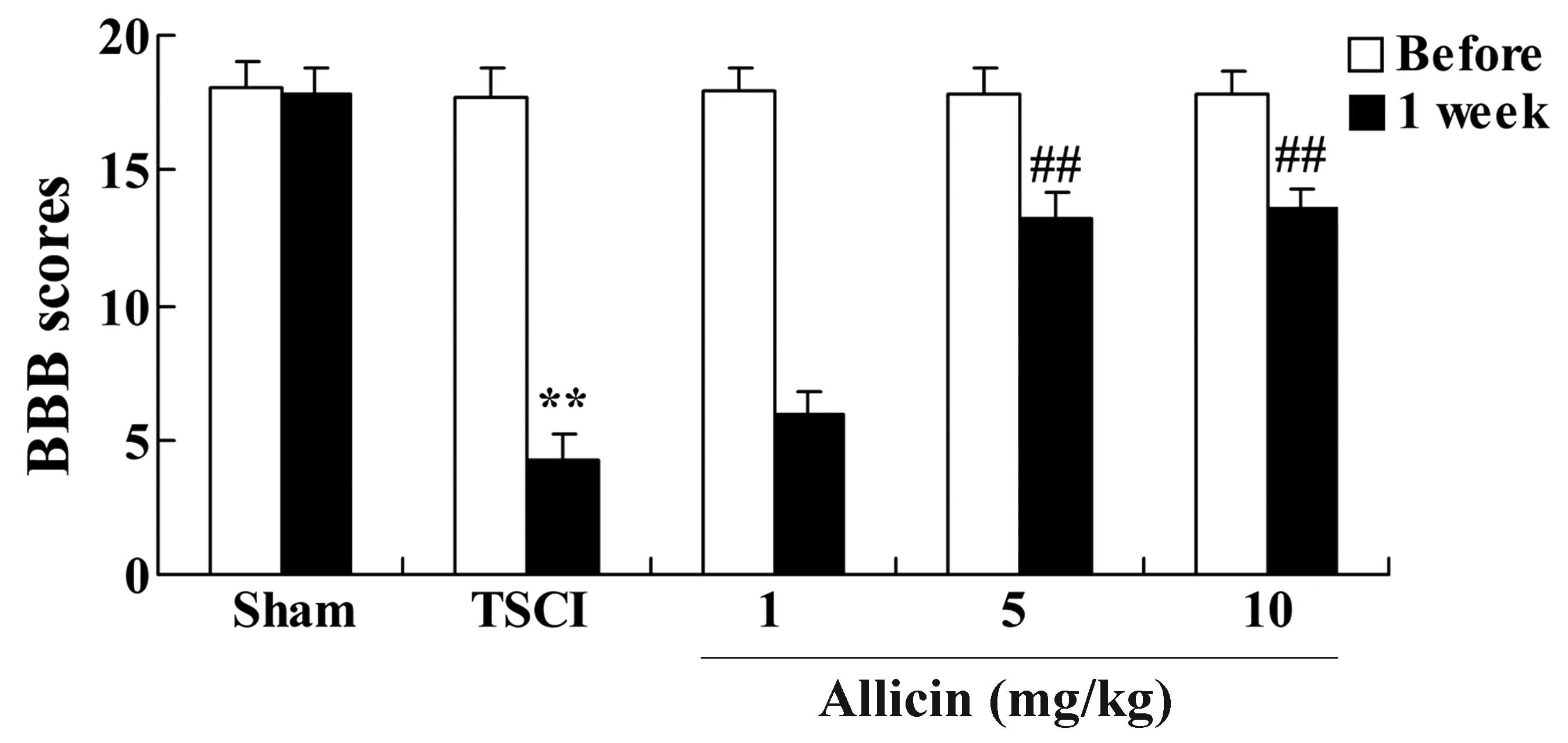
demonstrated in Fig. 2, 1 week
after TSCI, the BBB scores of TSCI mice were significantly reduced
compared with the sham group (P<0.001). Additionally, there was
a significant increase in the BBB scores following treatment with 5
or 10 mg/kg allicin compared with the TSCI model group (P=0.0023
and P=0.0017, respectively).
Protective effect of allicin on the
spinal cord water content in TSCI mice
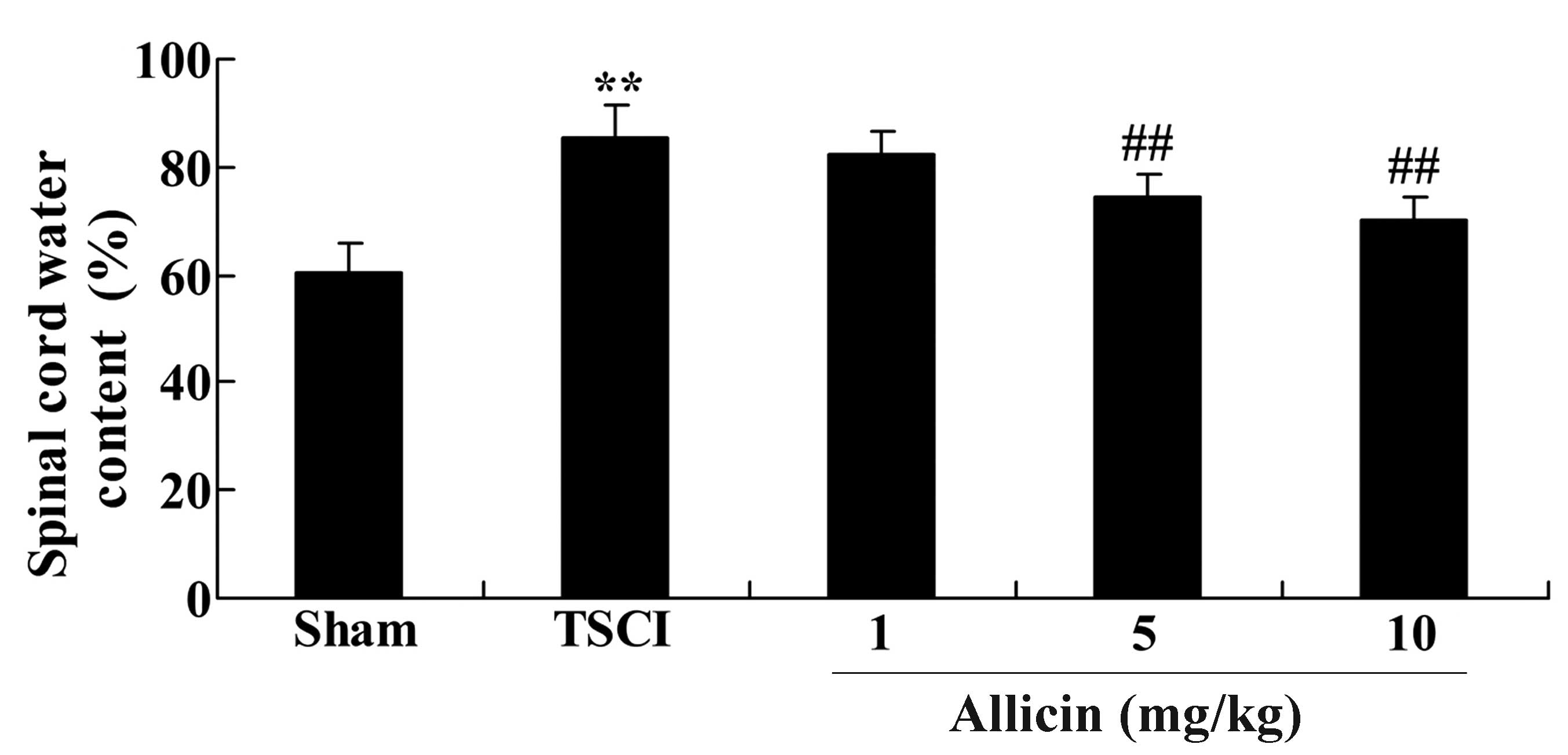
To confirm the effect of allicin on the spinal cord
water content in TSCI mice, the current study measured the spinal
cord water content following 1 week of treatment with allicin. The
results are presented in Fig. 3;
the spinal cord water content of TSCI mice was significantly
increased compared with the sham group (P=0.0041). After 1 week of
allicin treatment, the mice treated with 5 or 10 mg/kg allicin
exhibited a significant reduction in the spinal cord water content
when compared with the percentage water content in the TSCI model
group (P=0.0057 and P=0.0032, respectively).
Protective effect of allicin on oxidative
stress in TSCI mice
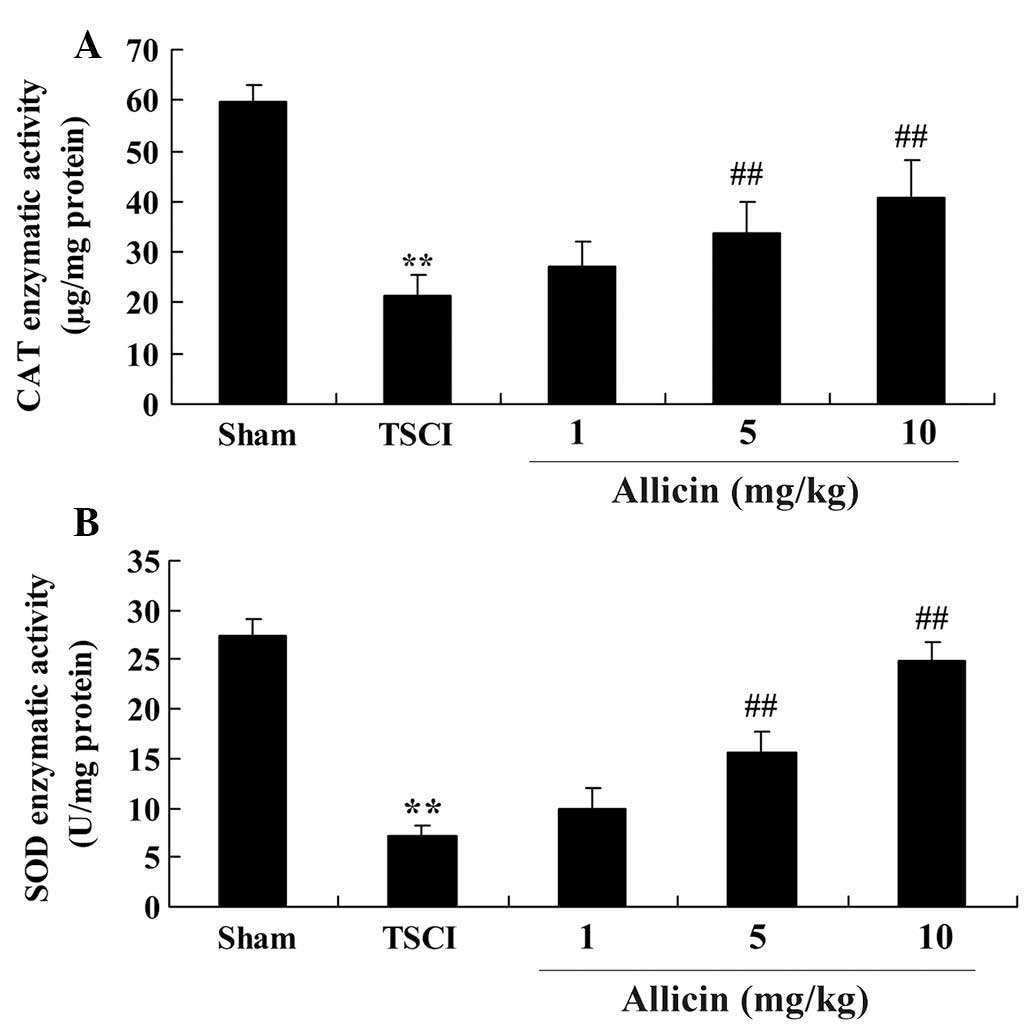
To determine whether the protective effect induced
by allicin was mediated by anti-oxidative activity, the enzymatic
activities of CAT and SOD were measured following allicin
treatment. As demonstrated in Fig.
4, the enzymatic activities of CAT and SOD were significantly
reduced in the TSCI group compared with the sham group (P<0.0001
and P=0.0001, respectively). CAT and SOD enzymatic activities were
increased by allicin treatment (5 or 10 mg/kg) compared with the
TSCI group (P=0.0023, P=0.0014, P=0.0005 and P=0.0003,
respectively).
Protective effect of allicin on
inflammation in TSCI mice
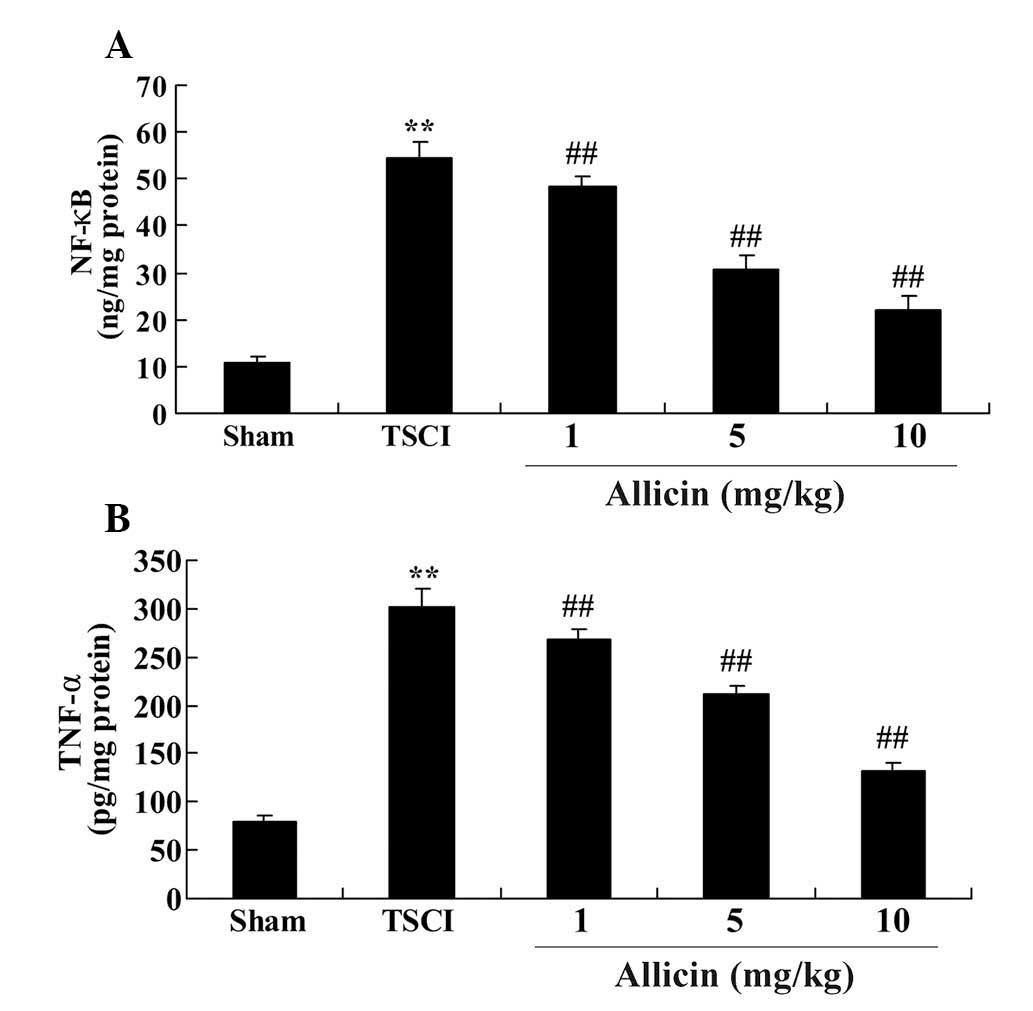
The current study also investigated the effect of
allicin on the anti-inflammatory signaling pathways. The results
demonstrated that the levels of NF-κB and TNF-α were increased in
the TSCI group compared with the sham group (each P<0.0001;
Fig. 5). Additionally, allicin
treatment, at all concentrations, significantly reduced the levels
of NF-κB and TNF-α compared with the TSCI model rats (P=0.0071,
P=0.0042, P=0.0052, P=0.0021, P=0.0030 and P=0.0016, respectively;
Fig. 5).
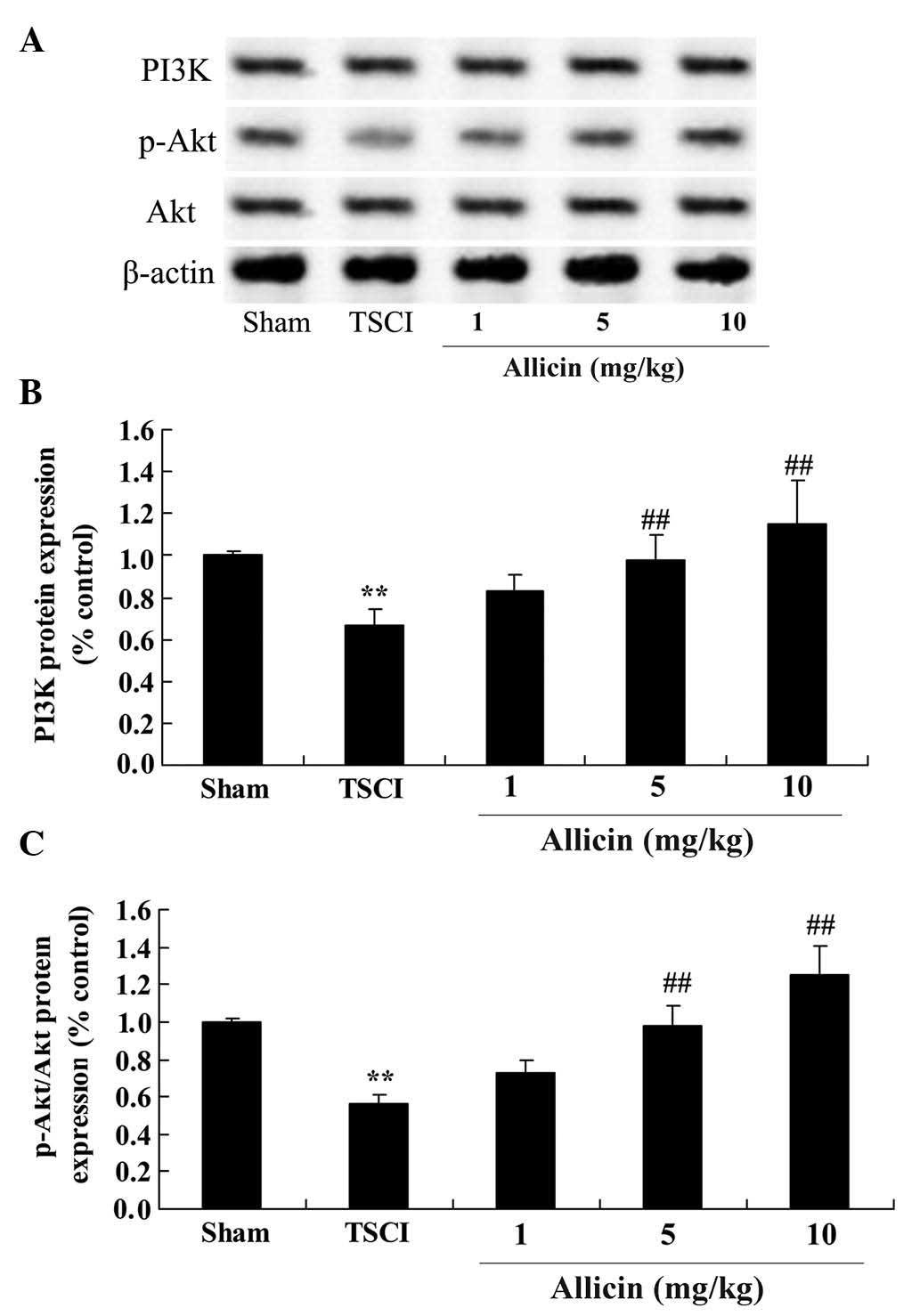
Protective effect of allicin on PI3K/Akt
in TSCI mice
To further investigate the protective effects of
allicin on TSCI, the current study investigated changes to the
PI3K/Akt pathways following allicin treatment of TSCI rats. As
demonstrated in Fig. 6, the
protein expression levels of PI3K and the phosphorylation of Akt in
TSCI mice were reduced compared with the sham group (P=0.0019 and
P=0.0006, respectively). However, treatment with 5 or 10 mg/kg
allicin significantly increased the protein expression levels of
PI3K and the phosphorylation of Akt compared with the TSCI mice
(P=0.0053, P=0.0022, P=0.0036 and P=0.0017, respectively).
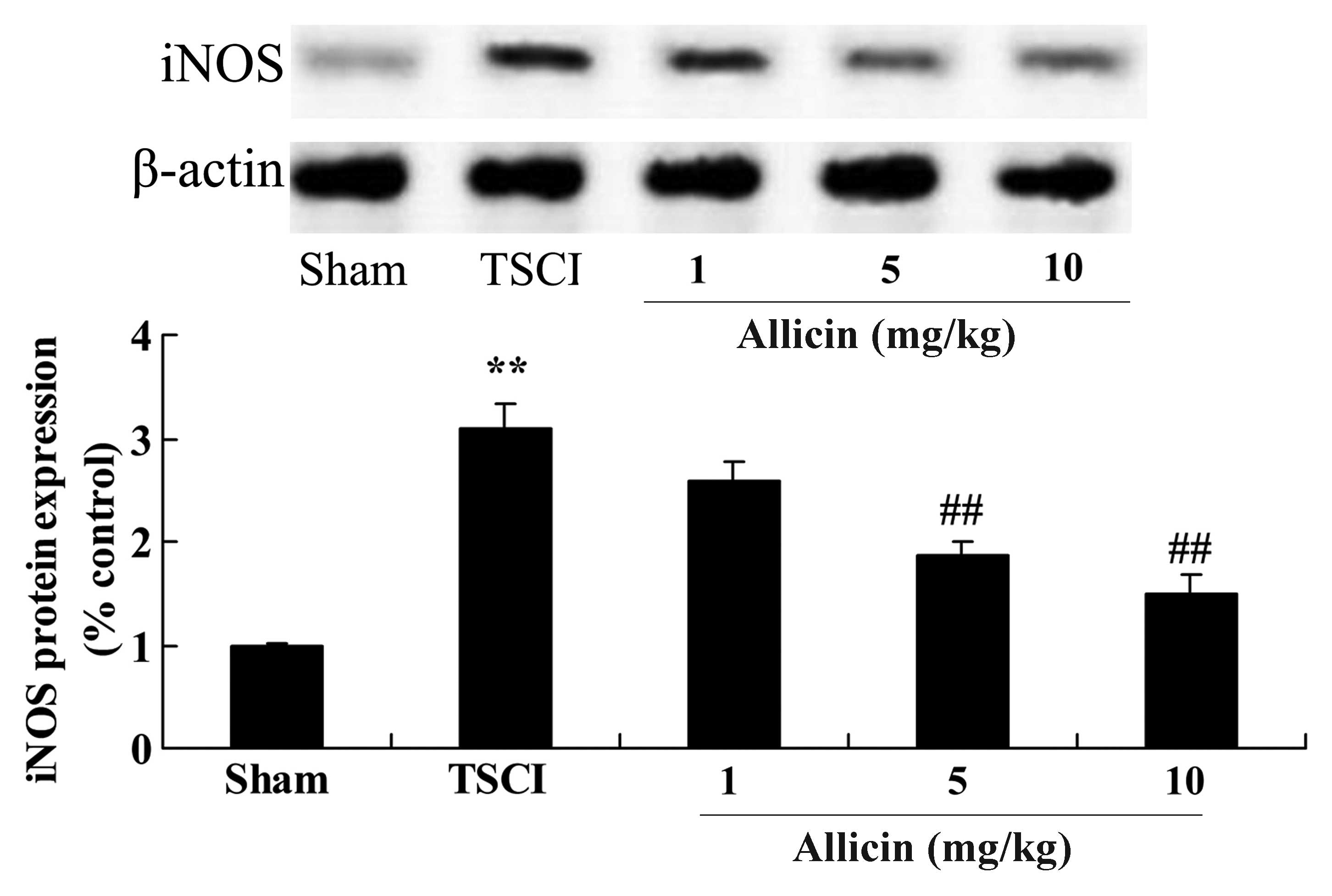
Protective effect of allicin on iNOS in
TSCI mice
To investigate the mechanisms that mediate the
protective effects of allicin against TSCI, iNOS protein expression
in TSCI mice was measured using western blot analysis. As presented
in Fig. 7, TSCI significantly
increased iNOS protein expression compared with the sham group
(P=0.0007). Administration of 5 and 10 mg/kg allicin significantly
inhibited the promotion of iNOS protein expression compared with
the TSCI model mice (P=0.0012 and P=0.0003, respectively).
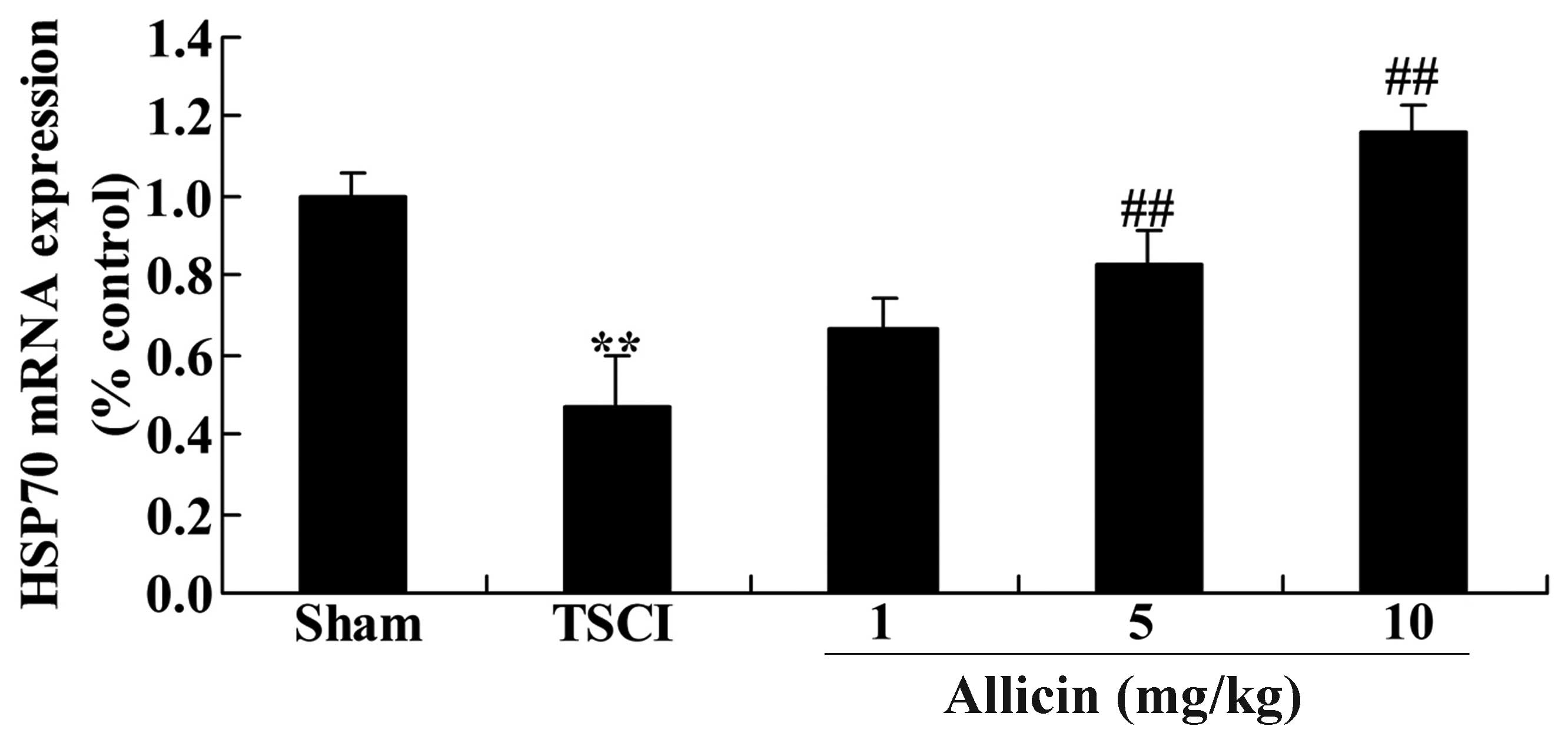
Protective effect of allicin on HSP70 in
TSCI mice
The present study also measured the HSP70 mRNA
expression levels to investigate the mechanisms that mediate the
effects of allicin on TSCI. As presented in Fig. 8, the HSP70 mRNA expression levels
were significantly reduced in the TSCI group compared with the sham
mice (P=0.0022). Furthermore, treatment with allicin (5 or 10
mg/kg) significantly increased the HSP70 mRNA expression levels
compared with the TSCI mice (P=0.0025 and P=0.0018, respectively;
Fig. 8).
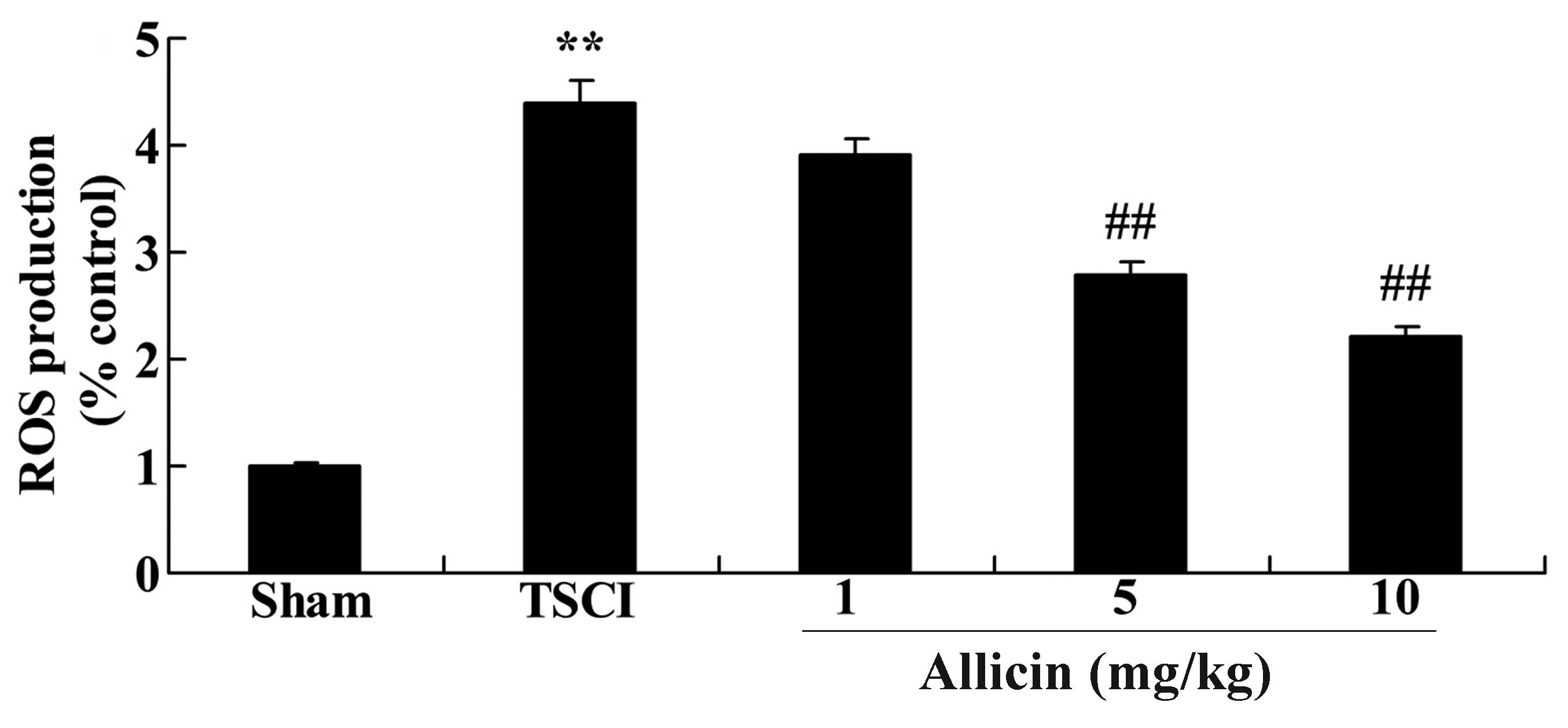
Protective effect of allicin on ROS
production in TSCI mice
To evaluate whether ROS are associated with the
protective effect of allicin on TSCI, ROS production was measured
in the experimental groups. The results of Fig. 9 demonstrate a significant increase
in ROS levels following TSCI compared with the sham group
(P<0.0001). Notably, treatment with 5 or 10 mg/kg allicin
significantly reduced the levels of ROS compared with the TSCI
group (P=0.0030 and P=0.0013, respectively).
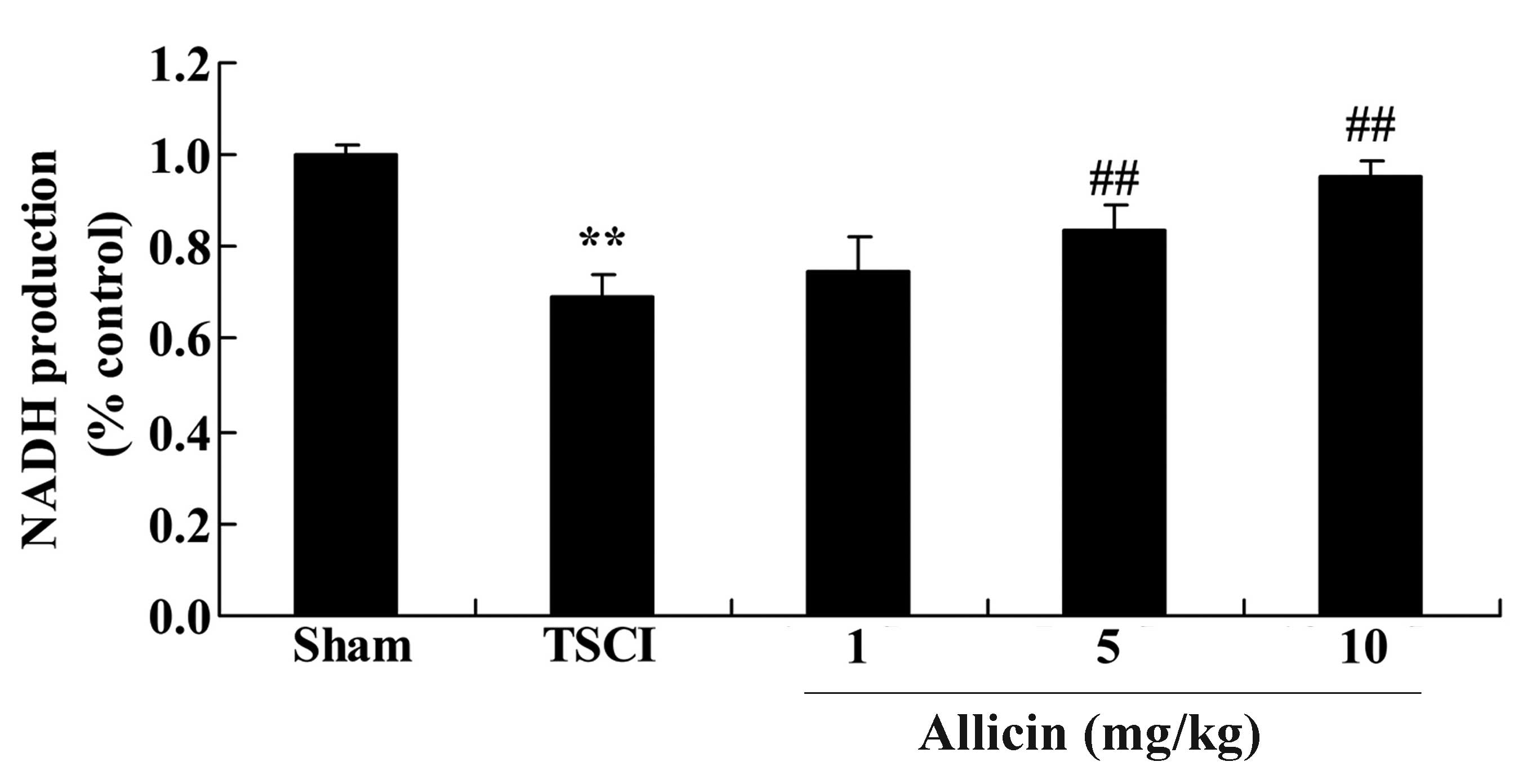
Protective effect of allicin on NADH
production in TSCI mice
To investigate the effects of allicin on NADH
production in TSCI mice, the NADH levels were analyzed following
TSCI and treatment with allicin. As demonstrated in Fig. 10, NADH levels were reduced in the
TSCI group compared with the sham group (P=0.0007). However,
allicin treatment (5 or 10 mg/kg) significantly elevated the NADH
levels compared with TSCI mice (P=0.0032 and P=0.0021).
Discussion
The development of the transportation and
construction industries has led to an increase in the number of
patients suffering from traumatic TSCI (14). TSCI places a heavy burden on the
families of patients and society, as the condition leads to high
disability rates, high costs, loss of labor forces and other
serious complications, including infection, bedsores and deep
venous thrombosis (15). Certain
countries have conducted numerous detailed studies on TSCI,
however, few have been performed in China (15). The current study confirms the
protective effect of allicin. Allicin treatment increased locomotor
recovery, and reduced the spinal cord water content, oxidative
stress and inflammation in TSCI mice. Liu et al (9) reported that allicin protects spinal
cord neurons via suppression of the oxidative stress pathway. Li
et al (16) indicated that
allicin alleviates inflammation in Caco-2 cells.
Akt is considered a vital regulatory factor for
signal transduction, and together with purine signals is important
in nervous physiological and pathological processes (17). Akt signaling facilitates the
growth, differentiation, survival and regeneration of neurons via
the P2 receptor following injury of the central nervous system, and
mediates the remodeling of impaired neural tissues (18). As a crucial mediator of protein
kinase cascades, following impairment of the central nervous
system, ATP can activate the PI3K/Akt pathways, which are important
for the survival and repair of neurons (19). In the present study, treatment with
allicin significantly increased the levels of PI3K and phospho-Akt
in TSCI rats. Liu et al (20) demonstrated that allicin protects
against cardiac hypertrophy and fibrosis through the activation of
the ROS-dependent Akt signaling pathways.
Numerous studies have suggested that injury of the
central nervous system caused by ischemia and other damage
increases the expression levels of NOS (21,22).
Excessive NO synthesis occurs, which directly impairs the function
of the nervous system. As a micromolecule and free radical, NO
possesses various bioactivities, including vasodilatation, neural
information transmission and cytotoxic effects (23). Excessive NO can induce cell
apoptosis, which is characteristic of the secondary changes
exhibited following TSCI (24). In
the current study, administration of allicin significantly
inhibited the promotion of iNOS protein expression in TSCI mice.
Liu et al (9) reported that
allicin protects spinal cord neurons through suppression of the NOS
pathway. Zhou et al (25)
also demonstrated that allicin protects against mechanical trauma
injury via Akt- and mitogen-activated protein kinase 1-mediated
regulation of the NOS pathway.
HSPs are a widely expressed, evolutionarily
conserved family of proteins. They function as molecular
chaperones, and are important for the transport and folding of
proteins during cellular stress. HSP70 proteins are considered to
be the principle family members (26). HSP70 protein expression is
stimulated following stress, including heat shock, ischemia, oxygen
deficit, viral infection and mechanical injuries (27). Previous studies on HSP70 in TSCI
have observed that, following injury, the HSP70 protein expression
levels were increased (26). In
the present study, treatment with allicin significantly increased
the levels of HSP70 mRNA expression in TSCI mice. Liu et al
(9) previously reported that
allicin protects spinal cord neurons through the regulation of the
HSP70 pathway.
Numerous studies have investigated the secondary
damage mechanisms following TSCI. It was previously demonstrated
that ROS are a crucial mediator of the secondary damages caused by
SCI. Following TSCI, a series of pathological changes occur in the
spinal cord tissue, including edema, bleeding, anoxia and ischemia
(28). These changes lead to
mitochondrial dysfunction and increased of ROS production (29). In the case of the rate of ROS
production exceeding the scavenging activity of the defense system,
ROS accumulates in cells. In the present study, allicin
significantly reduced the levels of ROS following TSCI. A previous
study demonstrated that allicin protects spinal cord neurons via
suppression of ROS production (9).
Chan et al (30)
demonstrated that allicin protects rat cardiomyoblasts by
inhibiting the generation of ROS.
NADH can protect cells from injury via oxidation and
peroxidation of heavy metal chromate-induced hemoglobin (31). Previous research has demonstrated
that NADH can effectively eliminate oxygen free radicals, inhibit
the production of ROS, stabilize the cytomembrane, activate
multi-enzyme systems, and enhance the synthesis and metabolism of
nucleic acids, proteins and polysaccharides. Furthermore, NADH can
regulate and improve metabolism (32). Additionally, NADH is important for
the production of the materials required for regeneration, repair
and protection. These results demonstrate the importance of NADH as
a mediator of the allicin-induced locomotor recovery of TSCI mice.
The present study demonstrated that allicin significantly increased
NADH production, which had been reduced by TSCI. Rabinkov et
al (33) reported that the
predominant biological action of allicin may be attributed to its
effect on NADH levels.
In conclusion, the findings of the current study
demonstrated that the beneficial effects of allicin following TSCI
are mediated via the regulation of oxidative stress and
inflammation. The protective effects of allicin were dependent on
the promotion of HSP70 protein expression and NADH levels, and the
inhibition of iNOS and ROS levels following TSCI. The results of
the present study indicate that the protective effects of allicin
may be useful in the treatment of TSCI.
References
|
1
|
Schwab JM, Zhang Y, Kopp MA, Brommer B and
Popovich PG: The paradox of chronic neuroinflammation, systemic
immune suppression, autoimmunity after traumatic chronic spinal
cord injury. Exp Neurol. 258:121–129. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Konya D, Gercek A, Akakin A, Akakin D,
Tural S, Cetinel S, Ozgen S and Pamir MN: The effects of
inflammatory response associated with traumatic spinal cord injury
in cutaneous wound healing and on expression of transforming growth
factor-beta1 (TGF-beta1) and platelet-derived growth factor
(PDGF)-A at the wound site in rats. Growth Factors. 26:74–79. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yin KJ, Kim GM, Lee JM, He YY, Xu J and
Hsu CY: JNK activation contributes to DP5 induction and apoptosis
following traumatic spinal cord injury. Neurobiol Dis. 20:881–889.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chan SC and Chan AP: Rehabilitation
outcomes following traumatic spinal cord injury in a tertiary
spinal cord injury centre: A comparison with an international
standard. Spinal Cord. 43:489–498. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Franceschini M, Di Clemente B, Citterio A
and Pagliacci MC: Follow-up in persons with traumatic spinal cord
injury: Questionnaire reliability. Eura Medicophys. 42:211–218.
2006.PubMed/NCBI
|
|
6
|
Dobkin BH, Apple D, Barbeau H, Basso M,
Behrman A, Deforge D, Ditunno J, Dudley G, Elashoff R, Fugate L, et
al: Methods for a randomized trial of weight-supported treadmill
training versus conventional training for walking during inpatient
rehabilitation after incomplete traumatic spinal cord injury.
Neurorehabil Neural Repair. 17:153–167. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van Weert KC, Schouten EJ, Hofstede J, van
de Meent H, Holtslag HR and van den Berg-Emons RJ: Acute phase
complications following traumatic spinal cord injury in Dutch level
1 trauma centres. J Rehabil Med. 46:882–885. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wong AM, Leong CP, Su TY, Yu SW, Tsai WC
and Chen CP: Clinical trial of acupuncture for patients with spinal
cord injuries. Am J Phys Med Rehabil. 82:21–27. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu SG, Ren PY, Wang GY, Yao SX and He XJ:
Allicin protects spinal cord neurons from glutamate-induced
oxidative stress through regulating the heat shock protein
70/inducible nitric oxide synthase pathway. Food Funct. 6:321–330.
2015. View Article : Google Scholar
|
|
10
|
Adetumbi MA and Lau BH: Allium sativum
(garlic)–a natural antibiotic. Med Hypotheses. 12:227–237. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Borlinghaus J, Albrecht F, Gruhlke MC,
Nwachukwu ID and Slusarenko AJ: Allicin: Chemistry and biological
properties. Molecules. 19:12591–12618. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang W, Wang Y, Cao YG, Qi HP, Li L, Bai
B, Liu Y and Sun HL: Antiarrhythmic effects and ionic mechanisms of
allicin on myocardial injury of diabetic rats induced by
streptozotocin. Naunyn Schmiedebergs Arch Pharmacol. 386:697–704.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sieber MW, Claus RA, Witte OW and Frahm C:
Attenuated inflammatory response in aged mice brains following
stroke. PLoS One. 6:e262882011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu Q, Ning GZ, Li YL, Feng HY and Feng SQ:
Factors affecting the length of stay of patients with traumatic
spinal cord injury in Tianjin, China. J Spinal Cord Med.
36:237–242. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang H, Xiang Q, Li C and Zhou Y:
Epidemiology of traumatic cervical spinal fractures and risk
factors for traumatic cervical spinal cord injury in China. J
Spinal Disord Tech. 26:E306–E313. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li C, Lun W, Zhao X, Lei S, Guo Y, Ma J
and Zhi F: Allicin alleviates inflammation of
trinitrobenzenesulfonic acid-induced rats and suppresses P38 and
JNK pathways in Caco-2 cells. Mediators Inflamm. 2015:4346922015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu L, Xie J, Xin N and Wang Z: Panax
notoginseng saponins promote wound repair of anterior cruciate
ligament through phosphorylation of PI3K, AKT and ERK. Int J Clin
Exp Pathol. 8:441–449. 2015.PubMed/NCBI
|
|
18
|
Yune TY, Park HG, Lee JY and Oh TH:
Estrogen-induced Bcl-2 expression after spinal cord injury is
mediated through phosphoinositide-3-kinase/Akt-dependent CREB
activation. J Neurotrauma. 25:1121–1131. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jung SY, Kim DY, Yune TY, Shin DH, Baek SB
and Kim CJ: Treadmill exercise reduces spinal cord injury-induced
apoptosis by activating the PI3K/Akt pathway in rats. Exp Ther Med.
7:587–593. 2014.PubMed/NCBI
|
|
20
|
Liu C, Cao F, Tang QZ, Yan L, Dong YG, Zhu
LH, Wang L, Bian ZY and Li H: Allicin protects against cardiac
hypertrophy and fibrosis via attenuating reactive oxygen
species-dependent signaling pathways. J Nutr Biochem. 21:1238–1250.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang Y, Gong FL, Zhao GB and Li J:
Chrysin suppressed inflammatory responses and the inducible nitric
oxide synthase pathway after spinal cord injury in rats. Int J Mol
Sci. 15:12270–12279. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Abbasi Habashi S, Sabouni F, Moghimi A and
Ansari Majd S: Modulation of lipopolysaccharide stimulated nuclear
factor kappa B mediated iNOS/NO production by bromelain in rat
primary microglial cells. Iran Biomed J. 20:33–40. 2016.
|
|
23
|
Ren B, Zhang YX, Zhou HX, Sun FW, Zhang
ZF, Wei Z, Zhang CY and Si DW: Tanshinone IIA prevents the loss of
nigrostriatal dopaminergic neurons by inhibiting NADPH oxidase and
iNOS in the MPTP model of Parkinson's disease. J Neurol Sci.
348:142–152. 2015. View Article : Google Scholar
|
|
24
|
Chen M, Xia X, Zhu X, Cao J, Xu D, Ni Y,
Liu Y, Yan S, Cheng X, Liu Y and Wang Y: Expression of SGTA
correlates with neuronal apoptosis and reactive gliosis after
spinal cord injury. Cell Tissue Res. 358:277–288. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhou YF, Li WT, Han HC, Gao DK, He XS, Li
L, Song JN and Fei Z: Allicin protects rat cortical neurons against
mechanical trauma injury by regulating nitric oxide synthase
pathways. Brain Res Bull. 100:14–21. 2014. View Article : Google Scholar
|
|
26
|
Sharma HS, Olsson Y and Westman J: A
serotonin synthesis inhibitor, p-chlorophenylalanine reduces the
heat shock protein response following trauma to the spinal cord: An
immunohistochemical and ultrastructural study in the rat. Neurosci
Res. 21:241–249. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iguchi M, Littmann AE, Chang SH, Wester
LA, Knipper JS and Shields RK: Heat stress and cardiovascular,
hormonal, and heat shock proteins in humans. J Athl Train.
47:184–190. 2012.PubMed/NCBI
|
|
28
|
Bains M and Hall ED: Antioxidant therapies
in traumatic brain and spinal cord injury. Biochim Biophys Acta.
1822:675–684. 2012. View Article : Google Scholar
|
|
29
|
Yeo JE, Kim JH and Kang SK: Selenium
attenuates ROS-mediated apoptotic cell death of injured spinal cord
through prevention of mitochondria dysfunction; in vitro and in
vivo study. Cell Physiol Biochem. 21:225–238. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chan JY, Tsui HT, Chung IY, Chan RY, Kwan
YW and Chan SW: Allicin protects rat cardiomyoblasts (H9c2 cells)
from hydrogen peroxide-induced oxidative injury through inhibiting
the generation of intracellular reactive oxygen species. Int J Food
Sci Nutr. 65:868–873. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jin W, Ming X, Hou X, Zhu T, Yuan B, Wang
J, Ni H, Jiang J, Wang H and Liang W: Protective effects of
erythropoietin in traumatic spinal cord injury by inducing the Nrf2
signaling pathway activation. J Trauma Acute Care Surg.
76:1228–1234. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee YS, Sindhu RK, Lin CY, Ehdaie A, Lin
VW and Vaziri ND: Effects of nerve graft on nitric oxide synthase,
NAD(P)H oxidase, and antioxidant enzymes in chronic spinal cord
injury. Free Radic Biol Med. 36:330–339. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rabinkov A, Miron T, Konstantinovski L,
Wilchek M, Mirelman D and Weiner L: The mode of action of allicin:
Trapping of radicals and interaction with thiol containing
proteins. Biochim Biophys Acta. 1379:233–244. 1998. View Article : Google Scholar : PubMed/NCBI
|