Introduction
The fungus Penicillium oxalicum
(Trichocomaceae) is known for its capacity to produce novel
acetogenins, sesquiterpenoids, diterpenoids, prostanoids and
steroids that exhibit bioactivities including antitumor,
antituberculosis, anti-inflammation, and antioxidant effects
(1,2). Penicillium oxalicum SCSGAF
0023 was isolated from the South China Sea gorgonian Muricella
flexuosa by Zhang et al (3). Previous studies on the
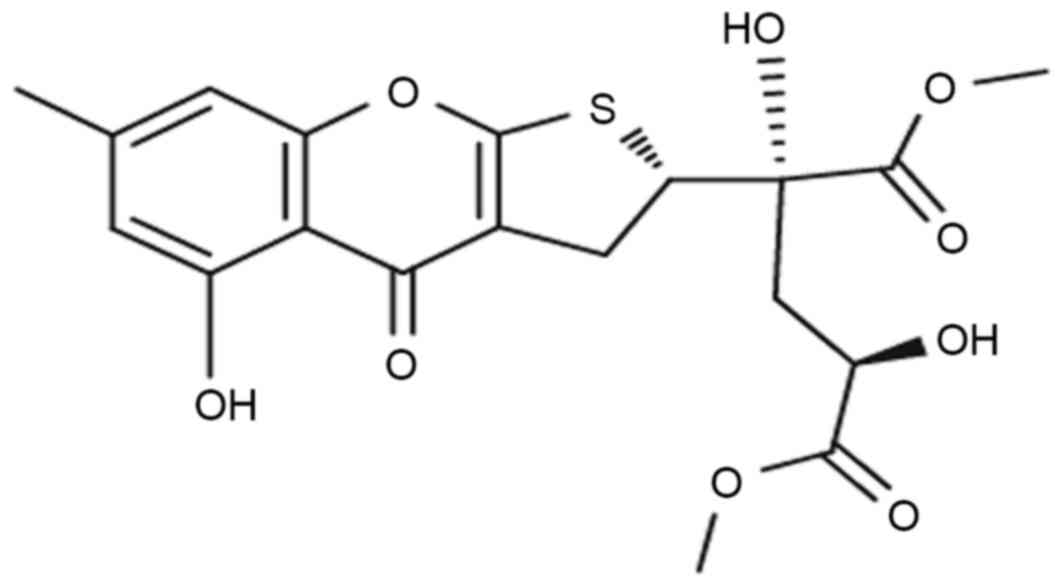
dihydrothiophene-condensed chromone oxalicumone A (POA; chemical
structure in Fig. 1), isolated
from a culture broth of Penicillium oxalicum SCSGAF 0023
(4), demonstrated that it exhibits
significant antitumor activity against several carcinoma cell
lines, including A375, SW-620 and HeLa, with IC50 values
of 8.9, 7.8 and 18.4 µM, respectively (5). This suggests that POA might serve as
a candidate for a novel antitumor drug. However, whether POA is
toxic to normal cells, in vivo or in vitro, has not
been reported to date, and this would be a major limitation to the
clinical application of POA. In the drug discovery and development
pipeline, toxicity information is important in guiding
pharmaceutical application and optimization. Thus, investigations
of the possible toxicity and its mechanisms are vital for POA
clinical development, in order to determine drug safety.
The function of renal proximal tubules is to
concentrate the glomerular filtrate by reabsorption of essential
molecules, and hence they are easily injured by drugs and chemicals
that are eliminated through the kidney (6). Since the proximal tubule is one of
the most common sites of injury by nephrotoxic drugs, screening and
understanding the toxicity potential of drug candidates on renal
proximal tubule cells is important as a first step to drug
discovery.
In the present study, several biological endpoints
were examined to assess the possible toxic effects of POA on human
kidney-2 (HK-2) cells in vitro and the underlying
mechanism.
Materials and methods
Materials
D/F12 medium and fetal bovine serum (FBS) were
purchased from Hyclone; GE Healthcare Life Sciences (Logan, UT,
USA) and Biological Industries (Kibbutz Beit-Haemek, Israel),
respectively. The Cell Counting Kit-8 (CCK-8) was purchased from
Dojindo Molecular Technologies, Inc. (Kumamoto, Japan). Trypsin,
dimethyl sulfoxide (DMSO), and Hoechst 33258 were purchased from
Sigma-Aldrich; Merck Millipore (Darmstadt, Germany). The Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) double
staining kit, DNA content quantitation assay kit,
5,5,6,6-tetra-chloro-1,1,3,3-tetraethylbenzimidazolyl-carbocyanine
iodide (JC-1) dye and caspase-3 activity assay kit were purchased
from Nanjing KeyGen Biotech. Co., Ltd. (Nanjing, China).
Glutathione (GSH; cat. no. CEA294Ge) and
N-acetyl-β-D-Glucosaminidase (NAG; cat. no. CSB-E07444 m) ELISA
kits were purchased from Uscn Life Science, Inc. (Wuhan, China) and
CUSABIO Biotech. Co., Ltd. (Wuhan, China), respectively.
Radioimmunoprecipitation assay (RIPA) lysis buffer and enhanced
chemiluminescence (ECL) kit were purchased from Biomiga, Inc. (San
Diego, CA, USA) and Beyotime Institute of Biotechnology (Haimen,
China), respectively. The bicinchoninic acid (BCA) protein assay
kit was purchased from BioTeke Corporation (Beijing, China). Fas
cell surface death receptor (Fas; dilution, 1:4,000; cat. no.
ab133619), B-cell lymphoma 2 apoptosis regulator (Bcl-2; dilution,
1:4,000; cat. no. ab182858), Bcl-2 associated protein X apoptosis
regulator (Bax; dilution, 1:4,000; cat. no. ab32503) and β-actin
(dilution, 1:4,000; cat. no. ab16039) antibodies were purchased
from Abcam (Cambridge, UK). Horseradish peroxidase-conjugated goat
anti-rabbit immunoglobulin G (dilution, 1:80,000; cat. no. IH-0011)
was obtained from Boster Systems, Inc. Pleasanton. CA, USA. All
other chemicals were obtained from Nanjing Jiancheng Bioengineering
Institute (Nanjing, China). POA was provided by the South China Sea
Institute of Oceanology (Guangzhou, China). The structure of POA
was determined by infrared, nuclear magnetic resonance and mass
spectrometry and its purity of >98% was determined by high
performance liquid chromatography. POA was dissolved in DMSO and
phosphate buffer saline (PBS) to obtain stock solutions (40 mM),
which were stored at −20°C. Prior to use in an experiment, the
stock solution was diluted to the indicated concentrations with
culture medium. During the experiments, the DMSO content in the
medium never exceeded 0.5% (v/v).
Cell culture
HK-2 cells were obtained from the American Type
Culture Collection (Manassas, VA, USA) and were grown in D/F12
supplemented with 10% FBS in a humidified incubator at 37°C in the
presence of 5% CO2. The culture medium was changed every
2 days. Cells for assays were detached by a solution of 0.25%
trypsin and 0.02% EDTA.
CCK-8 cell viability assay
HK-2 cell viability was evaluated by the CCK-8
assay. Briefly, HK-2 cells (1×104cells/well) were seeded
in 96-well microplates and then cultured in D/F12 growth medium for
24 h. Subsequently, the medium was replaced with D/F12 growth
medium containing 10, 20, 30, 40, 50, 60, 70, 80, 90 or 100 µM POA.
Cells containing equal volumes of cell culture medium but no POA (0
µM), were used as a control in each experiment throughout the
study. Following exposure to POA for 24, 48 or 72 h, 10 µl of the
CCK-8 assay solution was added into each well, followed by
incubation of the microplates at 37°C in 5% CO2/95% air
for 2 h. Finally, absorption was measured at 450 nm using a
microplate reader (PerkinElmer, Inc., Waltham, MA, USA), with a
reference wavelength of 650 nm (7). Three different experiments were
performed and the average value was calculated.
Morphological changes in the cell and
nucleus
Morphological changes in the HK-2 cells were
evaluated by phase contrast optical microscopy (Leica Microsystems
Gmbh, Wetzlar, Germany). Morphological changes of the cell nuclei
were evaluated by fluorescent visualization with Hoechst 33258
staining. Briefly, cells (4×104 cells/well) cultured on
slides were treated with 0, 20 or 40 µM POA for 24 h. Following
treatment, cells were washed with PBS, fixed with 4%
paraformaldehyde for 10 min and then incubated for 5 min with 5
mg/ml Hoechst 33258 fluorescent dye. The cells were then washed,
dried, and photographed using a fluorescence microscope.
Annexin V/PI staining assay
The early apoptosis rate was measured using Annexin
V-FITC/PI double staining and a Accuri™ C6 FACSCalibur flow
cytometer (BD Biosciences, San Jose, CA, USA) with BD CFlow
Software v.264.15 (BD Biosciences). Following treatment with 0, 20
or 40 µM POA for 24 h, 5×105 HK-2 cells were harvested
by centrifugation at 800 × g for 5 min at 4°C, washed twice
with ice-cold PBS and resuspended in 500 µl binding buffer,
followed by the addition of 5 µl Annexin V-FITC conjugate and 5 µl
PI buffer, according to the manufacturer's protocol. Following
incubation in the dark for 15 min at room temperature, the cells
were analyzed by flow cytometry. Each determination is based on the
acquisition of 10,000 events (8).
Cell cycle phase analysis
Following treatment with 0, 20 or 40 µM POA for 24
h, 5×105 cells were collected by centrifugation at 800 ×
g for 5 min at 4°C, washed twice with PBS, and then fixed
with 70% chilled ethanol for 12 h. Following fixation, cells were
washed twice with PBS and incubated in PBS containing 50 mg/ml PI,
1 mg/ml RNase A and Triton X-100 (0.5%) at 4°C for 30 min in the
dark. The fluorescence emitted from the PI-DNA complex was measured
using Accuri™ C6 FACScan flow cytometry (BD Biosciences) with BD
CFlow Software v.264.15 (BD Biosciences). The cells with nuclei
with sub-G1 content were considered apoptotic cells (9).
Activation of caspase 3
A caspase 3 activity assay kit was used to measure
caspase 3 activity, as previously described (10). In brief, 1×106 cells
were treated with 0, 20, 40, or 80 µM POA for 24 h, then cells were
harvested by centrifugation at 800 × g for 5 min at 4°C,
washed twice with ice-cold PBS, resuspended in lysis buffer and
left on ice for 60 min. The lysate was centrifuged at 12,000 ×
g at 4°C for 5 min. The cell supernatant was incubated with
the enzyme specific colorimetric substrate
acetyl-Asp-Gla-Val-Asp-phosphorylated nitroanilide (Ac-DEVD-pNA) in
assay buffer for 2 h at 37°C. The concentration of pNA from the
Ac-DEVD-pNA substrate was determined by the optical absorbance at
405 nm using a microplate reader (PerkinElmer, Inc., Waltham, MA,
USA).
Assays of antioxidant status
For assays of GSH, superoxide dismutase (SOD),
malondialdehyde (MDA), nitric oxide (NO),
N-acetyl-β-D-glucosaminidase (NAG), lactate dehydrogenase (LDH) and
reactive oxygen species (ROS), HK-2 cells (1×106
cells/well) were seeded in 6-well plates and then cultured in D/F12
growth medium for 24 h. Subsequently, the medium was replaced with
D/F12 growth medium containing 0, 20, 40, or 80 µM POA and
incubated for 24 h at 37°C. Following treatment, 1×106
cells were harvested by centrifugation at 800 × g for 5 min
at 4°C, then lysed on ice for 30 min using lysis buffer (50 mmol
Tris-HCl, 1.0 mmol/l EDTA, 150 mmol/l NaCl and 0.1% SDS), and
centrifuged once more at 5,000 × g for 10 min at 4°C. The
supernatant was used for the enzymatic assays. For assessment of
extracellularly released molecules, the culture medium was analyzed
following the 24 h POA treatment.
GSH content
The antioxidant enzyme GSH was measured using a GSH
ELISA kit, as per the manufacturer's instructions. GSH
concentration was determined by measurement of the absorbance at
450 nm using an ELISA Reader (PerkinElmer, Inc., Waltham, MA,
USA).
SOD activity
SOD is a scavenger of superoxide. SOD activity was
detected using the xanthine/xanthine oxidase method based on the
production of O2− anions (11). HK-2 cells and lysates were prepared
as described above, but in 25 cm2 culture flasks at
1×106 cells/flask. The SOD activity in the cell lysates
was determined using a formula calculation based on absorbance
values at 550 nm (11). Activity
of SOD is expressed as units per mg of cellular protein (U/mg
prot).
Determination of lipid
peroxidation
The concentration of MDA, an end product generated
from lipid peroxidation, was measured using an MDA detection kit
according to the manufacturer's instructions (12). Briefly, MDA reacts with
thiobarbituric acid (TBA) at 95–100°C in acidic conditions and the
reaction produces a pink MDA-TBA conjugate, which can be measured
using an EnSpire multi-mode microplate reader (PerkinElmer, Inc.,
Waltham, MA, USA) at 532 nm. The cellular MDA concentration was
expressed as nmol/mg of cellular protein (13).
Assessment of NO
NO, a potentially toxic molecule, is a labile,
diffusible product of mammalian cells. It serves as a short-lived
messenger molecule involved in diverse biological phenomena such as
cytotoxicity (14). NO levels were
estimated by measuring the accumulation of nitrite in the culture
medium, which is the resulting byproduct of NO metabolites. The
culture medium samples were tested with a NO detection kit,
according to the manufacturer's instruction. Absorbance at 550 nm
was measured with a microtiter plate reader (PerkinElmer, Inc.,
Waltham, MA, USA). A range of sodium nitrite concentrations were
used to generate the standard curve. The cellular NO content was
expressed as µmol/g of cellular protein.
NAG content
NAG is a lysosomal enzyme that is present in
proximal tubular cells. The activity of NAG is generally low in
HK-2 cells and increases as a consequence of the breakdown of renal
tubular cells (15). The culture
medium samples were tested for NAG by ELISA (16), according to the manufacturer's
instructions. Absorbance at 450 nm was measured with a microtiter
plate reader, and NAG concentration was determined based on a
standard curve.
LDH leakage assay
LDH is a cytoplasmic oxidoreductase, that is
important in maintaining cell membrane integrity. When cells are
damaged, LDH leaks into the culture medium. LDH leakage was
measured by testing the culture medium samples with an LDH ELISA,
according to the manufacturer's instructions. Absorbance at 450 nm
was measured with a microtiter plate reader and the concentration
of LDH was determined based on a standard curve.
Measurement of ROS production
ROS are oxygenic free radicals which can alter the
balance of endogenous protective systems, such as glutathione and
enzymatic antioxidant defense systems (17). ROS production in cells was detected
by a standard, cell permeable, fluorescent dye method,
2,7-dichlorofluorescin diacetate (DCFH-DA). Intracellular esterases
hydrolyze DCFH-DA to 2,7-dichlorofluorescin (DCFH), which is then
oxidized by ROS to dichlorofluorescein (DCF), and therefore amount
of fluorescence reflects ROS production in cells. Fluorescence
intensity was measured by Accuri™ C6 flow cytometry (BD
Biosciences) with BD CFlow Software v.264.15 (BD Biosciences) at
excitation and emission wavelengths of 485 nm and 530 nm,
respectively (18).
Mitochondrial membrane potential
(MMP)
Flow cytometry following JC-1 staining was used to
evaluate loss of MMP (19,20). HK-2 cells (5×105
cells/well) in 6-well culture plates were treated with 0, 20, 40 or
80 µM POA for 24 h. Cells were trypsinized, washed twice with
ice-cold PBS, resuspended in PBS and labeled with 5 µg/ml JC-1
cationic dye for 30 min in the dark at 37°C. Following labeling,
cells were washed twice with PBS, resuspended in PBS at a
concentration of 1 to 5×106 cells/ml and analyzed by
Accuri™ C6 flow cytometry (BD Biosciences) with BD CFlow Software
v.264.15 (BD Biosciences).
Western blot analysis
HK-2 cells cultured at a density of 1×106
cells/well in 6-well microplates were treated with 0, 20 or 40 µM
POA for 24 h. The treated cells were lysed in RIPA lysis buffer
containing 1% phenylmethane sulfonyl fluoride and incubated on ice
for 30 min. Lysates were centrifuged at 8,000 × g for 8 min
at 4°C, and the protein concentration in the supernatant was
determined by BCA assay. A total of 60 µg/lane protein was
separated by 12% SDS-PAGE and then electrotransferred to a
polyvinylidene fluoride membrane. The membrane was incubated in 5%
skim milk in TBST (0.1% Tween-20) buffer for 2 h, followed by
incubation with the primary antibody (diluted in TBST with 5% skim
milk) overnight at 4°C. The incubated membrane was washed 3 times
with TBST prior to incubation for 1 h at 25°C with the secondary
antibody which was diluted in TBST with 5% skim milk. Following 3
washes with TBST, the immune complexes were detected using an ECL
kit.
Statistical analysis
Data are presented as the mean ± standard deviation
of at least 3 independent experiments. Statistical differences
between means among multiple groups were analyzed with one way
analysis of variance followed by either the Bonferroni or Tamhane's
T2 post hoc test when appropriate. Statistical analysis was
performed using SPSS v.16.0 software (SPSS, Inc., Chicago, IL,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Cytotoxic effect of POA on HK-2
cells
CCK-8 assays were used to determine the effect of
POA on the growth of HK-2 cells. As presented in Fig. 2, cell growth became markedly
inhibited from dose concentrations of 70, 50 and 50 µM POA at 24,
48, or 72 h, respectively. Increased cytotoxicity was observed as
time and POA concentration increased (Fig. 2). DMSO, which was the vehicle
control for the treatments, did not exhibit any inhibitory effect
on the cell viability (0 µM POA; Fig.
2).
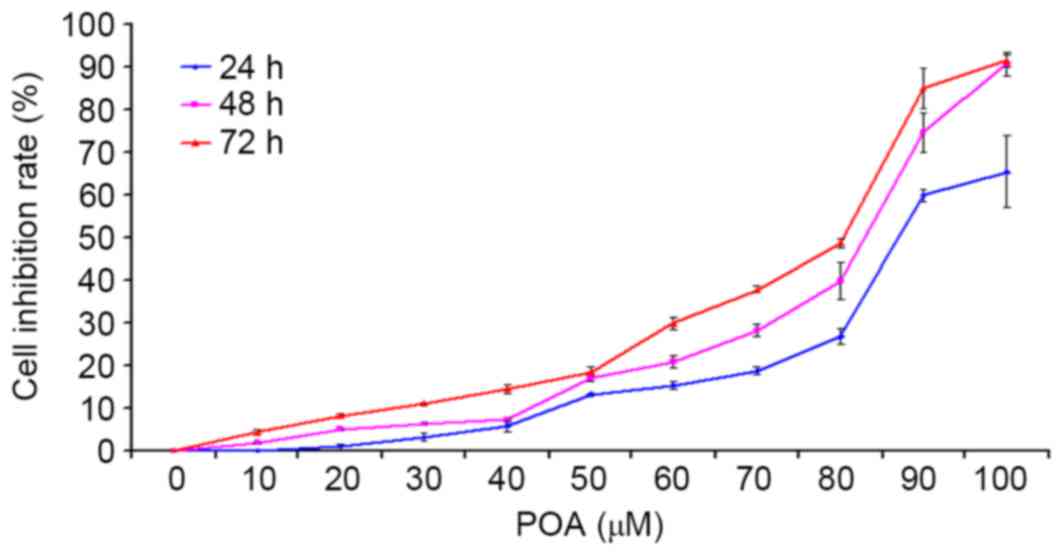 | Figure 2.Effect of POA on HK-2 cell viability.
HK-2 cells were treated with 0, 10, 20, 30, 40, 50, 60, 70, 80, 90,
or 100 µM POA for 24, 48 or 72 h, and cell viability was measured
by the Cell Counting Kit-8 assay. Data are presented as the mean ±
standard deviation of 3 independent experiments. POA, oxalicumone
A. |
Effect of POA on morphological changes
in HK-2 cells and their nuclei
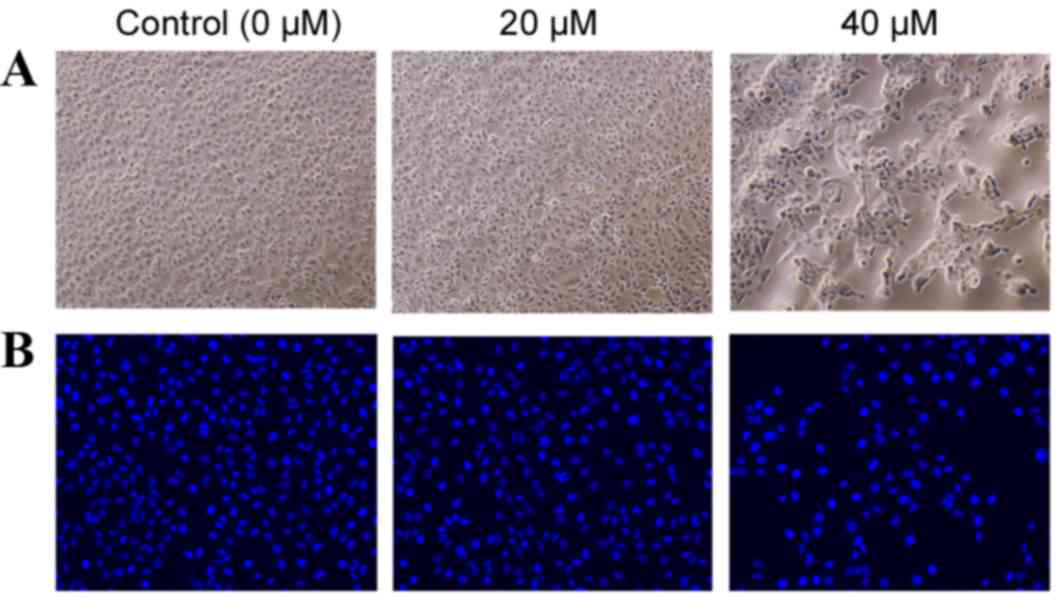
Using a phase contrast microscope, the untreated
HK-2 cells appeared to assume a common spindle cell shape with
intact nuclei (Fig. 3A). In the
presence of 20 µM POA for 24 h, there were slight morphological
alterations when compared with the untreated HK-2 cells, however,
in the presence of 40 µM POA for 24 h, the HK-2 cells contracted
and became rounded and detached from the substrate (Fig. 3A). HK-2 cells were stained with the
nuclear dye Hoechst 33258, which also serves as marker of
apoptosis. Control nuclei did not exhibit distinct changes
(Fig. 3B). In the presence of 40
µM POA, the cells' nuclei revealed brighter Hoechst 33258 staining,
karyopyknosis and fragmented chromatin, indicating that they might
be undergoing apoptosis. By contrast, there were no obvious
alterations in the nuclei with the 20 µM POA treatment (Fig. 3B).
Effect of POA on HK-2 cell
apoptosis
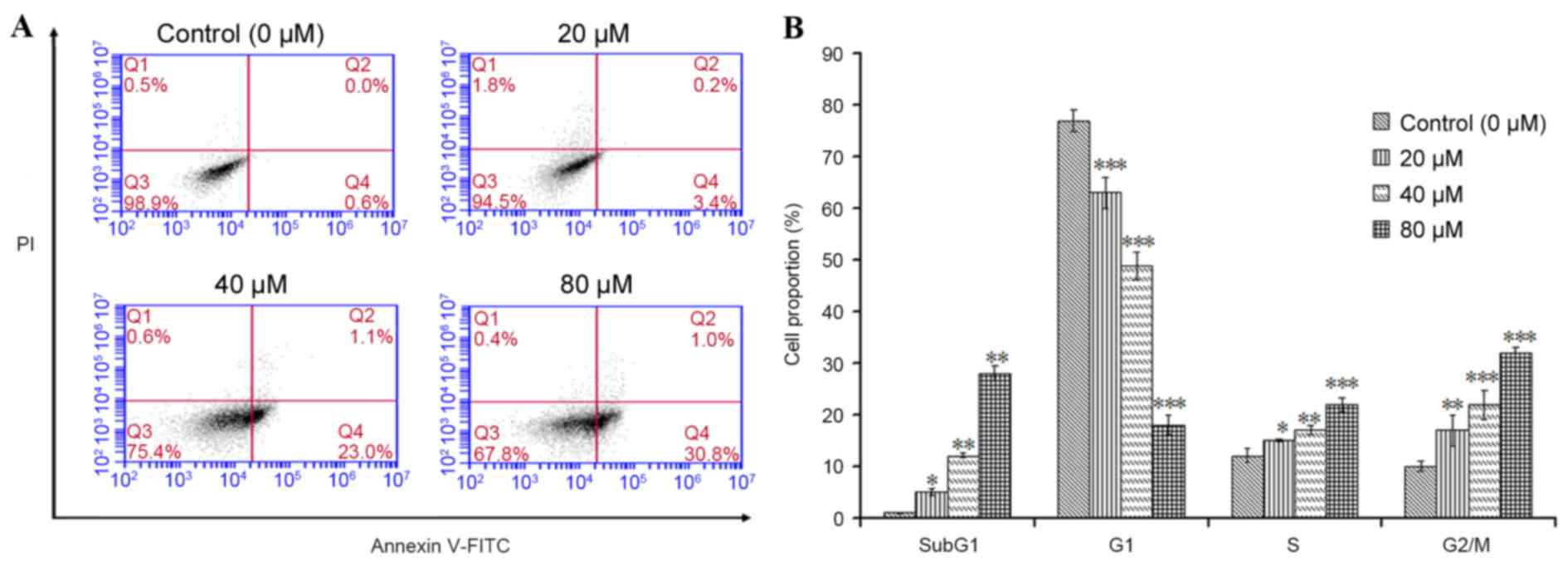
The apoptotic effect of POA on HK-2 cells was
further investigated by flow cytometric analysis. Annexin V-FITC/PI
double-labeling was used for the detection of phosphatidylserine
(PS) externalization, a characteristic of early apoptosis (21,22).
As demonstrated in Fig. 4A, a
significant increase in early apoptotic cells was observed in cells
treated with 20, 40 or 80 µM POA for 24 h (P<0.05), when
compared with untreated cells (Annexin V+/PI−
cells, as measured in the Q4 quadrant of the plots; Fig. 4A). To gain insights into the
mechanism of the growth inhibition activity of POA in HK-2 cells,
its effect on cell cycle distribution was examined by flow
cytometry. The number of apoptotic cells in each treatment group
was determined by observing the SubG1 apoptotic peaks, which are
attributed to the reduced DNA content. As demonstrated in Fig. 4B, treatment with POA for 24 h
resulted in a significant, dose-dependent, increase in the number
of apoptotic cells and a significant increase in the proportion of
cells in the S and G2/M phases compared with untreated cells. By
contrast, a significant reduction of the proportion of cells in the
G1 phase was observed in POA-treated cells compared with untreated
cells (Fig. 4B). In conclusion,
the results from the Annexin V-FITC/PI double-labeling assay and
the cell-cycle analysis indicated that POA-mediated growth
inhibition was accompanied by a dose-dependent increase in
apoptosis.
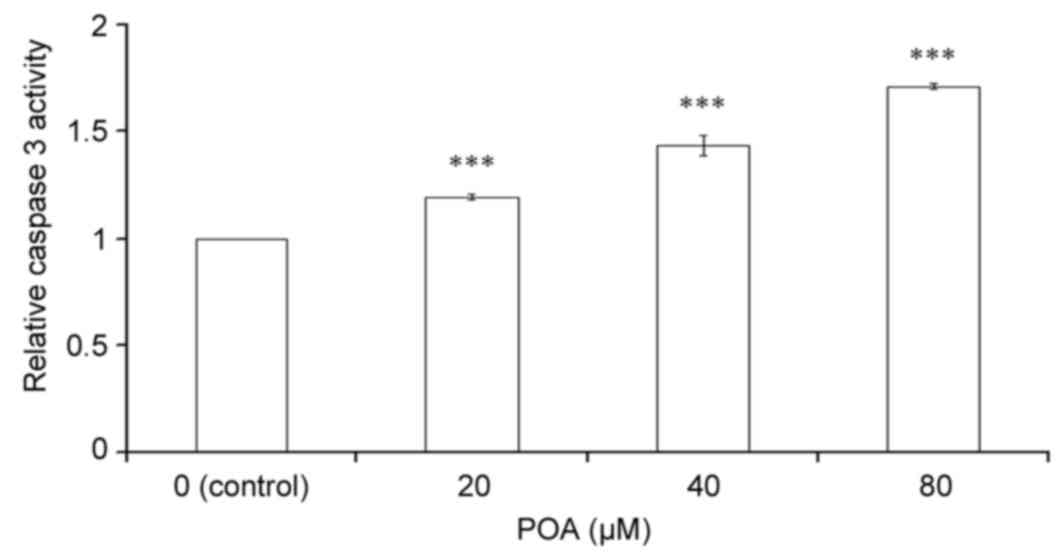
Effect of POA on caspase 3
activity
Caspase 3 is a key regulator of the terminal phase
of apoptosis (23). As
demonstrated in Fig. 5, a
significant, dose-dependent increase in caspase 3 activity was
observed in HK-2 cells treated with 20, 40 or 80 µM POA for 24 h
when compared with control untreated cells (P<0.001).
GSH content
The content of the crucial, cellular, non-enzymatic
antioxidant GSH was determined by ELISA. Treatment with 40 and 80
µM POA induced a significant decrease in cellular GSH content
(Fig. 6A). The level of GSH in the
control cells was 3.85 µg/ml, and this was reduced to 3.71
(P>0.05 vs. control), 3.19 (P<0.01 vs. control) and 2.81
(P<0.01 vs. control) µg/ml with 20, 40 and 80 µM POA treatment,
respectively (Fig. 6A).
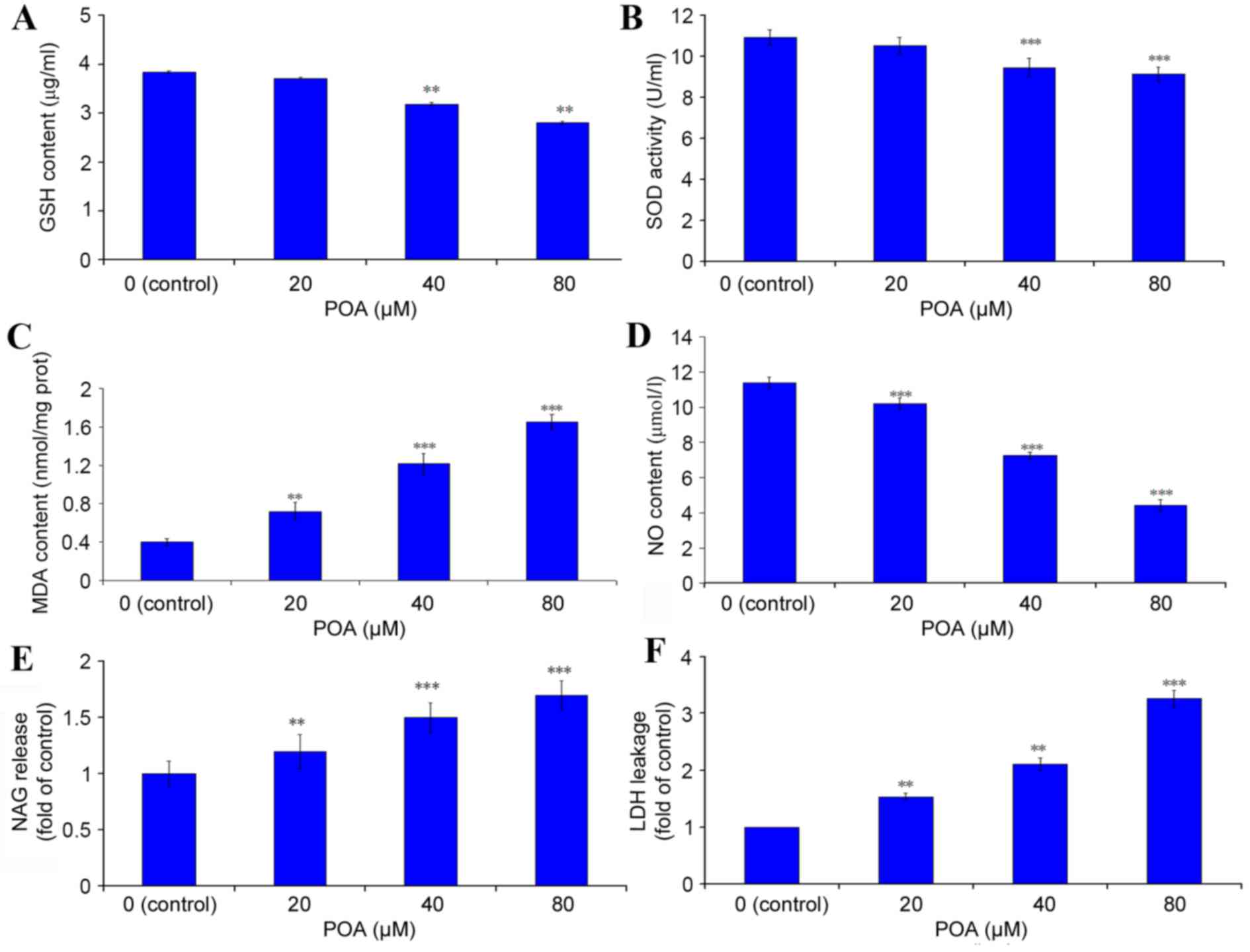 | Figure 6.Effect of POA on the antioxidant
status of HK-2 cells. HK-2 cells were treated with 0, 20, 40 or 80
µM POA for 24 h and the cell lysates (for endogenous molecules
assessment) or the culture medium (for assessment of molecules
released extracellularly) were tested by ELISA or specific dye
kits. (A) GSH content. (B) SOD activity. (C) MDA content. (D) NO
content. (E) NAG content. (F) LDH release. Data are presented as
the mean ± standard deviation of 3 independent experiments.
*P<0.05, **P<0.01 and ***P<0.001 vs. control. POA,
oxalicumone A; GSH, glutathione; SOD, superoxide dismutase; MDA,
malondialdehyde; NO, nitric oxide; NAG,
N-acetyl-β-D-Glucosaminidase; LDH, lactate dehydrogenase. |
SOD activity
HK-2 cells treated with 40 or 80 µM POA demonstrated
a significant decrease in SOD activity compared with control
(Fig. 6B). The SOD activity of the
control cells was 10.93 U/ml, which was reduced to 10.55 (P>0.05
vs. control), 9.47 (P<0.001 vs. control) and 9.12 (P<0.001
vs. control) U/ml in cells treated with 20, 40 and 80 µM POA,
respectively (Fig. 6B).
MDA content
MDA is a well-studied intermediate of oxidative
stress (24). HK-2 cells treated
with 20, 40 and 80 µM POA exhibited a significant increase in MDA
levels (Fig. 6C). MDA levels in
control cells were 0.40 nmol/mg of total protein, and these levels
were increased to 0.72 (P<0.01 vs. control), 1.22 (P<0.001
vs. control) and 1.65 nmol/mg (P<0.001 vs. control) with 20, 40
and 80 µM POA, respectively (Fig.
6C).
NO release
A significant effect of POA was observed on NO
production in HK-2 cells: In control cells, the concentration of NO
released in the culture medium was 11.42 µmol/l, and this was
significantly decreased to 10.20, 7.25 and 4.42 µmol/l with 20, 40
and 80 µM POA treatment, respectively (P<0.001 vs. control;
Fig. 6D).
NAG release
NAG is commonly regarded as a marker of proximal
tubular cell integrity, and measurement of its release is used as
an indicator of early kidney cells injury (25). As demonstrated in Fig. 6E, the NAG release level in HK-2
cells was significantly increased to 1.19-(P<0.01 vs. control),
1.40-(P<0.001 vs. control) and 1.70-fold (P<0.001 vs.
control) relative to the control cells, with 20, 40 and 80 µM POA,
respectively.
LDH leakage
Cellular LDH leakage in the culture medium was
measured as a key signal of damaged cells. LDH activity was
significantly increased to 1.48-(P<0.01 vs. control),
2.02-(P<0.01 vs. control) and 3.15-fold (P<0.001 vs. control)
relative to control cells with 20, 40 and 80 µM POA, respectively
(Fig. 6F).
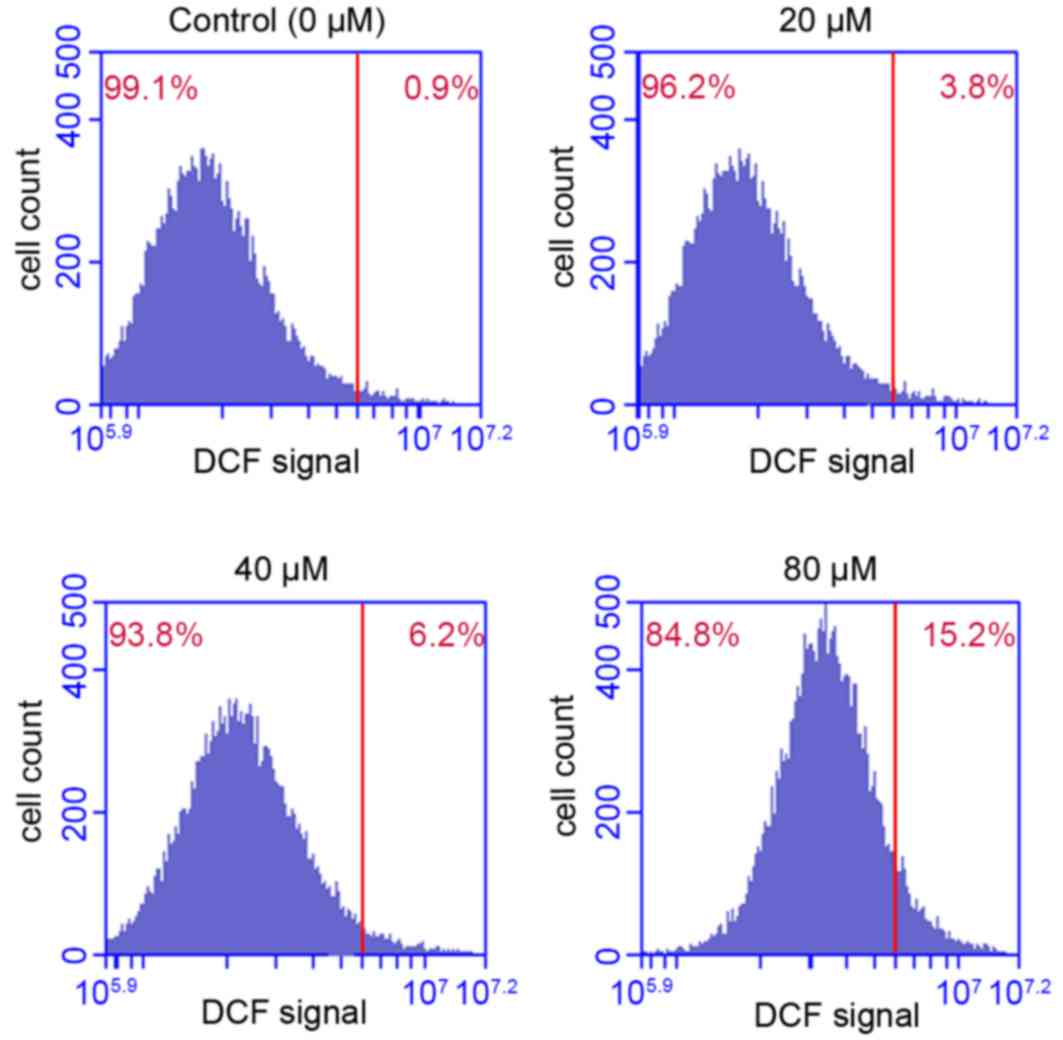
Effect of POA on ROS production
ROS is a critical regulator of cellular homeostasis,
and overproduction of ROS induces apoptosis and cell death
(26). The results presented in
Fig. 7 indicated that POA
treatment significantly promoted ROS accumulation in HK-2 cells, in
a dose-dependent manner. Treatment of HK-2 cells with 20, 40 or 80
µM POA for 24 h increased the % of cells that exhibit ROS
accumulation to 3.8, 6.2 and 15.2% of total, respectively, compared
with 0.9% of total in the control (Fig. 7).
Effect of POA on MMP
Disruption of mitochondrial integrity is one of the
early events leading to apoptosis (27). Accumulation of the cationic dye
JC-1 was assessed by flow cytometry to evaluate the effect of POA
on MMP. Following exposure of HK-2 cells to 20, 40 or 80 µM POA for
24 h, MMP disruption was detected in 14.8, 16.2 and 26.5% of total
cells, respectively, compared with 12.3% of total cells in the
untreated control group (Fig.
8).
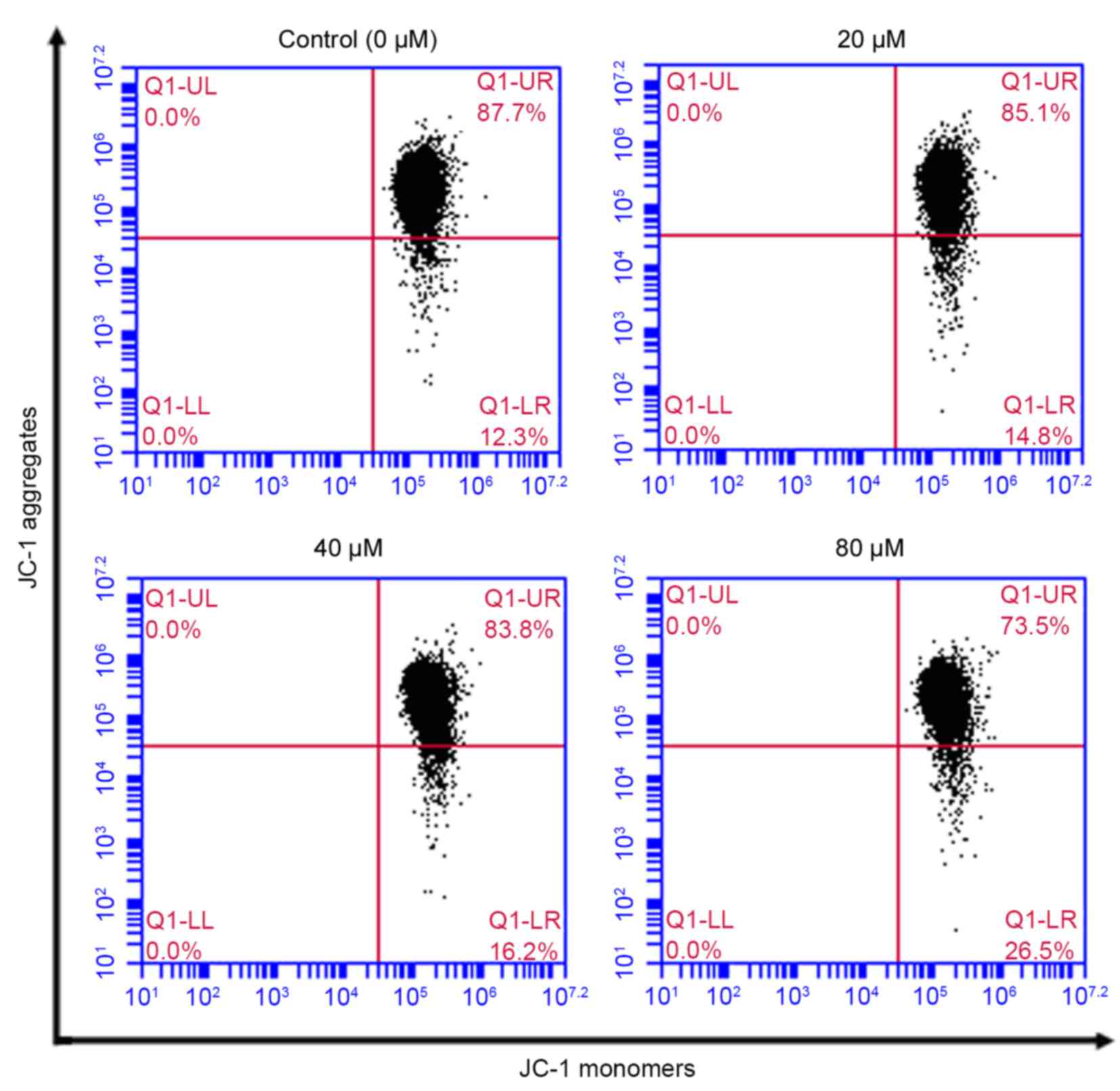 | Figure 8.Effect of POA on MMP depolarization
in HK-2 cells. Cells were treated with 0, 20, 40 or 80 µM POA for
24 h, and MMP depolarization was measured by JC-1 staining and flow
cytometry analysis. Data are presented as the mean ± standard
deviation of 3 independent experiments. POA, oxalicumone A; MMP,
mitochondrial membrane potential; JC-1,
5,5,6,6-tetra-chloro-1,1,3,3-tetraethylbenzimidazolyl- carbocyanine
iodide. |
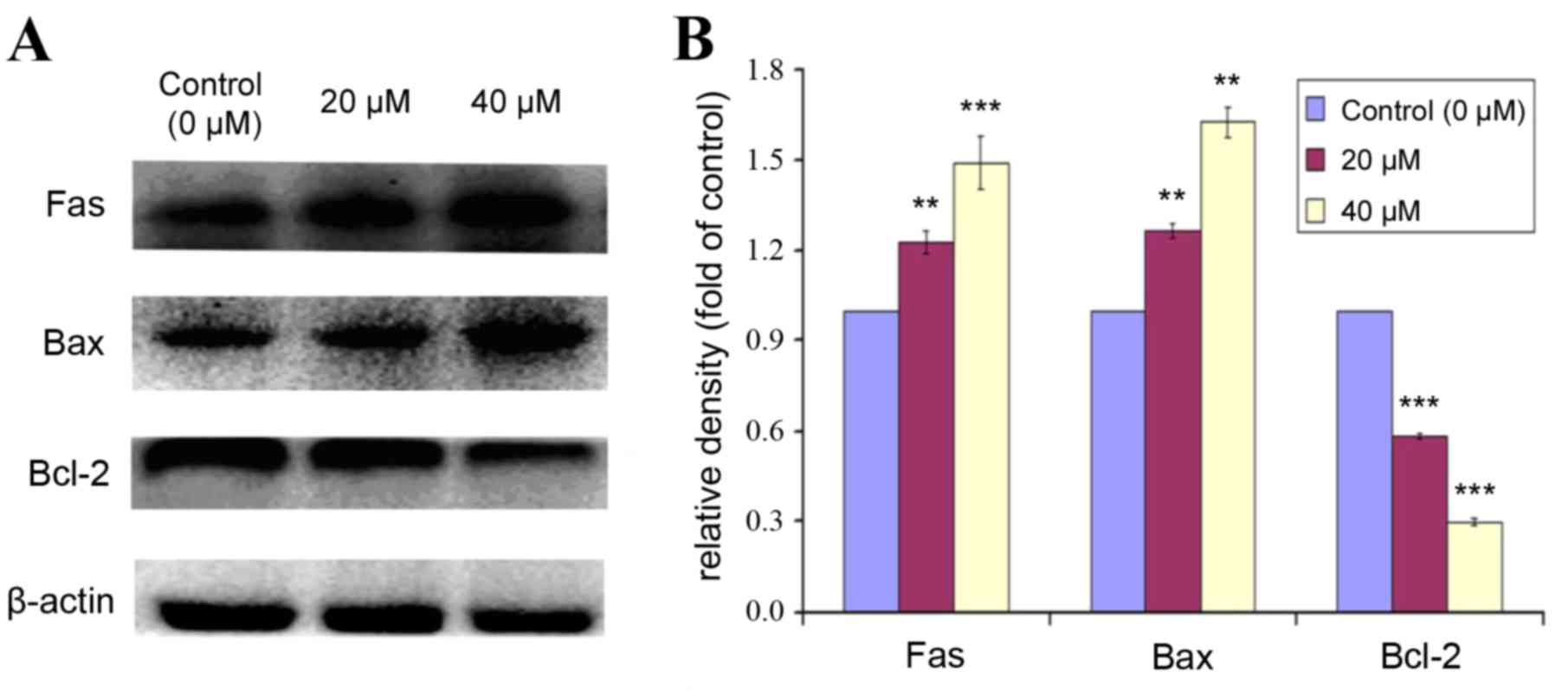
Effect of POA on expression of
apoptotic markers
Bcl-2 is a Bcl-2 family anti-apoptotic protein,
which helps cells to prevent apoptosis. Bax is a Bcl-2 family
pro-apoptotic protein, that translocates from the internal to the
outer mitochondrial membrane, inducing the release of pro-apoptotic
factors, and resulting in apoptosis. The expression levels of Fas,
Bax and Bcl-2 were analyzed by western blot in HK-2 cells treated
with 0, 20 or 40 µM POA for 24 h (Fig.
9). POA treatment significantly increased protein expression
levels of Fas and Bax compared with control (Fig. 9). By contrast, the expression of
Bcl-2 was significantly decreased with POA treatment, compared with
control (Fig. 9). These results
indicated that POA treatment resulted in an upregulation of the
pro-apoptotic proteins Fas and Bax and a downregulation of the
anti-apoptotic protein Bcl-2.
Discussion
Previous studies have demonstrated that POA exhibits
significant cytotoxicity against several carcinoma cell lines with
IC50≤10 µM (3,5) however, the pharmacological mechanism
remains unknown. The present study was designed to assess for the
first time the toxic effect of POA on proximal tubular cells. Using
a cell viability assay, it was demonstrated that POA treatment
inhibited proliferation of HK-2 cells in a dose-dependent manner
(Fig. 2), suggesting that POA
treatment can induce cytotoxicity in human renal epithelial
cells.
Apoptosis, a highly structured and ordered process,
is a fundamental form of cell death, and its purpose is to
eliminate superfluous, harmful, and metabolically perturbed cells
(28,29). In the present in vitro
study, cell nuclei stained by Hoechst 33258 displayed
karyopyknosis, deepened staining and karyorrhexis following POA
treatment, which is consistent with cellular shrinkage, chromatin
condensation, and nuclear fragmentation (Fig. 3). These morphological
characteristics are typical of apoptosis. Apoptosis was further
confirmed by flow cytometry experiments, where POA was demonstrated
to significantly increase the proportion of cells undergoing early
apoptosis in a dose-dependent manner, compared with control
(Fig. 4A). In addition, cell cycle
analysis revealed that POA treatment resulted in an increase of the
sub-G1 cell population, which is a hallmark of apoptosis (30), as well as an increase in the
S-phase cell population, compared with control (Fig. 4B). These results indicate that POA
treatment accelerated the cell cycle, caused abnormal proliferation
and induced apoptosis in the HK-2 cells. Furthermore, an increase
in POA-induced caspase 3 activity was demonstrated (Fig. 5), suggesting that the mechanism of
POA-mediated apoptosis on HK-2 cells was associated with the
activation of this key executor of apoptosis.
To examine whether the toxicity effect of POA on
HK-2 cells involves the mitochondria, the expression levels of
proteins that regulate apoptosis via the Fas, mitochondrial, and
endoplasmic reticulum signaling pathways were analyzed. It has been
reported that Fas is important in the initiation of the cell death
signaling pathway (31). As
demonstrated in Fig. 9, POA
treatment increased the expression of Fas, which suggests that
POA-induced apoptosis in HK-2 may be regulated via the
mitochondria-mediated intrinsic apoptotic pathway.
It is well-established that the mitochondrial death
pathway is regulated by members of the Bcl-2 family. This protein
family can be divided into either anti-apoptotic [such as Bcl-2 and
Bcl-2-like 1 protein extra-large (Bcl-XL)] or pro-apoptotic [such
as Bax, BH3 interacting domain death agonist (Bid) and Bcl-2-like
protein 11 (Bim)] members (32,33).
The Bax protein translocates to the outer membrane of mitochondria,
induces the release of pro-apoptotic factors and induces apoptosis
(34), while the Bcl-2 proteins
sequester in mitochondria, inhibit the release of pro-apoptotic
proteins and factors from liposomes, and suppress apoptosis
(35). The ratio of Bax/Bcl-2
proteins thus determines whether the cell will survive or undergo
apoptosis (36). In the present
study, POA treatment induced expression of the pro-apoptotic
protein Bax and reduced expression of the anti-apoptotic protein
Bcl-2 (Fig. 9). This results in a
significant increase of the Bax/Bcl-2 ratio, which indicates that
POA alters the balance between anti- and pro-apoptotic protein
members in the cell, and eventually induces apoptosis via the Bcl-2
mitochondrial signaling pathway.
Since POA treatment induced apoptosis, its effect on
ROS production was analyzed further. Excessive generation of ROS
compromises cellular function and integrity (37). As demonstrated in Fig. 7, POA treatment was demonstrated to
induce a significant and dose-dependent increase in ROS production
compared with control, which is in agreement with POA inducing
apoptosis and cytotoxicity. POA treatment also induced disruption
of MMP compared with control cells (Fig. 8). It has been reported that
alterations in the mitochondrial functions through increased ROS
generation and the disruption of MMP can lead to cell death
(38). Thus, the increased ROS
generation and the depolarization of MMP observed following POA
treatment further confirms its pro-apoptotic role via the
mitochondrial pathway.
Cellular redox balance is maintained by various
enzymatic and nonenzymatic antioxidant systems. The disruption of
this balance by exogenous substances results in cell damage
(39). The antioxidant systems
include antioxidant enzymes, including SOD and aldo-keto reductase,
and nonenzymatic molecules, including GSH, carotenoid and coenzyme
(40). The main physiological
function of GSH is scavenging free radicals and antioxidants
(41). In the present study, POA
treatment decreased the content of GSH in cells, compared with
control (Fig. 6A). SOD is a common
component of the cellular antioxidant systems, blocking cell damage
resulting from oxygen free radicals, and promptly repairing the
damaged cells. In the present study, POA treatment significantly
decreased SOD activity (Fig. 6B),
which is in agreement with the effect of POA on GSH. These results
further confirmed that POA induced oxidative stress on HK-2
cells.
It has been reported that membrane lipids are major
targets of free radicals (42). An
increase in the levels of lipid peroxidation products, such as MDA,
is an indication of membrane lipid damage (43). As demonstrated in Fig. 6C, POA treatment elevated MDA levels
compared with control, which suggests that POA induces lipid
peroxidation injury. These results confirm a POA-mediated oxidative
stress induction, consistent with the results from the SOD and GSH
measurements.
NO is a free radical that is a key participator in
both physiological and pathological processes. Similar to other
free radical species, small amounts of NO in the cell exhibit a
protective effect, while excessive amounts of NO induce cell damage
(44). LDH is a cytoplasmic,
glycolytic enzyme, that is highly expressed in kidney cells. When
cells are damaged, LDH leaks from the cell cytoplasm to the
extracellular space (45). The
present study revealed that NO and LDH levels were increased
following POA treatment compared with control, which indicated that
POA induced damage in HK-2 cells (Fig.
6D and F). NAG is generally considered a reliable and sensitive
enzyme marker of tubular epithelia injury (46,47).
POA treatment resulted in increased NAG release (Fig. 6E), which further confirmed the
cytotoxicity effect of POA on HK-2 cells.
In conclusion, using a wide variety of experimental
assays, the present study demonstrated that POA significantly
induced cytotoxicity and apoptosis in human renal epithelial cells
via the mitochondrial pathway. Analysis of antioxidant systems
activity indicated that POA induced damage to cellular antioxidant
enzymes, and changed the cellular antioxidant balance towards cell
toxicity. It remains unclear whether the toxicological mechanism
identified in the present study is the same as the pharmacological
mechanism, and this needs to be further investigated. The present
study evaluated for the first time (to the best of our knowledge)
POA-mediated toxicity and its mechanism in vitro, which is
valuable information for novel drug discovery. HK-2 cells are
suitable for toxicity studies in vitro, because the proximal
tubule cells of the kidney are the most common site of injury by
nephrotoxic drugs. Eventually, the present study may contribute
towards the potential use of POA in clinical application.
Acknowledgements
This work was financed by the National Marine Public
Welfare Research Project of China (grant no. 201305017), National
Natural Science Foundation of China (grant no. 81573638) and Xinhuo
Planning Project of Guangzhou University of Chinese Medicine (grant
no. XH20150107).
References
|
1
|
Blunt JW, Copp BR, Keyzers RA, Munro MH
and Prinsep MR: Marine natural products. Nat Prod Rep. 29:144–222.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Qi SH: Bioactive compounds from marine
gorgonian coralsStudies In Natural Products Chemistry. Rahman A:
38. 1st. Elsevier; Oxford: pp. 325–352. 2012, View Article : Google Scholar
|
|
3
|
Zhang XY, Bao J, Wang GH, He F, Xu XY and
Qi SH: Diversity and antimicrobial activity of culturable fungi
isolated from six species of the South China Sea gorgonians. Microb
Ecol. 64:617–627. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sun YL, He F, Liu KS, Zhang XY, Bao J,
Wang YF, Nong HX, Xu XY and Qi SH: Cytotoxic
dihydrothiophene-condensed chromones from marine-derived fungus
Penicillium oxalicum. Planta Med. 78:1957–1961. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sun YL, Bao J, Liu KS, Zhang XY, He F,
Wang YF, Nong XH and Qi SH: Cytotoxic dihydrothiophene-condensed
chromones from the marine-derived fungus Penicillium oxalicum.
Planta Med. 79:1474–1479. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu Y, Connors D, Barber L, Jayachandra S,
Hanumegowda UM and Adams SP: Multiplexed assay panel of
cytotoxicity in HK-2 cells for detection of renal proximal tubule
injury potential of compounds. Toxicol In Vitro. 23:1170–1178.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang J, Jia L, Kuang Z, Wu T, Hong Y, Chen
X, Leung WK, Xia J and Cheng B: The in vitro and in vivo antitumor
effects of clotrimazole on oral squamous cell carcinoma. PLoS One.
9:e988852014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tripathi M, Singh BK, Mishra C, Raisuddin
S and Kakkar P: Involvement of mitochondria mediated pathways in
hepatoprotection conferred by Fumaria parviflora Lam. extract
against nimesulide induced apoptosis in vitro. Toxicol In Vitro.
24:495–508. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nigam N, George J, Srivastava S, Roy P,
Bhui K, Singh M and Shukla Y: Induction of apoptosis by
(6)-gingerol associated with the modulation of p53 and involvement
of mitochondrial signaling pathway in B(a)P-induced mouse skin
tumorigenesis. Cancer Chemother Pharmacol. 65:687–696. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Terada Y, Inoshita S, Hanada S, Shimamura
H, Kuwahara M, Ogawa W, Kasuga M, Sasaki S and Marumo F:
Hyperosmolality activates Akt and regulates apoptosis in renal
tubular cells. Kidney Int. 60:553–567. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang JQ, Shen M, Zhu CC, Yu FX, Liu ZQ,
Ally N, Sun SC, Li K and Liu HL: 3-Nitropropionic acid induces
ovarian oxidative stress and impairs follicle in mouse. PLoS One.
9:e865892014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lund AK, Knuckles TL, Akata Obot C, Shohet
R, McDonald JD, Gigliotti A, Seagrave JC and Campen MJ: Gasoline
exhaust emissions induce vascular remodeling pathways involved in
atherosclerosis. Toxicol Sci. 95:485–494. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu CP, Hu W, Wu H and Hu X: No evident
dose-response relationship between cellular ROS level and its
cytotoxicity-a paradoxical issue in ROS-based cancer therapy. Sci
Rep. 4:50292014.PubMed/NCBI
|
|
14
|
Yuste JE, Tarragon E, Campuzano CM and
RosBernal F: Implications of glial nitric oxide in
neurodegenerative diseases. Front Cell Neurosci. 9:3222015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chiu JS: Models used to assess renal
function. Drug Dev Res. 32:247–255. 1994. View Article : Google Scholar
|
|
16
|
Ali RJ, Al-Obaidi FH and Arif HS: The role
of urinary N-acetyl Beta-D-glucosaminidase in children with
urological problems. Oman Med J. 29:285–288. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Prasad NR, Menon VP, Vasudev V and
Pugalendi KV: Radioprotective effect of sesamol on gamma-radiation
induced DNA damage, lipid peroxidation and antioxidants levels in
cultured human lymphocytes. Toxicology. 209:225–235. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sharikabad MN, Ostbye KM, Lyberg T and
Brørs O: Effect of extracellular Mg(2+) on ROS and Ca(2+)
accumulation during reoxygenation of rat cardiomyocytes. Am J
Physiol Heart Circ Physiol. 280:H344–H353. 2001.PubMed/NCBI
|
|
19
|
Cossarizza A, Baccarani-Contri M,
Kalashnikova G and Franceschi C: A new method for the
cytofluorimetric analysis of mitochondrial membrane potential using
the J-aggregate forming lipophilic cation
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolcarbocyanine
iodide (JC-1). Biochem Biophys Res Commun. 197:40–45. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yao J, Jiang Z, Duan W, Huang J, Zhang L,
Hu L, He L, Li F, Xiao Y, Shu B and Liu C: Involvement of
mitochondrial pathway in triptolide-induced cytotoxicity in human
normal liver L-02 cells. Biol Pharm Bull. 31:592–597. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fadok VA, Voelker DR, Campbell PA, Cohen
JJ, Bratton DL and Henson PM: Exposure of phosphatidylserine on the
surface of apoptotic lymphocytes triggers specific recognition and
removal by macrophages. J Immunol. 148:2207–2216. 1992.PubMed/NCBI
|
|
22
|
Zamai L, Falcieri E, Marhefka G and Vitale
M: Supravital exposure to propidium iodide identifies apoptotic
cells in the absence of nucleosomal DNA fragmentation. Cytometry.
23:303–311. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wilson MR: Apoptotic signal transduction:
Emerging pathways. Biochem Cell Biol. 76:573–582. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bonnes-Taourel D, Guérin MC and Torreilles
J: Is malonaldehyde a valuable indicator of lipid peroxidation.
Biochem Pharmacol. 44:985–988. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yamashita T, Doi K, Hamasaki Y, Matsubara
T, Ishii T, Yahagi N, Nangaku M and Noiri E: Evaluation of urinary
tissue inhibitor of metalloproteinase-2 in acute kidney injury: A
prospective observational study. Crit Care. 18:7162014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sinha K, Das J, Pal PB and Sil PC:
Oxidative stress: The mitochondria-dependent
andmitochondria-independent pathways of apoptosis. Arch Toxicol.
87:1157–1180. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ly JD, Grubb DR and Lawen A: The
mitochondrial membrane potential (deltapsi (m)) in apoptosis; an
update. Apoptosis. 8:115–128. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jacobson MD, Weil M and Raff MC:
Programmed cell death in the animal development. Cell. 88:347–354.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Scaffidi C, Kirchhoff S, Krammer PH and
Peter ME: Apoptosis signaling in lymphocytes. Curr Opin Immunol.
11:277–285. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pirocanac EC, Nassirpour R, Yang M, Wang
J, Nardin SR, Gu J, Fang B, Moossa AR, Hoffman RM and Bouvet M:
Bax-induction gene therapy of pancreatic cancer. J Surg Res.
106:346–351. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Waring P and Müllbacher A: Cell death
induced by the Fas/Fas ligand pathway and its role in pathology.
Immunol Cell Biol. 77:312–317. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Certo M, Del Gaizo Moore V, Nishino M, Wei
G, Korsmeyer S, Armstrong SA and Letai A: Mitochondria primed by
death signals determine cellular addiction to antiapoptotic BCL-2
family members. Cancer Cell. 9:351–365. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nakazawa Y, Kamijo T, Koike K and Noda T:
ARF tumor suppressor induces mitochondria-dependent apoptosis by
modulation of mitochondrial Bcl-2 family proteins. J Biol Chem.
278:27888–27895. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Antonsson B, Conti F, Ciavatta A,
Montessuit S, Lewis S, Martinou I, Bernasconi L, Bernard A, Mermod
JJ, Mazzei G, et al: Inhibition of Bax channel- forming activity by
Bcl-2. Science. 277:370–372. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Borner C: The Bcl-2 protein family:
Sensors and checkpoints for life-or-death decisions. Mol Immunol.
39:615–647. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lanave C, Santamaria M and Saccone C:
Comparative genomics: The evolutionary history of the Bcl-2 family.
Gene. 333:71–79. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zorov DB, Juhaszova M and Sollott SJ:
Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS
release. Physiol Rev. 94:909–950. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Urra FA, Cordova-Delgado M, Pessoa-Mahana
H, Ramírez-Rodriguez O, Weiss-Lopez B, Ferreira J and
Araya-Maturana R: Mitochondria: A promising target for anticancer
alkaloids. Curr Top Med Chem. 13:2171–2183. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rosas HD, Doros G, Bhasin S, Thomas B,
Gevorkian S, Malarick K, Matson W and Hersch SM: A systems-level
‘misunderstanding’: The plasma metabolome in Huntington's disease.
Ann Clin Transl Neurol. 2:756–768. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Brzović-Šarić V, Landeka I, Šarić B,
Barberić M, Andrijašević L, Cerovski B, Oršolić N and Đikić D:
Levels of selected oxidative stress markers in the vitreous and
serum of diabetic retinopathy patients. Mol Vis. 21:649–664.
2015.PubMed/NCBI
|
|
41
|
Schmitt B, Vicenzi M, Garrel C and Denis
FM: Effects of N-acetylcysteine, oral glutathione (GSH) and a novel
sublingual form of GSH on oxidative stress markers: A comparative
crossover study. Redox Biol. 6:198–205. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Valko M, Jomova K, Rhodes CJ, Kuca K and
Musílek K: Redox- and non-redox-metal-induced formation of free
radicals and their role in human disease. Arch Toxicol. 90:1–37.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dubner D, Gisone P, Jaitovich I and Perez
M: Free radicals production and estimation of oxidative stress
related to gamma irradiation. Biol Trace Elem Res. 47:265–270.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Förstermann U and Sessa WC: Nitric oxide
synthases: Regulation and function. Eur Heart J. 33:829–837. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xie H, Valera VA, Merino MJ, Amato AM,
Signoretti S, Linehan WM, Sukhatme VP and Seth P: LDH-A Inhibition,
a therapeutic strategy for treatment of hereditary leiomyomatosis
and renal cell cancer (HLRCC). Mol Cancer Ther. 8:626–635. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Laina A and Schwartz D: Renal tubular and
molecular events in acute renal failure. Nephron. 68:413–418.
1994.PubMed/NCBI
|
|
47
|
Jongman RM, van Klarenbosch J, Molema G,
Zijlstra JG, de Vries AJ and van Meurs M: Angiopoietin/Tie2
dysbalance is associated with acute kidney injury after cardiac
surgery assisted by cardiopulmonary bypass. PLoS One.
10:e01362052015. View Article : Google Scholar : PubMed/NCBI
|