Chloride channels (ClCs) are a type of permeable
channel protein for chloride ions or other anions on the cell
membrane, and ClC proteins are encoded by genes of the ClC family.
ClCs have nine family members, which are classified into three
distinct subfamilies: ClC-1, ClC-2, ClC-Ka/K1 and ClC-Kb/K2; ClC-3,
ClC-4 and ClC-5; and ClC-6 and ClC-7 (1). ClC-2 is one of the nine mammalian
members of the ClC family, and was initially isolated from the rat
heart and brain (2,3) and then from the rabbit heart
(4,5). ClC-2 is a two-pore homodimeric,
voltage-gated Cl− channel (5–8).
ClC-2 can be activated by hyperpolarization (3,9),
cell swelling (2,9), extracelluar hypotonicity (2) and extracellular acidification
(9,10). ClC-2 is almost ubiquitously
expressed (3,11), including in ureteric bud cells
(12), intestine (13–17),
gastric parietal cells (18,19),
the liver (20), lung (11,21–26),
rat retina (27), parotid acinar
cells (28), guinea pig cardiac
muscle (29), neuronal cells
(30), rat and human airways
(17), bovine trabecular meshwork
(31), human trabecular meshwork
(32,33) and rat trabecular meshwork (34). In addition, ClC-2 can regulate cell
volume (2,4,35),
control response to swelling (11,36–39),
and regulates post-synaptic responses to GABA and glycine (11,40,41).
Although ClC-2 has a wide variety of properties and
functions, repots of CLC-3 prior to 1994 are limited, with only six
published between 1953 and 1994. Between 1994 and 2015, ClC-2
attracted increasing attention as ClC-2 was understood to possess
several molecular, functional and physiological properties, and be
associated with several diseases, including degeneration of the
retina, Sjögren's syndrome, age-related cataracts, degeneration of
the testes, azoospermia, lung cancer, constipation, repair of
impaired intestinal mucosa barrier, leukemia, cystic fibrosis,
leukoencephalopathy, epilepsy and diabetes mellitus. However,
reviews on ClC-2 are limited. The present review aimed to discuss
the molecular, functional and physiological properties of ClC-2, in
addition to mechanisms involved in the regulation of ClC-2 and
diseases associated with ClC-2.
The ClC-2 protein is encoded by the CLCN2 gene,
which is composed of 898 amino acids and is located in chromosome
3q27.1. The major transcription start site of the ClC-2 gene has
been identified, and is localized 100 bp upstream of the putative
translation initiation codon (21). A previous study (42) on the possible evolution of ClC-2
gene suggested that ClC-2 may have evolved by gene duplication,
mutation and DNA rearrangement (27,43).
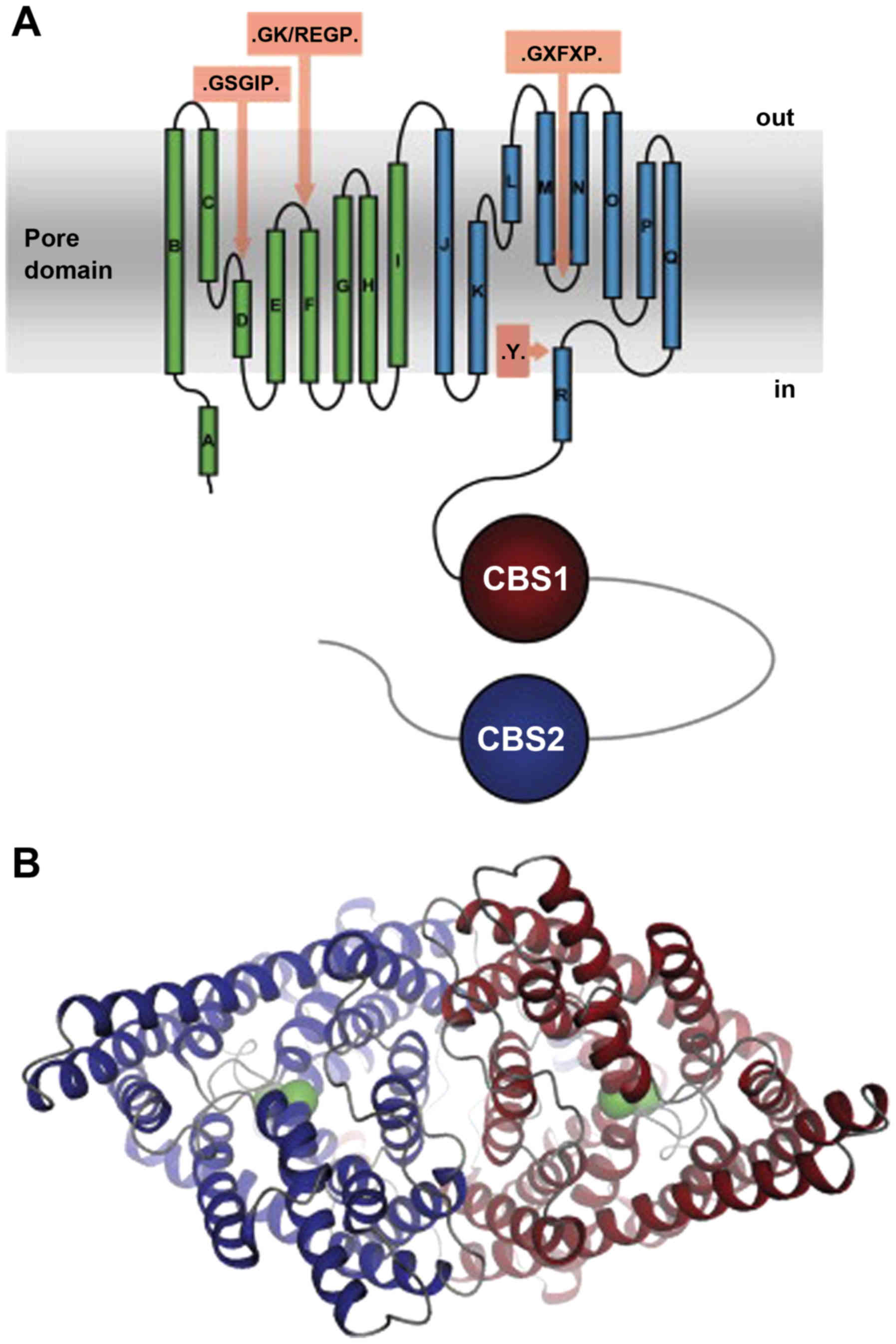
Despite wide functional diversity, ClC family
members share a conserved protein structure, including a
transmembrane region, which is involved in chloride anion
transport, and two intracellular copies of the
cystathionine-β-synthase (CBS) domain (CBS1 and CBS2) (5–8,45,46)
and an N-terminus. Therefore, the protein structure of ClC-2 is the
same in the other ClC family members (45,46),
as shown in Fig. 1.
Several other studies on the structural properties
of ClC-2 have reported that the ClC-2 dimer is the minimum
functional structure (47),
protons act independently from the possible effects of the
N-terminus on gating (48) and the
hetero-dimerization of ClC-2 can modify the unitary conductance of
protopores (49).
Previous studies have shown that ClC-2 has several
functional properties, including effects on cell volume (2,4,20,35,61),
the digestive system (11,16,18,19,62–64),
the eye (65–67), respiratory system (11,24,26,68–70),
circulatory system (35,71–75)
and nervous system (76). In
addition, several studies have reported the functions of the NH2
terminus (77) and carboxy
terminus (78) in ClC-2, and the
similar functional properties between chloride-selective ion
channels, including ClC-2, in protozoa (79–82).
The activation of ClC-2 in response to cell swelling
and low extracellular pH is involved in the dissociation of an
NH2-terminal region of ClC-2 (10). The dephosphorylation of
serine/threonine residues in the channel or an associated protein
appears to be necessary for current activation following cell
swelling in T84 cells (53) and
for a ClC-2-like current in ascidian embryo cells during the cell
cycle (83). Hypotonic cell
swelling activates an outwardly rectifying anionic conductance,
with different characteristics from those of ClC-2 previously
reported (84). The endogenous
currents suggest that protein tyrosine phosphorylation is involved
in the signal transduction pathway in certain cell types (85–87),
and that epidermal growth factor enhances the hypotonicity-induced
anionic efflux (85). However,
tyrosine kinase p56lck activates these
swelling-sensitive chloride channels without cell swelling, even
during cell shrinkage, in lymphocytes (88).
Previous studies have shown that CIC-2 has a
protective effect on trabecular cells under pressure stress
(65), can regulate the inward
rectification in Drosophila retinal photoreceptors (66), and can modulate cellular volume,
intracellular Cl− and other cellular functions in
trabecular meshwork cells (67).
ClC-2 is gestationally regulated and is
predominantly expressed in the fetal lung (26,68).
Cystic fibrosis transmembrane conductance regulator
(CFTR)-independent fluid accumulation is induced by keratinocyte
growth factor in fetal lung explants (69), and keratinocyte growth factor is
necessary for ClC-2-like pH-sensitive Cl− secretion in
the fetal airway epithelia (24).
It has been shown that ClC-2 affects fetal lung fluid production
and lung cyst morphology (70).
Estrogen is associated with a decrease in the
urinary excretion of sodium and chloride (150–153). A study by Nascimento et al
(90) showed that ClC-2 may be
involved in estrogen-induced Cl− transport in the rat
kidney. Another study (91) showed
that ClC-2 is also involved in the rat renal tubule transcellular
chloride transport regulated by arginine vasopressin, a
neurohypophysial hormone.
The functional expression and activation of ClC-2
is reduced by dynein (a protein complex) (101) and p34cdc2/cyclin B
(102), however, the inrerferon-γ
glycoprotein can activate ClC-2 in lung epithelial cells through
mRNA stabilization, and increase ClC-2 transcripts in Calu-3 cells
(107). ClC-2 is regulated by
protein phosphatase 1 (102),
interactions between the actin cytoskeleton (a filamentous protein
structure) and the N-terminus of ClC-2 (104). M phase-specific
p34cdc2/cyclin B can phosphorylate the ubiquitination of
ClC-2 (103). Heat shock protein
90 can increase ClC-2 current amplitude and the intracellular
Cl− concentration, and enhance channel sensitivity to
intracellular Cl− (105). ClC-2 is also associated with the
transmembrane glycoprotein, GlialCAM, in the brain, and GlialCAM
can involve ClC-2 in the homeostasis of myelin, which is defective
in leukodystrophy (106).
ClC-2 membrane abundance is increased by serum and
glucocorticoid inducible kinases (SGKs) and decreased by NEDD4-2,
an enzyme of the NEDD4 family (108). In rats, protein kinase A can
directly phosphorylate ClC-2, whereas protein kinase C and
Ca2+/calmodulin-dependent protein kinase II cannot
(113). PIKfyve is a FYVE
finger-containing phosphoinositide kinase (158) and is a potent stimulator of
ClC-2-activity (112), which can
contribute to the SGK1-dependent regulation of ClC-2 (112).
Janus kinase (JAK) 2 and JAK3 (tyrosine kinase) can
downregulate the activation of ClC-2 and offset Cl− exit
(109,111), however, their functions and
regulatory mechanism are different. JAK2 is involved in the
signaling of leptin (159),
growth hormones (160),
erythropoietin, thrombopoietin, granulocyte colony-stimulating
factor (161) and a variety of
cytokines (161,162). JAK2 inhibitors can treat
myeloproliferative disorders (163–170), and JAK2 can be activated by
hyperosmotic shock (171,172). By contrast, JAK3 can promote
proliferation and act against the apoptosis of lymphocytes and
tumor cells (173–177). It is also present in acute
megakaryoplastic leukemia (178,179). Therefore, cell proliferation and
apoptosis is associated with Cl− channel activity
(110). In addition, the
substitution of lysine by alanine can inactivate JAK3 (180). In addition to JAK2 and JAK3,
SPS1-related proline/alanine-rich kinase and oxidative
stress-responsive kinase 1 can also downregulate ClC-2 (114).
The Sp1 and Sp3 transcription factors can control
the rate of transcription of genetic information from DNA to
messenger RNA (181,182). Reducing interactions between Sp1
or Sp3 and the ClC-2 promoter can lead to a postnatal decrease in
the expression of ClC-2 in lung epithelia (116). In addition, the glycosylation of
SP1 produces the 105-kD isoform of SP1 and is involved in
regulating the expression of ClC-2 (115).
According to previous studies, in addition to the
above-mentioned regulatory mechanisms, scorpion venom (121), α1-adrenoceptor (122), Plasmodium berghei (123), adenosine triphosphate (124,125), ClH3 (137), GABAAR (40,126–128), permeant anions (138,139), membrane cholesterol (140) and β-cyclodextrin sensitive
clusters (141) can also regulate
or affect ClC-2. A peptide of scorpion venom can induce slower
activation kinetics of ClC-2 (121). α1-adrenoceptor activation can
significantly reduce protein levels of ClC-2 in the villus and
crypt epithelial cells from the acutely denervated jejunum, but not
the innervated jejunum (122).
The activation of ClC-2 is involved in the altered permeability
caused by Plasmodium berghei infection in Plasmodium
berghei-infected mouse CLCN2 (123). ATP not only decelerates ClC-2
common gating for sufficient electrical stability of neurons
(124), it also alters its
surface expression (125).
Intra-/extra-cellular permeant anions can affect the Vm-dependence
of ClC-2 (138). Specially,
Cl− as a permeant anion can increase pore occupancy,
thus obstructing the closure of the protopore gate of ClC-2
(139). In addition, alterations
in the (Cl−)i between 10 and 200 mM can
decelerate ClC-2 channel closing at a positive Vm (139). Membrane cholesterol can regulate
the activation of ClC-2, and the increased activation of ClC-2 is
involved in the relocalization of ClC-2 to detergent-soluble
microdomains (140).
β-cyclodextrin sensitive clusters with other molecules can mantain
ClC-2 activity (141).
Previous studies have shown that disruption and
abnormity of ClC-2 can cause a number of diseases, including
ophthalmological disease (11,183–185), otorhinolaryngological disease
(186), disease of the
reproductive system (11,187), disease of the respiratory system
(188), disease of the digestive
system (189–194), disease of the hematological
system (195), genetic diseases
(23,25,26,42,44,74,76,196–198), diseases of the nervous system
(30,40,41,168,199–212) and metabolic diseases (213). The majority of studies have
focused on diseases of the nervous system.
Severe degeneration of the retina occurs in
ClC-2-knockout mice, although no other notable eye abnormalities
are present (11). Possible
reasons for degeneration of the retina caused by ClC-2-knockout
include the following (11): i)
ClC-2 disruption causes the death of photoreceptor cells; ii)
depletion of ClC-2 may impair the transport and alter the ionic
environment of photoreceptors. Other studies on ophthalmological
diseases associated with ClC-2 include Sjögren's syndrome (183,184,214–218) and age-related cataracts (185).
Sjögren's syndrome is a chronic autoimmune disease,
and previous studies have shown that eye diseases caused by
Sjögren's syndrome are primarily through the destruction of
lacrimal glands (LGs) (214),
keratoconjunctivitis sicca (215). In addition, LG pathologic
properties in Sjögren's syndrome appear in rabbits induced by
autoimmune dacryoadenitis (216–218). Acini and interlobar ducts have
the lowest mRNA abundance of ClC2γ, and the intralobar duct has the
highest (183,184), and alterations in ClC2γ may alter
lacrimal secretion, particularly Cl− transport (184). The causes of age-related
cataracts include aging, inheritance, local nutritional disorders,
immune and metabolic abnormalities, trauma, poisoning and
radiation. According to a study by Ouyang (185), the expression level of ClC-2 is
associated with the development of age-related cataracts.
Reports on otorhinolaryngological diseases
associated with ClC-2 are limited, however, Li et al
reported on the pathogenesis of nasal polyps. The study (186) showed that ClC-2 was not expressed
in normal nasal mucosa, however, ClC-2 proteins were expressed in
epithelial cells and sub-epithelial mucous glands in patients
affected with nasal polyps. This suggested that ClC-2 is involved
in the pathogenesis of nasal polyps.
In previous studies, a number of diseases of the
reproductive system have been associated with ClC-2, including
degeneration of the testes (11)
and azoospermia (187,219,220).
It was shown that severe degeneration of the testes
appears in ClC-2-knockout mice, however, no other reproductive
abnormalities are observed (11).
A study by Bösl et al suggested that ClC-2 disruption causes
the death of germ cells, impaired transport and alterations to the
ionic environment of germ cells, and that germ cells rely on the
transepithelial transport mediated by Sertoli cells (11).
Azoospermia is a medical condition in which males
do not have a measurable level of sperm in their semen, and is
associated with low levels of fertility or infertility. Its
pathogenesis involves CFTR disruption in CF, affecting male
fertility (219). In addition,
the male sterility resulting from reduced fluid volume is
associated with disruption of Cl− channels from other
ClCN genes (220). Another study
(187) by Edwards et al
reported that ClCN2nmf240 homozygotes have azoospermic symptoms
severe degradation of spermatogenesis, and shortages of
spermatocytes, spermoblasts and sperm.
Reports on diseases of the respiratory system
associated with ClC-2 are limited. A previous study (188) reported that ClC-2 may be
important in the invasion, development and occurrence of lung
cancer, and that ClC-2 may be a novel molecular target for clinical
therapy in non-small cell lung cancer.
There have been several reports on diseases of
digestive system associated with ClC-2, including constipation
(18,26,189–193,221) and repair of impaired intestinal
mucosa barrier (194).
CFTR is major chloride channel regulating chloride
secretion in the small intestine (221), however, ClC-2 can also regulate
chloride secretion (18,26). Hypotonicity can also activate
chloride currents in the rat ileum (189). Several studies have reported that
ClC-2 is expressed in the intestinal epithelium, in T84 human
intestinal cells, rat intestinal tissue (3) and the murine duodenum (190). In addition, previous studies have
shown that ClC-2 can regulate chloride secretion in rodent neonatal
airways (26), and regulate
gastric chloride secretion (18).
However, another study (191)
described contradictory findings, reporting that ClC-2 was not
involved in Cl− secretion, but was involved in
Cl− absorption in the distal colon. According to these
studies, ClC-2 inhibitors may be used to treat constipation by
decreasing NaCl and water absorption in the colon (191).
Irritable bowel syndrome (IBS) is characterized by
chronic abdominal pain, discomfort and bloating associated with
altered bowel habits, including diarrhea and/or constipation.
Current therapy for constipation caused by IBS has side effects,
including deterioration of the condition or electrolyte
disturbances (192). Previous
studies have shown that, in addition to the above-mentioned side
effects, lubiprostone can also increase gut motility and frequency
of stool passage, relieve abdominal pain and discomfort (192), stimulate chloride secretion and
improve bowel function (193).
There have been few studies on diseases of the
hematological system associated with ClC-2. A previous study
reported the expression of swelling-and/or pH-regulated ClC-2
chloride channels in human leukemia (195). In addition, it was suggested that
the molecular identification of chloride channels may provide a
novel approach for the treatment of leukemia (195).
According to previous reports, there have been
several studies on the association between CF and ClC-2 (3,11,23,25,26,68,70,196–198,222,223). Several studies have reported that
ClC-2 may be an alternative pathway for chloride anion secretion in
CF (3,11,23),
and that ClC-2 is a potential target for therapy in CF (25,197). CF is a genetic disease, which
primarily affects the lungs, in addition to the pancreas, liver,
kidneys and intestine (222). CF
is caused by mutations in the gene encoding CFTR, the only member
of the ABC transporter family known to be a cAMP-activated chloride
channel (223). ClC-2 is a
candidate alternative chloride channel in respiratory epithelia
(196). ClC-2 is also involved in
lung morphogenesis (26,68), and can conduct chloride in mature
respiratory epithelia (23,25,197).
ClC-2 mRNA and protein are expressed in unaffected tissues in CF,
which may make up for defects in the expression of CFTR (68).
There have been more studies on diseases of the
nervous system associated with ClC-2, compared with other diseases
associated with ClC-2. Studies on diseases of the nervous system
associated with ClC-2 have predominantly focused on
leukoencephalopathy (199–202,224)
and epilepsy (30,40,203–212,225).
Leukoencephalopathy can refer specifically to any
of the following diseases: Progressive multifocal
leukoencephalopathy, toxic leukoencephalopathy, leukoencephalopathy
with vanishing white matter, leukoencephalopathy with neuroaxonal
spheroids, reversible posterior leukoencephalopathy syndrome,
megalencephalic leukoencephalopathy with subcortical cysts (MLC)
and hypertensive leukoencephalopathy. Among these, MLC is a rare
type of leukodystrophy, which is characterized by macrocephaly
emerging in the first years of life (224).
The disruption of ClC-2 can cause fluid
accumulation resulting in myelin vacuolation in mice, similar to
that observed in humans affected with MLC from mutations in MLC1 or
GlialCAM. GlialCAM is a ClC-2 binding partner and the first
auxiliary subunit of ClC-2 (199). According to a study by Jeworutzki
et al (199), neither the
stimulation of GlialCAM on ClC-2 currents, nor mislocalization of
this Cl− channel were found to damage glial
Cl− transport. Mutated GlialCAM in MLC can target ClC-2
to cell contacts in glia and activate its currents (200). A study by Jeworutzki et al
(200) showed that GlialCAM can
target the common gate deficient ClC-2 mutant, E211V/H816A, to cell
contacts without altering its function. Another study suggested
that ClC-2 is not important for MLC1 or GlialCAM localization in
the brain (201), but that it is
involved in the pathogenesis of MLC (201).
Epilepsy comprises a set of neurological diseases
characterized by epileptic seizures, and a heterogeneous disorder
characterized by recurrent unprovoked seizures, which affect ~1–3%
of the population during their lifetimes (225). ClC-2 mRNA and protein are found
in neurons and astrocytes (30,40,203). In addition, ClC-2 protein is
present at the end feet of astrocytes contacting blood vessels and
neurons close to inhibitory synapses (30). Inwardly rectifying
hyperpolarization-activated ClC-2-like currents are found in
hippocampal pyramidal cells (40,127) and in astrocytes (204–207). In hippocampal pyramidal cells and
astrocytes (208), ClC-2 may have
different effects. In neurons, ClC-2 can prevent the accumulation
of chloride anions above equilibrium due to the activation of ClC-2
by intracellular Cl− (41,127). Under these conditions,
GABAA R activity may become excitatory (208). A previous study (208) reported that loss of function
mutations of ClC-2 lead to increased excitability in certain
neurons, and that hyperpolarization-activated chloride currents are
detected in cortical astrocytes, but absent in tissues from
ClC-2-null mice (207). Another
study (211) showed that several
ClC-2 sequence abnormalities previously found in patients affected
with epilepsy are likely to represent innocuous polymorphisms,
detected by sequencing of a large collection of human DNA and
electrophysiological analysis.
Several other studies have reported that a
susceptibility locus for idiopathic generalized epilepsy (IGE) is
on chromosome 3q26 (the location of the CLCN2 gene) (209), and three mutations on ClC-2
cosegregated with IGE with autosomal dominant inheritance (210) have been identified.
In addition to the above-mentioned studies on the
association between epilepsy and ClC-2, an association between
temporal lobe epilepsy (TLE) and ClC-2 has been reported. TLE with
spontaneous recurrent attacks, and learning and memory
disabilities, is associated with neurodegeneration, abnormal
reorganization of the circuitry and loss of functional suppression
in hippocampus (212). A study by
Ge et al reported that ClC-2 contributes towards tonic
inhibition, modulated by α5 subunit-containing GABAA Rs
in the CA1 area (212).
Few studies on metabolic diseases associated with
ClC-2 have been performed, however, a study on diabetes mellitus
was found. Diabetes mellitus is a metabolic disease, which
generally causes chronic delayed wound healing. In a study by Pan
et al (213), a high
glucose concentration inhibited keratinocyte migration by
downregulating ClC-2, suggesting ClC-2 may be important during
delayed wound healing processes. In addition, the study reported
that ClC-2 is an important modulator of cell migration in
keratinocytes, although it did not discuss how ClC-2 is involved in
keratinocyte migration.
With the continuous progress in experimental and
clinical studies on the ClC family, it is clear that the activation
of ClC-2 occurs via hyperpolarization (3,9),
cell swelling (2,9), extracelluar hypotonicity (2) and extracellular acidification
(9,10), and that the expression of ClC-2 is
ubiquitous (3,11) in ureteric bud cells (12), the intestine (13–17),
gastric parietal cells (18,19),
the liver (20), the lungs
(21–25), rat retina (27), parotid acinar cells (28), guinea pig cardiac muscle (29), neuronal cells (30), rat and human airways (17), bovine trabecular meshwork (31), human trabecular meshwork (32,33)
and rat trabecular meshwork (34).
Although there has been progress in understanding the CLCN2 gene,
the molecular structure of the ClC-2 protein, the structure of
ClC-2 chloride channels, and the functional properties and
mechanisms regulating ClC-2, they remain to be fully elucidated and
there have been contradictions in previous studies (226). According to previous studies, it
is known that the disruption of ClC-2 can lead to several diseases.
Although ClC-2 is associated with the pathogenesis of several
diseases, the association between pathogenesis and ClC-2 remains to
be fully elucidated.
At present, >6,500 types of genetic disease have
been identified, among which ~3,000 are caused by a single gene
defect. In this review, the mechanisms underlying the association
between the CLCN2 gene and diseases, including azoospermia and IGE,
were discussed. In the future, investigations focused on the
determination of the CLCN2 gene may identify novel methods to treat
and prevent several diseases associated with ClC-2. In addition, as
the overexpression and underexpression of ClC-2 can cause diseases,
the development of specific ClC-2 activators and inhibitors, and
understanding the mechanism of action between the functional
properties of ClC-2 and these activators and inhibitors, is
required in future investigations to regulate the expression of
ClC-2.
As described above, ClC-2 can be regulated by
hormones, drugs and scorpion venom. Certain hormones in humans are
produced in small quantities, however, they have a substantial
effect on health, with underproduction or overproduction leading to
a variety of diseases. In addition, scorpion venom can be used to
treat cancer and lower blood pressure, and is applied for
hemostasis, anticoagulation, as an analgesic and a nerve growth
factor. Therefore, future investigations focused on the regulatory
mechanisms of hormones, drugs and scorpion venom may assist in
developing specific drugs for treating diseases associated with
ClC-2.
In conclusion, ClC-2 is important in several
diseases and, in order to fully elucidate the structure and
function of ClC-2, and the mechanisms regulating ClC-2 associated
with disease treatment, examining the associations between ClC-2
and regulators, including hormones, proteins, kinases,
transcription factors, scorpion venom, adenosine triphosphate,
clh-3, permeant anions, membrane cholesterol, tyrosine endocytosis
motif and α1-adrenoceptor is required to develop novel treatment
strategies. Therefore, further investigations are required in the
future.
The study was supported by the Library of Qiqihar
Medical University (grant no. 16TQB04; Qiqihar, China).
|
1
|
Strange K: Of mice and worms: Novel
insights into ClC-2 anion channel physiology. News Physiol Sci.
17:11–16. 2002.PubMed/NCBI
|
|
2
|
Gründer S, Thiemann A, Pusch M and Jentsch
TJ: Regions involved in the opening of ClC-2 chloride channel by
voltage and cell volume. Nature. 360:759–762. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thiemann A, Gründer S, Pusch M and Jentsch
TJ: A chloride channel widely expressed in epithelial and
non-epithelial cells. Nature. 356:57–60. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Furukawa T, Ogura T, Katayama Y and
Hiraoka M: Characteristics of rabbit ClC-2 current expressed in
Xenopus oocytes and its contribution to volume regulation. Am J
Physiol. 274:C500–C512. 1998.PubMed/NCBI
|
|
5
|
Bi MM, Hong S, Zhou HY, Wang HW, Wang LN
and Zheng YJ: Chloride channelopathies of ClC-2. Int J Mol Sci.
15:218–249. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Middleton RE, Pheasant DJ and Miller C:
Homodimeric architecture of a ClC-type chloride ion channel.
Nature. 383:337–340. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hsiao SH, Parrish AR, Nahm SS, Abbott LC,
McCool BA and Frye GD: Effects of early postnatal ethanol
intubation on GABAergic synaptic proteins. Brain Res Dev Brain Res.
138:177–185. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zúñiga L, Niemeyer MI, Varela D, Catalán
M, Cid LP and Sepulveda FV: The voltage-dependent ClC-2 chloride
channel has a dual gating mechanism. J Physiol. 555:671–682. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jentsch TJ, Stein V, Weinreich F and
Zdebik AA: Molecular structure and physiological function of
chloride channels. Physiol Rev. 82:503–568. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jordt SE and Jentsch TJ: Molecular
dissection of gating in the ClC-2 chloride channel. EMBO J.
16:1582–1592. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bösl MR, Stein V, Hübner C, Zdebik AA,
Jordt SE, Mukhophadhyay AK, Davidoff MS, Holstein AF and Jentsch
TJ: Male germ cells and photoreceptors, both depending on close
cell-cell interactions, degenerate upon ClC-2 Cl(−) channel
disruption. EMBO J. 20:1289–1299. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huber S, Braun G, Schröppel B and Horster
M: Chloride channels ClC-2 and ICln mRNA expression differs in
renal epithelial ontogeny. Kidney Int Suppl. 67:S149–S151. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gyömörey K, Yeger H, Ackerley C, Garami E
and Bear CE: Expression of the chloride channel ClC-2 in the murine
small intestine epithelium. Am J Physiol Cell Physiol.
279:C1787–C1794. 2000.PubMed/NCBI
|
|
14
|
Bali M, Lipecka J, Edelman A and Fritsch
J: Regulation of ClC-2 chloride channels in T84 cells by TGF-alpha.
Am J Physiol Cell Physiol. 280:C1588–C1598. 2001.PubMed/NCBI
|
|
15
|
Mohammad-Panah R, Gyomorey K, Rommens J,
Choudhury M, Li C, Wang Y and Bear CE: ClC-2 contributes to native
chloride secretion by a human intestinal cell line, Caco-2. J Biol
Chem. 276:8306–8313. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Catalán M, Cornejo I, Figueroa CD,
Niemeyer MI, Sepúlveda FV and Cid LP: ClC-2 in guinea pig colon:
mRNA. immunolabeling, and functional evidence for surface
epithelium localization. Am J Physiol Gastrointest Liver Physiol.
283:G1004–G1013. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lipecka J, Bali M, Thomas A, Fanen P,
Edelman A and Fritsch J: Distribution of ClC-2 chloride channel in
rat and human epithelial tissues. Am J Physiol Cell Physiol.
282:C805–C816. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Malinowska DH, Kupert EY, Bahinski A,
Sherry AM and Cuppoletti J: Cloning: Functional expression, and
characterization of a PKA-activated gastric Cl-channel. Am J
Physiol. 268:C191–C200. 1995.PubMed/NCBI
|
|
19
|
Sherry AM, Malinowska DH, Morris RE,
Ciraolo GM and Cuppoletti J: Localization of ClC-2 Cl- channels in
rabbit gastric mucosa. Am J Physiol Cell Physiol. 280:C1599–C1606.
2001.PubMed/NCBI
|
|
20
|
Roman RM, Smith RL, Feranchak AP, Clayton
GH, Doctor RB and Fitz JG: ClC-2 chloride channels contribute to
HTC cell volume homeostasis. Am J Physiol Gastrointest Liver
Physiol. 280:G344–G353. 2001.PubMed/NCBI
|
|
21
|
Cid LP, Montrose-Rafizadeh C, Smith DI,
Guggino WB and Cutting GR: Cloning of a putative human
voltage-gated chloride channel (ClC-2) cDNA widely expressed in
human tissues. Hum Mol Genet. 4:407–413. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sherry AM, Stroffekova K, Knapp LM, Kupert
EY, Cuppoletti J and Malinowska DH: Characterization of the human
pH- and PKA-activated ClC-2G(2 alpha) Cl- channel. Am J Physiol.
273:C384–C393. 1997.PubMed/NCBI
|
|
23
|
Schwiebert EM, Cid-Soto LP, Stafford D,
Carter M, Blaisdell CJ, Zeitlin PL, Guggino WB and Cutting GR:
Analysis of ClC-2 channels as an alternative pathway for chloride
conduction in cystic fibrosis airway cells. Proc Natl Acad Sci USA.
95:3879–3884. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Blaisdell CJ, Edmonds RD, Wang XT, Guggino
S and Zeitlin PL: pH-regulated chloride secretion in fetal lung
epithelia. Am J Physiol Lung Cell Mol Physiol. 278:L1248–L1255.
2000.PubMed/NCBI
|
|
25
|
Cuppoletti J, Tewari KP, Sherry AM, Kupert
EY and Malinowska DH: ClC-2 CI- channels in human lung epithelia:
Activation by arachidonic acid, amidation, and acid-activated
omeprazole. Am J Physiol Cell Physiol. 281:C46–C54. 2001.PubMed/NCBI
|
|
26
|
Murray CB, Morales MM, Flotte TR,
McGrath-Morrow SA, Guggino WB and Zeitlin PL: ClC-2: A
developmentally dependent chloride channel expressed in the fetal
lung and downregulated after birth. Am J Respir Cell Mol Biol.
12:597–604. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Enz R, Ross BJ and Cutting GR: Expression
of the voltage-gated chloride channel ClC-2 in rod bipolar cells of
the rat retina. J Neurosci. 19:9841–9847. 1999.PubMed/NCBI
|
|
28
|
Park K, Arreola J, Begenisich T and Melvin
JE: Comparison of voltage-activated CI- channels in rat parotid
acinar cells with ClC-2 in a mammalian expression system. J Membr
Biol. 163:87–95. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Britton FC, Hatton WJ, Rossow CF, Duan D,
Hume JR and Horowitz B: Molecular distribution of volume-regulated
chloride channels (ClC-2 and ClC-3) in cardiac tissues. Am J
Physiol Heart Circ Physiol. 279:H2225–H2233. 2000.PubMed/NCBI
|
|
30
|
Sik A, Smith RL and Freund TF:
Distribution of chloride channel-2-immunoreactive neuronal and
astrocytic processes in the hippocampus. Neuroscience. 101:51–65.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Srinivas SP, Maertens C, Goon LH, Goon L,
Satpathy M, Yue BY, Droogmans G and Nilius B: Cell volume response
to hyposmotic shock and elevated cAMP in bovine trabecular meshwork
cells. Exp Eye Res. 78:15–26. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Comes N, Gasull X, Gual A and Borrás T:
Differential expression of the human chloride channel genes in the
trabecular meshwork under stress conditions. Exp Eye Res.
80:801–813. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liang W and Zheng YJ: Effect of chloride
channel inhibitor in phagocytic process of human trabecular mesh
work cells. Chin J Gerontol. 30:906–908. 2010.
|
|
34
|
Wang JX, Wang SM and Liu HY: Expression of
ClC-2 Chloride channels in rat trabecular meshwork. Chin J
Biologicals. 23:274–276. 2010.
|
|
35
|
Xiong H, Li C, Garami E, Wang Y,
Ramjeesingh M, Galley K and Bear CE: ClC-2 activation modulates
regulatory volume decrease. J Membr Biol. 167:215–221. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Worrell RT, Butt AG, Cliff WH and Frizzell
RA: A volume-sensitive chloride conductance in human colonic cell
line T84. Am J Physiol. 256:C1111–C1119. 1989.PubMed/NCBI
|
|
37
|
Solc CK and Wine JJ: Swelling-induced and
depolarization-induced Cl-channels in normal and cystic fibrosis
epithelial cells. Am J Physiol. 261:C658–C674. 1991.PubMed/NCBI
|
|
38
|
Jackson PS and Strange K: Single-channel
properties of a volume-sensitive anion conductance. Current
activation occurs by abrupt switching of closed channels to an open
state. J Gen Physiol. 105:643–660. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Strange K, Emma F and Jackson PS: Cellular
and molecular physiology of volume-sensitive anion channels. Am J
Physiol. 270:C711–C730. 1996.PubMed/NCBI
|
|
40
|
Smith RL, Clayton GH, Wilcox CL, Escudero
KW and Staley KJ: Differential expression of an inwardly rectifying
chloride conductance in rat brain neurons: A potential mechanism
for cell-specific modulation of postsynaptic inhibition. J
Neurosci. 15:4057–4067. 1995.PubMed/NCBI
|
|
41
|
Staley K, Smith R, Schaack J, Wilcox C and
Jentsch TJ: Alteration of GABAA receptor function following gene
transfer of the CLC-2 chloride channel. Neuron. 17:543–551. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chu S and Zeitlin PL: Alternative mRNA
splice variants of the rat ClC-2 chloride channel gene are
expressed in lung: Genomic sequence and organization of ClC-2.
Nucleic Acids Res. 25:4153–4159. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chu S, Murray CB, Liu MM and Zeitlin PL: A
short CIC-2 mRNA transcript is produced by exon skipping. Nucleic
Acids Res. 24:3453–3457. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chu S, Blaisdell CJ, Liu MZ and Zeitlin
PL: Perinatal regulation of the ClC-2 chloride channel in lung is
mediated by Sp1 and Sp3. Am J Physiol. 276:L614–L624.
1999.PubMed/NCBI
|
|
45
|
Dutzler R: The ClC family of chloride
channels and transporters. Curr Opin Struct Biol. 16:439–446. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Dutzler R: A structural perspective on ClC
channel and transporter function. FEBS Lett. 581:2839–2844. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ramjeesingh M, Li C, Huan LJ, Garami E,
Wang Y and Bear CE: Quaternary structure of the chloride channel
ClC-2. Biochemistry. 39:13838–13847. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Varela D, Niemeyer MI, Cid LP and
Sepúlveda FV: Effect of an N-terminus deletion on voltage-dependent
gating of the ClC-2 chloride channel. J Physiol. 544:363–372. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Stölting G, Fischer M and Fahlke C: ClC-1
and ClC-2 form hetero-dimeric channels with novel protopore
functions. Pflugers Arch. 466:2191–2204. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fritsch J and Edelman A: Modulation of the
hyperpolarization-activated Cl- current in human intestinal T84
epithelial cells by phosphorylation. J Physiol. 490:115–128. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Flores CA, Niemeyer MI, Sepúlveda FV and
Cid LP: Two splice variants derived from a Drosophila melanogaster
candidate ClC gene generate ClC-2-type Cl- channels. Mol Membr
Biol. 23:149–156. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Qu C, Liang F, Smythe NM and Schulte BA:
Identification of ClC-2 and CIC-K2 chloride channels in cultured
rat type IV spiral ligament fibrocytes. J Assoc Res Otolaryngol.
8:205–219. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Fritsch J and Edelman A: Osmosensitivity
of the hyperpolarization-activated chloride current in human
intestinal T84 cells. Am J Physiol. 272:C778–C786. 1997.PubMed/NCBI
|
|
54
|
Speakem T, Kajitam H, Smith CP and Brown
PD: Inward-rectifying anion channels are expressed in the
epithelial cells of choroid plexus isolated from ClC-2 ‘knock-out’
mice. J Physiol. 539:385–390. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Valverde MA, Mintenig GM and Sepúlveda FV:
Differential effects of tamoxifen and I- on three distinguishable
chloride currents activated in T84 intestinal cells. Pflugers Arch.
425:552–554. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Chesnoy-Marchais D:
Hyperpolarization-activated chloride channels in Aplysi a neurons.
In Chloride Channels and Carriers in Nerve Muscle and Glial Cells
Alvarez-Leefmans. Russell JMFJ: USA: New York Plenum Press; 14. pp.
367–382. 1990
|
|
57
|
Arreola J, Park K, Melvin JE and
Begenisich T: Three distinct chloride channels control anion
movements in rat parotid acinar cells. J Physiol. 490:351–362.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Díaz M, Valverde MA, Higgins CF, Rucăreanu
C and Sepúlveda FV: Volume-activated chloride channels in HeLa
cells are blocked by verapamil and dideoxyforskolin. Pflugers Arch.
422:347–353. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Bond TD, Ambikapathy S, Mohammad S and
Valverde MA: Osmosensitive C1- currents and their relevance to
regulatory volume decrease in human intestinal T84 cells: Outwardly
vs. Inwardly rectifying currents. J Physiol. 511:45–54. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Furukawa T, Horikawa S, Terai T, Ogura T,
Katayama Y and Hiraoka M: Molecular cloning and characterization of
a novel truncated from (ClC-2 beta) of ClC-2 alpha (ClC-2G) in
rabbit heart. FEBS Lett. 375:56–62. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chamberlin ME and Strange K: Anisosmotic
cell volume regulation: A comparative view. Am J Physiol.
257:C159–C173. 1989.PubMed/NCBI
|
|
62
|
Nehrke K, Arreola J, Nguyen HV, Pilato J,
Richardson L, Okunade G, Baggs R, Shull GE and Melvin JE: Loss of
hyperpolarization-activated Cl(−) current in salivary acinar cells
from Clcn2 knockout mice. J Biol Chem. 277:23604–23611. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Nighot MP, Nighot PK, Ma TY, Malinowska
DH, Shull GE, Cuppoletti J and Blikslager AT: Genetic ablation of
the ClC-2 Cl- Channel disrupts mouse gastric parietal cell acid
secretion. PLoS One. 10:e01381742015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hori K, Takahashi Y, Horikawa N, Furukawa
T, Tsukada K, Takeguchi N and Sakai H: Is the ClC-2 chloride
channel involved in the Cl- secretory mechanism of gastric parietal
cells? FEBS Lett. 575:105–108. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Liang W, Zheng WX, Sun LX and Zheng YJ:
Effect of chloride channel 2 on trabecular meshwork cells under
pressure stress condition. Chin J Ophthalmol Otorhinolaryngol.
12:26–29. 2012.
|
|
66
|
Ugarte G, Delgado R, O'Day PM, Farjah F,
Cid LP, Vergara C and Bacigalupo J: Putative ClC-2 chloride channel
mediates inward rectification in Drosophila retinal photoreceptors.
J Membr Biol. 207:151–160. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Comes N, Abad E, Morales M, Borrás T, Gual
A and Gasull X: Identification and functional characterization of
ClC-2 chloride channels in trabecular meshwork cells. Exp Eye Res.
83:877–889. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Murray CB, Chu S and Zeitlin PL:
Gestational and tissue-specific regulation of C1C-2 chloride
channel expression. Am J Physiol. 271:L829–L837. 1996.PubMed/NCBI
|
|
69
|
Zhou L, Graeff RW, McCray PB Jr, Simonet
WS and Whitsett JA: Keratinocyte growth factor stimulates
CFTR-independent fluid secretion in the fetal lung in vitro. Am J
Physiol. 271:L987–L994. 1996.PubMed/NCBI
|
|
70
|
Blaisdell CJ, Morales MM, Andrade AC,
Bamford P, Wasicko M and Welling P: Inhibition of CLC-2 chloride
channel expression interrupts expansion of fetal lung cysts. Am J
Physiol Lung Cell Mol Physiol. 286:L420–L426. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Duan D, Ye L, Britton F, Horowitz B and
Hume JR: A novel anionic inward rectifier in native cardiac
myocytes. Circ Res. 86:E63–E71. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Britton FC, Wang GL, Huang ZM, Ye L,
Horowitz B, Hume JR and Duan D: Functional characterization of
novel alternatively spliced ClC-2 chloride channel variants in the
heart. J Biol Chem. 280:25871–25880. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Huang ZM, Prasad C, Britton FC, Ye LL,
Hatton WJ and Duan D: Functional role of CLC-2 chloride inward
rectifier channels in cardiac sinoatrial nodal pacemaker cells. Mol
Cell Cardiol. 47:121–132. 2009. View Article : Google Scholar
|
|
74
|
Komukai K, Brette F and Orchard CH:
Electrophysiological response of rat atrial myocytes to acidosis.
Am J Physiol Heart Circ Physiol. 283:H715–H724. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Komukai K, Brette F, Pascarel C and
Orchard CH: Electrophysiological response of rat ventricular
myocytes to acidosis. Am J Physiol Heart Circ Physiol.
283:H412–H422. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Földy C, Lee SH, Morgan RJ and Soltesz I:
Regulation of fast-spiking basket cell synapses by the chloride
channel ClC-2. Nat Neurosci. 13:1047–1049. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Cid LP, Niemeyer MI, Ramírez A and
Sepúlveda FV: Splice variants of a ClC-2 chloride channel with
differing functional characteristics. Am J Physiol Cell Physiol.
279:C1198–C1210. 2000.PubMed/NCBI
|
|
78
|
Garcia-Olivares J, Alekov A, Boroumand MR,
Begemann B, Hidalgo P and Fahlke C: Gating of human ClC-2 chloride
channels and regulation by carboxy-terminal domains. J Physiol.
586:5325–5336. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Eckmann L and Gillin FD: Microbes and
microbial toxins: Paradigms for microbial-mucosal interactions I.
Pathophysiological aspects of enteric infections with the lumen
dwelling protozoan pathogen Giardia lamblia. Am J Physiol
Gastrointest Liver Physiol. 280:G1–G6. 2001.PubMed/NCBI
|
|
80
|
Upcroft P and Upcroft JA: Drug targets and
mechanisms of resistance in the anaerobic protozoa. Clin Microbiol
Rev. 14:150–164. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Hawrelak J: Giardiasis: Pathophysiology
and management. Altern Med Rev. 8:129–142. 2003.PubMed/NCBI
|
|
82
|
Moreno-Galindo EG, Rodríguez-Elías JC,
Ramírez-Herrera MA, Sánchez-Chapula JA and Navarro-Polanco RA: The
principal conductance in Giardia lamblia trophozoites possesses
functional properties similar to the mammalian ClC-2 current.
Pflugers Arch. 466:915–924. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Villaz M, Cinniger JC and Moody WJ: A
voltage-gated chloride channel in ascidian embryos modulated by
both the cell cycle clock and cell volume. J Physiol. 488:689–699.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Okada Y: Volume expansion-sensing
outward-rectifier Cl- channel: Fresh start to the molecular
identity and volume sensor. Am J Physiol. 273:C755–C789.
1997.PubMed/NCBI
|
|
85
|
Tilly BC, van den Berghe N, Tertoolen LG,
Edixhoven MJ and de Jonge HR: Protein tyrosine phosphorylation is
involved in osmoregulation of ionic conductances. J Biol Chem.
268:19919–19922. 1993.PubMed/NCBI
|
|
86
|
Sorota S: Tyrosine protein kinase
inhibitors prevent activation of cardiac swelling-Induced chloride
current. Pflugers Arch. 431:178–185. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Voets T, Manolopoulos V, Eggermont J,
Ellory C, Droogmans G and Nilius B: Regulation of a
swelling-activated chloride current in bovine endothelium by
protein tyrosine phosphorylation and G proteins. J Physiol.
506:341–352. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Lepple-Wienhues A, Szabò I, Laun T, Kaba
NK, Gulbins E and Lang F: The tyrosine kinase p56lck mediates
activation of swelling-induced chloride channels in lymphocytes. J
Cell Biol. 141:281–286. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Santos Ornellas D, Grozovsky R, Goldenberg
RC, Carvalho DP, Fong P, Guggino WB and Morales M: Thyroid hormone
modulates ClC-2 chloride channel gene expression in rat renal
proximal tubules. J Endocrinol. 178:503–511. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Nascimento DS, Reis CU, Goldenberg RC,
Ortiga-Carvalho TM, Pazos-Moura CC, Guggino SE, Guggino WB and
Morales MM: Estrogen modulates ClC-2 chloride channel gene
expression in rat kidney. Pflugers Arch. 446:593–599. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Morales MM, Nascimento DS, Capella MA,
Lopes AG and Guggino WB: Arginine vasopressin regulates CFTR and
ClC-2 mRNA expression in rat kidney cortex and medulla. Pflugers
Arch. 443:202–211. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Moeser AJ, Haskell MM, Shifflett DE,
Little D, Schultz BD and Blikslager AT: ClC-2 chloride secretion
mediates prostaglandin-induced recovery of barrier function in
ischemia-injured porcine ileum. Gastroenterology. 127:802–815.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Moeser AJ, Nighot PK, Engelke KJ, Ueno R
and Blikslager AT: Recovery of mucosal barrier function in ischemic
porcine ileum and colon is stimulated by a novel agonist of the
ClC-2 chloride channel, lubiprostone. Am J Physiol Gastrointest
Liver Physiol. 292:G647–G656. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Cuppoletti J, Malinowska DH, Tewari KP, Li
QJ, Sherry AM, Patchen ML and Ueno R: SPI-0211 activates T84 cell
chloride transport and recombinant human ClC-2 chloride currents.
Am J Physiol Cell Physiol. 287:C1173–C1183. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Bao HF, Liu L, Self J, Duke BJ, Ueno R and
Eaton DC: A synthetic prostone activates apical chloride channels
in A6 epithelial cells. Am J Physiol Gastrointest Liver Physiol.
295:G234–G251. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Bijvelds MJ, Bot AG, Escher JC and De
Jonge HR: Activation of intestinal Cl- secretion by lubiprostone
requires the cystic fibrosis transmembrane conductance regulator.
Gastroenterology. 137:976–985. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Ao M, Venkatasubramanian J, Boonkaewwan C,
Ganesan N, Syed A, Benya RV and Rao MC: Lubiprostone activates Cl-
secretion via cAMP signaling and increases membrane CFTR in the
human colon carcinoma cell line, T84. Dig Dis Sci. 56:339–351.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Norimatsu Y, Moran AR and MacDonald KD:
Lubiprostone activates CFTR, but not ClC-2, via the prostaglandin
receptor (EP(4)). Biochem Biophys Res Commun. 426:374–379. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Jin Y, Pridgen TA and Blikslager AT:
Pharmaceutical activation or genetic absence of ClC-2 alters tight
junctions during experimental colitis. Inflamm Bowel Dis.
21:2747–2757. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Qiu SB, Liang Y and Liu P: Effect of ClC-2
ClC-3 CFTR expression in rats affected with the myocardial ischemia
reperfusion injury by Chinese native medicine compound
Guangxinkang. Chin J Integrative Med Cardio-/Cerebrovascular Dis.
11:1482–1485. 2013.
|
|
101
|
Dhani SU, Mohammad-Panah R, Ahmed N,
Ackerley C, Ramjeesingh M and Bear CE: Evidence for a functional
interaction between the ClC-2 chloride channel and the retrograde
motor dynein complex. J Biol Chem. 278:16262–16270. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Furukawa T, Ogura T, Zheng YJ, Tsuchiya H,
Nakaya H, Katayama Y and Inagaki N: Phosphorylation and functional
regulation of ClC-2 chloride channels expressed in Xenopus oocytes
by M cyclin-dependent protein kinase. J Physiol. 540:883–893. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Zheng YJ, Furukawa T, Ogura T, Tajimi K
and Inagaki NM: M phase-specific expression and
phosphorylation-dependent ubiquitination of the ClC-2 channel. J
Biol Chem. 277:32268–32273. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Ahmed N, Ramjeesingh M, Wong S, Varga A,
Garami E and Bear CE: Chloride channel activity of ClC-2 is
modified by the actin cytoskeleton. Biochem J. 352:789–794. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Hinzpeter A, Lipecka J, Brouillard F,
Baudoin-Legros M, Dadlez M, Edelman A and Fritsch J: Association
between Hsp90 and the ClC-2 chloride channel upregulates channel
function. Am J Physiol Cell Physiol. 290:C45–C56. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Pérez-Rius C, Gaitán-Peñas H, Estévez R
and Barrallo-Gimeno A: Identification and characterization of the
zebrafish ClC-2 chloride channel orthologs. Pflugers Arch.
467:1769–1781. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Chu S, Blaisdell CJ, Bamford P and Ferro
TJ: Interferon-gamma regulates ClC-2 chloride channel in lung
epithelial cells. Biochem Biophys Res Commun. 324:31–39. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Palmada M, Dieter M, Boehmer C, Waldegger
S and Lang F: Serum and glucocorticoid inducible kinases
functionally regulate ClC-2 channels. Biochem Biophys Res Commun.
321:1001–1006. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Hosseinzadeh Z, Bhavsar SK and Lang F:
Downregulation of ClC-2 by JAK2. Cell Physiol Biochem. 29:737–742.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Lang F, Föller M, Lang K, Lang P, Ritter
M, Vereninov A, Szabo I, Huber SM and Gulbins E: Cell volume
regulatory ion channels in cell proliferation and cell death.
Methods Enzymol. 428:209–225. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Warsi J, Elvira B, Hosseinzadeh Z,
Shumilina E and Lang F: Downregulation of chloride channel ClC-2 by
Janus kinase 3. J Membr Biol. 247:387–393. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Klaus F, Laufer J, Czarkowski K,
Strutz-Seebohm N, Seebohm G and Lang F: PIKfyve-dependent
regulation of the Cl- channel ClC-2. Biochem Biophys Res Commun.
381:407–411. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Park K, Begenisich T and Melvin JE:
Protein kinase A activation phosphorylates the rat ClC-2 Cl-
channel but does not change activity. J Membr Biol. 182:31–37.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Warsi J, Hosseinzadeh Z, Elvira B,
Bissinger R, Shumilina E and Lang F: Regulation of ClC-2 activity
by SPAK and OSR1. Kidney Blood Press Res. 39:378–387. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Vij N and Zeitlin PL: Regulation of the
ClC-2 lung epithelial chloride channel by glycosylation of SP1. Am
J Respir Cell Mol Biol. 34:754–759. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Holmes KW, Hales R, Chu S, Maxwell MJ,
Mogayzel PJ Jr and Zeitlin PL: Modulation of Sp1 and Sp3 in lung
epithelial cells regulates ClC-2 chloride channel expression. Am J
Respir Cell Mol Biol. 29:499–505. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Cuppoletti J, Chakrabarti J, Tewari KP and
Malinowska DH: Differentiation between human ClC-2 and CFTR Cl-
channels with pharmacological agents. Am J Physiol Cell Physiol.
307:C479–C492. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Thompson CH, Olivetti PR, Fuller MD,
Freeman CS, McMaster D, French RJ, Pohl J, Kubanek J and McCarty
NA: Isolation and characterization of a high affinity peptide
inhibitor of ClC-2 chloride channels. J Biol Chem. 284:26051–26062.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Zhao B, Quan H, Ma T, Tian Y, Cai Q and Li
H: 4,4′-Diisothiocyanostilbene-2,2′-disulfonic Acid (DIDS)
ameliorates ischemia-hypoxia-induced white matter damage in
neonatal rats through inhibition of the voltage-gated chloride
channel ClC-2. Int J Mol Sci. 16:10457–10469. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Liang W and Zheng YJ: Effect of interfered
ClC-2 gene expression on cell cycle of human trabecular mesh work
cells. Chin J Gerontol. 30:1070–1072. 2010.
|
|
121
|
Thompson CH, Fields DM, Olivetti PR,
Fuller MD, Zhang ZR, Kubanek J and McCarty NA: Inhibition of ClC-2
chloride channels by a peptide component or components of scorpion
venom. J Membr Biol. 208:65–76. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Baglole CJ, Sigalet DL and Meddings JB:
Alpha1-adrenoceptors down-regulate ClC-2 chloride channels in
epithelial cells from the acutely denervated jejunum. Eur J
Pharmacol. 565:202–206. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Huber SM, Duranton C, Henke G, Van De Sand
C, Heussler V, Shumilina E, Sandu CD, Tanneur V, Brand V,
Kasinathan RS, et al: Plasmodium induces swelling-activated ClC-2
anion channels in the host erythrocyte. J Biol Chem.
279:41444–41452. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Stölting G, Teodorescu G, Begemann B,
Schubert J, Nabbout R, Toliat MR, Sander T, Nürnberg P, Lerche H
and Fahlke C: Regulation of ClC-2 gating by intracellular ATP.
Pflugers Arch. 465:1423–1437. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Dhani SU, Kim Chiaw P, Huan LJ and Bear
CE: ATP depletion inhibits the endocytosis of ClC-2. J Cell
Physiol. 214:273–280. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Madison DV, Malenka RC and Nicoll RA:
Phorbol esters block a voltage-sensitive chloride current in
hippocampal pyramidal cells. Nature. 321:695–697. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Staley K: The role of an inwardly
rectifying chloride conductance in postsynaptic inhibition. J
Neurophysiol. 72:273–284. 1994.PubMed/NCBI
|
|
128
|
Rinke I, Artmann J and Stein V: ClC-2
voltage-gated channels constitute part of the background
conductance and assist chloride extrusion. J Neurosci.
30:4776–4786. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Schriever AM, Friedrich T, Pusch M and
Jentsch TJ: CLC chloride channels in Caenorhabditis elegans. J Biol
Chem. 274:34238–34244. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Nehrke K, Begenisich T, Pilato J and
Melvin JE: Into ion channel and transporter function.
Caenorhabditis elegans ClC-type chloride channels: Novel variants
and functional expression. Am J Physiol Cell Physiol.
279:C2052–C2066. 2000.PubMed/NCBI
|
|
131
|
Denton J, Nehrke K, Rutledge E, Morrison R
and Strange K: Alternative splicing of N- and C-termini of a C.
Elegans ClC channel alters gating and sensitivity to external Cl-
and H+. J Physiol. 555:97–114. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Zhang M, Chung SH, Fang-Yen C, Craig C,
Kerr RA, Suzuki H, Samuel AD, Mazur E and Schafer WR: A
self-regulating feed-forward circuit controlling C. Elegans
egg-laying behavior. Curr Biol. 18:1445–1455. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Desai C and Horvitz HR: Caenorhabditis
elegans mutants defective in the functioning of the motor neurons
responsible for egg laying. Genetics. 121:703–721. 1989.PubMed/NCBI
|
|
134
|
Lickteig KM, Duerr JS, Frisby DL, Hall DH,
Rand JB and Miller DM III: Regulation of neurotransmitter vesicles
by the homeodomain protein UNC-4 and its transcriptional
corepressor UNC-37/groucho in Caenorhabditis elegans cholinergic
motor neurons. J Neurosci. 21:2001–2014. 2001.PubMed/NCBI
|
|
135
|
Nathoo AN, Moeller RA, Westlund BA and
Hart AC: Identification of neuropeptide-like protein gene families
in Caenorhabditis elegans and other species. Proc Natl Acad Sci
USA. 98:14000–14005. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Kim K and Li C: Expression and regulation
of an FMRFamide-related neuropeptide gene family in Caenorhabditis
elegans. J Comp Neuro. 475:540–550. 2004. View Article : Google Scholar
|
|
137
|
Branicky R, Miyazaki H, Strange K and
Schafer WR: The voltage-gated anion channels encoded by clh-3
regulate egg laying in C. Elegans by modulating motor neuron
excitability. J Neurosci. 34:764–775. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Niemeyer MI, Cid LP, Yusef YR, Briones R
and Sepúlveda FV: Voltage-dependent and -independent titration of
specific residues accounts for complex gating of a ClC chloride by
extracellular protons. J Physiol. 587:1387–1400. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Sánchez-Rodríguez JE, De Santiago-Castillo
JA and Arreola J: Permeant anions contribute to voltage dependence
of ClC-2 chloride channel by interacting with the protopore gate. J
Physiol. 588:2545–2556. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Hinzpeter A, Fritsch J, Borot F, Trudel S,
Vieu DL, Brouillard F, Baudouin-Legros M, Clain J, Edelman A and
Ollero M: Membrane cholesterol content modulates ClC-2 gating and
sensitivity to oxidative stress. J Biol Chem. 282:2423–2432. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Cornejo I, Niemeyer MI, Zúñiga L, Yusef
YR, Sepúlveda FV and Cid LP: Rapid recycling of ClC-2 chloride
channels between plasma membrane and endosomes: Role of a tyrosine
endocytosis motif in surface retrieval. J Cell Physio. 221:650–657.
2009. View Article : Google Scholar
|
|
142
|
Light DB, Schwiebert EM, Karlson KH and
Stanton BA: Atrial natriuretic peptide inhibits a cation channel in
renal inner medullary collecting duct cells. Science. 243:393–395.
1999.
|
|
143
|
Masilamani S, Kim GH, Mitchell C, Wade JB
and Knepper MA: Aldosterone-mediated regulation of ENaC alpha,
beta, and gamma subunit proteins in rat kidney. J Clin Invest.
104:R19–R23. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Ewart HS and Klip A: Hormonal regulation
of the Na(+)-K(+)-ATPase: Mechanisms underlying rapid and sustained
changes in pump activity. Am J Physiol. 269:C295–C311.
1995.PubMed/NCBI
|
|
145
|
Morales MM, Brucoli HCP, Malnic G and
Lopes AG: Role of thyroid hormones in renal tubule acidification.
Mol Cell Biochem. 154:17–21. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Katz AI, Emmanouel DS and Lindheimer MD:
Thyroid hormone and the kidney. Nephron. 15:223–249. 1975.
View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Shirota T, Shinoda T, Yamada T and Aizawa
T: Alteration of renal function in hyperthyroidism: Increased
tubular secretion of creatinine and decreased distal tubule
delivery of chloride. Metabolism. 41:402–405. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Liu XM, Bai Y and Guo ZS: Study on urinary
function and metabolism of water and electrolytes in primary
hypothyroidism. Zhonghua Nei Ke Za Zhi. 29:299–302.318. 1990.(In
Chinese). PubMed/NCBI
|
|
149
|
Capasso G, De Tommaso G, Pica A, Anastasio
P, Capasso J, Kinne R and De Santo NG: Effects of thyroid hormones
on heart and kidney functions. Miner Electrolyte Metab. 25:57–74.
1999. View Article : Google Scholar
|
|
150
|
Weir RJ, Briggs E, Mack A, Naismith L,
Taylor L and Wilson E: Blood pressure in women taking oral
contraceptives. Br Med J. 1:533–535. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Meade TW, Haines AP, North WR, Chakrabarti
R, Howarth DJ and Stirling Y: Haemostatic, lipid, and
blood-pressure profiles of women on oral contraceptives containing
50 microgram or 30 microgram oestrogen. Lancet. 2:948–951. 1977.
View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Seeger H, Armbruster FP, Mueck AO and
Lippert TH: The effect of estradiol on urodilatin production in
postmenopausal women. Arch Gynecol Obstet. 262:65–68. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Brunette MG and Leclerc M: Effect of
estrogen on calcium and sodium transport by the nephron luminal
membranes. J Endocrinol. 170:441–450. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Ueno R, Osama H, Habe T, Engelke K and
Patchen M: Oral SPI-0211 increases intestinal fluid secretion and
chloride concentration without altering serum electrolyte levels
(Abstract). Gastroenterology. 126:A–298. 2004.
|
|
155
|
Johanson JF, Gargano M, Hollan PC, Patchen
ML and Ueno R: Phase III efficacy and safety of RU-0211, a novel
chloride channel activator, for the treatment of constipation.
Gastroenterology. 124:A482003. View Article : Google Scholar
|
|
156
|
Johanson JF, Gargano MA, Holland PC,
Patchen ML and Ueno R: Phase III randomized withdrawal study of
RU-0211 a novel chloride channel activator for the treatment of
constipation (Abstract). Gastroenterology. 126:A–100. 2004.
|
|
157
|
Lacy BE and Levy LC: Lubiprostone: A
chloride channel activator. J Clin Gastroenterol. 41:345–351. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Shisheva A, Sbrissa D and Lkonomov O:
Cloning, characterization, and expression of a novel Zn2+-binding
FYVE finger-containing phosphoinositide kinase in insulin-sensitive
cells. Mol Cell Biol. 19:623–634. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Morris DL and Rui L: Recent advances in
understanding leptin signaling and leptin resistance. Am J Physiol
Endocrinol Metab. 297:E1247–E1259. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Brooks AJ and Waters MJ: The growth
hormone receptor: Mechanism of activation and clinical
implications. Nat Rev Endocrinol. 6:515–525. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Spivak JL: Narrative review:
Thrombocytosis, polycythemia vera, and JAK2 mutations: The
phenotypic mimicry of chronic myeloproliferation. Ann Intern Med.
152:300–306. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Lopez AF, Hercus TR, Ekert P, Littler DR,
Guthridge M, Thomas D, Ramshaw HS, Stomski F, Perugini M, D'Andrea
R, et al: Molecular basis of cytokine receptor activation. IUBMB
Life. 62:509–518. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Tefferi A, Skoda R and Vardiman JW:
Myeloproliferative neoplasms: Contemporary diagnosis using
histology and genetics. Nat Rev Clin Oncol. 6:627–637. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Baskin R, Majumder A and Sayeski PP: The
recent medicinal chemistry development of Jak2 tyrosine kinase
small molecule inhibitors. Curr Med Chem. 17:4551–4558. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Ho K, Valdez F, Garcia R and Tirado CA:
JAK2 Translocations in hematological malignancies: Review of the
literature. J Assoc Genet Technol. 36:107–109. 2010.PubMed/NCBI
|
|
166
|
Oh ST and Gotlib J: JAK2 V617F and beyond:
Role of genetics and aberrant signaling in the pathogenesis of
myeloproliferative neoplasms. Expert Rev Hematol. 3:323–337. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Tefferi A: Novel mutations and their
functional and clinical relevance in myeloproliferative neoplasms:
JAK2, MPL, TET2, ASXL1, CBL, IDH and IKZF1. Leukemia. 24:1128–1138.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Pardanani A, Vannucchi AM, Passamonti F,
Cervantes F, Barbui T and Tefferi A: JAK inhibitor therapy for
myelofibrosis: Critical assessment of value and limitations.
Leukemia. 25:218–225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Santos FP and Verstovsek S: JAK2
inhibitors: What's the true therapeutic potential? Blood Rev.
25:53–63. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Mahfouz RA, Hoteit R, Salem Z, Bazarbachi
A, Mugharbel A, Farhat F, Ziyadeh A, Ibrahim A and Taher A: JAK2
V617F gene mutation in the laboratory work-up of myeloproliferative
disorders: Experience of a major referral center in Lebanon. Genet
Test Mol Biomarkers. 15:263–265. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Gatsios P, Terstegen L, Schliess F,
Häussinger D, Kerr IM, Heinrich PC and Graeve L: Activation of the
Janus kinase/signal transducer and activator of transcription
pathway by osmotic shock. J Biol Chem. 273:22962–22968. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Garnovskaya MN, Mukhin YV, Vlasova TM and
Raymond JR: Hypertonicity activates Na+/H+ exchange through Janus
kinase 2 and calmodulin. J Biol Chem. 278:16908–16915. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Uckun FM, Vassilev A, Dibirdik I and
Tibbles H: Targeting JAK3 tyrosine kinase-linked signal
transduction pathways with rationally-designed inhibitors.
Anticancer Agents Med Chem. 7:612–623. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
de Totero D, Meazza R, Capaia M, Fabbi M,
Azzarone B, Balleari E, Gobbi M, Cutrona G, Ferrarini M and Ferrini
S: The opposite effects of IL-15 and IL-21 on CLL B cells correlate
with differential activation of the JAK/STAT and ERK1/2 pathways.
Blood. 111:517–524. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Fainstein N, Vaknin I, Einstein O, Zisman
P, Ben Sasson SZ, Baniyash M and Ben-Hur T: Neural precursor cells
inhibit multiple inflammatory signals. Mol Cell Neurosci.
39:335–341. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Nakayama J, Yamamoto M, Hayashi K, Satoh
H, Bundo K, Kubo M, Goitsuka R, Farrar MA and Kitamura D: BLNK
suppresses pre-B-cell leukemogenesis through inhibition of JAK3.
Blood. 113:1483–1492. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Kim BH, Oh SR, Yin CH, Lee S, Kim EA, Kim
MS, Sandoval C, Jayabose S, Bach EA, Lee HK and Baeg GH: MS-1020 is
a novel small molecule that selectively inhibits JAK3 activity. Br
J Haematol. 148:132–143. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Walters DK, Mercher T, Gu TL, O'Hare T,
Tyner JW, Loriaux M, Goss VL, Lee KA, Eide CA, Wong MJ, et al:
Activating alleles of JAK3 in acute megakaryoblastic leukemia.
Cancer Cell. 10:65–75. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Malinge S, Ragu C, Della-Valle V, Pisani
D, Constantinescu SN, Perez C, Villeval JL, Reinhardt D,
Landman-Parker J, Michaux L, et al: Activating mutations in human
acute megakaryoblastic leukemia. Blood. 112:4220–4226. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Haan C, Rolvering C, Raulf F, Kapp M,
Drückes P, Thoma G, Behrmann I and Zerwes HG: Jak1 has a dominant
role over Jak3 in signal transduction through γc-containing
cytokine receptors. Chem Biol. 18:314–323. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Karin M: Too many transcription factors:
Positive and negative interactions. New Biol. 2:126–131.
1990.PubMed/NCBI
|
|
182
|
Latchman DS: Transcription factors: An
overview. Int J Biochem Cell Biol. 29:1305–1312. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Nandoskar P, Wang Y, Wei R, Liu Y, Zhao P,
Lu M, Huang J, Thomas P, Trousdale MD and Ding C: Changes of
chloride channels in the lacrimal glands of a rabbit model of
Sjögren syndrome. Cornea. 31:273–279. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Ding C, Parsa L, Nandoskar P, Zhao P, Wu K
and Wang Y: Duct system of the rabbit lacrimal gland: Structural
characteristics and role in lacrimal secretion. Invest Ophthalmol
Vis Sci. 51:2960–2967. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Ouyang WB: Correlation between ClC-2
chloride channel and age related cataract Master's thesis. Dalian
Med Univ Dalian. 2013.
|
|
186
|
Li HB, Han DM, Zhong B, Fan EZ and Liu ZY:
Expressions of chloride channel ClC-2 and ClC-3 in human nasal
polyps. J Clin Otorhinolaryngol. 17:266–277. 2003.
|
|
187
|
Edwards MM, de Evsikova Marín C, Collin
GB, Gifford E, Wu J, Hicks WL, Whiting C, Varvel NH, Maphis N, Lamb
BT, et al: Photoreceptor degeneration, azoospermia,
leukoencephalopathy, and abnormal RPE cellfunction in mice
expressing an early stop mutation in CLCN2. Invest Ophthalmol Vis
Sci. 51:3264–3272. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Xu ZN, Zheng XY, Zhao W, Xin H and Han ZG:
Expression and significance of chloride channel ClC-2 in
nonsmall-cell lung cancer. Chin J Lab Diagn. 16:60–62. 2012.
|
|
189
|
Diener M, Bertog M, Fromm M and Scharrer
E: Segmental heterogeneity of swelling-induced Cl- transport in rat
small intestine. Pflugers Arch. 432:293–300. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Joo NS, Clarke LL, Han BH, Forte LR and
Kim HD: Cloning of ClC-2 chloride channel from murine duodenum and
its presence in CFTR knockout mice. Biochim Biophys Acta.
1446:431–437. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Catalán MA, Flores CA, González-Begne M,
Zhang Y, Sepúlveda FV and Melvin JE: Severe defects in absorptive
ion transport in distal colons of mice that lack ClC-2 channels.
Gastroenterology. 142:346–354. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Lacy BE and Chey WD: Lubiprostone: Chronic
constipation and irritable bowel syndrome with constipation. Expert
Opin Pharmacother. 10:143–152. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Crowell MD, Harris LA, DiBaise JK and
Olden KW: Activation of type-2 chloride channels: A novel
therapeutic target for the treatment of chronic constipation. Curr
Opin Invest Drugs. 8:66–70. 2007.
|
|
194
|
Chen ZY, Wang YG, Yang P, Huang WG, Zhou
YS and Feng XS: Relationship between ClC-2 and intestinal mucosal
barrier in rats with obstructive jaundice. World Chin J
Digestology. 19:2829–2834. 2011.
|
|
195
|
Jiang B, Hattori N, Liu B, Kitagawa K and
Inagaki C: Expression of swelling- and/or pH-regulated chloride
channels (ClC-2,3,4 and 5) in human leukemic and normal immune
cells. Life Sci. 70:1383–1394. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Blaisdell CJ, Howard TD, Stern A, Bamford
P, Bleecker ER and Stine OC: CLC-2 single nucleotide polymorphisms
(SNPs) as potential modifiers of cystic fibrosis disease severity.
BMC Med Genet. 5:262004. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Cuppoletti J, Tewari KP, Sherry AM, Kupert
EY and Malinowska DH: Human ClC-2- chloride channels can be
activated Potential for therapy in cystic fibrosis. Faseb J.
15:A8472001.
|
|
198
|
Zdebik AA, Cuffe JE, Bertog M, Korbmacher
C and Jentsch TJ: Additional disruption of the ClC-2 Cl(−) channel
does not exacerbate the cystic fibrosis phenotype of cystic
fibrosis transmembrane conductance regulator mouse models. J Biol
Chem. 279:22276–22283. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Jeworutzki E, López-Hernández T,
Capdevila-Nortes X, Sirisi S, Bengtsson L, Montolio M, Zifarelli G,
Arnedo T, Müller CS, Schulte U, et al: GlialCAM, a protein
defective in a leukodystrophy, serves as a ClC-2 Cl(−) channel
auxiliary subunit. Neuron. 73:951–961. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Jeworutzki E, Lagostena L, Elorza-Vidal X,
López-Hernández T, Estévez R and Pusch M: GlialCAM a CLC-2 Cl(−)
channel subunit activates the slow gate of CLC chloride channels.
Biophys J. 107:1105–1116. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Hoegg-Beiler MB, Sirisi S, Orozco IJ,
Ferrer I, Hohensee S, Auberson M, Gödde K, Vilches C, de Heredia
ML, Nunes V, et al: Disrupting MLC1 and GlialCAM and ClC-2
interactions in leukodystrophy entails glial chloride channel
dysfunction. Nat Commun. 5:34752014. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Scheper GC, van Berkel CG, Leisle L, de
Groot KE, Errami A, Jentsch TJ and Van der Knaap MS: Analysis of
CLCN2 as candidate gene for megalencephalic leukoencephalopathy
with subcortical cysts. Genet Test Mol Biomarkers. 14:255–257.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Gulácsi A, Lee CR, Sik A, Viitanen T,
Kaila K, Tepper JM and Freund TF: Cell type-specific differences in
chloride-regulatory mechanisms and GABA(A) receptor-mediated
inhibition in rat substantia nigra. J Neurosci. 23:8237–8246.
2003.PubMed/NCBI
|
|
204
|
Ferroni S, Marchini C, Nobile M and
Rapisarda C: Characterization of an inwardly rectifying chloride
conductance expressed by cultured rat cortical astrocytes. Glia.
21:217–227. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Nobile M, Pusch M, Rapisarda C and Ferroni
S: Single-channel analysis of a ClC-2-like chloride conductance in
cultured rat cortical astrocytes. FEBS Lett. 479:10–14. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Makara JK, Petheö GL, Tóth A and Spät A:
pH-sensitive inwardly rectifying chloride current in cultured rat
cortical astrocytes. Glia. 34:52–58. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Makara JK, Rappert A, Matthias K,
Steinhäuser C, Spät A and Kettenmann H: Astrocytes from mouse brain
slices express ClC-2-mediated Cl- currents regulated during
development and after injury. Mol Cell Neurosci. 23:521–530. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Niemeyer MI, Yusef YR, Cornejo I, Flores
CA, Sepúlveda FV and Cid LP: Functional evaluation of human ClC-2
chloride channel mutations associated with idiopathic generalized
epilepsies. Physiol Genomics. 19:74–83. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
209
|
Sander T, Schulz H, Saar K, Gennaro E,
Riggio MC, Bianchi A, Zara F, Luna D, Bulteau C, Kaminska A, et al:
Genome search for susceptibility loci of common idiopathic
generalised epilepsies. Hum Mol Genet. 9:1465–1472. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Haug K, Warnstedt M, Alekov AK, Sander T,
Ramirez A, Poser B, Maljevic S, Hebeisen S, Kubisch C, Rebstock J,
et al: Mutations in CLCN2 encoding a voltage-gated chloride channel
are associated with idiopathic generalized epilepsies. Nat Genet.
33:527–532. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
211
|
Blanz J, Schweizer M, Auberson M, Maier H,
Muenscher A, Hübner CA and Jentsch TJ: Leukoencephalopathy upon
disruption of the chloride channel ClC-2. J Neurosci. 27:6581–6589.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Ge YX, Liu Y, Tang HY, Liu XG and Wang X:
ClC-2 contributes to tonic inhibition mediated by α5
subunit-containing GABA(A) receptor in experimental temporal lobe
epilepsy. Neuroscience. 186:120–127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Pan F, Guo R, Cheng W, Chai L, Wang W, Cao
C and Li S: High glucose inhibits ClC-2 chloride channels and
attenuates cell migration of rat keratinocytes. Drug Des Devel
Ther. 9:4779–4791. 2015.PubMed/NCBI
|
|
214
|
Borchers AT, Naguwa SM, Keen CL and
Gershwin ME: Immunopathogenesis of Sjögren's syndrome. Clin Rev
Allergy Immunol. 25:89–104. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Nocturne G and Mariette X: Advances in
understanding the pathogenesis of primary Sjogren's syndrome. Nat
Rev Rheumatol. 9:544–556. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Guo Z, Song D, Azzarolo AM, Schechter JE,
Warren DW, Wood RL, Mircheff AK and Kaslow HR: Autologous lacrimal
lymphoid mixed-cell reactions induce dacryoadenitis in rabbits. Exp
Eye Res. 71:23–31. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Zhu Z, Stevenson D, Schechter JE, Mircheff
AK, Atkinson R and Trousdale MD: Lacrimal histopathology and ocular
surface disease in a rabbit model of autoimmune dacryoadenitis.
Cornea. 22:25–32. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Thomas PB, Zhu Z, Selvam S, Samant DM,
Stevenson D, Mircheff AK, Schechter JE, Song SW and Trousdale MD:
Autoimmune dacryoadenitis and keratoconjunctivitis induced in
rabbits by subcutaneous injection of autologous lymphocytes
activated ex vivo against lacrimal antigens. J Autoimmun.
31:116–122. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Chan HC, Ruan YC, He Q, Chen MH, Chen H,
Xu WM, Chen WY, Xie C, Zhang XH and Zhou Z: The cystic fibrosis
transmembrane conductance regulator in reproductive health and
disease. J Physiol. 587:2187–2195. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
220
|
Yeung CH, Barfield JP and Cooper TG:
Chloride channels in physiological volume regulation of human
spermatozoa. Biol Reprod. 73:1057–1063. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Anderson MP, Sheppard DN, Berger HA and
Welsh M: Chloride channels in the apical membrane of normal and
cystic fibrosis airway and intestinal epithelia. Am J Physiol.
263:L1–L14. 1992.PubMed/NCBI
|
|
222
|
O'Sullivan BP and Freedman SD: Cystic
fibrosis. Lancet. 373:1891–1904. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
223
|
Riordan JR, Rommens JM, Kerem B, Alon N,
Rozmahel R, Grzelczak Z, Zielenski J, Lok S, Plavsic N, Chou JL, et
al: Identification of the cystic fibrosis gene: Cloning and
characterization of complementary. Science. 245:1066–1073. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
224
|
Van der Knaap MS, Barth PG, Stroink H, van
Nieuwenhuizen O, Arts WF, Hoogenraa F and Valk J:
Leukoencephalopathy with swelling and a discrepantly mild clinical
course in eight children. Ann Neurol. 37:324–334. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
225
|
Hauser WA, Annegers JF and Rocca WA:
Descriptive epidemiology of epilepsy: Contributions of
population-based studies from Rochester, Minnesota. Mayo Clin Proc.
71:576–586. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
226
|
Ji ZY: Research progress of chloride
channel ClC- 2. J Kunm Med Univ. 30:68–72. 2009.
|