Introduction
Squamous cell carcinoma (SCC) is derived from
malignant transformation of epithelial cells, including cells of
the epidermis. SCC is a common skin malignancy worldwide (1), and its occurrence has significantly
increased in recent years. SCC is highly aggressive, grows quickly
and has an elevated frequency of metastasis; Squamous cell
carcinoma of the skin is frequently treated by surgical excision,
Mohs surgery or electrodesiccation and curettage (2,3). The
disadvantages of these treatments are trauma and scar formation,
and are only applied to small skin lesions. Radiation therapy and
traditional chemotherapy is a primary treatment option for patients
where surgery is not feasible, and is an adjuvant therapy for those
with metastatic or high-risk cutaneous SCC (4). However, radiation is not suitable for
the distal part of the limbs, areas that difficult to heal and the
elderly (5), and traditional
chemotherapy has limited application due to the side effects that
it causes, including erosion, ulcer and erythema. Cryotherapy and
laser treatment are additionally used as for SCC in the clinical
setting; however, they have largely been replaced by surgery due to
the high recurrence rate (6,7).
Photodynamic therapy has received more attention as an effective
treatment for SCC of the skin (8),
although the curative effect is difficult to compare with other
treatments. Therefore, novel treatments targeting SCC are of utmost
interest for patients and clinicians (9).
Curcumin (CUR), which is a phenolic pigment
extracted from the Curcuma longa rhizome, is the main active
ingredient of turmeric, and has anti-inflammatory, antioxidant and
anticancer properties (10–13).
However, CUR is easily degraded in vitro and in vivo.
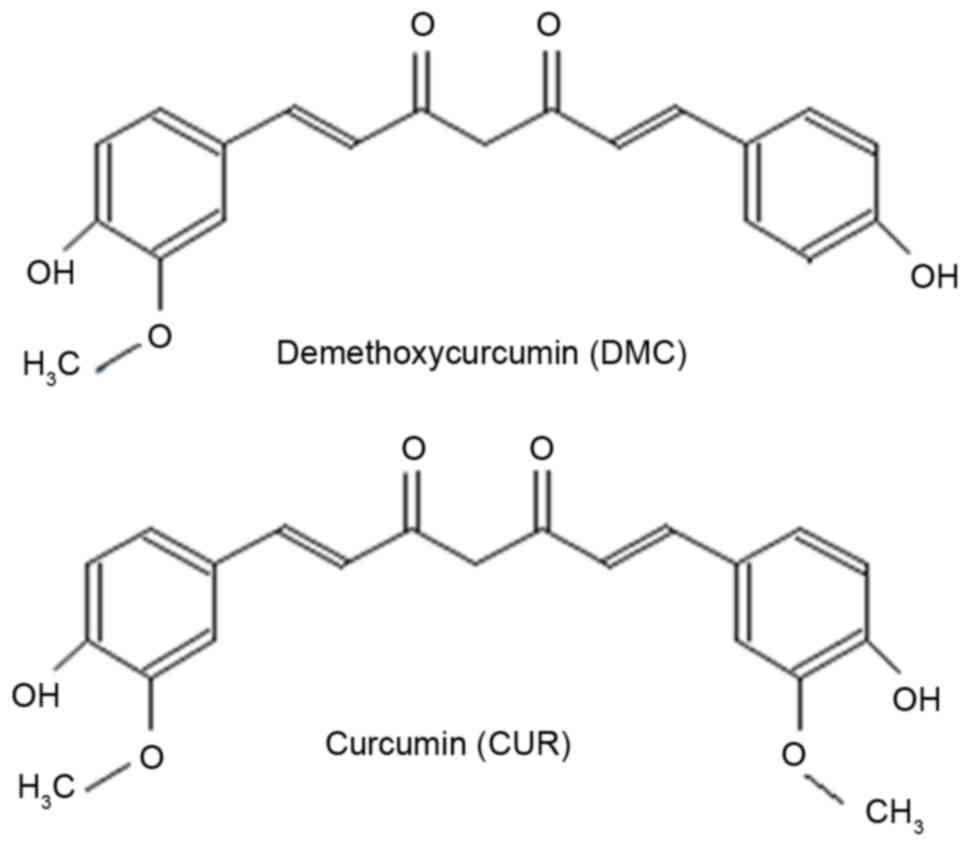
Demethoxycurcumin (DMC) is a derivative of CUR lacking a methoxy
group attached to a benzene ring (Fig.
1); it has similar biological properties to CUR, but is more
chemically stable (14).
Previous studies have demonstrated that DMC strongly
inhibits proliferation in numerous types of cancer, including
prostate cancer, kidney cancer and breast cancer cells (15–17).
However, the effects of DMC on skin cancer cells remain unknown. In
the present study, the effects of DMC treatment on the viability
and apoptosis of human skin squamous carcinoma A431 cells and human
keratinocyte HaCaT cells were examined. In addition, the molecular
mechanisms underlying these effects were explored. The present
findings provide a theoretical basis for the potential clinical
application of DMC in the treatment of skin cancer.
Materials and methods
Cell culture and reagents
A431 and HaCaT cells were purchased from the
Shanghai Cell Collection (Shanghai, China) and the Fuxiang
Institute of Biotechnology (Shanghai, China), respectively. Cells
were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) containing 10%
fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) and
1% penicillin and streptomycin, in a humidified incubator at 37°C
supplied with 5% CO2. Once cells reached 80% confluence,
they were passaged at a 1:3 ratio by dissociation with 0.25%
trypsin and 0.02% EDTA. Cells in the logarithmic growth phase were
used in all experiments. DMC (>99% purity) was obtained from the
YuanYe Institute of Biotechnology (Shanghai, China) and dissolved
in dimethyl sulfoxide (DMSO) to generate a 100 mmol/l stock
solution. The stock solution was stored at −20°C and freshly
diluted in complete culture medium prior to use. The final
concentration of DMSO applied to the cells was <0.1%. Rabbit
polyclonal primary antibodies against B-cell lymphoma 2 (Bcl-2;
cat. no. AB40415), Bcl-2-associated X protein (BAX; cat. no.
AB40636), caspase-3 (cat. no. AB42437), caspase-9 (cat. no.
AB32539) and cytochrome c (cat. no. AB133504) were purchased from
Biogot Technology Co., Ltd. (Nanjing, China). GAPDH (cat. no.
sc-32233) was used as a loading control (Santa Cruz Biotechnology,
Inc., Dallas, TX, USA). The goat-anti-rabbit secondary antibody
(cat. no. sc-362262) was purchased from Santa Cruz Biotechnology,
Inc.
Cell viability assay
The effects of DMC on A431 and HaCaT cell viability
were determined using a Cell Counting kit-8 (CCK-8) assay (Vicmed
Co., Ltd., Xuzhou, China). Cells were seeded in 96-well plates at a
density of 3×103 cells/well and incubated overnight in
DMEM containing 10% FBS. Cells were treated with various
concentrations of DMC (0, 5, 10, 20, 40 and 80 µМ) for 24, 48, 72
and 96 h, and were then incubated with CCK-8 reagent for 2 h at
room temperature. Absorbance at 450 nm was measured using a
microplate spectrophotometer (Thermo Labsystems, Santa Rosa, CA,
USA). Inhibition of viability was determined relative to the
control group measurement (untreated cells). The IC50
value was calculated using Graphpad Prism software (Graphpad
Software, Inc., San Diego, CA, USA). All experiments were performed
in triplicate.
Cell cycle distribution assay
Cells were seeded at 1×106 cells/well in
6-well plates, and allowed to grow for 24 h. Cells were treated in
triplicate with 0, 5, 10, 20, 40 and 80 µМ DMC for 48 h. Following
dissociation with 0.25% trypsin, cells were centrifuged at 2,000
rpm for 5 min at room temperature. The supernatant was discarded,
and pellets were washed once with PBS. Cell pellets were
resuspended in 500 µl 70% cold ethanol and fixed overnight at 4°C.
Following fixation, pellets were washed with PBS, 100 µl RNAse A
solution was added, and samples were incubated in a 37°C water bath
for 30 min. Subsequently, 400 µl propidium iodide (PI) (Nanjing
KeyGen Biotech Co., Ltd., Nanjing, China) was added, and samples
were incubated at 4°C in the dark for 30 min. Samples were analyzed
by flow cytometry (BD Biosciences, Franklin Lakes, NJ, USA), and
data were analyzed using FCS-express version 3 software (De Novo
software, Glendale, CA, USA). The experiment was performed in three
independent repeats.
Quantification of apoptosis by flow
cytometry
A431 and HaCaT cells were cultured in 6-well plates
for 12 h and then treated with 0, 5, 10, 20, 40 and 80 µM DMC for
48 h. Cells were dissociated, collected by centrifugation at 200.67
× g for 5 min at room temperature, and washed twice with PBS. Cells
(~1×106) were then resuspended in 500 µl binding buffer
containing 5 µl Annexin V and 5 µl PI reagents (Nanjing KeyGen
Biotech Co., Ltd.). Following a 15 min incubation at room
temperature in the dark, the samples were analyzed by flow
cytometry (BD Biosciences, Franklin Lakes, NJ, USA), and data were
analyzed using the FCS-express V3 software (de novo software,
Thornhill, Ontario, Canada).
Hoechst 33258 DNA staining
A431 and HaCaT cells were seeded in 6-well plates
and treated with 10, 20, 40 and 80 µM DMC for 48 h. Untreated cells
served as control. Following treatment, the medium was discarded,
the cells were washed twice with PBS, incubated with Hoechst 33258
(Nanjing KeyGen Biotech Co., Ltd.) for 5–10 min at room temperature
in the dark, washed twice with PBS, and finally observed under a
fluorescence microscope.
Western blotting
Adherent and floating cells were harvested at 200.67
× g for 5 min at room temperature and washed twice with ice-cold
PBS. Cells were homogenized in radioimmunoprecipitation assay lysis
buffer (Beyotime Institute of Biotechnology, Haimen, China)
containing 100 mM PMSF. Homogenates were centrifuged at 15,000 × g
at 4°C for 20 min and the supernatants were collected. Protein
concentrations were measured using the bicinchoninic acid method
(Nanjing KeyGen Biotech Co., Ltd.), using bovine serum albumin
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) to generate the
standard curve. A total of 100 µg protein of each sample was loaded
in SDS sample buffer and heat denatured at 100°C for 5 min. Protein
samples were then electrophoretically separated by 10–15% SDS-PAGE
and transferred onto nitrocellulose membranes (Promega Corporation,
Madison, WI, USA). Blocking was performed for 1 h at room
temperature in blocking buffer containing 5% non-fat dry milk.
Membranes were then incubated with primary antibodies at 4°C
overnight (1:1,000 dilution), followed by horseradish
peroxidase-conjugated secondary antibodies at room temperature for
2 h (1:5,000 dilution). Signals were detected with SuperSignal
enhanced chemiluminescence reagent (Pierce; Thermo Fisher
Scientific, Inc.). All experiments were performed in triplicate.
Densitometric analysis was performed on protein bands using
ImageQuant™ TL software version 8.1; Molecular Dynamics;
GE Healthcare Life Sciences, Chalfont, UK) (18).
Statistical analysis
Data were expressed as the mean ± standard
deviation. Statistical significance was determined by independent
samples t-test and one-way analysis of variance followed by the
Tukey's post-hoc test, as appropriate, using SPSS version 13.0 for
Windows (SPSS Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference. Statistical graphs
were drawn using GraphPad prism (Graph Pad Software, Inc.).
Results
DMC inhibits viability in A431 and
HaCaT cells
In order to investigate the effect of DMC treatment
on A431 and HaCaT cell viability, A431 and HaCaT cells were exposed
to various concentrations of DMC for 24, 48, 72 or 96 h, and then
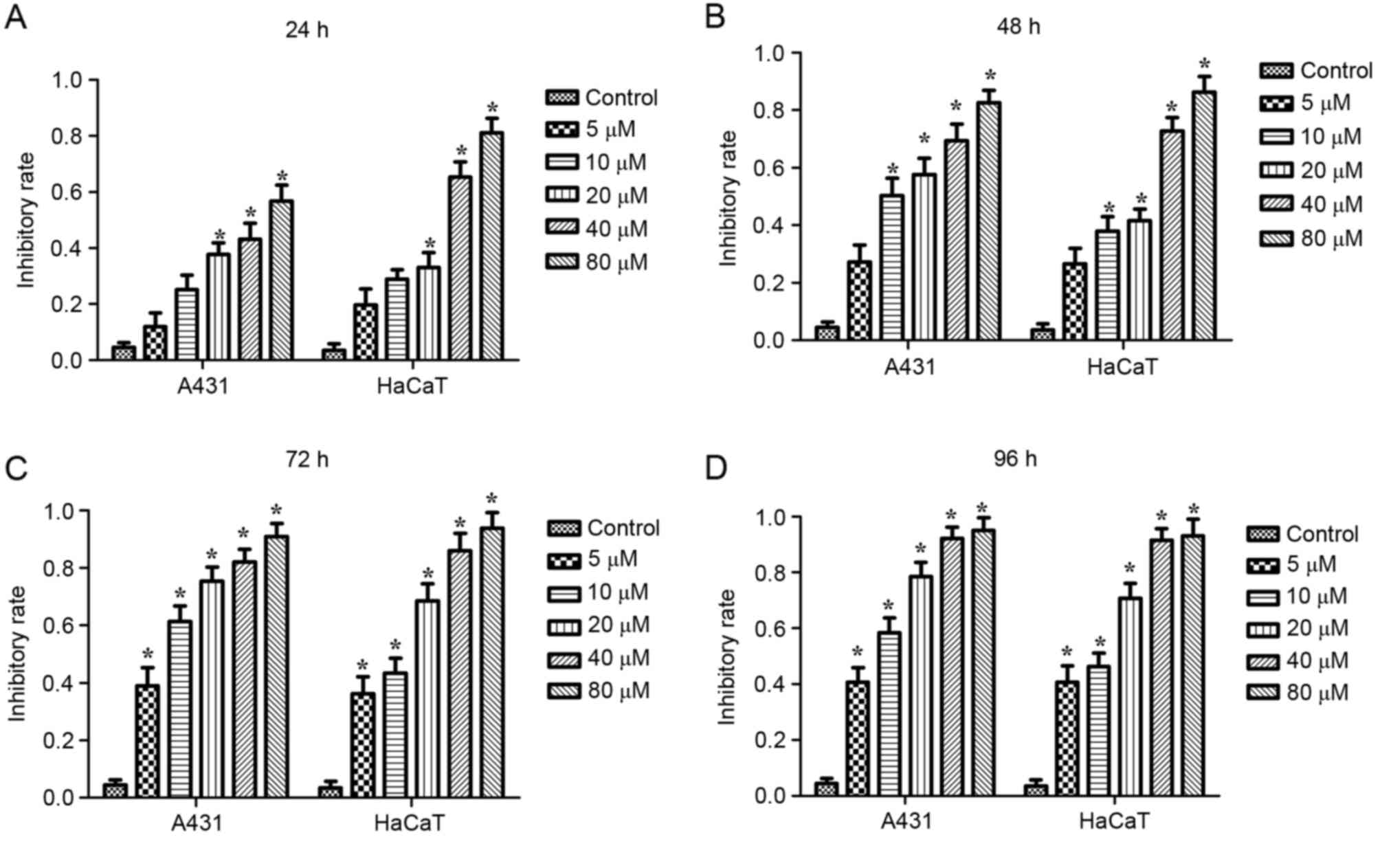
analyzed using a CCK-8 assay. As demonstrated in Fig. 2, the cell viability inhibitory rate
in DMC-treated cells was significantly increased compared with
control cells (P<0.05). The half maximal inhibitory
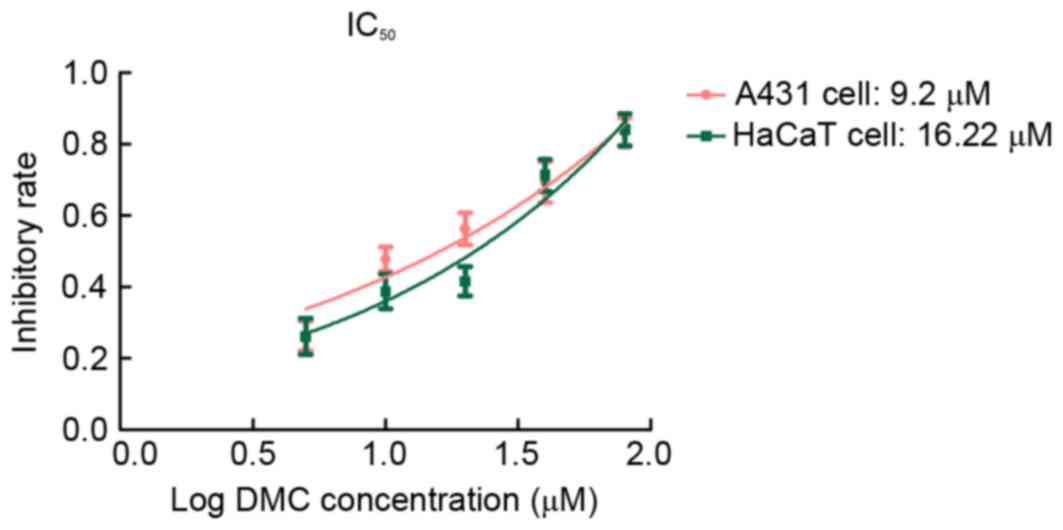
concentration values of DMC for A431 and HaCaT cells at 48 h were
9.2 and 16.22 µM, respectively (Fig.
3). These results indicated that DMC treatment decreased
viability of A431 and HaCaT cells in a dose-dependent manner.
DMC induces G2/M phase cell
cycle arrest in A431 and HaCaT cells
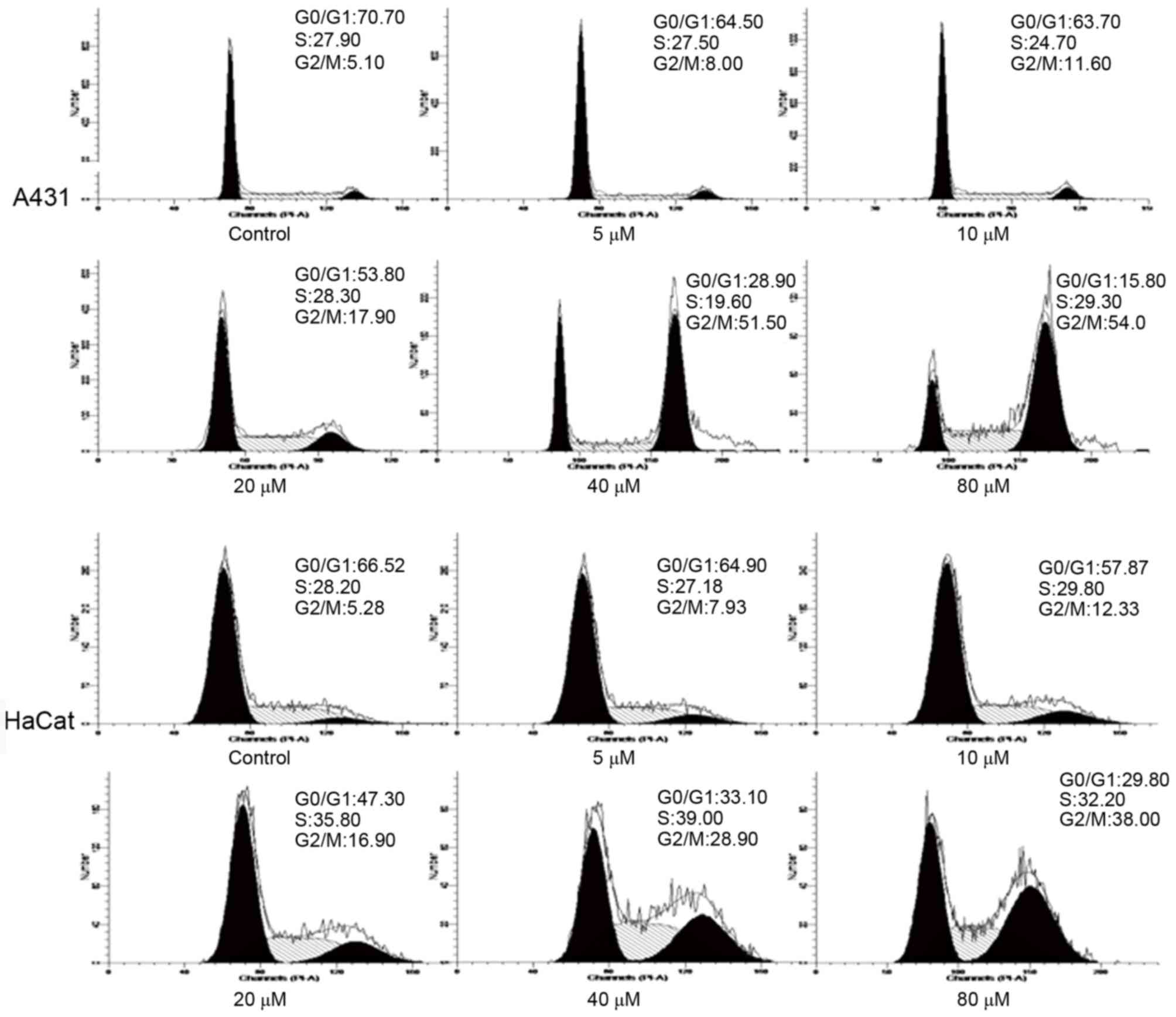
To further analyze the mechanism by which DMC
inhibits cell viability, A431 and HaCaT cells were treated with 5,
10, 20, 40 and 80 µM DMC for 48 h and then analyzed for cell cycle
phase distribution by flow cytometry. The results demonstrated that
the percentage of cells in the G0/G1 phase
was markedly decreased, whereas the percentage of cells in the G2/M
phase was markedly increased in the DMC-treated cells compared with
untreated cells (Fig. 4). These
results indicated that cell cycle arrest at G2/M may
contribute to the inhibitory effects of DMC on cell viability.
DMC induces apoptosis of A431 and
HaCaT cells
A431 and HaCaT cells were treated with 5, 10, 20, 40
and 80 µM DMC for 48 h and apoptosis was measured by Annexin V/PI
double staining and flow cytometric analysis. The apoptotic rate of
DMC-treated cells was significantly increased in a dose-dependent
manner compared with untreated cells (P<0.05; Fig. 5).
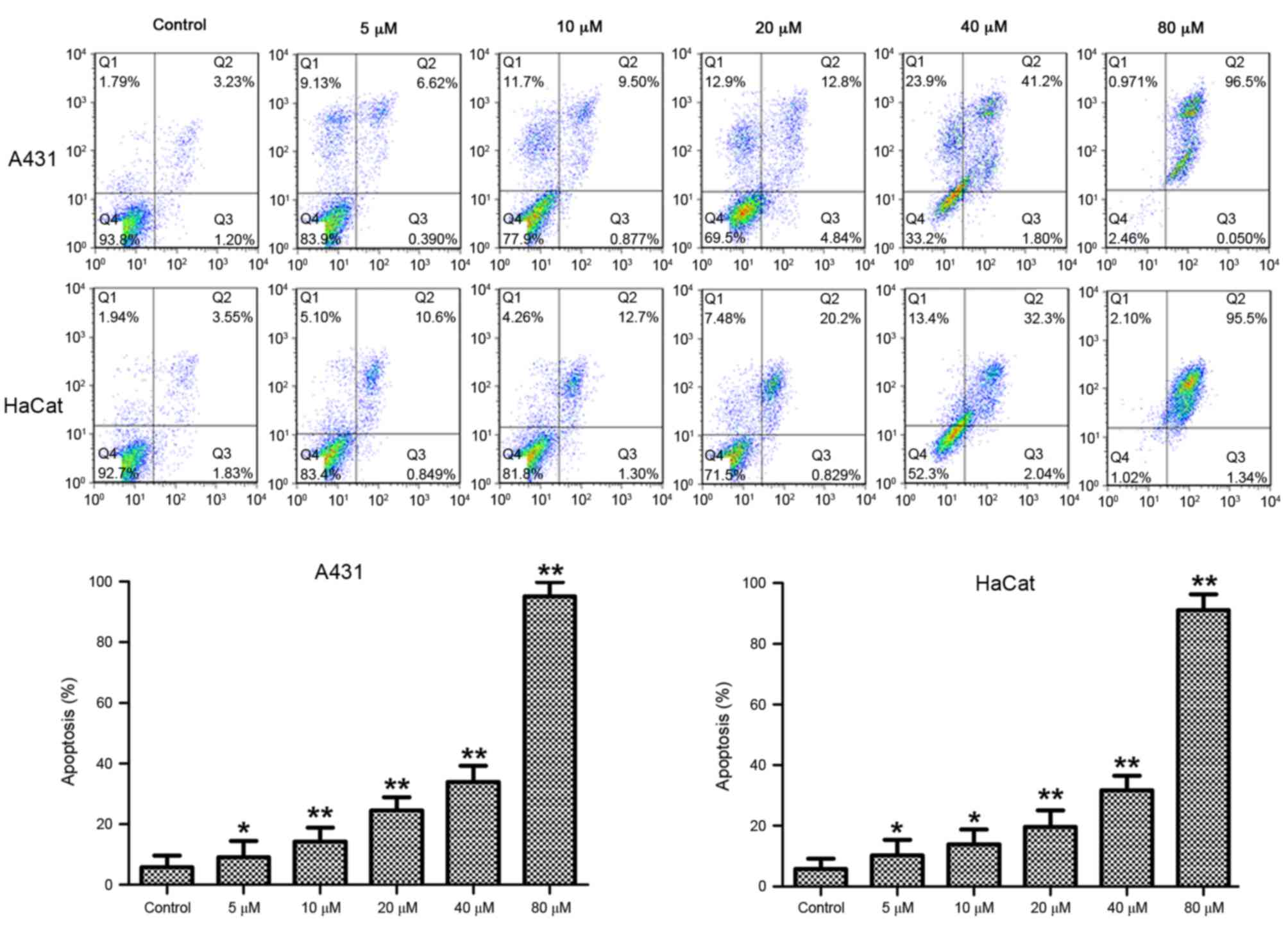 | Figure 5.Effects of DMC on apoptosis of A431
and HaCaT cells. Cells were treated with 5, 10, 20, 40 or 80 µM DMC
for 48 h, stained with Annexin V and PI, and analyzed by flow
cytometry. Untreated cells served as a control. Representative
plots are presented from at least three independent experiments.
Quantification is presented as mean percentage of apoptotic cells ±
standard deviation. *P<0.05 and **P<0.01 compared with
control. DMC, demethoxycurcumin; PI, propidium iodide; Q1, dead
cells; Q2, late apoptotic and necrotic cells; Q3, early apoptotic
cells; Q4, viable cells. |
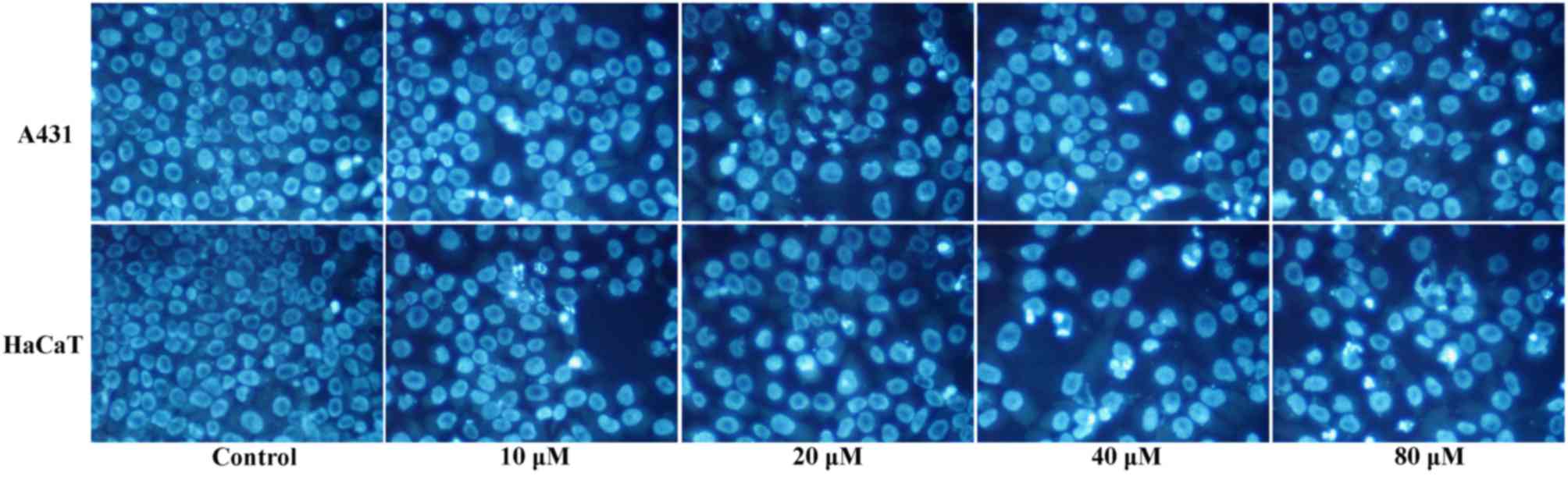
Morphological features of DMC-induced
apoptosis
To confirm apoptosis by cell morphological
observation, A431 and HaCaT cells were exposed to 0, 10, 20, 40 and
80 µM DMC for 48 h, stained with Hoechst 33258, and observed by
fluorescence microscopy. Control untreated cells displayed nuclei
with uniform staining (Fig. 6).
Conversely, shrinkage of the nuclei was observed in the cells
exposed to 20–80 µM DMC for 48 h (Fig.
6). The number of visibly apoptotic nuclei increased with
increasing concentrations of DMC, and various morphological
features of apoptosis were observed, including reduction of cell
numbers, nuclei with intensely bright staining and fragmented
nuclei (Fig. 6).
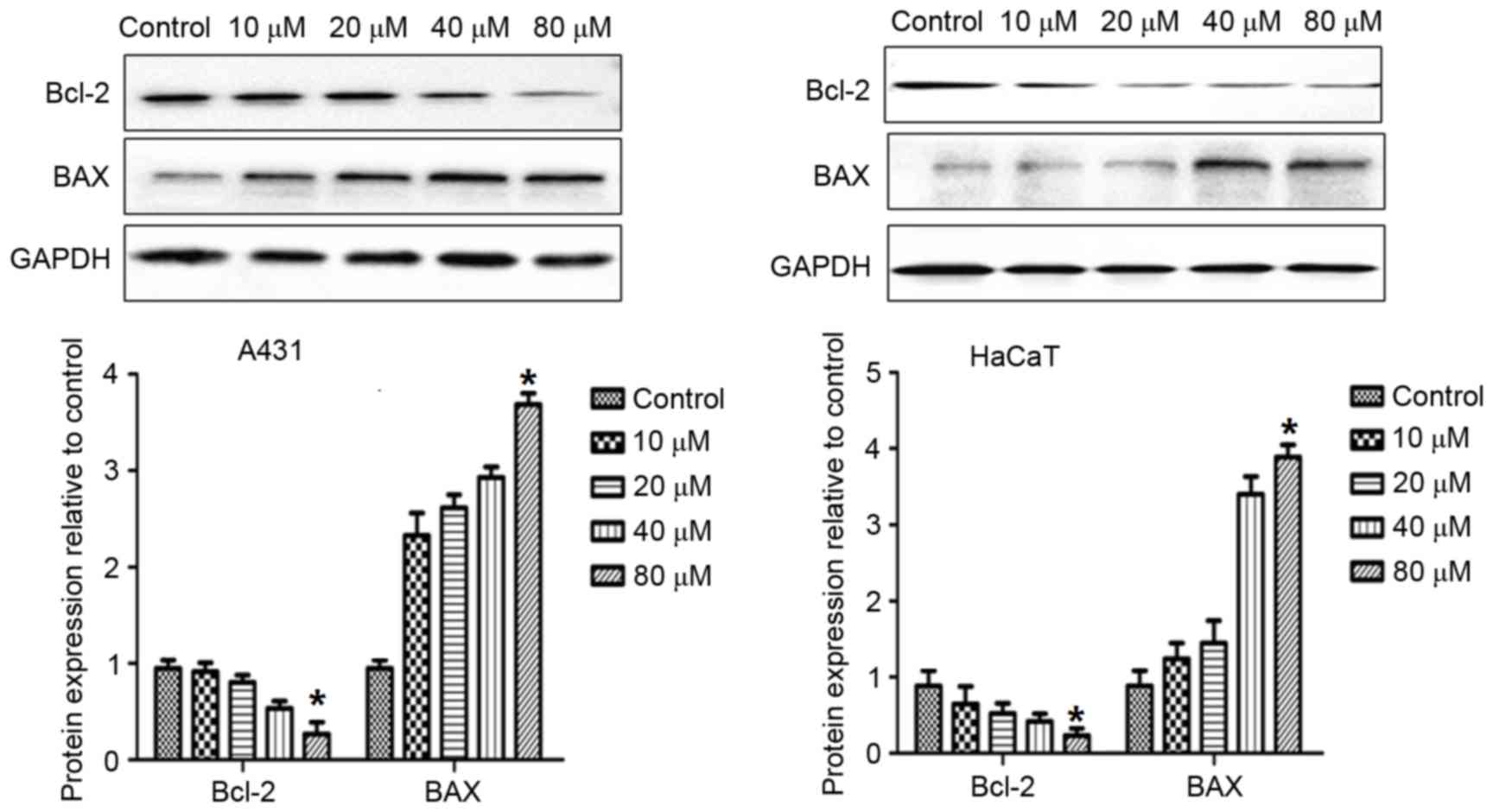
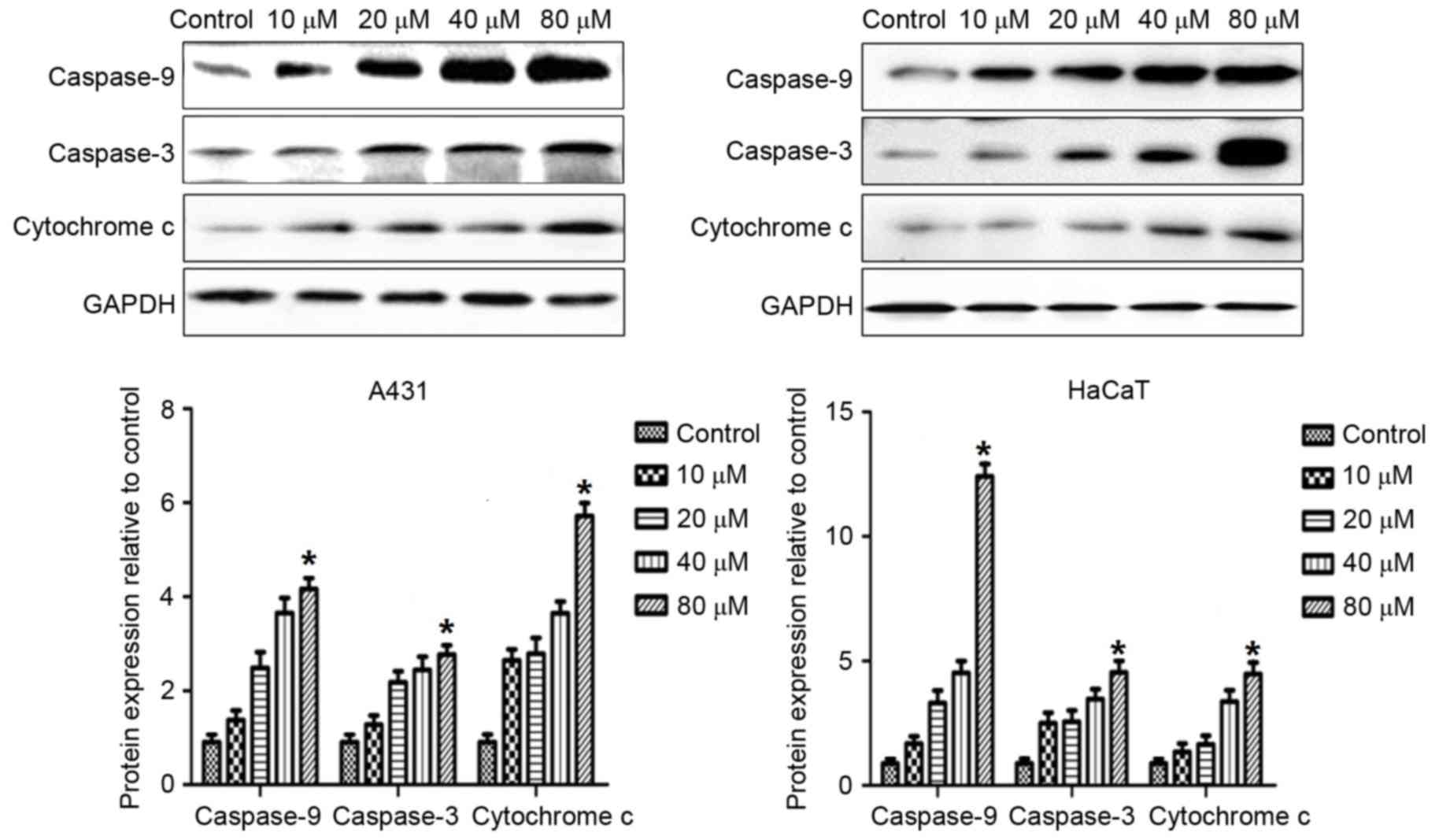
DMC increases expression of Bcl-2,
BAX, caspase-3, caspase-9 and cytochrome c proteins
In order to explore the mechanisms by which DMC
treatment may induce apoptosis, the effects of DMC on the protein
expression levels of various established apoptotic markers were
examined. A431 and HaCaT cells were treated with 10, 20, 40 and 80
µM DMC for 48 h and then analyzed by western blotting for protein
expression of Bcl-2, BAX, caspase-3, caspase-9 and cytochrome
c expression (Figs. 7 and
8). As demonstrated in Fig. 7, DMC treatment resulted in a
decrease in the protein expression levels of Bcl-2, and an increase
in BAX expression (Fig. 7). In
addition, DMC treatment caused an increase in the protein
expression levels of caspase-9, caspase-3 and cytochrome c
in a dose-dependent manner (Fig.
8).
Discussion
Previous studies have demonstrated that CUR exerts
inhibitory effects on various types of cancer, including bladder
cancer, prostate cancer, colon cancer, liver cancer and breast
cancer (19–22). DMC, which is a structural analogue
of CUR, also exhibits antitumor effects (14) and strongly inhibits proliferation
of prostate cancer cells (23).
The improved stability of DMC compared to CUR may significantly
prolong the time of action of DMC and extend its half-life,
suggesting that DMC may be an attractive compound to explore as an
anticancer agent.
It has previously been reported that CUR induces
cell cycle arrest in G2/M phase in liver cancer J5 cells
(24). In addition, DMC has been
demonstrated to induce G2/M cell cycle arrest in human
glioma U87 cells (25). In the
present study, the in vitro viability of A431 and HaCaT
cells treated with various concentrations of DMC was significantly
inhibited in a dose-dependent manner. Treatment with DMC reduced
the percentage of A431 and HaCaT cells in
G0/G1 phase in a dose-dependent manner,
whereas it increased the percentage of cells in S and
G2/M phases compared with untreated cells, indicating an
inhibition of mitosis. Evaluation of apoptosis by various methods,
including Annexin V/PI staining, Hoechst 33258 staining and
morphological observation, demonstrated that DMC treatment
increased apoptosis of A431 and HaCaT cells, which was consistent
with the DMC-mediated inhibition of cell viability.
The stimuli and pathways leading to cellular
apoptosis are diverse and complex, and regulation of apoptosis
involves proapoptotic and anti-apoptotic genes. The Bcl-2 family
proteins serve vital roles in the regulation of apoptosis (26–28).
The Bcl-2 family can be divided into two categories: pro- and
anti-apoptotic genes. These genes are key factors that determine if
a cell will commit to apoptosis or survival, and this decision is
determined by the ratio of BAX to Bcl-2 (26,29).
Bcl-2 family members alter mitochondrial membrane permeability and
regulate the release of cytochrome c from the mitochondria
to the cytoplasm. Cytochrome c then regulates cell apoptosis
through the transmission and amplification of apoptotic signals in
the cytoplasm. The close relationship between cytochrome c
and caspase family members has been detailed (30). Caspases are a specific kind of
protease (31). At present,
numerous caspases have been identified: Caspases-8, 9 and 10
initiate cellular apoptosis, whereas caspases-3, 6 and 7 are
involved in implementation of the apoptotic process. Cytochrome
c interacts with apoptotic peptidase activating factor 1 to
form an apoptotic complex (32,33).
Caspase-9 is recruited and activated by this apoptotic complex,
which then activates caspase-3. Caspase-3 is one of the most
important apoptotic execution factors in the caspase family, and
its activation is a sign of the irreversible commitment to
apoptosis (34). The activation of
caspases can in turn further promote the release of cytochrome
c from the mitochondria, resulting in an overall
amplification of the caspase cascade to promote apoptosis.
In the present study, treatment with DMC resulted in
a significant dose-dependent reduction in the protein expression
levels of Bcl-2, accompanied by a significant increase in BAX,
caspase-9, caspase-3 and cytochrome c. These results
indicated that DMC may regulate the release of cytochrome c
by downregulating Bcl-2 and upregulating BAX. The release of
cytochrome c can then activate downstream caspase-9 and
caspase-3, leading to an amplification of the caspase cascade
reaction, and ultimately to enhanced cell apoptosis.
In conclusion, treatment with DMC in the range of
10–80 µM inhibited the viability of A431 and HaCaT cells in a
dose-dependent manner. This was accompanied by cell cycle arrest in
G2/M phase and induction of apoptosis. These results
provide a putative mechanism of action for DMC regarding its
potential application as a therapeutic agent for the treatment of
skin malignancies.
Acknowledgements
The present study was supported by grants from the
Science and Technology Project of Xuzhou city (grant no. KC15SH010)
and Hubei Provincial Department of Education research project
(grant no. B2016126).
References
|
1
|
Prado R, Francis SO, Mason MN, Wing G,
Gamble RG and Dellavalle R: Nonmelanoma skin cancer
chemoprevention. Dermatol Surg. 37:1566–1578. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dréno B, Mansat E, Legoux B and Litoux P:
Skin cancer in transplant patients. Nephrol Dial Transplant.
13:1374–1379. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rajan N, Ryan J and Langtry JA: Squamous
cell carcinoma arising within a facial port-wine stain treated by
mohs micrographic surgical excision. Dermatol Surg. 32:864–866.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Smith TJ, Ryan LM, Douglass HO Jr, Haller
DG, Dayal Y, Kirkwood J, Tormey DC, Schutt AJ, Hinson J and Sischy
B: Combined chemoradiotherapy vs. radiotherapy alone for early
stage squamous cell carcinoma of the esophagus: A study of the
Eastern Cooperative Oncology Group. Int J Radiat Oncol Biol Phys.
42:269–276. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Moreno G, Chia AL, Lim A and Shumack S:
Therapeutic options for Bowen's disease. Australas J Dermatol.
48:1–8; quiz 9–10. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Giuliano EA, Johnson PJ, Delgado C, Pearce
JW and Moore CP: Local photodynamic therapy delays recurrence of
equine periocular squamous cell carcinoma compared to cryotherapy.
Vet Ophthalmol. 17:(Suppl 1). S37–S45. 2014. View Article : Google Scholar
|
|
7
|
Canis M, Ihler F, Martin A, Matthias C and
Steiner W: Transoral laser microsurgery for T1a glottic cancer:
Review of 404 cases. Head Neck. 37:889–895. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Szeimies RM, Morton CA, Sidoroff A and
Braathen LR: Photodynamic therapy for non-melanoina skin cancer.
Acta Derm Venereol. 85:483–490. 2005.PubMed/NCBI
|
|
9
|
Shimizu I, Cruz A, Chang KH and Dufresne
RG: Treatment of squamous cell carcinoma in situ: A review.
Dermatol Surg. 37:1394–1411. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fang J, Lu J and Holmgren A: Thioredoxin
reductase is irreversibly modified by curcumin: A novel molecular
mechanism for its anticancer activity. J Biol Chem.
280:52284–52290. 2005. View Article : Google Scholar
|
|
11
|
Jurenka JS: Anti-inflammatory properties
of curcumin, a major constituent of curcum longa: A review of
preclinical and clinical research. Altern Med Rev. 14:141–153.
2009.PubMed/NCBI
|
|
12
|
LoTempio MM, Veena MS, Steele HL,
Ramamurthy B, Ramalingam TS, Cohen AN, Chakrabarti R, Srivatsan ES
and Wang MB: Curcumin suppresses growth of head and neck squamous
cell carcinoma. Clin Cancer Res. 11:6994–7002. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gupta SC, Patchva S, Koh W and Aggarwal
BB: Discovery of Curcumin, a component of golden spice, and its
miraculous biological activities. Clin Exp Pharmacol Physiol.
39:283–299. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tamvakopoulos C, Dimas K, Sofianos ZD,
Hatziantoniou S, Han Z, Liu ZL, Wyche JH and Pantazis P: Metabolism
and anticancer activity of the curcumin analogue,
dimethoxycurcumin. Clin Cancer Res. 13:1269–1277. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin SS, Lai KC, Hsu SC, Yang JS, Kuo CL,
Lin JP, Ma YS, Wu CC and Chung JG: Curcumin inhibits the migration
and invasion of human A549 lung cancer cells through the inhibition
of matrix metalloproteinase-2 and-9 and vascular endothelial growth
factor (VEGF). Cancer Lett. 285:127–133. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen HW, Lee JY, Huang JY, Wang CC, Chen
WJ, Su SF, Huang CW, Ho CC, Chen JJ, Tsai MF, et al: Curcumin
inhibits lung cancer cell invasion and metastasis through the tumor
suppressor HLJ1. Cancer Res. 68:7428–7438. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ni X, Zhang A, Zhao Z, Shen Y and Wang S:
Demethoxycurcumin inhibits cell proliferation, migration and
invasion in prostate cancer cells. Oncol Rep. 28:85–90.
2012.PubMed/NCBI
|
|
18
|
Schneider CA, Rasband WS and Eliceiri KW:
NIH Image to ImageJ: 25 years of image analysis. Nat Methods.
9:671–675. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Teiten MH, Gaascht F, Cronauer M, Henry E,
Dicato M and Diederich M: Anti-proliferative potential of curcumin
in androgen-dependent prostate cancer cells occurs through
modulation of the wingless signaling pathway. Int J Oncol.
38:603–611. 2011.PubMed/NCBI
|
|
20
|
Lee YK, Park SY, Kim Ym and Park OJ:
Regulatory effect of the AmPK-COX-2 signaling pathway in
curcumin-induced apoptosis in HT-29 colon cancer cells. Ann N Y
Acad Sci. 1171:489–494. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yodkeeree S, Ampasavate C, Sung B,
Aggarwal BB and Limtrakul P: Demethoxycurcumin suppresses migration
and invasion of MDA-MB-231 human breast cancer cell line. Eur J
Pharmacol. 627:8–15. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sharma RA, Steward WP and Gescher AJ:
Pharmacokinetics and pharmacodynamics of curcumin. Adv Exp Med
Biol. 595:453–470. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ni X, Zhang A, Zhao Z, Shen Y and Wang S:
Demethoxycurcumin inhibits cell proliferation, migration and
invasion in prostate cancer cells. Oncol Rep. 28:85–90.
2012.PubMed/NCBI
|
|
24
|
Cheng CY, Lin YH and Su CC: Curcumin
inhibits the proliferation of human hepatocellular carcinoma J5
cells by inducing endoplasmic reticulum stress and mitochondrial
dysfunction. Int J Mol Med. 26:673–678. 2010.PubMed/NCBI
|
|
25
|
Luthra PM, Kumar R and Prakash A:
Demethoxycurcumin induces Bcl-2 mediated G2/M arrest and apoptosis
in human glioma U87 cells. Biochem Biophys Res Commun. 384:420–425.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liang CZ, Zhang JK, Shi Z, Liu B, Shen CQ
and Tao HM: Matrine induces caspase-depedent apoptosis in human
osteosarcoma cells intro and in vivo through the upregulation of
Bax and Fas/FasL and downregulation of Bcl-2. Cancer Chemother
Pharmacol. 69:317–331. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ghate NB, Hazra B, Sarkar R, Chaudhuri D
and Mandal N: Alteration of Bax/Bcl-2 ratio contributes to
Terminalia belerica-induced apoptosis in human lung and breast
carcinoma. In Vitro Cell Dev Biol Anim. 50:527–537. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zeng H, Kong X, Peng H, Chen Y, Cai S, Luo
H and Chen P: Apoptosis and Bcl-2 family proteins, taken to chronic
obstructive pulmonary disease. Eur Rev Med Pharmacol Sci.
16:711–727. 2012.PubMed/NCBI
|
|
29
|
Cory S, Huang DC and Adams JM: The Bcl-2
family: Roles in cell survival and oncogenesis. Oncogene.
22:8590–8607. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hilchie AL, Furlong SJ, Sutton K,
Richardson A, Robichaud MR, Giacomantonio CA, Ridgway ND and Hoskin
DW: Curcumin-induced apoptosis in PC3 prostate carcinoma cells is
caspase-independent and involves cellular ceramide accumulation and
damage to mitochondria. Nutr Cancer. 62:379–389. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Y, Zhang S, Geng JX and Hu XY: Curcumin
inhibits human non-small cell lung cancer A549 cell proliferation
through regulation of Bcl-2/Bax and cytochrome C. Asian Pac J
Cancer Prev. 14:4599–4602. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cain K, Brmton SB, Langlais C, Walker G,
Brown DG, Sun XM and Cohen GM: Apaf-1 oligomerizes into
biologically active approximately 700-kDa and inactive
approximately 1.4-MDa apoptosome complexes. J Biol Chem.
275:6067–6070. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Balasubramanian S and Eckert RL: Curcumin
suppresses AP1 transcription factor-dependent differentiation and
activates apoptosis in human epidermal keratinocytes. J Biol Chem.
282:6707–6715. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mazumder S, Plesea D and Almasan A:
Caspase-3 activation is acritical determinant of genotoxic
stress-induced apoptosis. Methods Mol Biol. 414:13–21.
2008.PubMed/NCBI
|