Introduction
It is well accepted that exercise is beneficial for
impaired cardiac function and can improve survival, decrease
complications, and abrogate increases in muscle sympathetic nerve
activity (1,2). Moderate-intensity endurance exercise
is also capable of leading to cardiac hypertrophy in humans, which
may worsen cardiac function. Furthermore, patients with impaired
cardiac function may have decreased capacity for exercise and
increased risk for exercise-related mortality (3).
The local rennin angiotensin system has been
identified in the brain and cardiovascular system. Angiotensin II
(AngII) serves an important role in the pathology and physiology of
cardiac hypertrophy, myocardial injury, hypertension and
atherosclerosis. By binding to the AngII type 1 receptor
(AT1R), AngII activates a G-protein-coupled receptor,
leading to activation of the gene activity and mRNA synthesis,
stimulating the protein synthesis and cell growth, and then
transforming physical cardiac hypertrophy to pathological cardiac
hypertrophy (4). In the
pathological alterations of cardiac hypertrophy, the number and
affinity characteristics of AT1R are markedly changed.
However, to the best of our knowledge, no studies have examined the
changes of the cardiac pathological hypertrophy signaling pathway
(AngII/AT1R/Gq signaling pathway) on the condition of
exercise-induced cardiac hypertrophy. Therefore, the effects of
moderate-intensity exercise on the expression and distribution of
cardiac local AngII and AT1R were examined in the
present study.
Materials and methods
Animals
A total of 40 male Sprague-Dawley rats (age, 3
months; weight, 180–210 g) were purchased from Sichuan Provincial
People's Hospital (no. SCXK2008; specific-pathogen-free). The rats
were divided into the endurance exercise group (E group; n=20) and
control group (C group; n=20). Each group included two sub-groups:
Morphological detection group (C1/E1group; n=10) and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
detection group (C2/E2 group; n=10). The rats were housed under
specific pathogen-free conditions in an animal facility and given a
pelleted regular rodent diet (standard laboratory animal medicine)
and water ad libitum. The room temperature was 22–25°C,
humidity was 45±10%, and a 12-h light-dark diurnal cycle was used.
The current study was approved by the Animal Care and Use Committee
of the Sichuan Provincial People's Hospital (Sichuan, China).
Exercise protocol
The endurance exercise group was trained on an
animal treadmill. The animals underwent adaptive training for 1
week, the speed of animal treadmill was 10 m/min. The endurance
exercise training protocol used was adapted from that of the
training protocol from Fenning et al (5), combined with the control method of
maximal oxygen uptake of Wisløff et al (6). The exercise intensity remained at
60–65%VO2max. The exercise training lasted for 8 weeks, 6 days a
week. The exercise duration was gradually increased from 15 to 50
min. The speed was gradually increased from 10 to 35 m/mins.
Immunohistochemistry
All rats were sacrificed at 24 h following the last
session of exercise. The rats were anesthetized by abdominal
administration of pentobarbital sodium (4 mg/100 g weight).
Subsequent to opening the chest and removing out the whole heart,
heart samples of groups C1 and E1 were embedded in paraffin and cut
with a standard microtome. Histological sections were cut in 5-µm
thick slices and analyzed by immunohistochemical staining for AngII
and AT1R expression (Epitomics; Abcam, Cambridge, UK). Briefly,
sections were de-waxed and rehydrated. Peroxidase ABC-reagent and
3,3′-diaminobenzidine chromogenic substrate were applied. Treatment
with 0.3% H2O2 was used to block endogenous
peroxidase activity, for 15–20 min at 25°C, followed by washing
with PBS (0.01 mol/l) three times for 5 min. Complex digestive
solution was applied for 15 min to increase the permeability of the
tissues and cells, followed by washing with distilled water three
times for 5 min. Rabbit serum (cat. no. AR1010; Wuhan Boster
Biological Technology, Ltd., Wuhan, China) was applied and tissues
were incubated for 20 min at 37°C to reduce nonspecific background
staining. Afterwards, the slides were washed in Tris-buffered
saline and incubated with diaminobenzidine as substrate and
counterstained with hematoxylin-eosin. Negative controls without
primary antibody were included in each experiment to verify
antibody specificity. Primary antibody (1:200, mouse anti-human;
cat. no. sc-8431; Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
was incubated overnight at 4°C, followed by three washes (5 mins)
with PBS (0.01 mol). Biotin-labeled secondary antibody (goat
anti-mouse; 1:200; cat. no. SC-1022; Santa Cruz Biotechnology,
Inc.) was incubated for 1 h at 37°C, followed by four washes with
PBS (0.01 mol).
The sections were observed and images were acquired
under alight microscope camera. Using the image analysis software
(Image-ProPlus; version 6; Media Cybernetics, Inc., Rockville, MD,
USA), the image analysis parameters were automatically generated. A
total of 20 slices were selected for each group, and 5 fields of
vision were selected to analyze the images. The system measurements
used were as follows: i) The average optical density of the
positive reaction region in each field of vision; ii) the total
area of the test positive region.
RT-qPCR analysis
Total RNA was extracted from heart tissue according
to the protocol of the TRIzol reagent (cat. no. BA0290; Wuhan
Boster Biological Technology, Ltd.). DNA was extracted according to
the protocol of AxyPrep™ Multisource Genomic DNA Miniprepkit
(Axygen; Corning Incorporated, Corning, NY, USA). The quality and
concentration of the nucleic acids were measured using Gene Quant
pro (Biochrom Ltd., Cambridge, UK; version 80-2115-04). cDNA was
synthesized as described previously (7).
AngII and AT1R were amplified by the PCR
reaction in a 25 µl reaction volume containing 10XPCR buffer, 2.5
mM dNTPs, 50 mM MgCl2, 10 µM forward and 10 µM reverse
primers, 0.2 U Platinum® Taq and 10 mg DNA. The PCR
cycling conditions were as follows: 94°C for 5 min; 45 cycles of
94°C for 30 sec, 57°C for 30 sec, 72°C for 1.5 min; followed by
70°C for 10 min. RT-qPCR was performed using the Applied Biosystems
StepOne™ Real-Time PCR detection system. PCR was conducted in a 25
µl reaction volume containing 10 µl SYBR Green Supermix (Bio-Rad
Laboratories, Inc., Hercules, CA, USA), 1 µl each of the specific
primer mix (100 nM as final concentration of each primer), 2 µl
cDNA and 6 µl nuclease-free water (8). The primer sequences and details of
the target genes are presented in Table I.
 | Table I.Sequence and details of target
genes. |
Table I.
Sequence and details of target
genes.
| Genes | Forward primer | Reverse primer | Tm (°C) |
|---|
| Angiotensin II |
GCAGTCCGTTCCTGTTCG |
CTCTGACGCTGGCCTTGT | 56 |
| Angiotensin II type 1
receptor |
AAAATGAGCACGCTTTCTTACCGGC |
GAACCTGTCACTCCACCT | 60 |
| β-actin |
ATGGAGCCACCGATCCACA |
CATCCGTAAAGACCTCTATGCCAAC | 56 |
To obtain the RT-qPCR detection limits for AngII and
AT1R, a standard curve based on the quantification cycle
(Cq) for a dilution series of pure AngII and AT1R (1,
0.5, 1×10−1, 1×10−2, 1×10−3, and
1×10−4 ng/µl, diluted with 0.1X
Tris-ethylenediaminetetraacetic acid buffer) was constructed and
triplicate qPCR was performed. In order to quantitatively detect
AngII and AT1R in the standard panel samples, another
standard curve based on the Cq (8)
for the same dilution series.
Statistical analysis
Data were expressed as mean ± standard deviation.
The group differences were analyzed using the independent-sample
t-test. All data analyses were performed using the SPSS statistical
package (version 17.0; SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
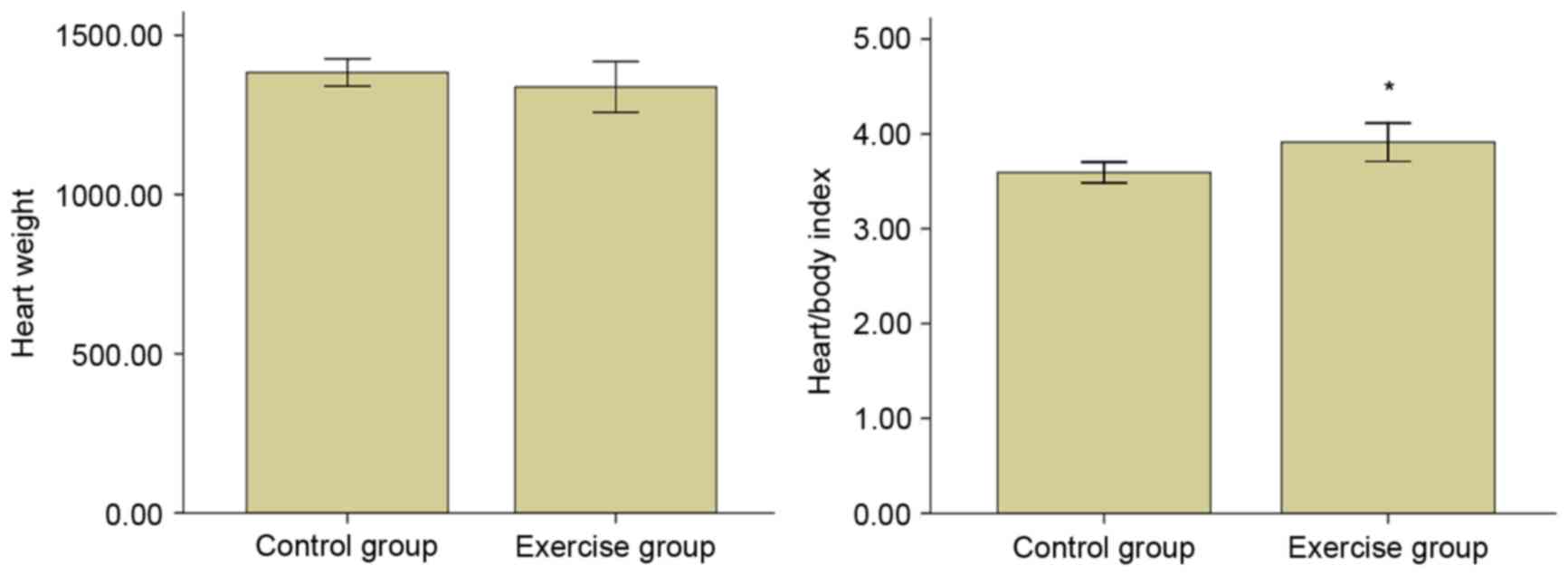
The heart weight and heart/body index
of each group
As presented in Fig.
1, no significant differences were identified in the heart
weight between the two groups. However, the heart/weight index
increased significantly (P<0.05). This indicates that the
moderate-intensity endurance training resulted in cardiac
hypertrophy.
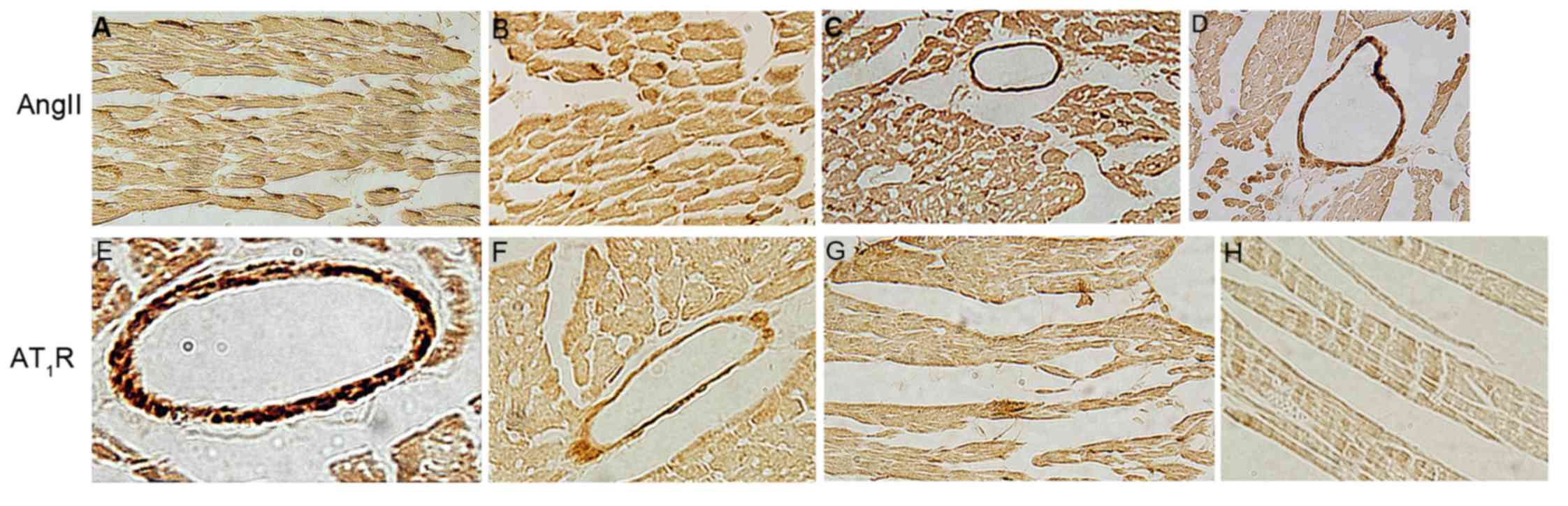
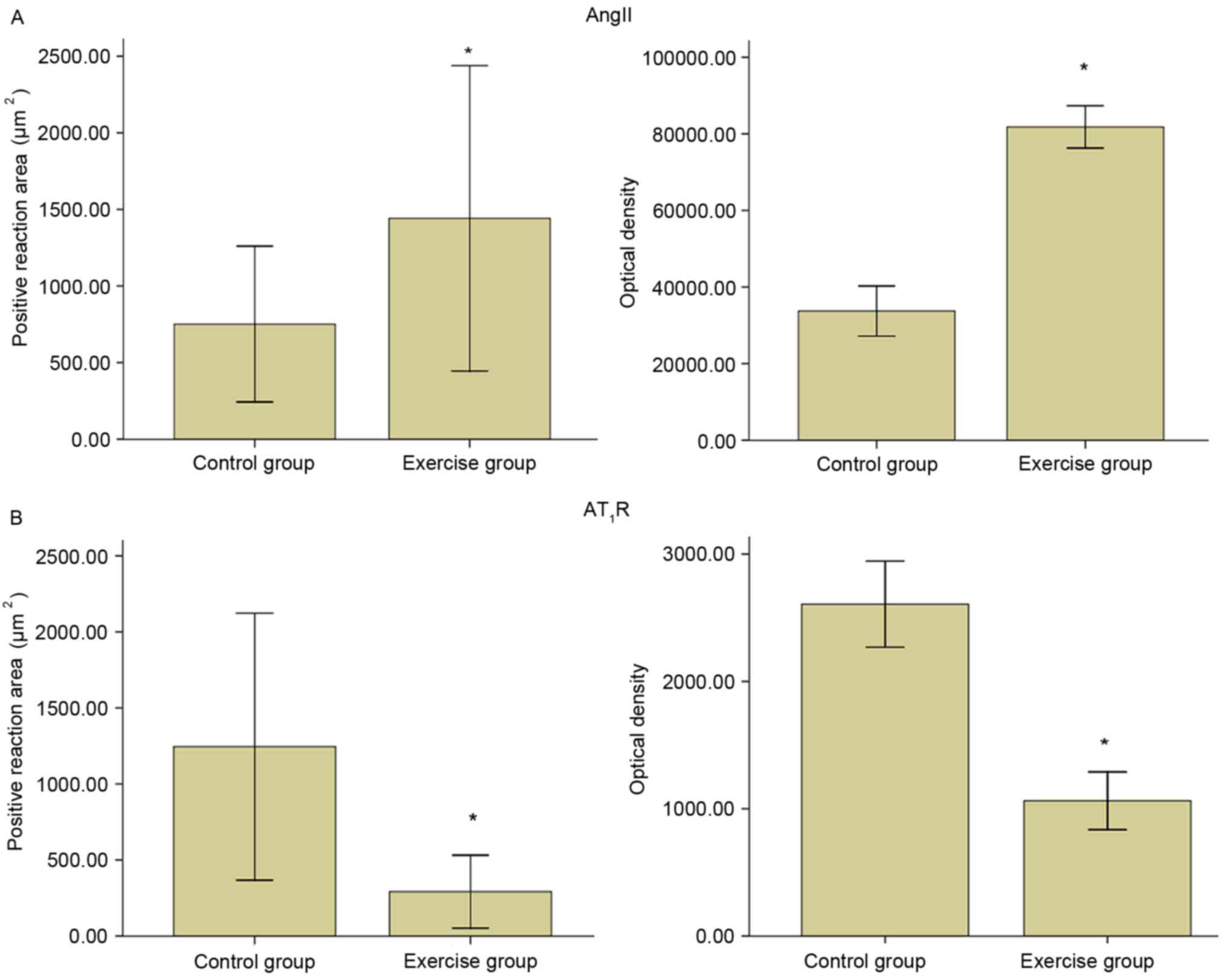
Immunohistochemical observation of
cardiac AngII
The AngII of control group was primarily expressed
in the myocardial cell cytoplasm and cell membrane, less in the
coronary vessel smooth muscle (Figs.
2 and 3). The immune reaction
of AngII of exercise group in myocardial cell and coronary vessel
smooth muscle enhanced, the staining density increased (Figs. 2 and 3).
Immunohistochemical observation of
cardiac AT1R
The AT1R of group C1 was primarily expressed in the
coronary vessel wall smooth muscle cells, and to a lesser extent in
the myocardial cell membrane (Figs.
2 and 3). The immune reaction
of AT1R of group E1 in myocardial cell and coronary vessel smooth
muscle decreased, thus the staining density reduced (Figs. 2 and 3).
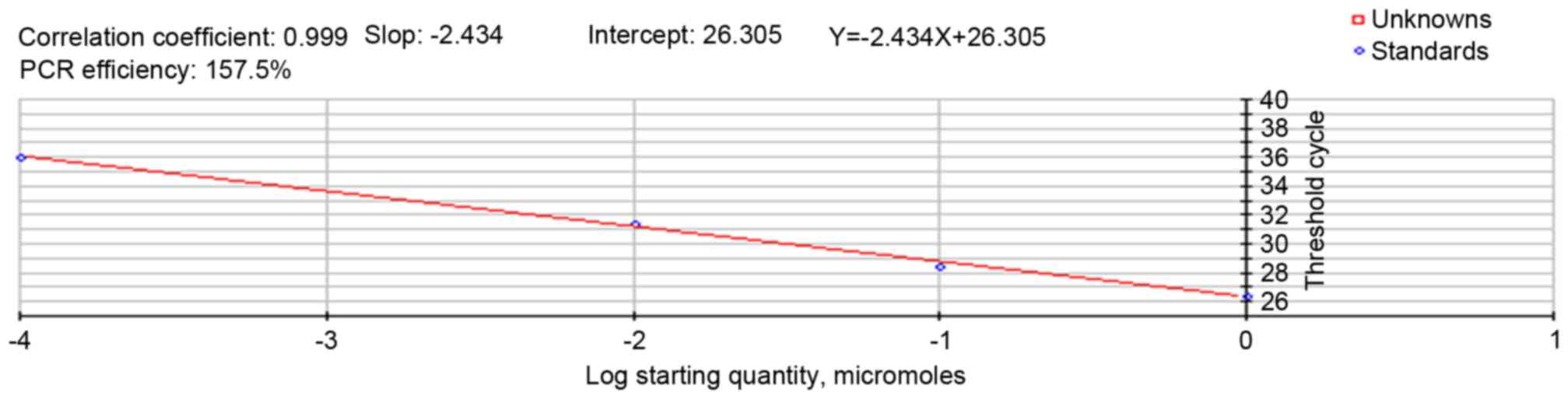
Effects of moderate-intensity
endurance exercise on Angiotensin II mRNA expression
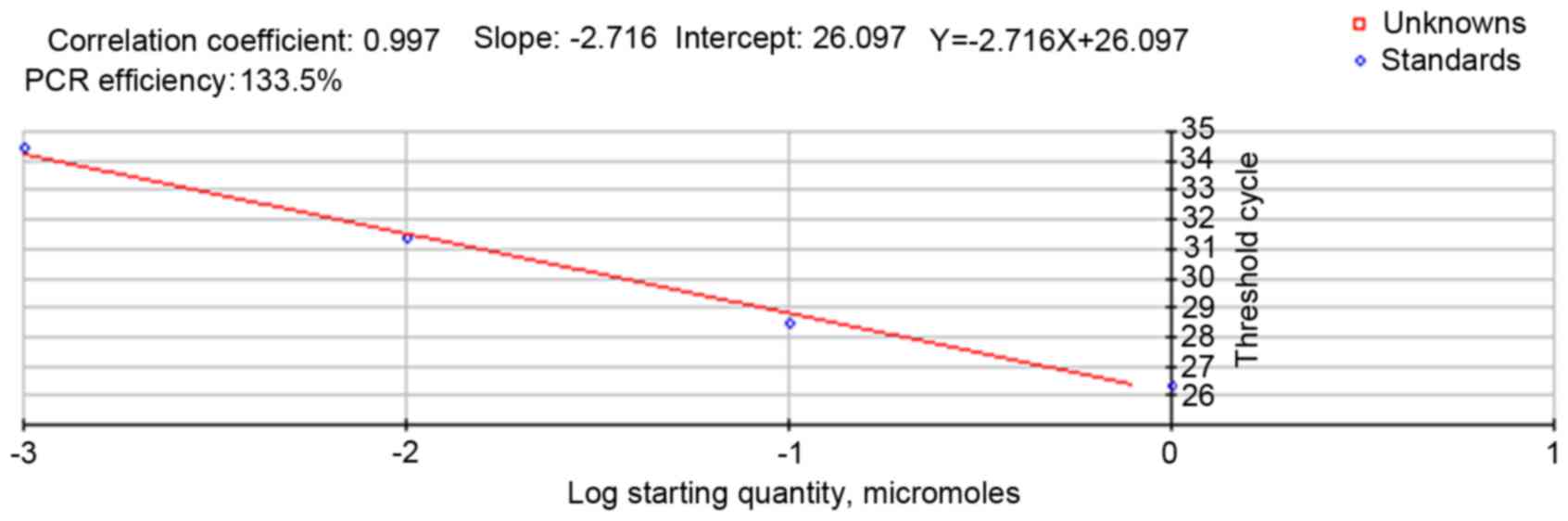
The standard curve of RT-qPCR for the AngII gene and
its regression equation is presented in Fig. 4, which indicated that the fragment
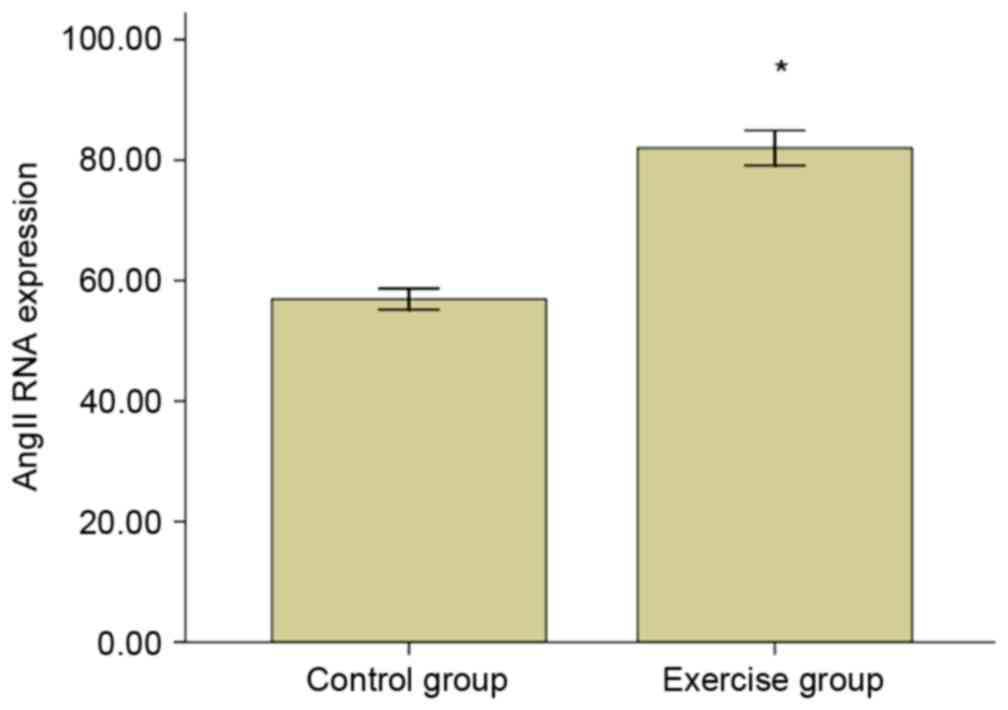
had high specificity. The AngII mRNA relative expression level is
presented in Fig. 5, demonstrating
that the expression for AngII mRNA of the exercise group was
increased significantly when compared with the control group
(P<0.05).
Effect of moderate-intensity endurance
exercise on cardiac AT1R mRNA expression
The standard curve of RT-qPCR for the AT1R gene and
its regression equation is presented in Fig. 6, indicating that the fragment had
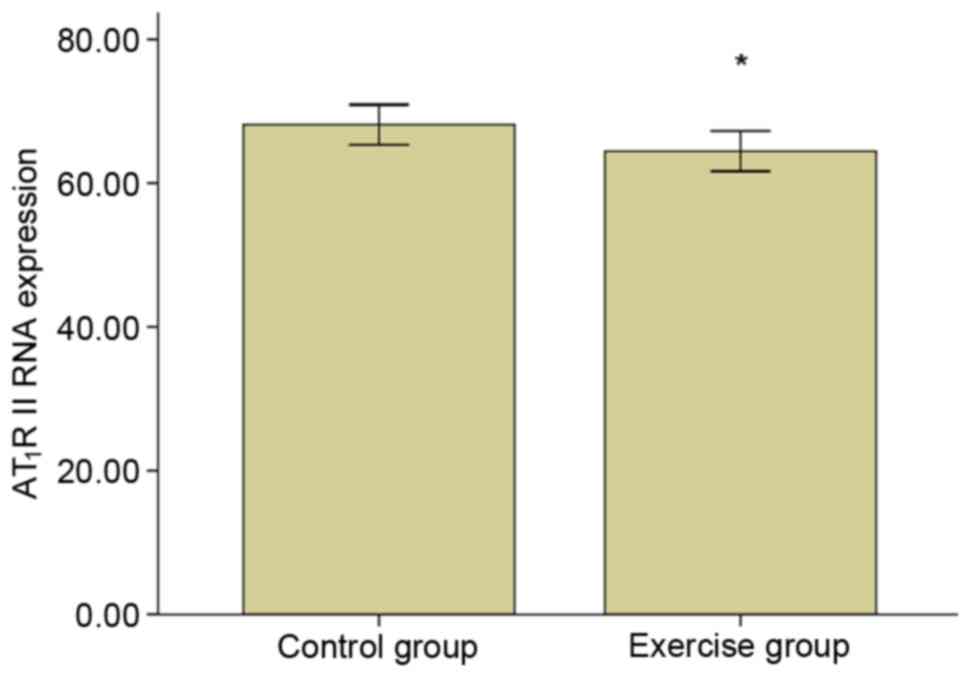
high specificity. The AT1R mRNA relative expression level is
exhibited in Fig. 7, which
demonstrated that the expression for AT1R II mRNA in the exercise
group was significantly decreased compared with the control group
(P<0.05).
Discussion
Following the 8-week endurance training, the body
weight of E group was significantly reduced compared with the C
group. The heart weight/body weight coefficient of group E
increased significantly compared with the C group. These
observations indicated that the moderate intensity endurance
exercise could cause endurance exercise-induced cardiac
hypertrophy. Histological analysis demonstrated that the myocardial
blood and oxygen supply was not impaired without pathological
cardiac hypertrophy.
Subsequent to the 8-week endurance exercise, the
immunohistochemistry staining of AngII was observed in cardiac
myocytes and coronary vascular wall muscle, while the positive
brown staining of myocardial cells was predominantly distributed in
the cytoplasm and cell membrane, and less frequently in the cell
membrane of the smooth muscle of coronary vessel wall. The
expression of cardiac local AngII mRNA increased significantly.
These results indicate that AngII is predominantly secreted by
cardiac myocytes and coronary vascular smooth muscle cells, and
moderate endurance exercise can improve the content of AngII in the
heart. Cardiac hypertrophy is a compensatory mechanism in case of
chronic long-term overload, and the elevated expression of AngII
serves an important role in the pathological myocardial hypertrophy
(9). The results demonstrated that
the 8-week moderate intensity endurance training led to cardiac
hypertrophy, histological examination exhibited no pathological
changes, indicating that AngII increase, which was stimulated by
moderate endurance exercise, is a benign adaptation to movement.
When the exercise intensity was increased, the volume load and
pressure load will be larger. Myocardial cells were hypertrophied
and the cardiac local AngII content and
angiotensin-converting-enzyme activity increases, which suggests
that there are correlations between the secretion of cardiac local
AngII and the training load (10);
the positive correlation suggests that under the condition of
overload exercise stimulation, the heart will be transformed from a
physiological cardiac hypertrophy to decompensate pathological
hypertrophy, and the changes of cardiac local AngII may be
important factors.
The AngII/AT1R signaling pathway is necessary for
cardiac renin-angiotensin system function, and the biological
function of local AngII is implemented by AT1R (11). AT1R undergoes rapid
internalization and desensitization when stimulated by AngII; on
the one hand, it can stimulate endothelial cells to produce
endothelin, contract the coronary artery, decrease myocardial
capillary density and aggravate myocardial ischemic injury
(12). In addition,
AT1R can couple with Gq proteins, activate phospholipase
C-β, and hydrolysis the phosphatidylinositol biphosphate to
diacylglyceroland inositol triphosphate (13). Subsequently, it serves an important
role in the myocardial cell apoptosis, necrosis, and cardiac
pathological hypertrophy. When the expression and characteristics
of AT1R change, simultaneously, the cardiac endogenous
active peptides that can induce cardiac hypertrophy (e.g. atrial
natriuretic factor or insulin like growth factor 1) will
correspondingly change (14). This
indicates that the alteration of the activity of the receptor is an
important for AngII to induce cardiac hypertrophy, the change of
the quantity of AngII does not appear to be an important factor.
The experimental results indicated that following eight weeks of
moderate intensity endurance exercise, the cardiac local
AT1R mRNA expression of the rats was significantly
reduced. Immunohistochemical testing indicated that moderate
intensity endurance training, the immune positive reaction and
optical density of AT1R in all levels of coronary
vascular smooth muscle and cardiac cells were markedly reduced.
Therefore, it is suggested that after eight weeks of moderate
intensity endurance training, heart local AT1R
expression is significantly reduced. It is hypothesized that the
main reason for AT1R downregulation may be that AngII
has a positive moderating effect on the myocardium, it can induce
cardiac hypertrophy, and the AT1R distribution gradually
reduces. This is the protective mechanism to avoid producing
pathological cardiac hypertrophy changes. The receptor distribution
reduction in coronary vascular smooth muscle can prevent coronary
excessive vasoconstriction; prevent cardiac ischemia and hypoxia,
so as to ensure the energy metabolism required for the heart to
function. In addition, moderate intensity endurance exercise
induces increased AngII feedback, which can stimulate
AT1R to AT2R transformation, which has a
contrasting effect on cell growth, proliferation, vascular tension
and vasopressin release (15); or
is decomposed by angiotensin converting enzyme 2 to Ang(1–7),
which leads to indirect cardioprotective effects. It is suggested
that after the 8-weeks exercise, upregulation of cardiac local
AngII and downregulation of AT1R is protective towards
the heart, which can prevent transforming pathological cardiac
hypertrophy and coronary vascular excessive vasoconstriction, and
is also important in maintaining the cardiac load demand and energy
metabolism under the physiological state (16). In the current study, the cardiac
AngII expression was increased after the 8-week moderate intensity
endurance training, which indicated that cardiac AngII is involved
in the heart physiological function via autocrine, paracrine and
intracrine mechanisms. It is suggested that AngII may participate
in the physiological functions of other organs via blood
circulation. Cardiac hypertrophy was induced when the rat cardiac
AT1R expression was downregulated, which suggests that
AngII through the AT1R signaling pathway may not serve a
major role in endurance training-induced cardiac hypertrophy.
Meanwhile, there is a significant difference between the
downregulation of AT1R expression and the upregulation
of AT1R expression under pathological cardiac
hypertrophy, which indicates that receptor number and/or affinity
changes are the key factors for physiological cardiac hypertrophy
becoming pathological cardiac hypertrophy, the expression change of
AngII itself does not lead to the characteristics of cardiac
hypertrophy.
AngII was predominantly distributed in the cell
cytoplasm and cell membrane, and rarely in coronary vascular wall
smooth muscle cells. AT1R was predominantly distributed
in the coronary vessel wall smooth muscle cells, however was rarely
present in cardiac cells. Following the 8-week endurance training
period, the AngII expression was increased and the AT1R
expression was decreased. AT1R may expand the coronary
artery, thereby increasing coronary blood flow and ensuring the
energy supply of the heart during exercise. The expression changes
in AngII do not reflect the character of cardiac hypertrophy. The
exercise-induced change in the expression of AngII and
AT1R may be a protective mechanism to avoid cardiac
pathological hypertrophy.
Acknowledgements
The current study was supported by grants from the
Fundamental Research Funds for the Central Universities, China
(grant no. 12JCY07), Chengdu University Funding (grant no.
2012ZJZ01) and the 2013 Annual Research Projects from Education
Department of Sichuan Province (grant no. 13ZB0340).
References
|
1
|
Haack KK, Engler CW, Papoutsi E, Pipinos
II, Patel KP and Zucker IH: Parallel changes in neuronal AT1R and
GRK5 expression following exercise training in heart failure.
Hypertension. 60:354–361. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rajani R, Rimington H and Chambers JB:
Treadmill exercise in apparently asymptomatic patients with
moderate or severe aortic stenosis: Relationship between cardiac
index and revealed symptoms. Heart. 96:689–695. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lou H, Danelisen I and Singal PK:
Involvement of mitogen-activated protein kinases in
adriamycin-induced cardiomyopathy. Am J Physiol Heart Circ Physiol.
288:H1925–H1930. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Haudek SB, Cheng J, Du J, Wang Y,
Hermosillo-Rodriguez J, Trial J, Taffet GE and Entman ML: Monocytic
fibroblast precursors mediate fibrosis in angiotensin-II-induced
cardiac hypertrophy. J Mol Cell Cardiol. 49:499–507. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fenning A, Harrison G, Dwyer D, Rose'Meyer
R and Brown L: Cardiac adaptation to endurance exercise in rats.
Mol Cell Biochem. 251:51–59. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wisløff U, Helgerud J, Kemi OJ and
Ellingsen O: Intensity-controlled treadmill running in rats: VO(2
max) and cardiac hypertrophy. Am J Physiol Heart Circ Physiol.
280:H1301–H1310. 2001.PubMed/NCBI
|
|
7
|
Xin LI: Effect of moderate-intensity
endurance exercise on the expression of cardiac signaling pathway.
East China Normal Univ. 2011.
|
|
8
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kohli S, Ahuja S and Rani V: Transcription
factors in heart: Promising therapeutic targets in cardiac
hypertrophy. Curr Cardiol Rev. 7:262–271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Belardinelli R: Exercise training in
chronic heart failure: How to harmonize oxidative stress,
sympathetic outflow, and angiotensin II. Circulation.
115:3042–3044. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang GX, Ohmori K, Nagai Y, Fujisawa Y,
Nishiyama A, Abe Y and Kimura S: Role of AT1 receptor in
isoproterenol-induced cardiac hypertrophy and oxidative stress in
mice. J Mol Cell Cardiol. 42:804–811. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jugdutt BI and Sawicki G: AT1 receptor
blockade alters metabolic, functional and structural proteins after
reperfused myocardial infarction: Detection using proteomics. Mol
Cell Biochem. 263:179–188. 2004. View Article : Google Scholar
|
|
13
|
Bai H, Wu LL, Xing DQ, Liu J and Zhao YL:
Angiotensin II induced upregulation of G alpha q/11, phospholipase
C beta 3 and extracellular signal-regulated kinase 1/2 via
angiotensin II type 1 receptor. Chin Med J (Engl). 117:88–93.
2004.PubMed/NCBI
|
|
14
|
Anavekar NS and Solomon SD: Angiotensin II
receptor blockade and ventricular remodelling. J Renin Angiotensin
Aldosterone Syst. 6:43–48. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang XY, Gao GD, Du XJ, Zhou J, Wang XF
and Lin YX: The signalling of AT2 and the influence on the collagen
metabolism of AT2 receptor in adult rat cardiac fibroblasts. Acta
Cardiol. 62:429–438. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fatini C, Guazzelli R, Manetti P,
Battaglini B, Gensini F, Vono R, Toncelli L, Zilli P, Capalbo A,
Abbate R, et al: RAS genes influence exercise-induced left
ventricular hypertrophy: An elite athletes study. Med Sci Sports
Exerc. 32:1868–1872. 2000. View Article : Google Scholar : PubMed/NCBI
|