Introduction
Intestinal ischemia-reperfusion (II/R) is a common
pathophysiological basis for numerous clinical diseases (1–3). It
has been demonstrated that II/R is a multifactorial and complex
pathophysiologic process that contributes to multi organ failure
and is associated with high mortality rates (4). In addition to local damage to the
bowel, II/R leads to remote organ dysfunction (5,6),
particularly in the lung, resulting in acute lung injury (ALI)
(3,7), characterized by an excess elevation
of pro-inflammatory cytokines and activated neutrophils (8–10).
Animal models and clinical data support the concept
that interleukin (IL)-1, IL-6, tumor necrosis factor (TNF), and
IL-8 are typical cytokines involved in acute inflammation (7). Furthermore, it had been demonstrated
that high levels of IL-6, a multifunctional cytokine, may be a
marker of severity of inflammatory responses and are associated
with inflammatory and immune diseases (11–14).
IL-6 contributes to the inflammatory responses associated with II/R
and serves important roles in the acute phase reaction and in the
progression of ALI (15–17). It has been demonstrated that
pretreatment with dexmedetomidine hydrochloride is useful for
reducing lung damage caused by II/R, and this treatment was
associated with decreased levels of TNF-β, IL-6, toll-like receptor
4 (TLR4) and myeloid differentiation primary response gene 88
(18). Similarly, various studies
have revealed that decreased levels of IL-6 are associated with
attenuation of the inflammatory response in ALI (18–20).
Additional evidence has suggested that IL-6 is directly involved in
the mediation of inflammation via regulation of other inflammatory
cytokine responses and neutrophil extravasation (17). In addition, a recent study
demonstrated that IL-6 regulated intestinal epithelial tight
junction permeability, and this effect was mediated by c-jun
N-terminal kinase, activation of activator protein-1 (AP-1) and
subsequent AP-1 activation of the claudin-2 gene (21). However, for the diseases caused by
II/R, IL-6 instead was demonstrated to have potent
anti-inflammatory roles in the liver following II/R injury, and may
inhibit endotoxin-induced local acute inflammation of the trachea
(22,23). Therefore, as IL-6 has been
suggested to be involved in the development of almost all chronic
inflammatory diseases (14), and
may exert a potential protective effect on II/R-associated
injuries, the exact role of IL-6 in acute inflammatory diseases
requires further elucidation. Furthermore, whether IL-6 knockdown
may serve as an effective strategy for the treatment of these
diseases, and clarification of the potential underlying mechanisms
requires additional further investigation. Previous studies have
suggested that IL-10 may serve a protective role in II/R-induced
ALI (24–26); thus, this requires further
examination.
The RNA interference (RNAi) technique has become a
reliable and powerful technique to inhibit the expression of
targeted genes in both in vitro and in vivo (27–30).
Accordingly, the present study aimed to test the hypothesis that
blocking the proinflammatory cytokine IL-6 by RNAi may alleviate
ALI following II/R, via upregulation of IL-10 expression.
Materials and methods
Animals and ethical statement
The present study used 176 adult male Sprague Dawley
rats (weight, 200–220 g), provided by the Experimental Animal
Center of Sichuan University (Chengdu, China). Study protocols were
followed according to the guidelines for laboratory animal care and
safety as issued by the United States National Institutes of
Health. All animals were raised in plastic cages (n=2/cage) with
soft bedding and free access to food and water in a temperature
(21–25°C) and humidity (45–50%)-controlled room. Animal care and
all experimental protocols were approved by the Animal Care &
Welfare Committee of Kunming Medical University (Kunming,
China).
Experiments
To investigate the function of IL-6 in the lungs of
rats following II/R, human immunodeficiency virus (HIV) based
lentiviral vectors were used to deliver double-stranded short
hairpin RNA (shRNA) sequences to affect RNAi-mediated target gene
knockdown in tissue. The recombinant IL-6 lentiviral vectors along
with the specific shRNA sequences were designed and constructed by
GeneCopoeia™ (Guangzhou, China).
Screening for effective RNAi
sequences
To screen for sequence segments that effectively
mediate IL-6 knockdown, IL-6 gene sequences were obtained from the
National Center for Biotechnology Information database (Bethesda,
MD, USA), and three potential shRNAs sequences targeting IL-6 mRNA
and one nonsense shRNA as a negative control were designed and
purchased from GeneCopoeia. To test their knockdown efficacy in
vitro, four 6-well plates of PC12 cells purchased from the
Animal Research Institute of the Chinese Academy of Medical
Sciences (Beijing, China) were seeded at 8×104/well and
cultured at 37°C in 5% CO2. When the cells reached
50–60% confluency, the culture medium was replaced. Cells were then
transfected with 1 µg shRNA fragment and 3 µl SuperFectin™ II DNA
Transfection reagent (Shanghai Pufei Biological Technology Co.,
Ltd, Shanghai, China) according to the manufacturer's protocol.
After an 18 h transfection, the medium was replaced with Dulbecco's
modified Eagle's medium (DMEM; Hyclone; GE Healthcare Life
Sciences, Logan, UT, USA) supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
50 U/ml penicillin-streptomycin (Hyclone; GE Healthcare Life
Sciences). Subsequently, total RNA of PC12 cells was extracted 48 h
post-transfection and the interference efficiency of IL-6 shRNA was
examined by reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) amplification as described in the following
section. The most effective shRNA was selected to produce the
recombinant lentivirus (31).
Production of the recombinant
lentivirus
The procedure used to produce lentivirus was based
on the Lenti-Pac™ HIV Expression Packaging kit user manual as
previously described (31). In
addition, mCherry fluorescent protein (mCherryFP) was fused to the
plasmid vector. Briefly, 293Tα lentiviral packaging cells
(GeneCopeia Co., Guangzhou, China) were cultured in DMEM
supplemented with 10% heat-inactivated FBS and incubated at 37°C in
5% CO2. Following this, 1.25 µg lentiviral expression
plasmid, 2.5 µl (0.5 µg/µl) Lenti-Pac HIV Expression Packaging mix
and EndoFectin™ Lenti transfection reagent (all from GeneCopeia
Co.) were diluted with Opti-Minimal Essential medium (Invitrogen;
Thermo Fisher Scientific, Inc.) for 25 min at room temperature.
When cell confluence reached 70–80%, the mixture was added to the
culture medium of 293Tα cells. The culture medium was replaced 8 h
after transfection with DMEM supplemented with 10% FBS. In
addition, 10 µl TiterBoost reagent (GeneCopeia Co.) was added to
improve the percentage of virus generation. A total of 72 h
post-transfection, the culture medium was collected, centrifuged at
4°C, 3,000 × g for 30 min, and the supernatant was filtered.
Lentiviral stocks were aliquoted and stored at −80°C for further
use.
II/R model and animal grouping
Animals were randomly divided into the following
groups as indicated in Table I:
Sham, II/R (II/R without any injections), negative-control (NC;
II/R+Lv-NC vector) and IL-6 shRNA (II/R+RSH048925-HIVmU6).
 | Table I.Animal grouping and samples used. |
Table I.
Animal grouping and samples used.
| Group | Lung wet/dry
ratio | H&E/IF | WB/RT-qPCR |
|---|
| Sham | 8 | 8 | 8 |
| II/R | 40 | 40 | 40 |
|
Negative-control | N | 8 | 8 |
| IL-6 shRNA | N | 8 | 8 |
II/R was generated by blocking the superior
mesenteric (SMA) and coeliac (CA) arteries as described previously
(32,33). Rats were fasted with no restriction
of water access for 24 h prior to surgery. Following this, the rats
were anesthetized by intraperitoneal injection with
ketamine-xylazine, which was provided by Kunming Medical University
(Yunnan, China) (100 and 20 mg/kg, respectively) and placed in a
supine position. The intestines were separated gently until the SMA
and CA were isolated, and traumatic microvascular clips were used
to clamp the SMA and CA for 40 min. After 40 min ischemia, the
artery clamps were removed and intestinal perfusion was induced for
0, 8, 16, 24 or 48 h. Sham control animals underwent the same
surgical procedure with the exception of the SMA and CA
clamping.
Lentivirus injection
For IL-6 RNAi, the respective prepared lentivirus
constructs were injected into the right lung tissue of 16 subject
rats over 3 min immediately following arterial clamping. After
clamping arteries for 40 min, the SMA and CA clamps were loosened
and the skin was sutured. Subsequently, 0.5 ml saline was injected
into the enterocoelia. A total of 16 II/R NC rats were injected
with the Lv-NC vector as a control.
Tissue collection
At the end of each experimental time point,
experimental and sham animals were sacrificed by cutting the
abdominal aorta following intraperitoneal injection of
ketamine-xylazine (100 and 20 mg/kg, respectively). Following this,
the lung tissues were removed immediately for further analysis.
Measurement of lung edema
Lung edema was determined by the lung wet/dry weight
ratio as described previously (34). The harvested lung tissue was
immediately weighed to obtain the wet weight. Subsequently, the
tissue was dried in an oven at 90°C for 24 h and reweighed to
obtain the dry weight.
Histological analysis
Histological analysis of the lung tissue was
performed by hematoxylin-eosin (H&E) staining. At the end point
of the various reperfusion times, all rats were sacrificed by
cutting the abdominal aorta following intraperitoneal injection of
ketamine-xylazine (100 and 20 mg/kg, respectively), and harvested
lung tissues were fixed in 4% paraformaldehyde in 0.1 M ice-cold
phosphate buffer, pH 7.4, for >72 h at 4°C. Subsequently, fixed
lung tissues were embedded in paraffin and sectioned at a thickness
of 5 µm, transferred to glass slides, and stained with H&E.
Finally, the stained sections were observed under a light
microscope to detect morphological alterations. The degree of lung
injury was scored at 0 to 4 according to the severity of edema,
neutrophil infiltration, hemorrhage and hyaline membrane formation
as previously described (33).
Furthermore, the number of red cells seeping into the alveolar
interstitium and alveolar space from four fields of each section (3
sections/animal and 8 animals/group) were quantitatively analyzed.
Each slide was evaluated by three investigators blinded to the
experimental information.
RT-qPCR
At the end of the reperfusion period, the
ischemic/reperfused lung tissues were frozen immediately and stored
at −80°C until further required. The mRNA expression of IL-10 and
IL-6 were assayed using RT-qPCR. Total RNA from the lung tissues
was isolated using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol,
and reverse transcribed to cDNA with the RevertAid™
First Strand cDNA Synthesis kit (Takara Biotechnology, Co., Ltd.,
Dalian, China). Subsequently, RT-qPCR of cDNA was performed using
the following primer sequences: Forward, 5′-AGAAGACCAGAGCAGATTTT-3′
and reverse, 5′-GAGAAAAGAGTTGTGCAATG-3′ for IL-6; annealing
temperature, 52°C. Forward, 5′-CAGAAATCAAGGAGCATTTG-3′ and reverse,
5′-CTGCTCCACTGCCTTGCTTT-3′ for IL-10; annealing temperature, 50°C.
Forward, 5′-GAAGATCAAGATCATTGCTCCT-3′ and reverse,
5′-TACTCCTGCTTGCTGATCCA-3′ for β-actin; annealing temperature,
52°C. β-actin served as an internal control. PCR cycling conditions
were as follows: Initial denaturation at 95°C for 2 min,
denaturation at 95°C for 15 sec and amplification at 53°C for 20
sec, followed by extension at 60°C for 30 sec for a total of 40
cycles. The quantitation cycle (Cq) of each sample was recorded,
and data were analyzed by normalization to β-actin values using the
2−∆∆Cq method (35).
The experiment was replicated three times.
Immunofluorescence staining
To examine the location of IL-6 and IL-10 following
II/R, a comparative analysis of immunofluorescence staining was
performed in lung tissue sections. Following routine
de-paraffinization and rehydration, the slices were permeated in
PBS containing 3% goat serum (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) for 30 min at 37°C and incubated overnight at 4°C with
specific rabbit primary antibodies against IL-6 (Ab6672; 1:500;
Abcam, Cambridge, UK) or IL-10 (Ab9969; 1:100; Abcam). Negative
controls were performed by adding PBS instead of the primary
antibody. The next day, sections were rinsed with PBS and incubated
in the dark with an anti-rabbit cy3-labeled secondary antibody
(111–165-003; 1:200; Jackson Laboratories, Bar Harbor, ME, USA),
for 30 min at 37°C. The sections were subsequently washed three
times with PBS, followed by incubation with DAPI (Beyotime
Institute of Biotechnology, Shanghai, China) for 5 min to visualize
the cell nuclei. Finally, sections were observed under a
fluorescence microscope (Leica Microsystems GmbH, Wetzlar,
Germany).
Western blot analysis
Lung samples were frozen immediately and stored at
−80°C until further required. To examine whether the protein
expression levels of IL-6 and IL-10 altered following II/R and IL-6
shRNA transfection, protein was extracted from lung samples from
each group and lysed with radioimmunoprecipitation assay lysis
buffer (Beyotime Institute of Biotechnology) containing 2%
inhibitor cocktail (Roche Diagnostics, Basel, Switzerland). Samples
were centrifuged at 12,000 × g for 15 min at 4°C and the
supernatant was collected. A Bicinchoninic Acid protein assay kit
(Beyotime Institute of Biotechnology) was used to detect the
protein concentration. Protein (100 µg) was separated by sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (15%) at 60 V
for 30 min and 100 V for 1.5 h, following which proteins were
transferred onto polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA) for 4 h at 350 mA. The membranes
were subsequently blocked in 5% skimmed milk in TBS containing
Tween-20 (TBST) for 1 h at room temperature and incubated with
rabbit anti-rat primary antibodies against IL-6 (1:800) or IL-10
(1:2,500) overnight at 4°C. β-actin (ABM40028; 1:1,000; Abbkine
Scientific Co., Ltd., Wuhan, China) served as an internal control.
Subsequently, the membranes were rinsed four times with TBST and
incubated with a horseradish peroxidase conjugated goat anti-rabbit
IgG secondary antibody (ab6721; 1:5,000; Abcam) for 1.5 h. Finally,
membranes were rinsed four times with TBST and the immune complexes
were detected using ChemiDoc XRS System with Image Lab Software
version 2.0 (Bio-Rad Laboratories, Inc., Hercules, CA, USA) with
enhanced chemiluminescence reagent (BL520A; Bio-Rad Laboratories,
Inc.).
Statistical analysis
Statistical analysis was performed using SPSS
software version 18.0 (SPSS, Inc., Chicago, IL, USA). Data are
expressed as the mean ± standard deviation and were analyzed using
a Student's t-test with a two-tailed distribution. For
multiple group comparisons, one-way analysis of variance followed
by Tukey's post hoc test was used. P<0.05 was considered to
indicate a statistically significant difference.
Results
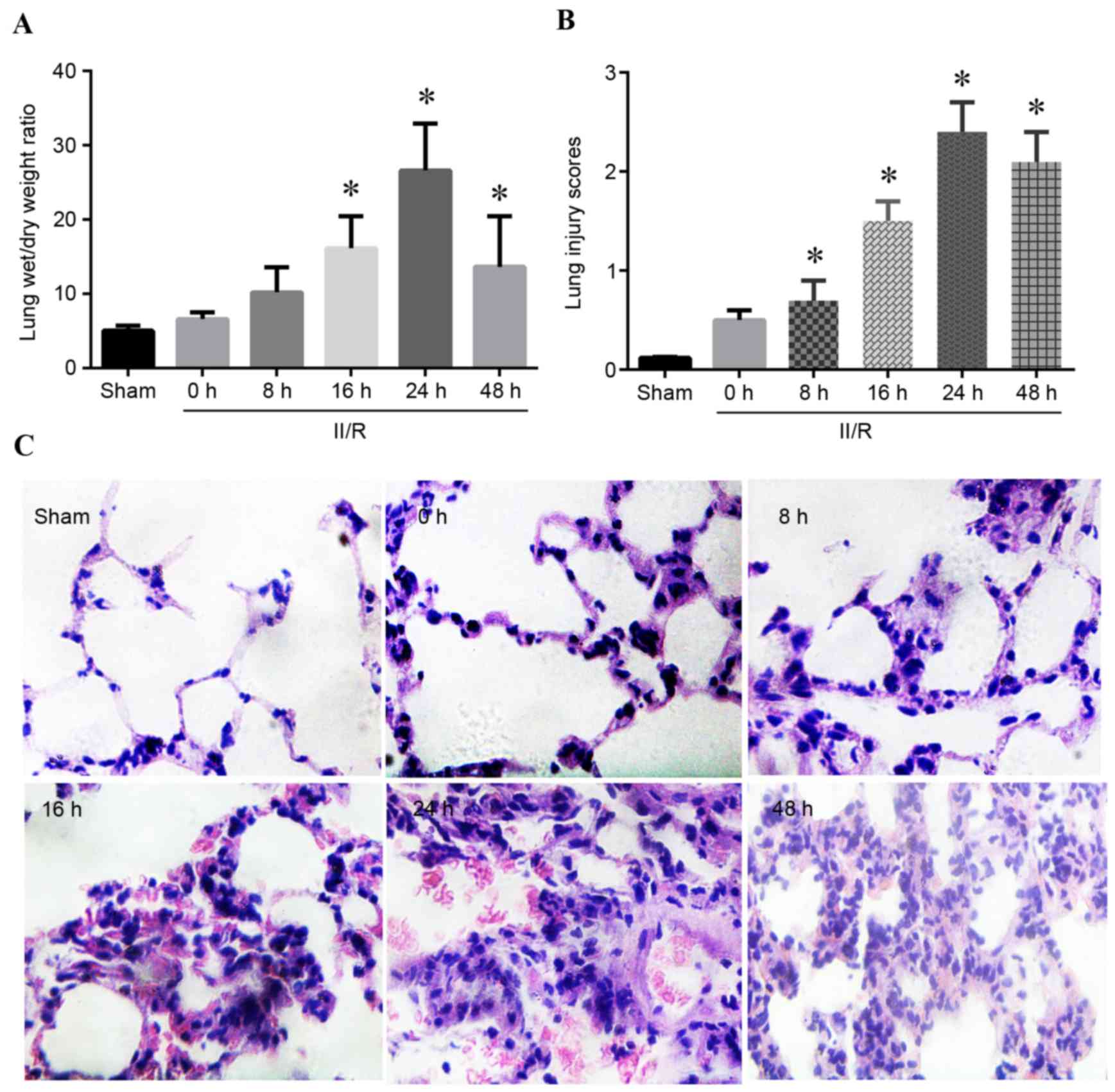
Lung edema and damaged morphology
induced by II/R
Lung edema and morphological alterations were
detected by lung wet/dry weight ratio and H&E staining
following II/R. As presented in Fig.
1A, compared with the sham group, rats subjected to II/R injury
demonstrated significantly increased lung wet/dry weight ratio,
reaching the highest level 24 h post-reperfusion (P<0.05). The
lung injury scores of the II/R rats were significantly higher
compared with the sham group at 8, 16, 24 and 48 h post-reperfusion
(P<0.05; Fig. 1B). H&E
staining revealed that II/R injured rats exhibited histological
evidence of ALI based on a grading system that assessed congestion,
intra-alveolar cellular infiltration and hemorrhage (Fig. 1C).
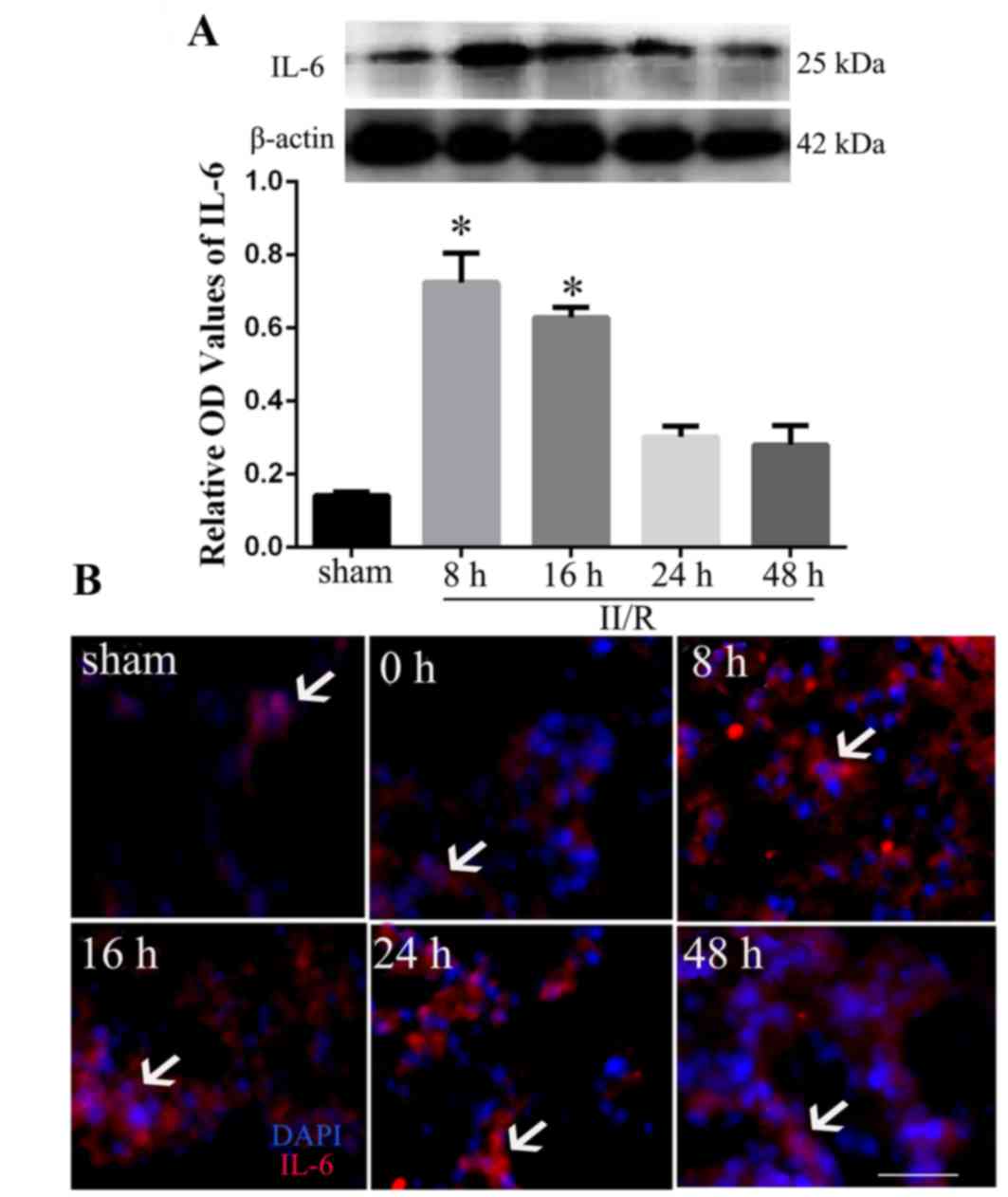
Expression of IL-6 in lung tissues
following II/R
To determine whether IL-6 may be associated with the
ALI following II/R, the protein expression levels of IL-6 were
measured in the lungs by western blot analysis 8, 16, 24 and 48 h
after reperfusion. The results revealed that IL-6 protein
expression levels significantly increased in rats subjected to II/R
compared with sham operated rats, with the highest level observed 8
h post-reperfusion (P<0.05; Fig.
2A). Furthermore, immunofluorescence staining identified that
II/R induced significantly increased intensities of IL-6
immunofluorescence in invading neutrophils, with the strongest
intensity observed at 8 and 16 h after reperfusion (Fig. 2B).
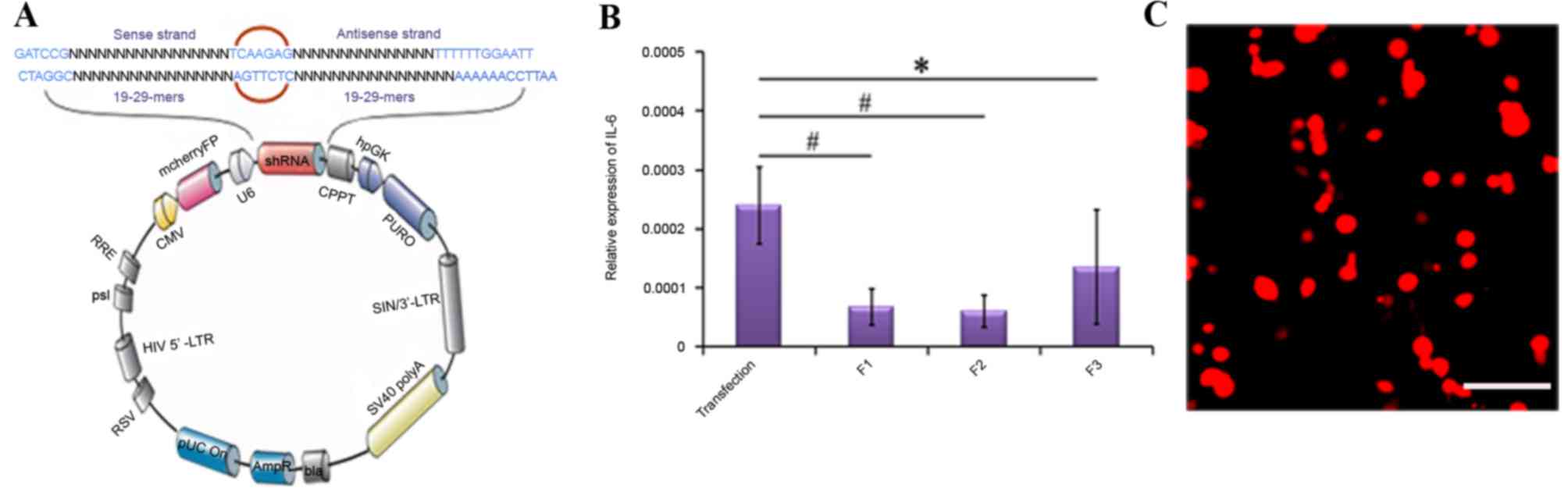
Identification of lentiviral
recombinants
Lentivirus-introduced IL-6 RNAi was used to regulate
the expression of IL-6. Details of the lentivirus constructs with
the inserted shRNA sequence are presented in Fig. 3A. Compared with the transfection
(transfection reagent+random sequence) or negative control groups,
IL-6 mRNA expression levels were reduced in the IL-6 shRNA plasmid
transfection groups, and the shRNA-F2 sequence exhibited the
highest RNAi efficiency (P<0.01; Fig. 3B). Therefore, the shRNA-F2 segment
was used to construct a plasmid and generate a lentiviral
recombinant, following which a recombinant containing the targeted
gene with mCherryFP (IL-6-shRNA) was packaged into 293Tα cells.
Immunofluorescence detection revealed that 293Tα cells emitted red
fluorescence, confirming successful transfection (Fig. 3C). These results confirmed the
efficacy of the lentivirus-mediated IL-6 RNAi in these experimental
conditions.
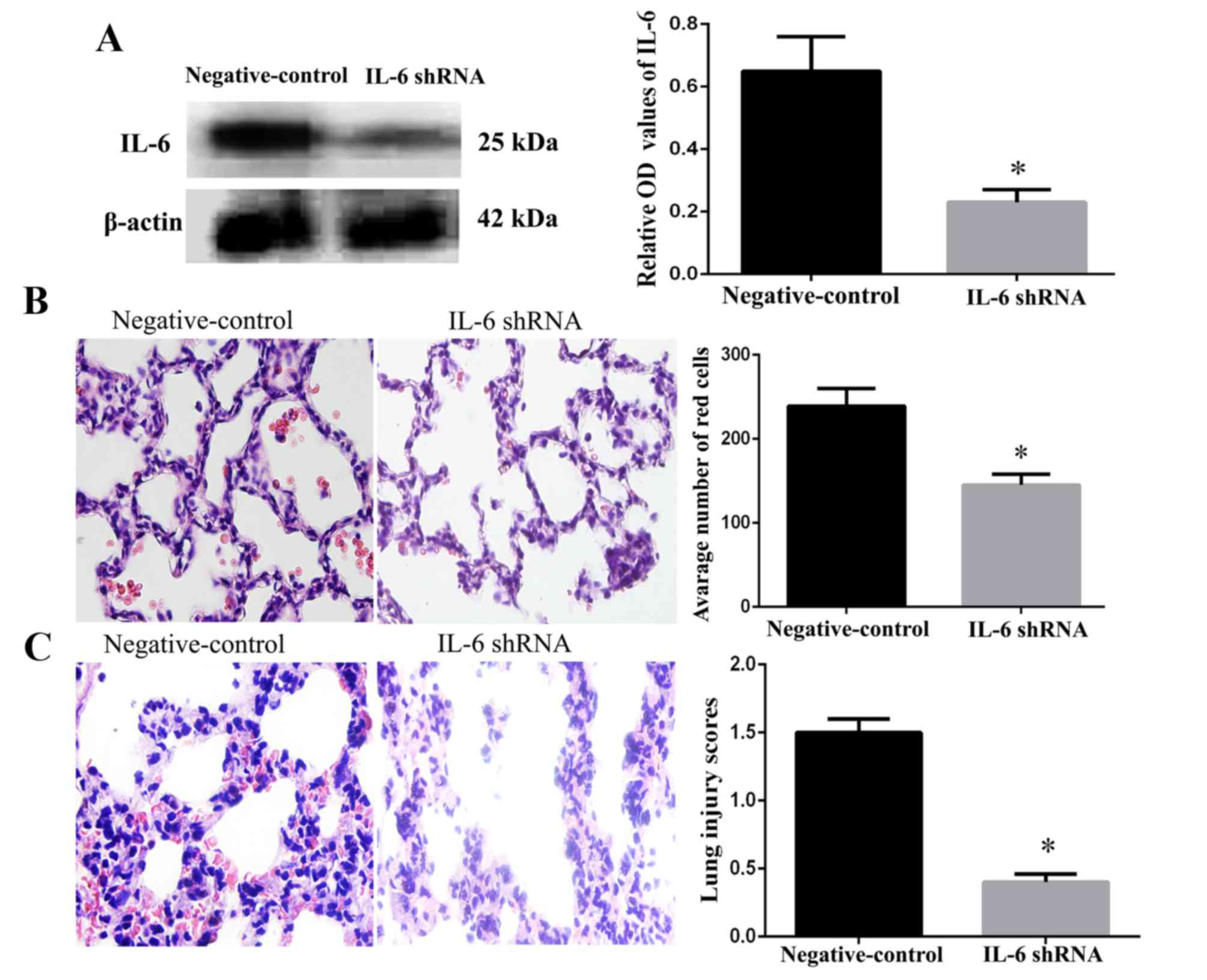
Protective role of IL-6 knockdown on
lung injury
As presented in Fig.
4A, the addition of the IL-6 shRNA lentivirus significantly
decreased the expression of IL-6. The distribution of red cells
(biconcave red in the alveolar interstitium and alveolar space) in
the two groups was identified by H&E staining. A total of 16 h
after reperfusion, the number of red cells in the pulmonary alveoli
in the IL-6 shRNA group was dramatically decreased compared with
the negative control group, which had a large number of red cells
(P<0.05; Fig. 4B). As a result,
the lung injury scores of rats were improved in the IL-6 shRNA
group compared with the negative control group (P<0.05; Fig. 4C). These results supported the
hypothesis that transfection of a shRNA lentivirus may downregulate
the expression of IL-6 and alleviate the effects of the
inflammatory response.
Role of IL-6 knockdown on the
expression of IL-10
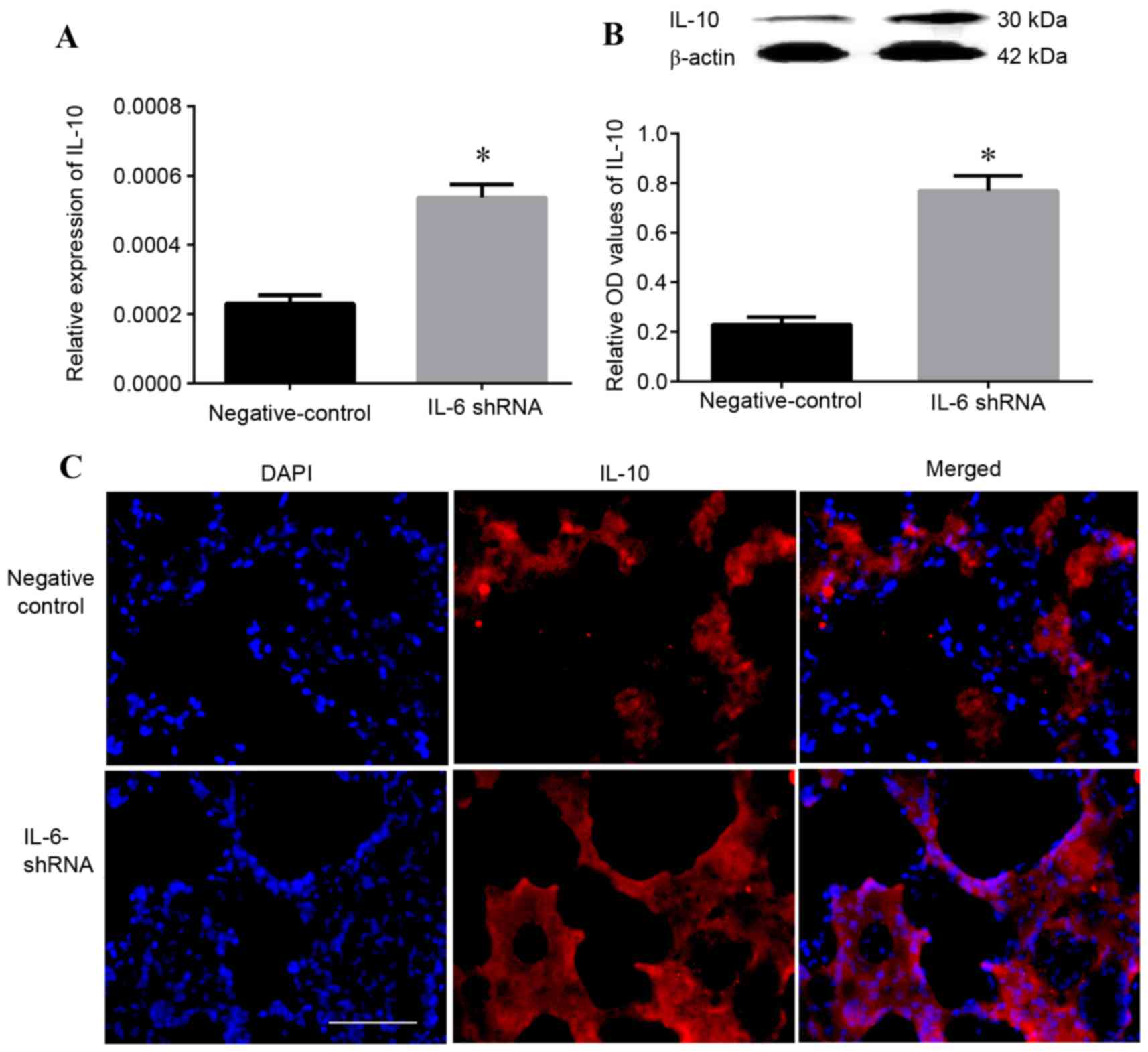
To investigate the effect of IL-6 on the expression
of IL-10, the mRNA (Fig. 5A) and
protein (Fig. 5B) expression
levels of IL-10 were analyzed by RT-qPCR and western blot analysis,
respectively. The results revealed that 16 h after reperfusion,
mRNA and protein expression levels of IL-10 were increased compared
with the negative control group (P<0.05). Furthermore,
immunofluorescence analysis demonstrated that following IL-6 shRNA
transfection, the density of IL-10 immunoreactivity in lung tissue
was significantly increased compared with the control
Lv-NC-transfected group (Fig.
5C).
Discussion
An II/R model was used in the present study to
induce ALI, including lung edema and elevated lung injury scores,
and IL-6 expression levels were demonstrated to be elevated. To
understand the physiological role of IL-6 in mediating
II/R-associated lung injuries, an IL-6 shRNA lentiviral construct
was used to specifically suppress IL-6 expression in the lungs of
experimental animals. RT-qPCR and western blot analysis revealed
that IL-6 silencing contributed to a progressive and parallel
reduction in IL-6 mRNA and protein expression levels, which was
followed by improved lung injury scores and increased IL-10
expression. Therefore, these results provided experimental support
for the clinical application of IL-6 knockdown in the treatment of
ALI caused by II/R.
The results of the histological analyses performed
in this study closely associate with those identified in previous
studies (36–38), thus confirming the presence of ALI
caused by II/R and the success of this rat model. In the present
study, subsequent analyses were performed at various time intervals
to determine the period at which the observed phenomena were most
significant. From this, it was determined that the most severe lung
injury occurred 24 h post-reperfusion, as indicated by lung edema
and increased lung injury scores. Furthermore, it was observed that
the mRNA and protein expression levels of IL-6 increased in lung
tissues following II/R, suggesting that elevated IL-6 may be
associated with lung injury caused by II/R (18). In support of this, previous studies
have reported that intra-alveolar cell infiltrates, alveolar
hemorrhages and excessive elevation of proinflammatory cytokines
including IL-1, IL-6, TNF and IL-8 are important factors in the
occurrence and development of II/R-mediated ALI (9,10,39).
In addition, TLR4 has been demonstrated to serve an important role
in the pathogenesis of II/R-induced ALI and inflammation, and p38
kinase and nuclear factor-κB may be involved in TLR4
signaling-mediated lung inflammatory processes during II/R
(3,40), illustrating the complexity of this
physiological response.
IL-6 is a pleiotropic acute reactant cytokine
involved in inflammatory responses (17) that may be promptly and transiently
produced in response to infections and tissue injuries,
contributing to host defenses via stimulation of acute phase
responses, hematopoiesis, and immune reactions (40). Furthermore, IL-6 exerts various
effects in addition to those on hepatocytes and lymphocytes that
are frequently detected in chronic inflammatory diseases (14,41,42).
The present study primarily investigated acute inflammatory disease
following II/R treatment, which differs from previous studies that
focused on the chronic inflammatory process (14).
Conversely, IL-6 may have an anti-inflammatory
effect in vitro and in vivo, as IL-6 administration
has been demonstrated to inhibit TNF-α and IL-1β signaling, reduce
aggregation of polymorphonuclear cells (PMNs), and protect against
the damage caused by toxins in vivo (43). Together, these disparate findings
indicated that the function of IL-6 on the process of ALI is
complex; therefore, further confirmation of the exact role of IL-6
in acute inflammatory diseases associated with II/R is
required.
The present study demonstrated that IL-6 knockdown
led to significantly increased expression of IL-10, and markedly
reduced lung edema scores and leakage of red blood cells to the
pulmonary alveoli in II/R model rats compared with the control
group. These results are particularly relevant as no specific
therapeutic treatments are currently available for ALI caused by
II/R. Based on the observation that oxidative stress and mast cells
interact together and promote II/R-induced ALI (44), previous studies have revealed that
osthol, valproic acid, protocatechuic acid and cyclic
arginine-glycine-aspartate peptide are able to affect survival and
the development of ALI in a rat model of II/R. These factors may
impact recovery following lung injury via anti-oxidant and
anti-inflammatory effects (33,45–48).
Support for the potential use of IL-6 knockdown in II/R-associated
ALI, as studied here, is provided by numerous reports. Goodman
et al (49) injected IL-6
into the healthy endotrachea of mice, and demonstrated that this
led to PMN aggregation and pulmonary edema in murine lung tissue,
suggesting that IL-6 may cause lung injury. Conversely, Bhatia
et al (50) confirmed that
injection of an IL-6 antibody may decrease the level of C-reactive
protein (CRP) in patients with sepsis to healthy levels, suggesting
that IL-6 inhibits CRP. Furthermore, another study reported that
IL-6 serves a protective role in inflammation, as increased IL-6
was negatively correlated with sepsis mortality (17). However, few studies have placed
emphasis on the protective effect of IL-6 knockdown and its
regulation of inflammatory cytokines in ALI induced by II/R.
Therefore, to the best of our knowledge, this is the first study to
demonstrate a protective effect of IL-6 RNA knockdown on ALI after
II/R, and further, to demonstrate the potential underlying
mechanism of IL-10 upregulation. These data provide the foundation
for the potential application of IL-6 RNAi for the treatment of
II/R-associated ALI.
In conclusion, the present study demonstrated that
II/R may induce ALI and enhance the expression of IL-6 in an II/R
rat model. In addition, IL-6 inhibition in lungs by an
IL-6-shRNA-lentivirus alleviated the inflammatory response. This
protective role was associated with upregulated IL-10 expression in
lung tissues. Therefore, the results of the present study may serve
as a basis for the development of novel treatment strategies for
ALI and associated clinical diseases including systemic
inflammatory response syndrome and multiple organ dysfunction
syndrome evoked by II/R.
Acknowledgements
The present study was supported by the Program for
Innovative Research Team (in Science and Technology) in University
of Yunnan Province and a grant from the Key Natural Science
Foundation of Yunnan (grant no. 2013FZ264).
References
|
1
|
Tadros T, Traber DL, Heggers JP and
Herndon DN: Effects of interleukin-1alpha administration on
intestinal ischemia and reperfusion injury, mucosal permeability
and bacterial translocation in burn and sepsis. Ann Surg.
237:101–109. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kumar S, Boehm J and Lee JC: p38 MAP
kinases: Key signalling molecules as therapeutic targets for
inflammatory diseases. Nat Rev Drug Discov. 2:717–726. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tendler DA: Acute intestinal ischemia and
infarction. Semin Gastrointest Dis. 14:66–76. 2003.PubMed/NCBI
|
|
4
|
Kim K, Li YQ, Jin G, Chong W, Liu BL, Lu
J, Lee K, Demoya M, Velmahos G and Alam HB: Effect of valproicacid
on acute lung injury in a rodent model of intestinal ischemia
reperfusion. Resuscitation. 83:243–248. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Harward TR, Brooks DL, Flynn TC and Seeger
JM: Multiple organ dysfunction after mesenteric artery
revascularization. J Vasc Surg. 18:459–469. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cui T, Miksa M, Wu R, Komura H, Zhou M,
Dong W, Wang Z, Higuchi S, Chaung W, Blau SA, et al: Milk fat
globule epidermal growth factor 8 attenuates acute lung injury in
mice after intestinal ischemia and reperfusion. Am J Respir Crit
Care Med. 181:238–246. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Narita K, Kuwabara Y and Fujii Y: Lung
injury after intestinal ischemia-reperfusion may be avoided by the
reduced absorption of locally produced cytokines. Surg Today.
34:937–942. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sayan H, Ozacmak VH, Sen F, Cabuk M, Atik
DY, Igdem AA and Ozacmak ID: Pharmacological preconditioning with
erythropoietin reduces ischemia-reperfusion injury in the small
intestine of rats. Life Sci. 84:364–371. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sukhotnik I, Slijper N, Pollak Y,
Chemodanov E, Shaoul R, Coran AG and Mogilner JG: Parenteral
omega-3 fatty acids (Omegaven) modulate intestinal recovery after
intestinal ischemia-reperfusion in a rat model. J Pediatr Surg.
46:1353–1360. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Grommes J and Soehnlein O: Contribution of
neutrophils to acute lung injury. Mol Med. 17:293–307. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Damas P, Ledoux D, Nys M, Vrindts Y, De
Groote D, Franchimont P and Lamy M: Cytokine serum level during
severe sepsis in human IL-6 as a marker of severity. Ann Surg.
215:356–362. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Farmer DG, McDiarmid SV, Kuniyoshi J,
Robert ME, Shaked A and Busuttil RW: Intragraft expression of
messenger RNA for interleukin-6 and tumor necrosis factoralpha is a
predictor of rat small intestine transplant rejection. J Surg Res.
57:138–142. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
McDiarmid SV, Farmer DG, Kuniyoshi JS,
Robert M, Khadavi A, Shaked A and Busuttil RW: The correlation of
intragraft cytokine expression with rejection in rat small
intestine transplantation. Transplantation. 58:690–697. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tanaka T, Narazaki M and Kishimoto T: IL-6
in inflammation, immunity, and disease. Cold Spring Harb Perspect
Biol. 6:a0162952014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Molmenti EP, Ziambaras T and Perlmutter
DH: Evidence for an acute phase response in human intestinal
epithelial cells. J Biol Chem. 268:14116–14124. 1993.PubMed/NCBI
|
|
16
|
Beagley KW, Eldridge JH, Lee F, Kiyono H,
Everson MP, Koopman WJ, Hirano T, Kishimoto T and McGhee JR:
Interleukins and IgA synthesis. Human and murine interleukin 6
induce high rate IgA secretion in IgA-committed B cells. J Exp Med.
169:2133–2148. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kimizuka K, Nakao A, Nalesnik MA, Demetris
AJ, Uchiyama T, Ruppert K, Fink MP, Stolz DB and Murase N:
Exogenous IL-6 inhibits acute inflammatory responses and prevents
ischemia/reperfusion injury after intestinal transplantation. Am J
Transplant. 4:482–494. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shen J and Fu G, Jiang L, Xu J, Li L and
Fu G: Effect of dexmedetomidine pretreatment on lung injury
following intestinal ischemia-reperfusion. Exp Ther Med.
6:1359–1364. 2013.PubMed/NCBI
|
|
19
|
He DK, Shao YR, Zhang L, Shen J, Zhong ZY,
Wang J and Xu G: Adenovirus-delivered angiopoietin-1 suppresses
NF-κB and p38 MAPK and attenuates inflammatory responses in
phosgene-induced acute lung injury. Inhal Toxicol. 26:185–192.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
O'Dea KP, Dokpesi JO, Tatham KC, Wilson MR
and Takata M: Regulation of monocyte subset proinflammatory
responses within the lung microvasculature by the p38 MAPK/MK2
pathway. Am J Physiol Lung Cell Mol Physiol. 301:L812–L821. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Al-Sadi R, Ye D, Boivin M, Guo S, Hashimi
M, Ereifej L and Ma TY: Interleukin-6 modulation of intestinal
epithelial tight junction permeability is mediated by JNK pathway
activation of claudin-2 gene. PLoS One. 9:e853452014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ulich TR, Yin S, Guo K, Yi ES, Remick D
and del Castillo J: Intratracheal injection of endotoxin and
cytokines. II. Interleukin-6 and transforming growth factor beta
inhibit acute inflammation. Am J Pathol. 138:1097–1101.
1991.PubMed/NCBI
|
|
23
|
Camargo CA Jr, Madden JF, Gao W, Selvan RS
and Clavien PA: Interleukin-6 protects liver against warm
ischemia/reperfusion injury and promotes hepatocyte proliferation
in the rodent. Hepatology. 26:1513–1520. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wei D, Huang ZH, Zhang RH, Wang CL, Xu MJ,
Liu BJ, Wang GH and Xu T: Roles of p38 MAPK in the regulation of
the inflammatory response to swine influenza virus-induced acute
lung injury in mice. Acta Virol. 58:374–379. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guido BC, Zanatelli M, Tavares-de-Lima W,
Oliani SM and Damazo AS: Annexin-A1 peptide down-regulates the
leukocyte recruitment and up-regulates interleukin-10 release into
lung after intestinal ischemia-reperfusion in mice. J Inflamm
(Lond). 10:102013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gloor B, Todd KE, Lane JS, Rigberg DA and
Reber HA: Mechanism of increased lung injury after acute
pancreatitis in IL-10 knockout mice. J Surg Res. 80:110–114. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Napoli C, Lemieux C and Jorgensen R:
Introduction of a chimeric chalcone synthase gene into petunia
results in reversible co-suppression of homologous genes in trans.
Plant Cell. 2:279–289. 1990. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Younis A, Siddique MI, Kim CK and Lim KB:
RNA interference (RNAi) induced gene silencing: A promising
approach of hi-tech plant breeding. Int J Biol Sci. 10:1150–1158.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bosher JM and Labouesse M: RNA
interference: Genetic wand and genetic watchdog. Nat Cell Biol.
2:E31–E36. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim DH and Rossi JJ: Strategies for
silencing human disease using RNA interference. Nat Rev Genet.
8:173–184. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu R, Zhao W, Zhao Q, Liu SJ, Liu J, He
M, Xu Y, Wang W, Liu W, Xia QJ, et al: Endoplasmic reticulum
protein 29 protects cortical neurons from apoptosis and promoting
corticospinal tract regeneration to improve neural behavior via
caspase and Erk signal in rats with spinal cord transection. Mol
Neurobiol. 50:1035–1048. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Crisafulli C, Mazzon E, Galuppo M,
Paterniti I, Caminiti R and Cuzzocrea S: Olprinone attenuates the
development of ischemia/reperfusion injury of the gut. Intensive
Care Med. 36:1235–1247. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim K, Li Y, Jin G, Chong W, Liu B, Lee K,
Demoya M, Velmahos GC and Alam HB: Effect of valproic acid on acute
lung injury in a rodent model of intestinal ischemia reperfusion.
Resuscitation. 83:243–248. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pei YH, Cai XM, Chen J, Sun BD, Sun ZR,
Wang X and Qian XM: The role of p38 MAPK in acute paraquat-induced
lung injury in rats. Inhal Toxicol. 26:880–884. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C (T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zheng DY, Zhou M, Jin J, He M, Wang Y, Du
J, Xiao XY, Li PY, Ye AZ, Liu J and Wang TH: Inhibition of P38 MAPK
downregulates the expression of IL-1β to protect lung from acute
injury in intestinal ischemia reperfusion rats. Mediators Inflamm.
2016:93480372016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang L, Tan Y, Tian J, Ma HY, Li JT and
Luo CZ: Morphological character of lung injury and its functional
implication in adult rats subjected to brain ischemia. Ibrain.
1:1–8. 2015.
|
|
38
|
Zhou M and Wang TH: Temporal and spatial
pattern of lung injury in rats subjected to intestinal ischemia
reperfusion. Idiscovery. 1:1–8. 2015.
|
|
39
|
Zarubin T and Han J: Activation and
signaling of the p38 MAP kinase pathway. Cell Res. 15:11–18. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ben DF, Yu XY, Ji GY, Zheng DY, Lv KY, Ma
B and Xia ZF: TLR4 mediates lung injury and inflammation in
intestinal ischemia-reperfusion. J Surg Res. 174:326–333. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hirano T, Akira S, Taga T and Kishimoto T:
Biological and clinical aspects of interleukin 6. Immunol Today.
11:443–449. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Akira S, Taga T and Kishimoto T:
Interleukin-6 in biology and medicine. Adv Immunol. 54:1–78. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Riffo-Vasquez Y and Spina D: Role of
cytokines and chemokines in bronchial hyperresponsiveness and
airway inflammation. Pharmacol Ther. 94:185–211. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhao W, Gan X, Su G, Wanling G, Li S, Hei
Z, Yang C and Wang H: The interaction between oxidative stress and
mast cell activation plays a role in acute lung injuries induced by
intestinal ischemia-reperfusion. J Surg Res. 187:542–552. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mo LQ, Chen Y, Song L, Wu GM, Tang N,
Zhang YY, Wang XB, Liu KX and Zhou J: Osthole prevents intestinal
ischemia-reperfusion-induced lung injury in a rodent model. J Surg
Res. 189:285–294. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zabot GP, Carvalhal GF, Marroni NP,
Hartmann RM, da Silva VD and Fillmann HS: Glutamine prevents
oxidative stress in a model of mesenteric ischemia and reperfusion.
World J Gastroenterol. 20:11406–11414. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Matsuo S, Yang WL, Aziz M, Jacob A and
Wang P: Cyclic arginine-glycine-aspartate attenuates acute lung
injury in mice after intestinal ischemia/reperfusion. Crit Care.
17:R192013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang GZ, Yao JH, Jing HR, Zhang F, Lin MS,
Shi L, Wu H, Gao DY, Liu KX and Tian XF: Suppression of the p66shc
adapter protein by protocatechuic acid prevents the development of
lung injury induced by intestinal ischemia reperfusion in mice. J
Trauma Acute Care Surg. 73:1130–1137. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Goodman RB, Pugin J, Lee JS and Matthay
MA: Cytokine-mediated inflammation in acute lung injury. Cytokine
Growth Factor Rev. 14:523–535. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Bhatia M and Moochhala S: Role of
inflammatory mediators in the pathophysiology of acute respiratory
distress syndrome. J Pathol. 202:145–156. 2004. View Article : Google Scholar : PubMed/NCBI
|