Introduction
Resveratrol is a natural polyphenolic compound that
is present in grapes, berries and red wine. Accumulating evidence
indicates that resveratrol may provide various health benefits,
including protective properties against cardiovascular disease and
cancer (1,2). The majority of the anti-apoptotic or
anti-inflammatory effects of resveratrol are associated with the
activation of sirtuin 1 (SIRT1), which is known as a nicotinamide
adenine dinucleotide (NAD+)-dependent deacetylase that
activates genes associated with longevity and survival (3,4). The
French population reportedly tends to consume high levels of
saturated fatty acids in their meals however, their circulating
levels remain relatively low and they have low mortality rates
associated with coronary heart disease, when compared with other
countries that have a similar consumption of saturated fats. This
may be due to their frequent consumption of red wine containing
abundant resveratrol (5).
As for the effects of resveratrol on bone cells,
resveratrol is reported to stimulate the differentiation of
osteoblasts (6,7), which conduct bone formation and also
bone resorption through the expression of receptor activator of
nuclear factor-κB (RANK)-ligand (RANKL), which binds to RANK
expressed on osteoclasts (8).
Previous reports from this lab have demonstrated that, in
osteoblast-like MC3T3-E1 cells, resveratrol suppresses the
synthesis of osteoprotegerin (OPG), a decoy receptor of RANKL
(8), induced by prostaglandin
F2α (PGF2α), PGD2,
PGE1, PGE2 or basic fibroblast growth factor
(FGF-2) (9–13). However, resveratrol increases bone
morphogenetic protein-4 (BMP-4)-induced OPG synthesis (14). In addition, synthesis of vascular
endothelial growth factor (VEGF) induced by BMP-4 or transforming
growth factor-β (TGF-β) is inhibited by resveratrol in MC3T3-E1
cells (15,16). These findings indicate that
resveratrol may orchestrate stimuli from numerous bone remodeling
agents, resulting in the modulation of bone metabolism. However,
the exact mechanism underlying the effects of resveratrol on
osteoblasts remains to be elucidated.
Osteocalcin, which is synthesized in osteoblasts, is
the most abundant non-collagenous protein and is also a marker of
mature osteoblast phenotype (17).
Osteocalcin undergoes post-translational modification whereby
glutamic acid residues are carboxylated to form γ-carboxyglutamic
acid (Gla) residues by vitamin K-dependent γ-carboxylase (17). It has been reported that
osteocalcin-deficient mice present higher bone mass and bone
strength, indicating that osteocalcin is a determining factor in
bone formation (18). In addition,
it was previously proposed that uncarboxylated osteocalcin released
from osteoblasts functions as a hormone, which regulates energy
metabolism by stimulation of insulin secretion from pancreatic
β-cells and by upregulation of insulin sensitivity through
adiponectin production by adipocytes (19). Thus, the evidence indicates that
bone may also have a crucial role as an endocrine organ, regulating
energy metabolism through the production of osteocalcin by
osteoblasts (20).
In addition to a role in the modulation of
whole-body metabolism, thyroid hormone is an important modulator of
skeletal function. Excess levels of thyroid hormone, which is
termed hyperthyroidism, accelerates metabolic turnover rate and
increases the ratio of bone resorption to bone formation, which may
lead to osteoporosis (21,22). It was previously reported that bone
mineral density is significantly decreased and the risk of fracture
increases in untreated patients with hyperthyroidism (23). The thyroid hormone receptor is a
member of the steroid hormone receptor superfamily (24). It is established that thyroid
hormone, like other steroid hormones, binds to its specific
intracellular receptors and the complex subsequently induces the
expression of the gene network (24). In our previous study (25), triiodothyronine (T3)
stimulated osteocalcin synthesis in osteoblast-like MC3T3-E1 cells
and p38 mitogen-activated protein (MAP) kinase positively regulated
the synthesis. The present study investigated the effect of
resveratrol on T3-stimulated osteocalcin synthesis in
MC3T3-E1 cells. The results of the present study indicate that
resveratrol may suppress T3-stimulated osteocalcin
synthesis at a point upstream of transcription in osteoblast-like
MC3T3-E1 cells, and that the inhibition by resveratrol is mediated,
at least partially, by SIRT1 activation.
Materials and methods
Materials
T3 was obtained from Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany). Resveratrol and SRT1720 were obtained from
EMD Millipore (Billerica, MA, USA). The mouse osteocalcin ELISA kit
(cat. no. BT-470) was obtained from Alfa Aesar; Thermo Fisher
Scientific, Inc. (Lancashire, UK). Phospho-specific p38 MAP kinase
(cat. no. #4511) and p38 MAP kinase (cat. no. #9212) antibodies
were obtained from Cell Signaling Technology, Inc. (Danvers, MA,
USA). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (cat. no.
sc-25778) antibody was purchased from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA). An enhanced chemiluminescence (ECL) western
blotting detection system was obtained from GE Healthcare Life
Sciences (Chalfont, UK). Other materials and chemicals were
obtained from commercial sources. T3 was dissolved in 0.1 M NaOH.
Resveratrol and SRT1720 were dissolved in dimethyl sulfoxide. The
maximum concentration of dimethyl sulfoxide was 0.1%, which did not
affect the assay for osteocalcin, reverse
transcription-quantitative polymerase chain reaction (RT-qPCR),
luciferase reporter assay or western blot analysis.
Cell culture
Cloned osteoblast-like MC3T3-E1 cells that were
derived from newborn mouse calvaria (26) were maintained as previously
described (27). Briefly, the
cells were cultured in α-minimum essential medium (α-MEM)
containing 10% fetal bovine serum (FBS; cat. no. 12483-020; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) at 37°C in a
humidified atmosphere of 5% CO2/95% air. The cells were
seeded into 35 mm diameter dishes (5×104 cells/dish) or
90 mm diameter dishes (2×105 cells/dish) in α-MEM
containing 10% FBS. After 5 days, the medium was exchanged for
α-MEM containing 0.3% FBS; the cells were used for experiments 48 h
following this exchange.
Assay for osteocalcin
Cultured cells were pretreated with various doses of
resveratrol (0, 10, 50 and 70 µM), or SRT1720 at 37°C for 60 min,
and subsequently stimulated by 10 nM of T3 or vehicle in 1 ml of
α-MEM containing 0.3% FBS at 37°C for 0, 48 or 96 h. In the
experiment investigating the effect of various resveratrol doses on
osteocalcin release, cells were stimulated by T3 or vehicle for 96
h, whereas cells pretreated with SRT170 were stimulated by T3 or
vehicle for 0, 48 or 96 h. The conditioned medium was collected at
the end of incubation and the osteocalcin concentration in the
medium was measured using the mouse osteocalcin ELISA kit according
to the manufacturer's protocol.
RT-qPCR
Cultured cells were pretreated with 50 µM of
resveratrol or vehicle at 37°C for 60 min, and subsequently
stimulated by 10 nM of T3 or vehicle in 1 ml of α-MEM containing
0.3% FBS for 48 h. Total RNA was isolated and transcribed into cDNA
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
and Omniscript® RT kit (Qiagen, Inc., Valencia, CA,
USA), respectively. qPCR was performed using a
LightCycler® Fast Start DNA Master SYBR Green I kit in
capillaries (Roche Diagnostics, Basel, Switzerland). The forward
and reverse primers for mouse osteocalsin mRNA were synthesized
based on the reports by Zhang et al (28) and were obtained from Greiner
Bio-One Co., Ltd. (Tokyo, Japan). These primer sequences (listed
5′-3′) were as follows: forward, TTC TGC TCA CTC TGC TGACC and
reverse, TTT GTA GGC GGT CTT CAA GC. The forward and reverse
primers for mouse GAPDH mRNA were synthesized based on the report
by Simpson et al (29) and
were obtained from Sigma-Aldrich; Merck KGaA. These primer
sequences (listed 5′-3′) were as follows: forward, AAC GAC CCC TTC
ATT GAC and reverse, TCC ACG ACA TAC TCA GCAC. The reaction
mixtures were incubated at 95°C for 10 min, followed by 40 cycles
at 60°C for 5 sec and 72°C for 7 sec. The amplified products were
determined by melting curve analysis (30), according to the system protocol.
The data were analyzed using the second derivative maximum method
and LightCycler3 Data Analysis software (version 3.5.28; Roche
Diagnostics). The osteocalcin mRNA levels were normalized to those
of GAPDH mRNA using the pfaffl method (31).
Luciferase reporter assay
A reporter plasmid, pDR4 (thyroid
hormone-responsive element)-Luc was purchased from
Stratagene (Agilent Technologies, Inc., Santa Clara, CA, USA). The
cultured cells were pretreated with 50 µM resveratrol or vehicle at
37°C for 6 h after the transfection with the pDR4-Luc
reporter plasmid (1 µg/dish) using UniFector transfection reagent
at 37°C for 6 h (B-Bridge International, Inc., Santa Clara, CA,
USA). The cells were cotransfected with pRL-CMV (Renilla
luciferase; 0.1 µg/dish; Promega Corporation, Madison, WI, USA) as
an internal standard to normalize transfection efficiency.
Following pretreatment, cells were stimulated by 10 nM T3 or
vehicle at 37°C for 48 h. Luciferase activity of the cell lysates
was measured using a Dual-Luciferase® Reporter Assay
system (Promega Corporation) according to the manufacturer's
protocol.
Western blot analysis
The cultured cells were pretreated with various
doses of resveratrol (0, 10, 30 and 50 µM) at 37°C for 60 min and
subsequently stimulated by 10 nM of T3 in a-MEM containing 0.3% FBS
at 37°C for 120 min. Cells were washed twice with PBS and lysed,
homogenized and sonicated in a lysis buffer containing 62.5 mM
Tris/HCl (pH 6.8), 2% SDS, 50 mM dithiothreitol and 10% glycerol.
SDS-PAGE was performed as described by Laemmli (32) on 10% polyacrylamide gels. Protein
quantification was performed using a Pierce Bicinchoninic Acid
Protein Assay kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Protein (10 µg/lane) was fractionated and
transferred onto an Immun-Blot® PVDF membrane (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The membranes were blocked
with 5% fat-free dry milk in TBS-Tween-20 (TBS-T; 20 mM Tris-HCl,
pH 7.6, 137 mM NaCl, 0.1% Tween-20) at 37°C for 1 h prior to
incubation with primary antibodies. Western blot analysis was
performed as described previously (33) using phospho-specific p38 MAP kinase
antibodies, p38 MAP kinase antibodies or GAPDH as primary
antibodies, with peroxidase-labeled antibodies raised in goat
against rabbit IgG (KPL, Inc., Gaithersburg, MD, USA) used as
secondary antibodies. The primary and secondary antibodies were
diluted at 1:1,000 with 5% fat-free dry milk in TBS-T. The
peroxidase activity on the PVDF sheet was visualized on X-ray film
by means of the ECL western blotting detection system (GE
Healthcare Life Sciences) according to the manufacturer's
protocol.
Statistical analysis
Results were analyzed by one-way analysis of
variance followed by the Bonferroni method for multiple comparisons
between pairs, using Microsoft Office Excel 2013 for Windows
(Microsoft Corporation, Redmond, WA, USA) and P<0.05 was
considered to indicate a statistically significant difference.
Results are presented as the mean ± standard error of the mean of
triplicate results from three independent cell preparations.
Results
Effect of resveratrol on the
T3-stimulated osteocalcin release in MC3T3-E1 cells
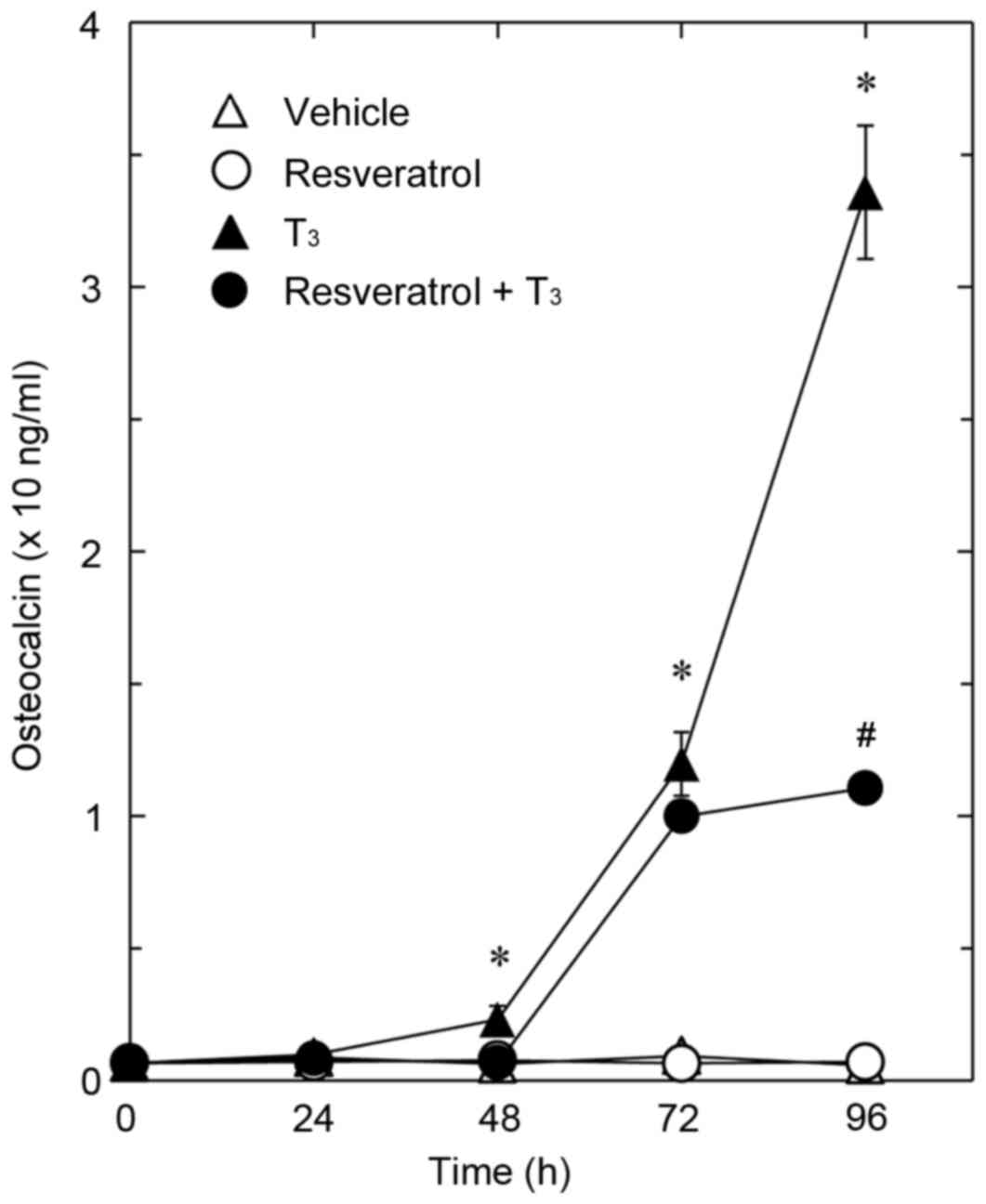
The present study first investigated the effect of
resveratrol on T3-stimulated osteocalcin release in osteoblast-like
MC3T3-E1 cells. As previously described (25), T3 stimulated secretion of
osteocalcin after 48 h. Resveratrol, which had a limited effect on
the release of osteocalcin alone, significantly reduced
T3-stimulated release of osteocalcin compared with cells treated
with T3 only (Fig. 1). The
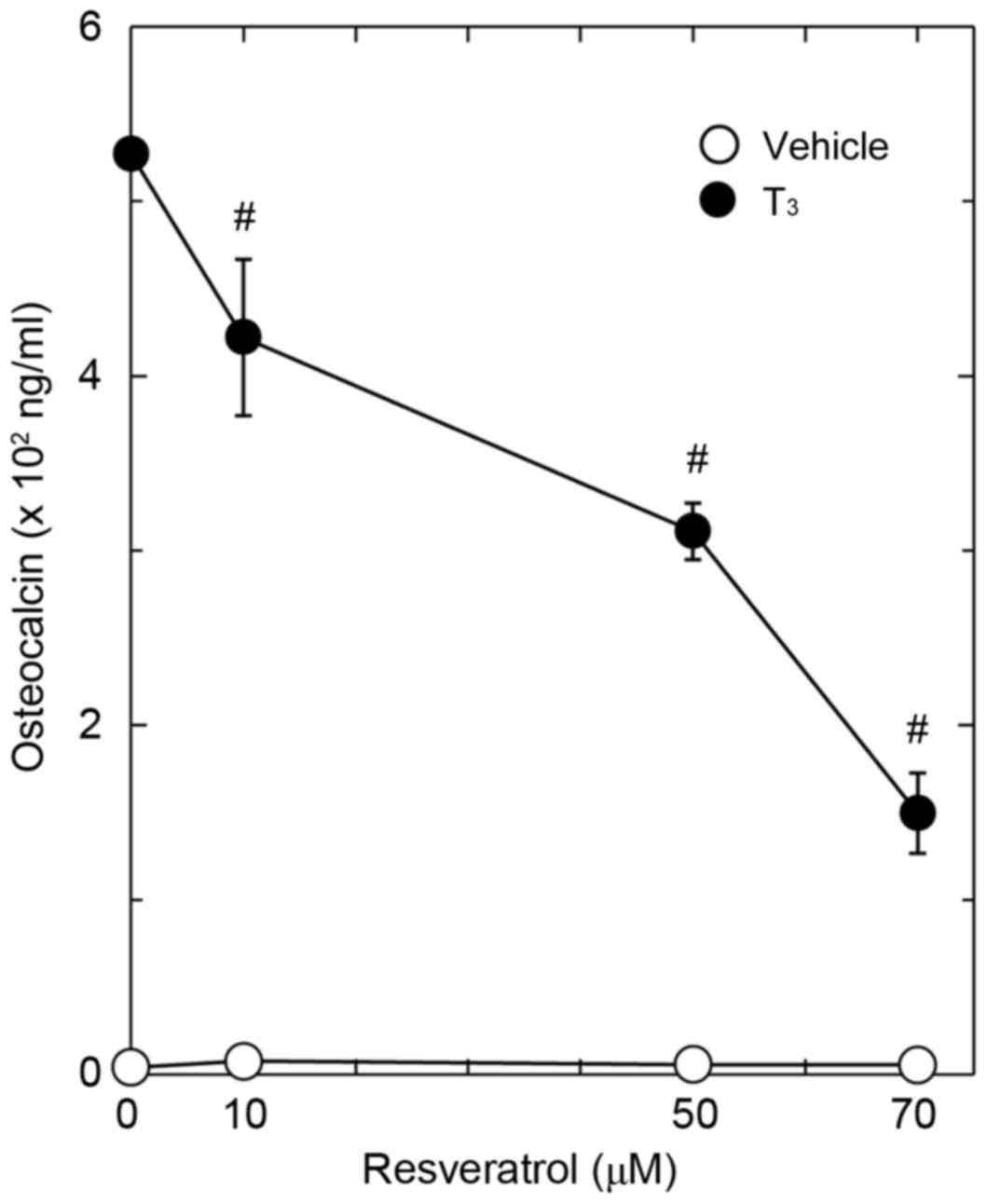
suppressive effect of resveratrol on the T3-stimulated osteocalcin
release was dose-dependent in the range between 10 and 70 µM
(Fig. 2). The maximum effect of
resveratrol was observed at 70 mM, which caused a ~70% decrease in
the T3-effect.
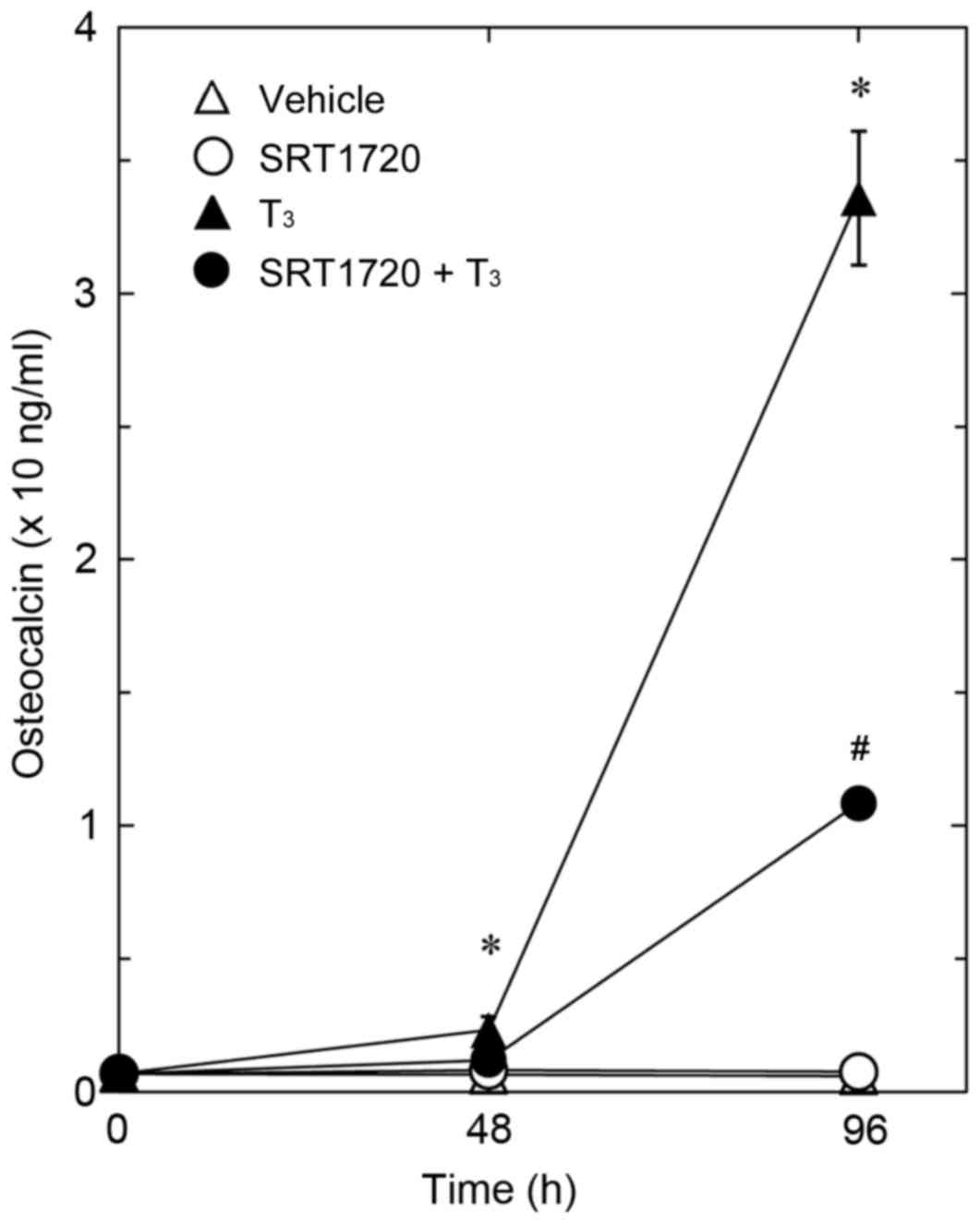
Effect of SRT1720 on the
T3-stimulated osteocalcin release in MC3T3-E1 cells
It has been previously reported that resveratrol
exerts its biological effects through the activation of SIRT1
(1,2). Thus, the present study investigated
the effect of SRT1720, which is a specific and potent synthetic
activator of SIRT1 (34), on
T3-stimulated osteocalcin release in osteoblast-like MC3T3-E1
cells. Similarly to resveratrol, SRT1720 significantly reduced
T3-stimulated osteocalcin synthesis compared with cells treated
with T3 only (Fig. 3).
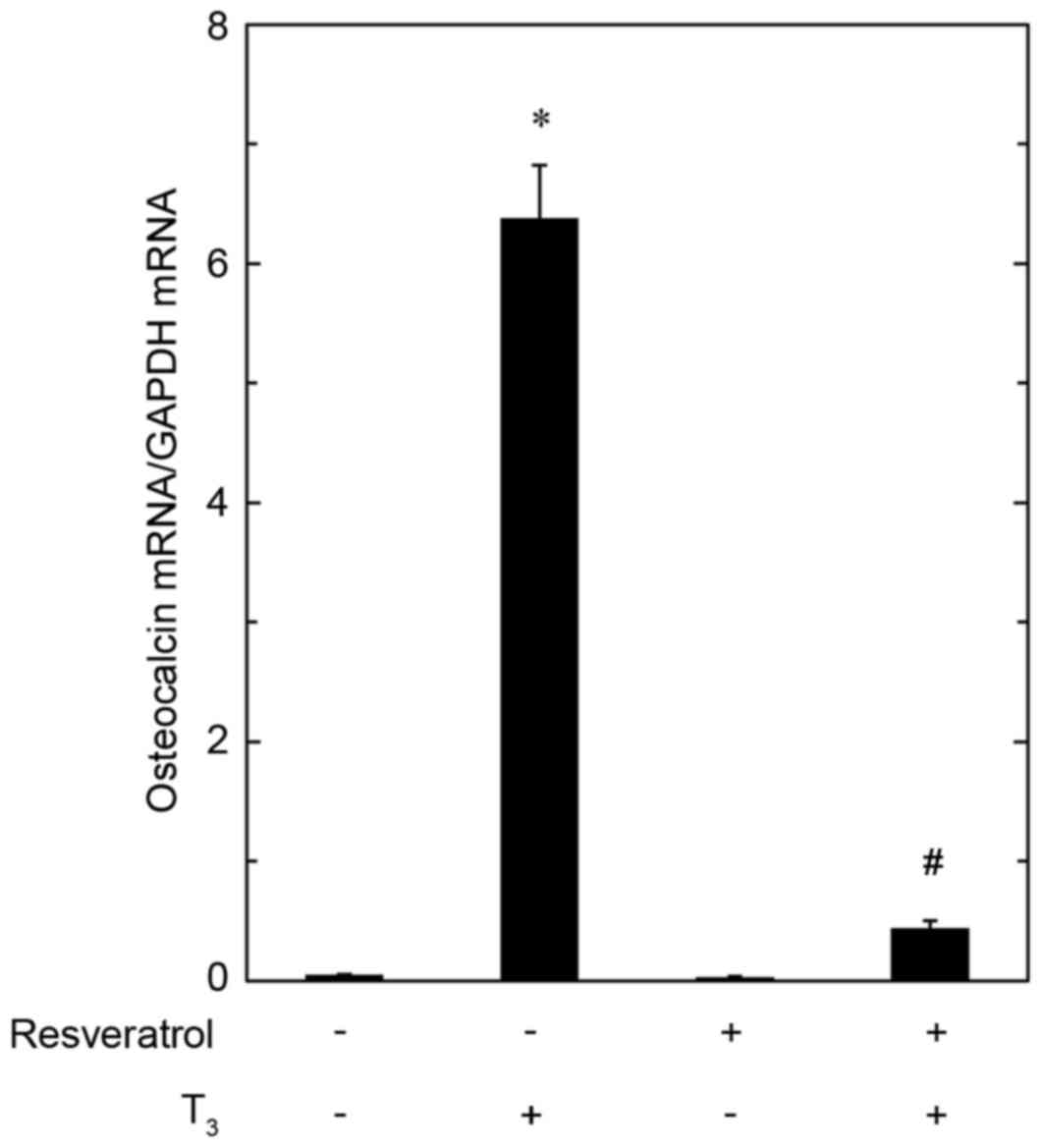
Effect of resveratrol on
T3-induced expression of osteocalcin mRNA in MC3T3-E1
cells
In order to investigate whether the inhibitory
effect of resveratrol on the T3-stimulated osteocalcin release is
exerted through transcriptional events or not, the present study
examined the effect of resveratrol on T3-induced osteocalcin mRNA
expression by RT-qPCR. Resveratrol, which had no effect on basal
levels of osteocalcin mRNA when applied alone, significantly
reduced the expression level of osteocalcin mRNA induced by T3
compared with cells treated with T3 only (Fig. 4).
Effect of resveratrol on
T3-induced transactivation activity of thyroid
hormone-responsive element in MC3T3-E1 cells
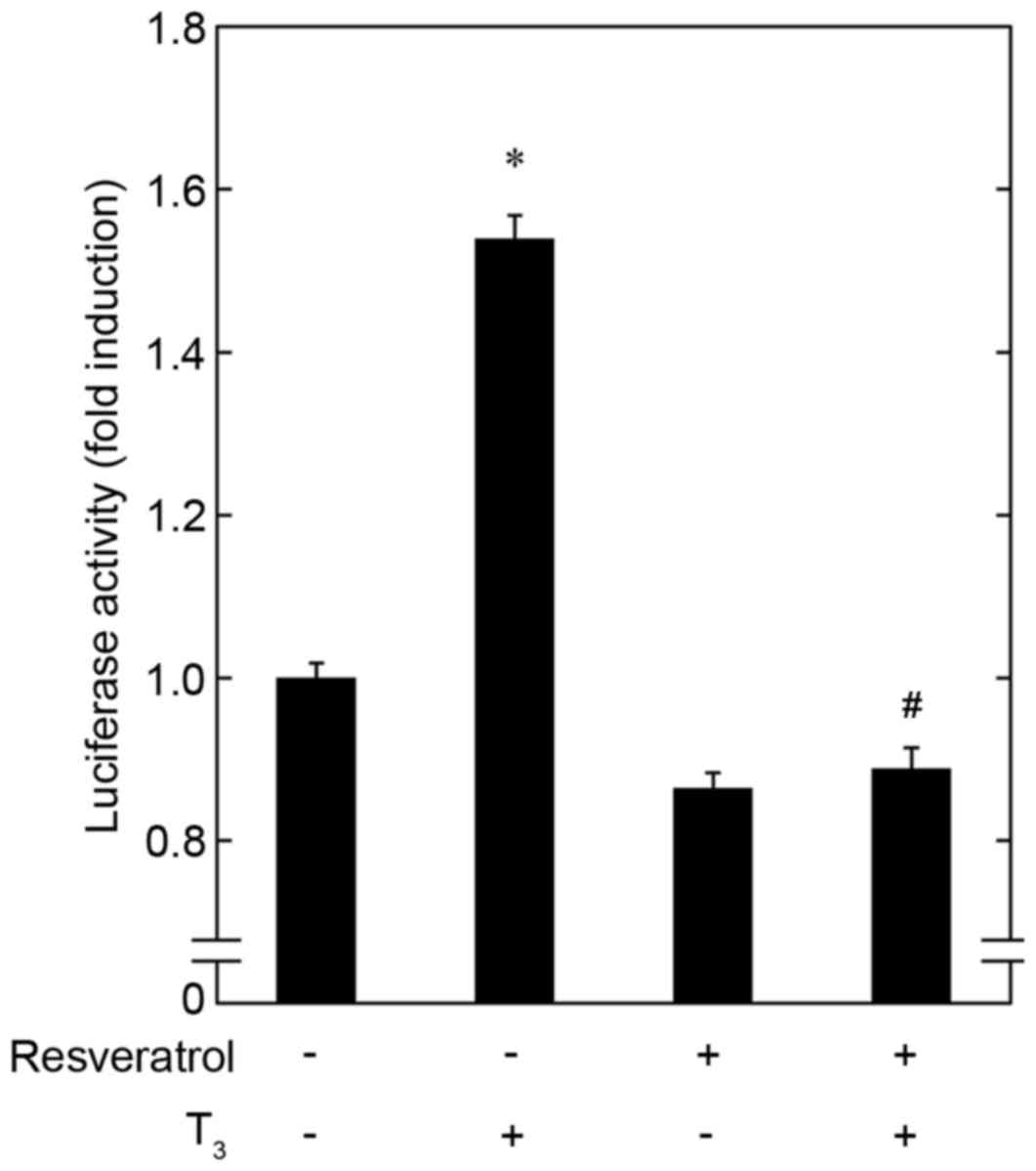
In addition, the effect of resveratrol on
T3-stimulated transactivation activity of thyroid
hormone-responsive element in osteoblast-like MC3T3-E1 cells was
investigated using a luciferase reporter assay. Pretreatment with
resveratrol alone had a limited effect on the luciferase activity
of thyroid hormone-responsive element compared with untreated cells
(Fig. 5). However, resveratrol
significantly inhibited the activity induced by T3 compared with
cells treated with T3 only (Fig.
5).
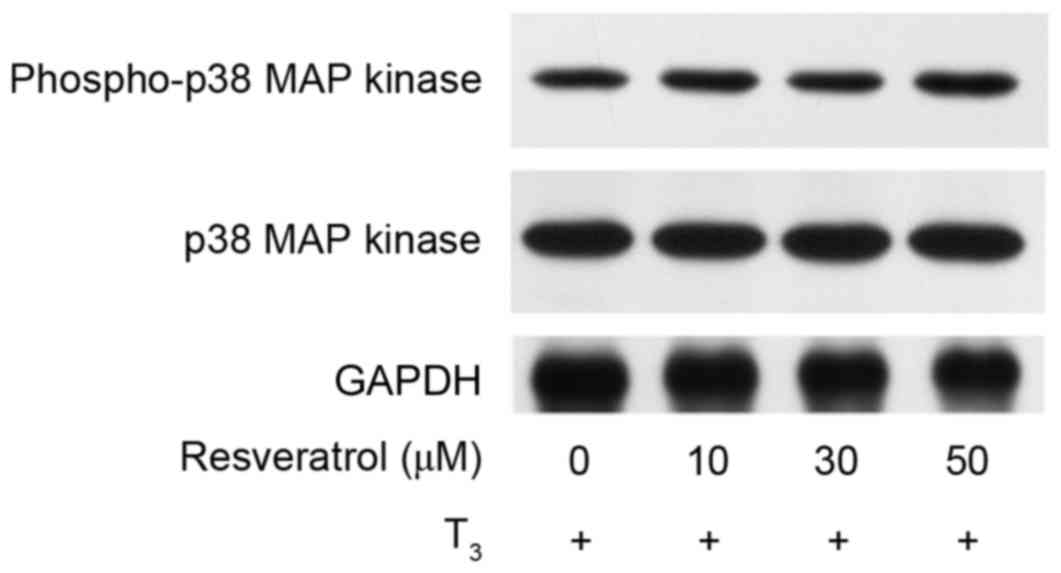
Effect of resveratrol on the
T3-induced phosphorylation of p38 MAP kinase in MC3T3-E1
cells
In our previous study, T3-stimulated osteocalcin
synthesis was demonstrated to be positively regulated through the
p38 MAP kinase pathway in osteoblast-like MC3T3-E1 cells (25). In order to clarify whether the
activation of p38 MAP kinase is implicated in the
resveratrol-effect on T3-induced osteocalcin synthesis in these
cells, the present study investigated the effect of resveratrol on
the T3-induced phosphorylation of p38 MAP kinase. However, the
results demonstrated that resveratrol did not affect the
T3-stimulated phosphorylation of p38 MAP kinase (Fig. 6).
Discussion
The present study demonstrated that resveratrol
significantly decreased T3-stimulated osteocalcin
release in osteoblast-like MC3T3-E1 cells. Additionally, it was
observed that resveratrol significantly reduced osteocalcin mRNA
levels upregulated by T3. Therefore, the inhibitory
effect of resveratrol on the T3-induced osteocalcin
synthesis may occur upstream of transcriptional levels in
osteoblast-like MC3T3-E1 cells. A number of biological effects of
resveratrol are reported to be dependent on SIRT1 activation
(1,2). Therefore, in order to investigate
whether the suppression of T3-stimulated osteocalcin
synthesis by resveratrol is mediated through SIRT1 in MC3T3-E1
cells, the effect of SRT1720, a synthetic compound that activates
SIRT1 with a potency 1,000-fold greater than resveratrol (34), on the release of osteocalcin was
examined. The results demonstrated that SRT1720 mimicked the
suppressive effect of resveratrol on T3-induced release
of osteocalcin. Based on the results, the inhibitory effect of
resveratrol on osteocalcin synthesis induced by T3 may
be mediated, at least partially, by the activation of SIRT1 in
osteoblast-like MC3T3-E1 cells.
It is established that the effects of thyroid
hormone, a member of the nuclear receptor superfamily, are exerted
via its binding to specific receptors in the nucleus, and that the
receptor-hormone complex subsequently activates target gene
expression (35). Therefore, the
present study investigated the effect of resveratrol on
T3-induced transactivation activity of thyroid
hormone-responsive element by a luciferase reporter assay in
MC3T3-E1 cells. Resveratrol significantly reduced the
T3-induced transactivation activity of thyroid
hormone-responsive element, in addition to the expression of
osteocalcin mRNA in MC3T3-E1 cells. The results indicate that the
inhibitory effect of resveratrol on T3-induced
osteocalcin synthesis may be exerted upstream of gene transcription
in osteoblast-like MC3T3-E1 cells.
In the MAP kinase superfamily, it is established
that p44/p42 MAP kinase, p38 MAP kinase and stress-activated
protein kinase/c-Jun N-terminal kinase have pivotal roles in a
variety of cellular functions, including proliferation,
differentiation and survival (36). Our previous studies (25,37)
reported that T3 stimulates the activation of p44/p42
MAP kinase and p38 MAP kinase in osteoblast-like MC3T3-E1 cells,
and that p38 MAP kinase, and not p44/p42 MAP kinase, functions as a
positive regulator in osteocalcin synthesis stimulated by
T3. However, the present study demonstrated that
resveratrol did not affect the T3-induced
phosphorylation of p38 MAP kinase in these cells. Based on the
results, it seems unlikely that the effect of resveratrol on
T3-stimulated osteocalcin synthesis is exerted at a
point upstream of p38 MAP kinase in osteoblast-like MC3T3-E1
cells.
Osteocalcin is synthesized specifically in mature
osteoblasts and stored in the bone matrix (17). The presence of the three Gla
residues is critical for the structure and function of osteocalcin,
which allows it to bind to hydroxyapatite with a high affinity in
their fully carboxylated state, which subsequently regulates the
maintenance of adequate bone mass (18). Therefore, the results of the
present study, which demonstrate suppression of
T3-stimulated osteocalcin synthesis by resveratrol, may
indicate a novel role for the polyphenol in the modulation of bone
metabolism. We have recently demonstrated that resveratrol
modulates the synthesis of OPG stimulated by PGF2α,
PGD2, PGE1, PGE2, FGF-2 or BMP-4,
and regulates VEGF synthesis induced by BMP-4 or TGF-β in
osteoblast-like MC3T3-E1 cells (9–16).
Taking these findings into account, resveratrol may support the
maintenance of skeletal conditions via orchestrating osteoblast
functions elicited by various stimuli, including prostaglandins,
cytokines and growth factors, which may explain the lower hip bone
fracture risk observed in wine drinkers (6). In addition, osteocalcin has recently
been recognized as a potent bone-derived hormone that regulates
energy or lipid metabolism (20).
Based on the results of the current study, as
T3-stimulated osteocalcin synthesis was suppressed by
resveratrol in osteoblast-like MC3T3-E1 cells, resveratrol may
modulate whole body energy metabolism through the regulation of
osteocalcin synthesis in osteoblasts. Further investigation is
required to clarify the exact mechanism underlying the effects of
resveratrol in osteoblasts. In conclusion, the results of the
present study indicate that resveratrol suppresses
T3-stimulated osteocalcin synthesis upstream of
transcription in osteoblasts, and that the inhibitory effect of
resveratrol is partially mediated by SIRT1.
Acknowledgements
We are grateful to Mrs. Yumiko Kurokawa (Department
of Pharmacology, Gifu University Graduate School of Medicine, Gifu,
Japan) for her excellent technical assistance. The present study
was supported in part by a Grant-in-Aid for Scientific Research
(grant no. 19591042) from the Ministry of Education, Culture,
Sports, Science and Technology of Japan, a Grant-in-Aid for
Scientific Research (grant no. H25-Aging-General-004) from the
Ministry of Health, Labour and Welfare of Japan, and the Research
Funding for Longevity Sciences (grant no. 25-4, 26-12) from
National Center for Geriatrics and Gerontology (Obu, Japan).
References
|
1
|
Blander G and Guarente L: The Sir2 family
of protein deacetylases. Annu Rev Biochem. 73:417–435. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Koo SH and Montminy M: In vino veritas: A
tale of two Sirt1s? Cell. 127:1091–1093. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Howitz KT, Bitterman KJ, Cohen HY, Lamming
DW, Lavu S, Wood JG, Zipkin RE, Chung P, Kisielewski A, Zhang LL,
et al: Small molecule activators of sirtuins extend
Saccharomyces cerevisiae lifespan. Nature. 425:191–196.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baur JA, Pearson KJ, Price NL, Jamieson
HA, Lerin C, Kalra A, Prabhu VV, Allard JS, Lopez-Lluch G, Lewis K,
et al: Resveratrol improves health and survival of mice on a
high-calorie diet. Nature. 444:337–342. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Biagi M and Bertelli AA: Wine, alcohol and
pills: What future for the French paradox? Life Sci. 131:19–22.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kubo JT, Stefanick ML, Robbins J,
Wactawski-Wende J, Cullen MR, Freiberg M and Desai M: Preference
for wine is associated with lower hip fracture incidence in
post-menopausal women. BMC Women's Health. 13:362013. View Article : Google Scholar
|
|
7
|
Mizutani K, Ikeda K, Kawai Y and Yamori Y:
Resveratrol stimulates the proliferation and differentiation of
osteoblastic MC3T3-E1 cells. Biochem Biophys Res Commun.
253:859–863. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Suda T, Takahashi N, Udagawa N, Jimi E,
Gillespie MT and Martin TJ: Modulation of osteoclast
differentiation and function by the new members of the tumor
necrosis factor receptor and ligand families. Endocr Rev.
20:345–357. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kuroyanagi G, Tokuda H,
Matsushima-Nishiwaki R, Kondo A, Mizutani J, Kozawa O and Otsuka T:
Resveratrol suppresses prostaglandin F(2α)-induced osteoprotegerin
synthesis in osteoblasts: Inhibition of the MAP kinase signaling.
Arch Biochem Biophys. 542:39–45. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kuroyanagi G, Mizutani J, Kondo A,
Yamamoto N, Matsushima-Nishiwaki R, Otsuka T, Kozawa O and Tokuda
H: Suppression by resveratrol of prostaglandin D2-stimulated
osteoprotegerin synthesis in osteoblasts. Prostaglandins Leukot
Essent Fatty Acids. 91:73–80. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yamamoto N, Otsuka T, Kuroyanagi G, Kondo
A, Kainuma S, Nakakami A, Matsushima-Nishiwaki R, Kozawa O and
Tokuda H: Resveratrol reduces prostaglandin E1-stimulated
osteoprotegerin synthesis in osteoblasts: Suppression of
stress-activated protein kinase/c-Jun N-terminal kinase.
Prostaglandins Other Lipid Mediat. 116–117:57–63. 2015. View Article : Google Scholar
|
|
12
|
Yamamoto N, Tokuda H, Kuroyanagi G,
Mizutani J, Matsushima-Nishiwaki R, Kondo A, Kozawa O and Otsuka T:
Regulation by resveratrol of prostaglandin E2-stimulated
osteoprotegerin synthesis in osteoblasts. Int J Mol Med.
34:1439–1445. 2014.PubMed/NCBI
|
|
13
|
Kuroyanagi G, Otsuka T, Yamamoto N,
Matsushima-Nishiwaki R, Nakakami A, Mizutani J, Kozawa O and Tokuda
H: Down-regulation by resveratrol of basic fibroblast growth
factor-stimulated osteoprotegerin synthesis through suppression of
Akt in osteoblasts. Int J Mol Sci. 15:17886–17900. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kuroyanagi G, Tokuda H, Yamamoto N,
Matsushima-Nishiwaki R, Mizutani J, Kozawa O and Otsuka T:
Resveratrol amplifies BMP-4-stimulated osteoprotegerin synthesis
via p38 MAP kinase in osteoblasts. Mol Med Rep. 12:3849–3854.
2015.PubMed/NCBI
|
|
15
|
Kondo A, Otsuka T, Kuroyanagi G, Yamamoto
N, Matsushima-Nishiwaki R, Mizutani J, Kozawa O and Tokuda H:
Resveratrol inhibits BMP-4-stimulated VEGF synthesis in
osteoblasts: Suppression of S6 kinase. Int J Mol Med. 33:1013–1018.
2014.PubMed/NCBI
|
|
16
|
Kuroyanagi G, Otsuka T, Yamamoto N,
Matsushima-Nishiwaki R, Kozawa O and Tokuda H: Resveratrol
suppresses TGF-β-induced VEGF synthesis in osteoblasts: Inhibition
of the p44/p42 MAPKs and SAPK/JNK pathways. Exp Ther Med.
9:2303–2310. 2015.PubMed/NCBI
|
|
17
|
Hauschka PV, Lian JB, Cole DE and Gundberg
CM: Osteocalcin and matrix Gla protein: Vitamin K-dependent
proteins in bone. Physiol Rev. 69:990–1047. 1989.PubMed/NCBI
|
|
18
|
Ducy P, Desbois C, Boyce B, Pinero G,
Story B, Dunstan C, Smith E, Bonadio J, Goldstein S, Gundberg C, et
al: Increased bone formation in osteocalcin-deficient mice. Nature.
382:448–452. 1996. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee NK and Karsentry G: Reciprocal
regulation of bone and energy metabolism. Trends Endocrinol Metab.
19:161–166. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Oldknow KJ, MacRae VE and Farquharson C:
Endocrine role of bone: Recent and emerging perspectives beyond
osteocalcin. J Endocrinol. 225:R1–R19. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gogakos AI, Bassett JH Duncan and Williams
GR: Thyroid and bone. Arch Biochem Biophys. 503:129–136. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gorka J, Taylor-Gjevre RM and Arnason T:
Metabolic and clinical consequences of hyperthyroidism on bone
density. Int J Endocrinol. 2013:6387272013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vestergaard P and Mosekilde L:
Hyperthyroidism, bone mineral, and fracture risk-a meta-analysis.
Thyroid. 13:585–593. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cheng SY, Leonard JL and Davis PJ:
Molecular aspects of thyroid hormone actions. Endocr Rev.
31:139–170. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ishisaki A, Tokuda H, Yoshida M, Hirade K,
Kunieda K, Hatakeyama D, Shibata T and Kozawa O: Activation of p38
mitogen-activated protein kinase mediates thyroid
hormone-stimulated osteocalcin synthesis in osteoblasts. Mol Cell
Endocrinol. 214:189–195. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sudo H, Kodama HA, Amagai Y, Yamamoto S
and Kasai S: In vitro differentiation and calcification in a new
clonal osteogenic cell line derived from newborn mouse calvaria. J
Cell Biol. 96:191–198. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kozawa O, Tokuda H, Miwa M, Kotoyori J and
Oiso Y: Cross-talk regulation between cyclic AMP production and
phosphoinositide hydrolysis induced by prostaglandin E2 in
osteoblast-like cells. Exp Cell Res. 198:130–134. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang W, Yang N and Shi XM: Regulation of
mesenchymal stem cell osteogenic differentiation by
glucocorticoid-induced leucine zipper (GILZ). J Biol Chem.
283:4723–4729. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Simpson DA, Feeney S, Boyle C and Stitt
AW: Retinal VEGF mRNA measured by SYBR Green I fluorescence: A
versatile approach to quantitative PCR. Mol Vis. 6:178–183.
2000.PubMed/NCBI
|
|
30
|
Pryor RJ and Wittwer CT: Real-time
polymerase chain reaction and melting curve analysis. Methods Mol
Biol. 336:19–32. 2006.PubMed/NCBI
|
|
31
|
Pfaffl MW: A new mathematical model for
relative quantification in real-time RT-PCR. Nucleic Acids Res.
29:e452001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kato K, Ito H, Hasegawa K, Inaguma Y,
Kozawa O and Asano T: Modulation of the stress-induced synthesis of
hsp27 and alpha B-crystallin by cyclic AMP in C6 rat glioma cells.
J Neurochem. 66:946–950. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Milne JC, Lambert PD, Schenk S, Carney DP,
Smith JJ, Gagne DJ, Jin L, Boss O, Perni RB, Vu CB, et al: Small
molecule activators of SIRT1 as therapeutics for the treatment of
type 2 diabetes. Nature. 450:712–716. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mullur R, Liu YY and Brent GA: Thyroid
hormone regulation of metabolism. Physiol Rev. 94:355–382. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kyriakis JM and Avruch J: Mammalian
mitogen-activated protein kinase signal transduction pathways
activated by stress and inflammation. Physiol Rev. 81:807–869.
2001.PubMed/NCBI
|
|
37
|
Kozawa O, Hatakeyama D, Yoshida M, Kamiya
Y, Kondo C, Matsuno H and Uematsu T: Activation of p44/p42
mitogen-activated protein kinase limits triiodothyronine-stimulated
alkaline phosphatase activity in osteoblasts. Biochem Biophys Res
Commun. 286:1140–1143. 2001. View Article : Google Scholar : PubMed/NCBI
|