Embryonic stem cells (ESCs) are characterized by
unlimited proliferation (self-renewal) and the ability to
differentiate into three primary germ layers, namely the endoderm,
mesoderm and ectoderm (pluripotency) (1–4). It
has been established that complicated regulatory networks are
present in ESCs that critically maintain the state of self-renewal
and pluripotency for later development (5,6).
Several transcription factors (TFs), including octamer-binding
transcription factor 4 (OCT4), SRY-box 2 (SOX2) and homeobox
protein NANOG (NANOG) are known to sit at the top of the regulatory
hierarchy, regulating the expression of various downstream target
genes (7,8). Among them, OCT4 serves an
indispensable role in maintaining the pluripotency of ESCs
(9,10) and in reprogramming the
terminally-differentiated somatic cells back into the ESC-like
cells (11–13). Furthermore, OCT4 can mediate the
differentiation of murine ESCs induced by retinoic acid or
Wnt/β-catenin in a manner that is independent of and distinct from
other core TFs (14), indicating
that OCT4 may have unique and non-substitutable roles in
controlling the self-renewal, pluripotency and differentiation of
ESCs.
Cell cycle progression is required for ESCs to
proliferate and avoid staying in a quiescent state. Multiple
studies have demonstrated that cell cycle-associated proteins can
regulate various core TFs or differentiation markers (15). In a reciprocal manner, several TFs,
such as NANOG and c-MYC proto-oncogene protein, can control the
expression levels of multiple cell cycle-associated target genes
(16,17). This review will be focused on
reciprocal interplays between OCT4 and cell cycle checkpoints and
their connections with the ESC pluripotency.
Cell cycle comprises four different phases; the S
phase for DNA replication, the M phase for cell mitosis, and two
gap phases between S phase and M phase (G1 phase for synthesis of
proteins and lipids, and G2 phase for checking DNA integrity).
Ample evidence has revealed that the duration of cell cycle in
murine somatic cells is relatively long (>16 h), which is
dominated by the G1 phase (18);
in contrast, the cell cycle of murine ESCs progresses faster (~8–10
h) (19), which is characterized
by a truncated G1 phase and a prolonged S phase (20). Although the duration of cell cycle
in human ESCs is significantly lengthened (~32–38 h) (21), the time spent at G1 phase is
minimal (3 h in human ESCs vs. 10 h in human somatic cells)
(15,22), indicating that the cell cycle
dynamics may crucially impact on the differentiation potential of
pluripotent stem cells. Indeed, ~1–5% of the total proteins differ
their expression levels between ESCs and induced pluripotent (iPS)
cells, and the majority of them are cell cycle proteins (23).
There is mounting evidence demonstrating that
lengthening the G1 phase in ESCs contributes to inducing
differentiation (24–27), and distinct G1 phase profiles will
lead to different lineage fates. Human ESCs in early G1 phase can
only differentiate into endoderm, whereas in late G1 phase they
were limited to neuroectodermal differentiation (28). In fact, all-trans retinoic acid, a
common differentiation inducer, can regulate the gene expression of
Cyclin D1 (29,30) and result in G1 phase accumulation
(31–33). It is therefore reasonable to
propose that during the G1 phase, ESCs sense and integrate various
extracellular and intracellular signals to make the decisions on
the timing and the fate of differentiation. A shortened G1 phase
may minimize the exposure of ESCs to various signals, thereby
preserving their pluripotency. In addition, it was demonstrated in
a recent study that G2 cell cycle arrest is also required for
endodermal development (34);
furthermore, specific disruption of S and G2 phases will affect the
pluripotent state of human ESCs in a G1 phase-independent way
(35–37). Gamma-ray-induced DNA damage induces
G2/M blockage and the differentiation of ESCs (38,39).
It is important that ESCs have a long enough G2 phase to check and
restore the fidelity of the genome as a result of G1/S checkpoint
deficiency.
The expression of Cyclin-dependent kinase 4/6
(CDK4/6) and Cyclin D is increased in early G1 phase in somatic
cells. Although the lack of Cyclin D expression was reported in
murine ESCs (40), the mRNA levels
of CDK4 and Cyclin D2 were increased in human ESCs (22,41).
Further studies demonstrated that Cyclin D expression is enhanced
in late G1 and G1/S phases in human ESCs. Notably, knocking down
Cyclin D induces endodermal differentiation, whereas its
overexpression promoted neuroectodermal differentiation by
inhibiting mothers against decapentaplegic (SMAD) 2/3 nuclear
translocation (28). In addition,
Cyclin D can also recruit transcriptional co-regulators to
development-assocaited gene loci and modify the epigenetics of
target genes (42). There is
evidence demonstrating that a proper level of Cyclin D is necessary
for maintaining the pluripotent state of ESCs, while overexpression
of them may induce reprogramming of epidermal cells into stem-like
cells with higher expression levels of OCT4 and NANOG (43). In contrast, in adult stem cells or
cancer cells, OCT4 can directly bind to the promoter region of
Cyclin D1, thereby regulating its transcription and controlling
G1/S transition (44–46). Meanwhile, OCT4 can bind with the
conserved promoter of microRNA (miR)-302 (47), increasing the level of
p16(Ink4a)/p19(Ink4d) and inhibiting the interaction between CDK4/6
and Cyclin D (48). Furthermore,
OCT4 can also interact with SMAD2/3 to control the pluripotent
state of ESCs (49,50). Taken together, these studies
suggested that OCT4 is involved in the transcriptional regulation
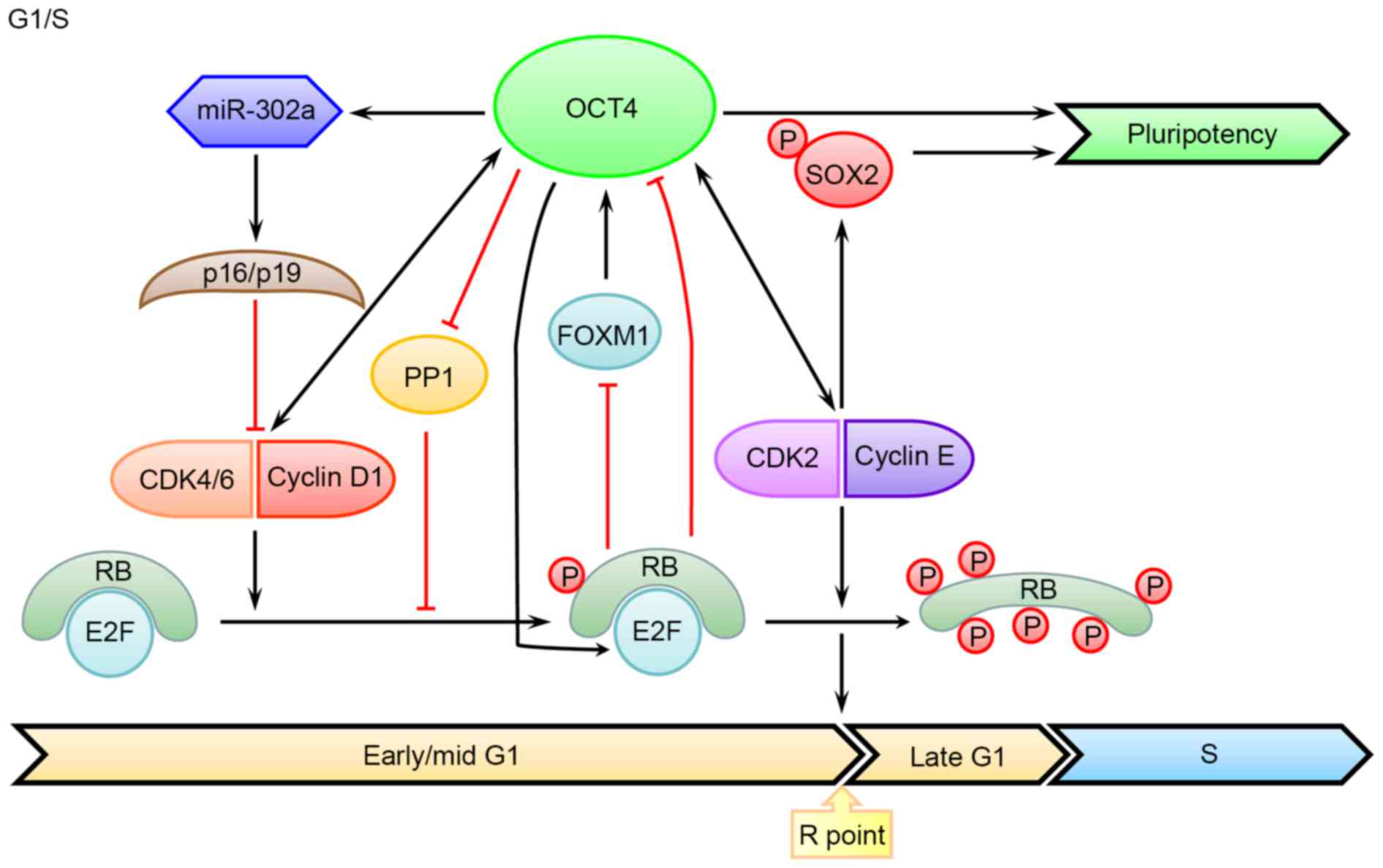
of Cyclin D as well as other target genes (Fig. 1).
CDK2-Cyclin E is constitutively expressed and
involved in the progression of G1/S transition (26). In human ESCs, inhibition of CDK2
will lead to G1 phase arrest, which is accompanied with apoptosis
or differentiation. Inhibition of CDK2 can induce sustained genomic
damage and elicit DNA damage response, thus contributing to
apoptosis of impaired ESCs (51,52).
As demonstrated in further studies, OCT4 expression can be
suppressed by downregulating CDK2 (53,54),
while CDK2 can enhance reprogramming efficiency by phosphorylating
SOX2 at Ser-39 and Ser-253 sites (55). Although the regulation of
CDK2-Cyclin A/E by OCT4 in ESCs has not been reported, OCT4 can
promote tumor proliferation by activating Cyclin E (56). Thus, it remains possible that OCT4
may regulate the expression of CDK2-Cyclin A/E in ESCs.
Retinoblastoma (RB) protein is a downstream target
of CDK4/6-Cyclin D, which can inhibit the transcription activity of
E2F transcription factor 1 (E2F) in its hypophosphorylated state.
After being hyperphosphorylated by CDK2-Cyclin E, RB can release
E2F for the ultimate regulation of a number of targets involved in
G1 phase progression and S phase entry (Fig. 1). Therefore, it came as no surprise
that the activity of RB-E2F can influence the ESC self-renewal and
pluripotency (57,58). In fact, activated RB can directly
bind to the promoter regions of OCT4 and SOX2, leading to their
transcriptional suppression and a declined reprogramming efficiency
(59); in contrast, the inactive
RB allows for generation of iPS cells in the absence of exogenous
SOX2 expression (60).
Furthermore, RB can also regulate OCT4 level by suppressing the
expression of forkhead box protein M1, which is a transcription
factor promoting OCT4 expression (61,62).
In addition, E2f will switch from an active state in stem cells to
a suppressed state in differentiated cells through forming a
complex with RB (63). Conversely,
in murine ESCs, OCT4 maintains the hypo-phosphorylated state of RB
by inhibiting the activity of protein phosphatase 1 (64), which is well-known for its role in
triggering mitotic exit (65).
Additionally, OCT4 can also directly bind to the promoter region of
E2f3a and increase its expression level in murine ESCs, which
contributes to relieving the cell growth retardation caused by OCT4
knockdown (66). As inhibition of
E2F2 can impair self-renewal and cell cycle progression in human
ESCs, the pluripotency is preserved in E2F2 silencing cells
(67). Therefore, the effects of
RB on the pluripotency of ESCs are unlikely mediated by E2F. The
other roles of RB in ESCs will be discussed later.
In somatic cells, CDK1-Cyclin A/B is a critical cell
cycle regulator that can promote G2/M transition. As has been
demonstrated in multiple studies, CDK1-Cyclins serve critical roles
in the self-renewal and development of ESCs. The expression level
of Cyclin A, the first cloned Cyclin protein, is higher in ESCs in
G2 phase than that in fibroblast cells (68), and resetting its expression level
in early-passage iPS cells can improve the pluripotency and reduce
the tumorigenicity (23). In
addition, the Cyclin B1 level is also upregulated in ESCs in G2
phase compared with that in somatic cells. Increased expression of
Cyclin B1 in G2 phase can delay the dissolution of pluripotent
state in human ESCs, while knockdown of Cyclin B1 induces markedly
declined expression of pluripotent markers in human ESCs (36). The same is true for CDK1. In human
ESCs, down-regulating CDK1 leads to loss of pluripotency, increased
differentiation markers, accumulation of double-strand breaks, as
well as the inability to arrest at G2 phase and commit to apoptosis
(69,70). CDK1 can enhance the binding of OCT4
to the promoter and suppress the transcription of homeobox protein
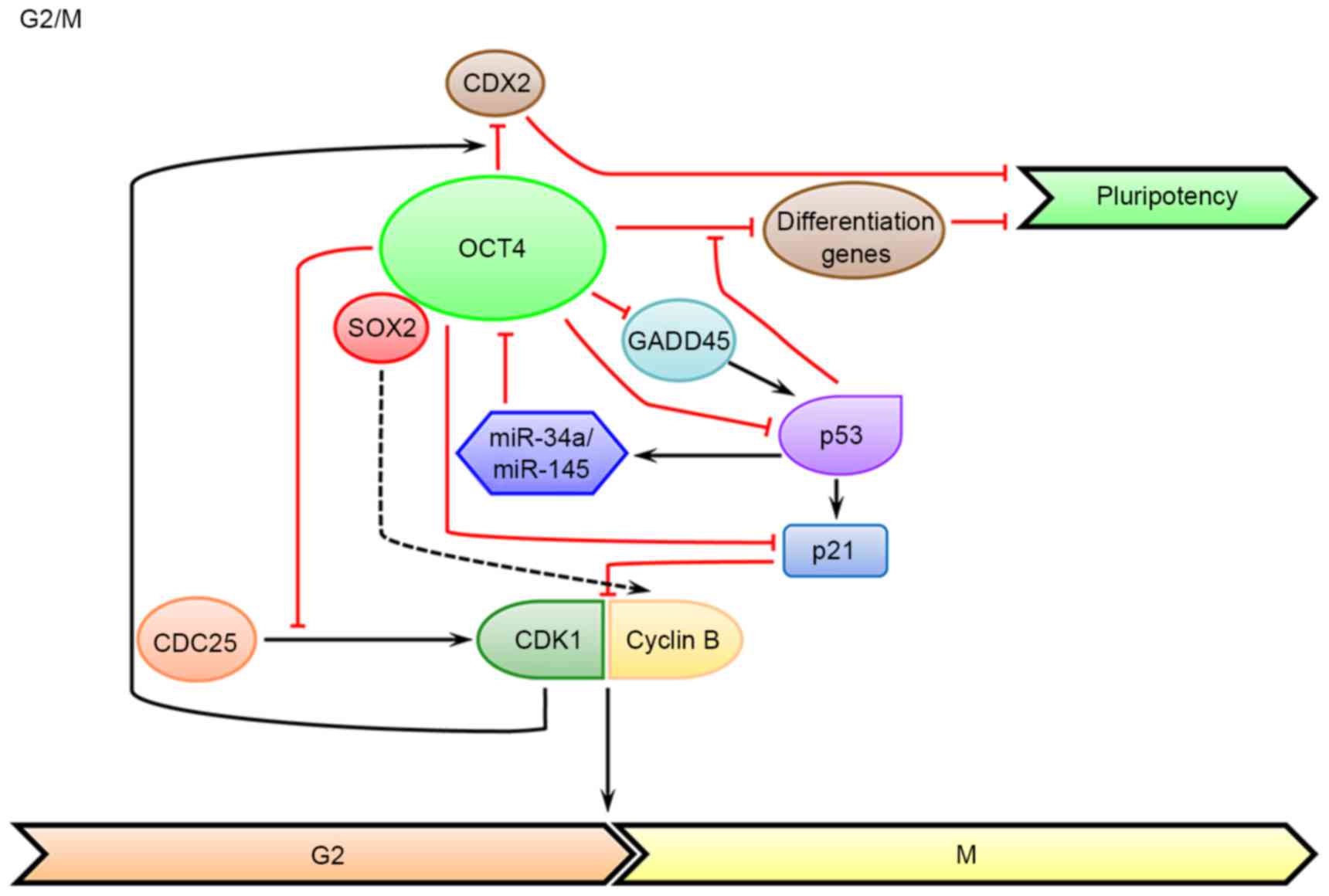
CDX2, a classic differentiation marker (71). Furthermore, several markers of G2/M
are expressed during the meso- and endodermal differentiation
(e.g., WEE1 G2 checkpoint kinase blocks entry into mitosis by
phosphorylating CDK1 at Y15), rather than the ectodermal
differentiation (34). In
contrast, OCT4 can inhibit the activation of CDK1 by cell division
cycle 25 phosphorylation, which is independent of its
transcriptional activity (Fig. 2).
Thus, ESCs have to express more CDK1 to overcome the inhibitory
effect of OCT4. Inhibition of CDK1 by OCT4 will lead to a prolonged
duration of G2 phase, which allows for subsequent checking of
genome integrity and reducing chromosomal mis-segregation (72). Indeed, inhibition of CDK1 can
activate the response to DNA damage and promote nuclear
translocation and activation of p53, thereby maintaining the
survival of ESCs (73). The
potential connection between OCT4 and Cyclin A/B has not been
elucidated in any study yet, but there is evidence that SOX2, a
core TF frequently associated with OCT4, can promote the expression
of Cyclin A/B in cancer cells (74–76).
The direct regulation of CDK1-Cyclin by OCT4 warrants further
investigation.
Growth arrest and DNA-damage-inducible protein 45
(GADD45), which includes several isoforms, is crucial for
protecting genome stability in G2/M transition by suppressing cell
cycle and repairing DNA. GADD45ag morpholino knockdown in
Xenopus can induce differentiation of neural embryonic cells
by inducing various cell cycle related inhibitors, such as p53, p21
and Cyclin G1. Additionally, GADD45ag morphants exhibit increased
expression of Xenopus OCT4 homologs, indicating that
GADD45ag is required for early embryonic cells to exit pluripotency
and enter differentiation (77).
In addition, GADD45a can bind to the OCT4 promoter and promote its
demethylation in Xenopus oocytes, which is accompanied with
DNA repair (78,79). Furthermore, studies in human cells
indicated that GADD45 G is a downstream target of OCT4, which is
significantly increased in the OCT4 knockdown system (80,81).
As discussed above, RB is a tumor-suppressor gene
controlling the activity of transcription factor of E2F family,
which serves an indispensable role in G1/S transition. Increased
activity of RB can trigger cell cycle arrest, differentiation or
death of ESCs (82). However, the
inactivation of RB family in ESCs can also induce G2/M arrest and
cell death (57), which may be
attributed to the loss of its function in maintaining the genetic
stability (83–85). These findings indicated that the
expression level of RB needs to be tightly controlled at a proper
level, so that the pluripotency and self-renewal of ESCs can be
maintained. Furthermore, overexpression of RB in S phase can lead
to G2 phase arrest (86).
Additionally, RB can directly bind to cohesin and condensin II,
which can regulate centromere functions and control mitosis
(87–91).
The p53-p21 signaling pathway is a major checkpoint
in cell cycle of G1/S and G2/M transition. The expression level of
p53 is kept low in ESCs, which is predominantly present in the
cytoplasm. The extremely low level of p53 in the nucleus is also
inactivated. p53 will translocate to cell nucleus and initiate the
transcription of its target genes in the event of DNA damage
(92). In addition, p53 can
promote the translocation of active Bcl-2-associated X protein from
the Golgi to mitochondria to initiate apoptosis under DNA damage
stresses (93). It is demonstrated
that p53 deficiency will lead to genomic instability in ESCs
(94). In contrast, the activated
p53 in ESCs will result in differentiation (31,95,96)
or apoptosis (73,97). However, it has also been
demonstrated in other studies that p53 has anti-differentiation
effects in ESCs (98), indicating
that p53 exerts its functions in a context-dependent manner, and
that proper intracellular levels and subcellular localization of
p53 are critical for its roles in maintaining the pluripotent state
in ESCs.
In addition, p53 can regulate the expression of
various key TFs in ESCs. For example, knockdown of p53 can lead to
downregulated NANOG expression (99). As a common differentiation inducer
of ESCs, p53 expression is activated after exposure to retinoic
acid, which drives the expression of miR-34a and miR-145 and
reduces the OCT4 expression (31).
In addition, the differentiation-activated p53 can recruit UTX and
lysine-specific demethylase 6B (JMJD3), the H3K27me3-specific
demethylases, bind to the promoter regions of developmental
transcription factors that are repressed by OCT4, and increase the
expression of various differentiation genes (100). p53 is also the downstream target
of OCT4 (Fig. 2). Studies have
revealed that silencing OCT4 will lead to p53 activation and induce
differentiation (101–103). For instance, silencing OCT4
significantly reduces the expression of SIRT1, a deacetylase known
to inhibit p53 activity and the differentiation of ESCs, leading to
increased acetylation of p53 at lysine 120 and 164 that is required
for its stabilization and functionality (104). In addition, OCT4 can bind to the
promoter region of CD49f (integrin subunit α6), which can also
decrease the level of p53 (105).
p21, a downstream target of p53, can inhibit the
activation of CDKs and result in cell cycle arrest (Fig. 2); in addition, it can also be
regulated in a p53-independent way. It has been revealed in studies
that p21 is involved in DNA repair, transcriptional regulation,
differentiation and apoptosis. In ESCs, the expression level of p21
is compromised due to epigenetic modification (106), and the lack of p21 function is
required for maintaining the pluripotent state (107). Ionizing radiation-induced DNA
damage can lead to elevated p21 mRNA level and cell cycle arrest at
G2 phase (108). Upregulation of
p21 in human ESCs will induce G1 phase arrest and subsequent
differentiation into multiple lineages (109). This result is consistent with the
finding that p21 has multiple fuctions in both G1/S and G2/M
checkpoints (110,111). p21 can also mediate apoptosis in
murine ESCs that are exposed to dihydrolipoic acid (112). In addition, increased p21
expression leads to decreased reprogramming efficiency in somatic
cells (113). Conversely, OCT4
can inhibit the activity of p21 by directly binding to its promoter
region or by indirectly up-regulating DNA
(cytosine-5)-methyltransferase 1, a DNA methyltransferase, which
can inhibit lineage differentiation (114–116).
A large amount of energy is generated in ESCs to
meet the requirements for biosynthesis and cell cycle progression.
The energy metabolism mode of primed ESCs is similar to that of
other adult stem cells or cancer cells with a high glycolytic flux
rather than oxidative phosphorylation (OXPHOS), which is known as
the ‘Warburg effect’ (117–121). This phenomenon can be partly
attributed to the immature structure and function of mitochondria
and a hypoxic niche (5% of physiological level) (122,123). Though glycolysis produces less
ATPs than OXPHOS, it has faster rate of ATP generation, which makes
it competent to support active cell proliferation. Additionally,
pyruvate, the product of glycolysis, together with other
intermediate products of tricarboxylic acid (TCA) cycle, can be
used for biosynthesis (such as DNA, protein and lipid) in ESCs as
well as in cancer cells for shortening the G1 phase (123–125). A high glycolytic flux metabolism
in hypoxia may reduce the damages to DNA caused by reactive oxide
species (ROS), which may impair the pluripotency ESCs and induce
their differentiation (126,127).
Initial evidence indicated OCT4 may be involved in
regulating metabolism as its knockdown resulted in increases in TCA
cycle activity and decreases in glycolytic flux (117). Further studies demonstrated that
OCT4 can directly regulate the transcription of hexokinase 2 (HK2)
and pyruvate kinase (PK) M2, the two key glycolytic enzymes that
determine the rate of glycolysis. Overexpression of HK2 and PKM2
contributes to sustaining the high glycolysis level and preserving
the pluripotency of ESCs (128).
Notably, PKM2 can directly bind to OCT4 and enhance OCT4-mediated
transcription (129,130).
It has been known for a while that ESCs are
characterized by an abbreviated G1 phase and a prolonged S phase.
However, the underlying mechanisms remain largely elusive. Emerging
evidence has implicated a direct role of the master pluripotency
factor OCT4 in controlling the transcription of several key cell
cycle regulators. In general, OCT4 appears to directly or
indirectly activate the transcription of cell cycle machineries
that promote G1/S transition and avoid differentiation (Fig. 1). Meanwhile, by suppressing
multiple cell cycle genes, OCT4 controls proper duration of G2
phase to ensure the genomic integrity via both the
transcription-dependent and -independent mechanisms (Fig. 2). Reciprocally, the cell cycle
regulators especially CDK1 can directly interact with OCT4 and
promote its suppressive binding to the differentiation genes and
thereby maintaining the ESC pluripotency.
Another important feature of ESCs is their high
glycolytic metabolism under hypoxic conditions that may minimize
the oxidative damage of ROS to genetic material. Recent studies
revealed that OCT4 can promote glycolysis by transcriptionally
upregulating the expression of several key glycolytic enzymes,
directly linking ESC metabolism to their self-renewal and
pluripotency. Given the convergence of ESC pluripotency and cell
cycle control on OCT4, it would be of interest to investigate in
future studies how OCT4 and other master pluripotency factors
coordinate ESC metabolism with their cell cycle progression.
The rapid cell cycle progression of ESCs requires
high-fidelity DNA replication and repair mechanisms. The
investigation into the potential connection between ESC cell cycle
control and DNA replication/repair is just at its infancy, and it
remains to be seen if the master pluripotency factors such as OCT4
may also serve a role in these events.
The present review was supported by the National Key
Research and Development Program of China (grant no.
2016YFA0100303) and the National Natural Science Foundation of
China (grant no. 31601103).
|
1
|
Wu J and Belmonte JC Izpisua: Dynamic
pluripotent stem cell states and their applications. Cell Stem
Cell. 17:509–525. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Smith AG: Embryo-derived stem cells: Of
mice and men. Annu Rev Cell Dev Biol. 17:435–462. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thomson JA, Itskovitz-Eldor J, Shapiro SS,
Waknitz MA, Swiergiel JJ, Marshall VS and Jones JM: Embryonic stem
cell lines derived from human blastocysts. Science. 282:1145–1147.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Martin GR: Isolation of a pluripotent cell
line from early mouse embryos cultured in medium conditioned by
teratocarcinoma stem cells. Proc Natl Acad Sci USA. 78:7634–7638.
1981; View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yousefi M, Hajihoseini V, Jung W,
Hosseinpour B, Rassouli H, Lee B, Baharvand H, Lee K and Salekdeh
GH: Embryonic stem cell interactomics: The beginning of a long road
to biological function. Stem Cell Rev. 8:1138–1154. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang L and Chen YG: Signaling control of
differentiation of embryonic stem cells toward mesendoderm. J Mol
Biol. 428:1409–1422. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Martello G and Smith A: The nature of
embryonic stem cells. Annu Rev Cell Dev Biol. 30:647–675. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Das S and Levasseur D: Transcriptional
regulatory mechanisms that govern embryonic stem cell fate. Methods
Mol Biol. 1029:191–203. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nichols J, Zevnik B, Anastassiadis K, Niwa
H, Klewe-Nebenius D, Chambers I, Schöler H and Smith A: Formation
of pluripotent stem cells in the mammalian embryo depends on the
POU transcription factor Oct4. Cell. 95:379–391. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Niwa H, Miyazaki J and Smith AG:
Quantitative expression of Oct-3/4 defines differentiation,
dedifferentiation or self-renewal of ES cells. Nat Genet.
24:372–376. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Takahashi K and Yamanaka S: Induction of
pluripotent stem cells from mouse embryonic and adult fibroblast
cultures by defined factors. Cell. 126:663–676. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu T, Wang H, He J, Kang L, Jiang Y, Liu
J, Zhang Y, Kou Z, Liu L, Zhang X and Gao S: Reprogramming of
trophoblast stem cells into pluripotent stem cells by Oct4. Stem
Cells. 29:755–763. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsai SY, Bouwman BA, Ang YS, Kim SJ, Lee
DF, Lemischka IR and Rendl M: Single transcription factor
reprogramming of hair follicle dermal papilla cells to induced
pluripotent stem cells. Stem Cells. 29:964–971. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Simandi Z, Horvath A, Wright LC,
Cuaranta-Monroy I, De Luca I, Karolyi K, Sauer S, Deleuze JF, Gudas
LJ, Cowley SM and Nagy L: OCT4 Acts as an integrator of
pluripotency and signal-induced differentiation. Mol Cell.
63:647–661. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kareta MS, Sage J and Wernig M: Crosstalk
between stem cell and cell cycle machineries. Curr Opin Cell Biol.
37:68–74. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bretones G, Delgado MD and León J: Myc and
cell cycle control. Biochim Biophys Acta. 1849:506–516. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang X, Neganova I, Przyborski S, Yang C,
Cooke M, Atkinson SP, Anyfantis G, Fenyk S, Keith WN, Hoare SF, et
al: A role for NANOG in G1 to S transition in human embryonic stem
cells through direct binding of CDK6 and CDC25A. J Cell Biol.
184:67–82. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
White J, Stead E, Faast R, Conn S,
Cartwright P and Dalton S: Developmental activation of the Rb-E2F
pathway and establishment of cell cycle-regulated cyclin-dependent
kinase activity during embryonic stem cell differentiation. Mol
Biol Cell. 16:2018–2027. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stead E, White J, Faast R, Conn S,
Goldstone S, Rathjen J, Dhingra U, Rathjen P, Walker D and Dalton
S: Pluripotent cell division cycles are driven by ectopic Cdk2,
cyclin A/E and E2F activities. Oncogene. 21:8320–8333. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Singh AM and Dalton S: The cell cycle and
Myc intersect with mechanisms that regulate pluripotency and
reprogramming. Cell Stem Cell. 5:141–149. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ohtsuka S and Dalton S: Molecular and
biological properties of pluripotent embryonic stem cells. Gene
Ther. 15:74–81. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Becker KA, Ghule PN, Therrien JA, Lian JB,
Stein JL, van Wijnen AJ and Stein GS: Self-renewal of human
embryonic stem cells is supported by a shortened G1 cell cycle
phase. J Cell Physiol. 209:883–893. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
McLenachan S, Menchón C, Raya A, Consiglio
A and Edel MJ: Cyclin A1 is essential for setting the pluripotent
state and reducing tumorigenicity of induced pluripotent stem
cells. Stem Cells Dev. 21:2891–2899. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sela Y, Molotski N, Golan S,
Itskovitz-Eldor J and Soen Y: Human embryonic stem cells exhibit
increased propensity to differentiate during the G1 phase prior to
phosphorylation of retinoblastoma protein. Stem Cells.
30:1097–1108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Calder A, Roth-Albin I, Bhatia S, Pilquil
C, Lee JH, Bhatia M, Levadoux-Martin M, McNicol J, Russell J,
Collins T and Draper JS: Lengthened G1 phase indicates
differentiation status in human embryonic stem cells. Stem Cells
Dev. 22:279–295. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Filipczyk AA, Laslett AL, Mummery C and
Pera MF: Differentiation is coupled to changes in the cell cycle
regulatory apparatus of human embryonic stem cells. Stem Cell Res.
1:45–60. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Coronado D, Godet M, Bourillot PY,
Tapponnier Y, Bernat A, Petit M, Afanassieff M, Markossian S,
Malashicheva A, Iacone R, et al: A short G1 phase is an intrinsic
determinant of naïve embryonic stem cell pluripotency. Stem Cell
Res. 10:118–131. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pauklin S and Vallier L: The cell-cycle
state of stem cells determines cell fate propensity. Cell.
155:135–147. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Delacroix L, Moutier E, Altobelli G,
Legras S, Poch O, Choukrallah MA, Bertin I, Jost B and Davidson I:
Cell-specific interaction of retinoic acid receptors with target
genes in mouse embryonic fibroblasts and embryonic stem cells. Mol
Cell Biol. 30:231–244. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jirmanova L, Afanassieff M, Gobert-Gosse
S, Markossian S and Savatier P: Differential contributions of ERK
and PI3-kinase to the regulation of cyclin D1 expression and to the
control of the G1/S transition in mouse embryonic stem cells.
Oncogene. 21:5515–5528. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jain AK, Allton K, Iacovino M, Mahen E,
Milczarek RJ, Zwaka TP, Kyba M and Barton MC: p53 regulates cell
cycle and microRNAs to promote differentiation of human embryonic
stem cells. PLoS Biol. 10:e10012682012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Giuliano CJ, Kerley-Hamilton JS, Bee T,
Freemantle SJ, Manickaratnam R, Dmitrovsky E and Spinella MJ:
Retinoic acid represses a cassette of candidate pluripotency
chromosome 12p genes during induced loss of human embryonal
carcinoma tumorigenicity. Biochim Biophys Acta. 1731:48–56. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu YY, Tachiki KH and Brent GA: A
targeted thyroid hormone receptor alpha gene dominant-negative
mutation (P398H) selectively impairs gene expression in
differentiated embryonic stem cells. Endocrinology. 143:2664–2672.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Van Oudenhove JJ, Grandy RA, Ghule PN, Del
Rio R, Lian JB, Stein JL, Zaidi SK and Stein GS: Lineage-specific
early differentiation of human embryonic stem cells requires a G2
cell cycle pause. Stem Cells. 34:1765–1775. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gonzales KA and Liang H: Transcriptomic
profiling of human embryonic stem cells upon cell cycle
manipulation during pluripotent state dissolution. Genom Data.
6:118–119. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gonzales KA, Liang H, Lim YS, Chan YS, Yeo
JC, Tan CP, Gao B, Le B, Tan ZY, Low KY, et al: Deterministic
restriction on pluripotent state dissolution by cell-cycle
pathways. Cell. 162:564–579. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Islam MS, Stemig ME, Takahashi Y and Hui
SK: Radiation response of mesenchymal stem cells derived from bone
marrow and human pluripotent stem cells. J Radiat Res. 56:269–277.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rebuzzini P, Pignalosa D, Mazzini G, Di
Liberto R, Coppola A, Terranova N, Magni P, Redi CA, Zuccotti M and
Garagna S: Mouse embryonic stem cells that survive γ-rays exposure
maintain pluripotent differentiation potential and genome
stability. J Cell Physiol. 227:1242–1249. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rebuzzini P, Fassina L, Mulas F, Bellazzi
R, Redi CA, Di Liberto R, Magenes G, Adjaye J, Zuccotti M and
Garagna S: Mouse embryonic stem cells irradiated with γ-rays
differentiate into cardiomyocytes but with altered contractile
properties. Mutat Res. 756:37–45. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fluckiger AC, Marcy G, Marchand M, Négre
D, Cosset FL, Mitalipov S, Wolf D, Savatier P and Dehay C: Cell
cycle features of primate embryonic stem cells. Stem Cells.
24:547–556. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Becker KA, Stein JL, Lian JB, van Wijnen
AJ and Stein GS: Establishment of histone gene regulation and cell
cycle checkpoint control in human embryonic stem cells. J Cell
Physiol. 210:517–526. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pauklin S, Madrigal P, Bertero A and
Vallier L: Initiation of stem cell differentiation involves cell
cycle-dependent regulation of developmental genes by Cyclin D.
Genes Dev. 30:421–433. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhao A, Yang L, Ma K, Sun M, Li L, Huang
J, Li Y, Zhang C, Li H and Fu X: Overexpression of cyclin D1
induces the reprogramming of differentiated epidermal cells into
stem cell-like cells. Cell Cycle. 15:644–653. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Su C: Survivin in survival of
hepatocellular carcinoma. Cancer Lett. 379:184–190. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bai M, Yuan M, Liao H, Chen J, Xie B, Yan
D, Xi X, Xu X, Zhang Z and Feng Y: OCT4 pseudogene 5 upregulates
OCT4 expression to promote proliferation by competing with miR-145
in endometrial carcinoma. Oncol Rep. 33:1745–1752. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Han SM, Han SH, Coh YR, Jang G, Ra J Chan,
Kang SK, Lee HW and Youn HY: Enhanced proliferation and
differentiation of Oct4- and Sox2-overexpressing human adipose
tissue mesenchymal stem cells. Exp Mol Med. 46:e1012014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Card DA, Hebbar PB, Li L, Trotter KW,
Komatsu Y, Mishina Y and Archer TK: Oct4/Sox2-regulated miR-302
targets cyclin D1 in human embryonic stem cells. Mol Cell Biol.
28:6426–6438. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lin SL and Ying SY: Mechanism and method
for generating tumor-free iPS cells using intronic microRNA miR-302
induction. Methods Mol Biol. 936:295–312. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sun LT, Yamaguchi S, Hirano K, Ichisaka T,
Kuroda T and Tada T: Nanog co-regulated by Nodal/Smad2 and Oct4 is
required for pluripotency in developing mouse epiblast. Dev Biol.
392:182–192. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li P, Ma X, Adams IR and Yuan P: A tight
control of Rif1 by Oct4 and Smad3 is critical for mouse embryonic
stem cell stability. Cell Death Dis. 6:e15882015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Neganova I, Vilella F, Atkinson SP, Lloret
M, Passos JF, von Zglinicki T, O'Connor JE, Burks D, Jones R,
Armstrong L and Lako M: An important role for CDK2 in G1 to S
checkpoint activation and DNA damage response in human embryonic
stem cells. Stem Cells. 29:651–659. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Bárta T, Vinarský V, Holubcová Z,
Dolezalová D, Verner J, Pospísilová S, Dvorák P and Hampl A: Human
embryonic stem cells are capable of executing G1/S checkpoint
activation. Stem Cells. 28:1143–1152. 2010.PubMed/NCBI
|
|
53
|
Deshpande AM, Dai YS, Kim Y, Kim J, Kimlin
L, Gao K and Wong DT: Cdk2ap1 is required for epigenetic silencing
of Oct4 during murine embryonic stem cell differentiation. J Biol
Chem. 284:6043–6047. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kallas A, Pook M, Trei A and Maimets T:
Assessment of the potential of CDK2 inhibitor NU6140 to influence
the expression of pluripotency markers NANOG, OCT4, and SOX2 in
2102Ep and H9 cells. Int J Cell Biol. 2014:2806382014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ouyang J, Yu W, Liu J, Zhang N, Florens L,
Chen J, Liu H, Washburn M, Pei D and Xie T: Cyclin-dependent
kinase-mediated Sox2 phosphorylation enhances the ability of Sox2
to establish the pluripotent state. J Biol Chem. 290:22782–22794.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Koo BS, Lee SH, Kim JM, Huang S, Kim SH,
Rho YS, Bae WJ, Kang HJ, Kim YS, Moon JH and Lim YC: Oct4 is a
critical regulator of stemness in head and neck squamous carcinoma
cells. Oncogene. 34:2317–2324. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Conklin JF, Baker J and Sage J: The RB
family is required for the self-renewal and survival of human
embryonic stem cells. Nat Commun. 3:12442012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Tsai SY, Opavsky R, Sharma N, Wu L, Naidu
S, Nolan E, Feria-Arias E, Timmers C, Opavska J, de Bruin A, et al:
Mouse development with a single E2F activator. Nature.
454:1137–1141. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kareta MS, Gorges LL, Hafeez S, Benayoun
BA, Marro S, Zmoos AF, Cecchini MJ, Spacek D, Batista LF, O'Brien
M, et al: Inhibition of pluripotency networks by the Rb tumor
suppressor restricts reprogramming and tumorigenesis. Cell Stem
Cell. 16:39–50. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Vilas JM, Ferreirós A, Carneiro C, Morey
L, Da Silva-Álvarez S, Fernandes T, Abad M, Di Croce L,
García-Caballero T, Serrano M, et al: Transcriptional regulation of
Sox2 by the retinoblastoma family of pocket proteins. Oncotarget.
6:2992–3002. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kelleher FC and O'Sullivan H: FOXM1 in
sarcoma: Role in cell cycle, pluripotency genes and stem cell
pathways. Oncotarget. 7:42792–42804. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wierstra I and Alves J: Transcription
factor FOXM1c is repressed by RB and activated by cyclin D1/Cdk4.
Biol Chem. 387:949–962. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chong JL, Wenzel PL, Sáenz-Robles MT, Nair
V, Ferrey A, Hagan JP, Gomez YM, Sharma N, Chen HZ, Ouseph M, et
al: E2f1-3 switch from activators in progenitor cells to repressors
in differentiating cells. Nature. 462:930–934. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Schoeftner S, Scarola M, Comisso E,
Schneider C and Benetti R: An Oct4-pRb axis, controlled by MiR-335,
integrates stem cell self-renewal and cell cycle control. Stem
Cells. 31:717–728. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Doonan JH and Morris NR: The bimG gene of
Aspergillus nidulans, required for completion of anaphase, encodes
a homolog of mammalian phosphoprotein phosphatase 1. Cell.
57:987–996. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kanai D, Ueda A, Akagi T, Yokota T and
Koide H: Oct3/4 directly regulates expression of E2F3a in mouse
embryonic stem cells. Biochem Biophys Res Commun. 459:374–378.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Suzuki DE, Nakahata AM and Okamoto OK:
Knockdown of E2F2 inhibits tumorigenicity, but preserves stemness
of human embryonic stem cells. Stem Cells Dev. 23:1266–1274. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kalaszczynska I, Geng Y, Iino T, Mizuno S,
Choi Y, Kondratiuk I, Silver DP, Wolgemuth DJ, Akashi K and
Sicinski P: Cyclin A is redundant in fibroblasts but essential in
hematopoietic and embryonic stem cells. Cell. 138:352–365. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Neganova I, Tilgner K, Buskin A,
Paraskevopoulou I, Atkinson SP, Peberdy D, Passos JF and Lako M:
CDK1 plays an important role in the maintenance of pluripotency and
genomic stability in human pluripotent stem cells. Cell Death Dis.
5:e15082014. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Van Hoof D, Muñoz J, Braam SR, Pinkse MW,
Linding R, Heck AJ, Mummery CL and Krijgsveld J: Phosphorylation
dynamics during early differentiation of human embryonic stem
cells. Cell Stem Cell. 5:214–226. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Li L, Wang J, Hou J, Wu Z, Zhuang Y, Lu M,
Zhang Y, Zhou X, Li Z, Xiao W and Zhang W: Cdk1 interplays with
Oct4 to repress differentiation of embryonic stem cells into
trophectoderm. FEBS Lett. 586:4100–4107. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhao R, Deibler RW, Lerou PH, Ballabeni A,
Heffner GC, Cahan P, Unternaehrer JJ, Kirschner MW and Daley GQ: A
nontranscriptional role for Oct4 in the regulation of mitotic
entry. Proc Natl Acad Sci USA. 111:15768–15773. 2014; View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Huskey NE, Guo T, Evason KJ, Momcilovic O,
Pardo D, Creasman KJ, Judson RL, Blelloch R, Oakes SA, Hebrok M and
Goga A: CDK1 inhibition targets the p53-NOXA-MCL1 axis, selectively
kills embryonic stem cells, and prevents teratoma formation. Stem
Cell Reports. 4:374–389. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Lee SH, Oh SY, Do SI, Lee HJ, Kang HJ, Rho
YS, Bae WJ and Lim YC: SOX2 regulates self-renewal and
tumorigenicity of stem-like cells of head and neck squamous cell
carcinoma. Br J Cancer. 111:2122–2130. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Tompkins DH, Besnard V, Lange AW, Keiser
AR, Wert SE, Bruno MD and Whitsett JA: Sox2 activates cell
proliferation and differentiation in the respiratory epithelium. Am
J Respir Cell Mol Biol. 45:101–110. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hou Z, Zhao W, Zhou J, Shen L, Zhan P, Xu
C, Chang C, Bi H, Zou J4, Yao X, et al: A long noncoding RNA Sox2ot
regulates lung cancer cell proliferation and is a prognostic
indicator of poor survival. Int J Biochem Cell Biol. 53:380–388.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Kaufmann LT and Niehrs C: Gadd45a and
Gadd45g regulate neural development and exit from pluripotency in
Xenopus. Mech Dev. 128:401–411. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Barreto G, Schäfer A, Marhold J, Stach D,
Swaminathan SK, Handa V, Döderlein G, Maltry N, Wu W, Lyko F and
Niehrs C: Gadd45a promotes epigenetic gene activation by
repair-mediated DNA demethylation. Nature. 445:671–675. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Schäfer A, Schomacher L, Barreto G,
Döderlein G and Niehrs C: Gemcitabine functions epigenetically by
inhibiting repair mediated DNA demethylation. PLoS One.
5:e140602010. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Awe JP, Crespo AV, Li Y, Kiledjian M and
Byrne JA: BAY11 enhances OCT4 synthetic mRNA expression in adult
human skin cells. Stem Cell Res Ther. 4:152013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Jung M, Peterson H, Chavez L, Kahlem P,
Lehrach H, Vilo J and Adjaye J: A data integration approach to
mapping OCT4 gene regulatory networks operative in embryonic stem
cells and embryonal carcinoma cells. PLoS One. 5:e107092010.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Mushtaq M, Gaza HV and Kashuba EV: Role of
the RB-interacting proteins in stem cell biology. Adv Cancer Res.
131:133–157. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Zheng L, Flesken-Nikitin A, Chen PL and
Lee WH: Deficiency of Retinoblastoma gene in mouse embryonic stem
cells leads to genetic instability. Cancer Res. 62:2498–2502.
2002.PubMed/NCBI
|
|
84
|
Eguchi T, Takaki T, Itadani H and Kotani
H: RB silencing compromises the DNA damage-induced G2/M checkpoint
and causes deregulated expression of the ECT2 oncogene. Oncogene.
26:509–520. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
van Harn T, Foijer F, van Vugt M, Banerjee
R, Yang F, Oostra A, Joenje H and te Riele H: Loss of Rb proteins
causes genomic instability in the absence of mitogenic signaling.
Genes Dev. 24:1377–1388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Karantza V, Maroo A, Fay D and Sedivy JM:
Overproduction of Rb protein after the G1/S boundary causes G2
arrest. Mol Cell Biol. 13:6640–6652. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Sage J and Straight AF: RB's original CIN?
Genes Dev. 24:1329–1333. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Kagey MH, Newman JJ, Bilodeau S, Zhan Y,
Orlando DA, van Berkum NL, Ebmeier CC, Goossens J, Rahl PB, Levine
SS, et al: Mediator and cohesin connect gene expression and
chromatin architecture. Nature. 467:430–435. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Hu G, Kim J, Xu Q, Leng Y, Orkin SH and
Elledge SJ: A genome-wide RNAi screen identifies a new
transcriptional module required for self-renewal. Genes Dev.
23:837–848. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Ding L, Paszkowski-Rogacz M, Nitzsche A,
Slabicki MM, Heninger AK, de Vries I, Kittler R, Junqueira M,
Shevchenko A, Schulz H, et al: A genome-scale RNAi screen for Oct4
modulators defines a role of the Paf1 complex for embryonic stem
cell identity. Cell Stem Cell. 4:403–415. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Fazzio TG and Panning B: Condensin
complexes regulate mitotic progression and interphase chromatin
structure in embryonic stem cells. J Cell Biol. 188:491–503. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Solozobova V, Rolletschek A and Blattner
C: Nuclear accumulation and activation of p53 in embryonic stem
cells after DNA damage. BMC Cell Biol. 10:462009. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Dumitru R, Gama V, Fagan BM, Bower JJ,
Swahari V, Pevny LH and Deshmukh M: Human embryonic stem cells have
constitutively active Bax at the Golgi and are primed to undergo
rapid apoptosis. Mol Cell. 46:573–583. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Song H, Chung SK and Xu Y: Modeling
disease in human ESCs using an efficient BAC-based homologous
recombination system. Cell Stem Cell. 6:80–89. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Maimets T, Neganova I, Armstrong L and
Lako M: Activation of p53 by nutlin leads to rapid differentiation
of human embryonic stem cells. Oncogene. 27:5277–5287. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Hadjal Y, Hadadeh O, Yazidi CE, Barruet E
and Binétruy B: A p38MAPK-p53 cascade regulates mesodermal
differentiation and neurogenesis of embryonic stem cells. Cell
Death Dis. 4:e7372013. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Heo SH, Cha Y and Park KS: Hydroxyurea
induces a hypersensitive apoptotic response in mouse embryonic stem
cells through p38-dependent acetylation of p53. Stem Cells Dev.
23:2435–2442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Lee KH, Li M, Michalowski AM, Zhang X,
Liao H, Chen L, Xu Y, Wu X and Huang J: A genomewide study
identifies the Wnt signaling pathway as a major target of p53 in
murine embryonic stem cells. Proc Natl Acad Sci USA. 107:69–74.
2010; View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Abdelalim EM and Tooyama I: Knockdown of
p53 suppresses Nanog expression in embryonic stem cells. Biochem
Biophys Res Commun. 443:652–657. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Akdemir KC, Jain AK, Allton K, Aronow B,
Xu X, Cooney AJ, Li W and Barton MC: Genome-wide profiling reveals
stimulus-specific functions of p53 during differentiation and DNA
damage of human embryonic stem cells. Nucleic Acids Res.
42:205–223. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Zhen HY, Zhou J, Wu HN, Yao C, Zhang T, Wu
T, Quan CS and Li YL: Lidamycin regulates p53 expression by
repressing Oct4 transcription. Biochem Biophys Res Commun.
447:224–230. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Ng WL, Chen G, Wang M, Wang H, Story M,
Shay JW, Zhang X, Wang J, Amin AR, Hu B, et al: OCT4 as a target of
miR-34a stimulates p63 but inhibits p53 to promote human cell
transformation. Cell Death Dis. 5:e10242014. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Chen T, Du J and Lu G: Cell growth arrest
and apoptosis induced by Oct4 or Nanog knockdown in mouse embryonic
stem cells: A possible role of Trp53. Mol Biol Rep. 39:1855–1861.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Zhang ZN, Chung SK, Xu Z and Xu Y: Oct4
maintains the pluripotency of human embryonic stem cells by
inactivating p53 through Sirt1-mediated deacetylation. Stem Cells.
32:157–165. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Yu KR, Yang SR, Jung JW, Kim H, Ko K, Han
DW, Park SB, Choi SW, Kang SK, Schöler H and Kang KS: CD49f
enhances multipotency and maintains stemness through the direct
regulation of OCT4 and SOX2. Stem Cells. 30:876–887. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Itahana Y, Zhang J, Göke J, Vardy LA, Han
R, Iwamoto K, Cukuroglu E, Robson P, Pouladi MA, Colman A and
Itahana K: Histone modifications and p53 binding poise the p21
promoter for activation in human embryonic stem cells. Sci Rep.
6:281122016. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Suvorova II, Grigorash BB, Chuykin IA,
Pospelova TV and Pospelov VA: G1 checkpoint is compromised in mouse
ESCs due to functional uncoupling of p53-p21Waf1 signaling. Cell
Cycle. 15:52–63. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Filion TM, Qiao M, Ghule PN, Mandeville M,
van Wijnen AJ, Stein JL, Lian JB, Altieri DC and Stein GS: Survival
responses of human embryonic stem cells to DNA damage. J Cell
Physiol. 220:586–592. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Zhu H, Hu S and Baker J: JMJD5 regulates
cell cycle and pluripotency in human embryonic stem cells. Stem
Cells. 32:2098–2110. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Niculescu AB III, Chen X, Smeets M, Hengst
L, Prives C and Reed SI: Effects of p21(Cip1/Waf1) at both the G1/S
and the G2/M cell cycle transitions: pRb is a critical determinant
in blocking DNA replication and in preventing endoreduplication.
Mol Cell Biol. 18:629–643. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Karimian A, Ahmadi Y and Yousefi B:
Multiple functions of p21 in cell cycle, apoptosis and
transcriptional regulation after DNA damage. DNA Repair (Amst).
42:63–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Chan WH, Houng WL, Lin CA, Lee CH, Li PW,
Hsieh JT, Shen JL, Yeh HI and Chang WH: Impact of dihydrolipoic
acid on mouse embryonic stem cells and related regulatory
mechanisms. Environ Toxicol. 28:87–97. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Tahmasebi S, Alain T, Rajasekhar VK, Zhang
JP, Prager-Khoutorsky M, Khoutorsky A, Dogan Y, Gkogkas CG,
Petroulakis E, Sylvestre A, et al: Multifaceted regulation of
somatic cell reprogramming by mRNA translational control. Cell Stem
Cell. 14:606–616. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Zhen HY, He QH, Li Y, Zhou J, Yao C, Liu
YN and Ma LJ: Lidamycin induces neural differentiation of mouse
embryonic carcinoma cells through down-regulation of transcription
factor Oct4. Biochem Biophys Res Commun. 421:44–50. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Tsai CC, Su PF, Huang YF, Yew TL and Hung
SC: Oct4 and Nanog directly regulate Dnmt1 to maintain self-renewal
and undifferentiated state in mesenchymal stem cells. Mol Cell.
47:169–182. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Lee J, Go Y, Kang I, Han YM and Kim J:
Oct-4 controls cell-cycle progression of embryonic stem cells.
Biochem J. 426:171–181. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Abu Dawud R, Schreiber K, Schomburg D and
Adjaye J: Human embryonic stem cells and embryonal carcinoma cells
have overlapping and distinct metabolic signatures. PLoS One.
7:e398962012. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Kondoh H, Lleonart ME, Nakashima Y, Yokode
M, Tanaka M, Bernard D, Gil J and Beach D: A high glycolytic flux
supports the proliferative potential of murine embryonic stem
cells. Antioxid Redox Signal. 9:293–299. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Margineantu DH and Hockenbery DM:
Mitochondrial functions in stem cells. Curr Opin Genet Dev.
38:110–117. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Folmes CD and Terzic A: Energy metabolism
in the acquisition and maintenance of stemness. Semin Cell Dev
Biol. 52:68–75. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Folmes CD, Ma H, Mitalipov S and Terzic A:
Mitochondria in pluripotent stem cells: Stemness regulators and
disease targets. Curr Opin Genet Dev. 38:1–7. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
St John JC: Mitochondrial DNA copy number
and replication in reprogramming and differentiation. Semin Cell
Dev Biol. 52:93–101. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Lees JG, Rathjen J, Sheedy JR, Gardner DK
and Harvey AJ: Distinct profiles of human embryonic stem cell
metabolism and mitochondria identified by oxygen. Reproduction.
150:367–382. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Heiden MG Vander, Cantley LC and Thompson
CB: Understanding the Warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Lunt SY and Heiden MG Vander: Aerobic
glycolysis: Meeting the metabolic requirements of cell
proliferation. Annu Rev Cell Dev Biol. 27:441–464. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Bigarella CL, Liang R and Ghaffari S: Stem
cells and the impact of ROS signaling. Development. 141:4206–4218.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Wanet A, Arnould T, Najimi M and Renard P:
Connecting mitochondria, metabolism, and stem cell fate. Stem Cells
Dev. 24:1957–1971. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Kim H, Jang H, Kim TW, Kang BH, Lee SE,
Jeon YK, Chung DH, Choi J, Shin J, Cho EJ and Youn HD: Core
pluripotency factors directly regulate metabolism in embryonic stem
cell to maintain pluripotency. Stem Cells. 33:2699–2711. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Christensen DR, Calder PC and Houghton FD:
GLUT3 and PKM2 regulate OCT4 expression and support the hypoxic
culture of human embryonic stem cells. Sci Rep. 5:175002015.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Lee J, Kim HK, Han YM and Kim J: Pyruvate
kinase isozyme type M2 (PKM2) interacts and cooperates with Oct-4
in regulating transcription. Int J Biochem Cell Biol. 40:1043–1054.
2008. View Article : Google Scholar : PubMed/NCBI
|