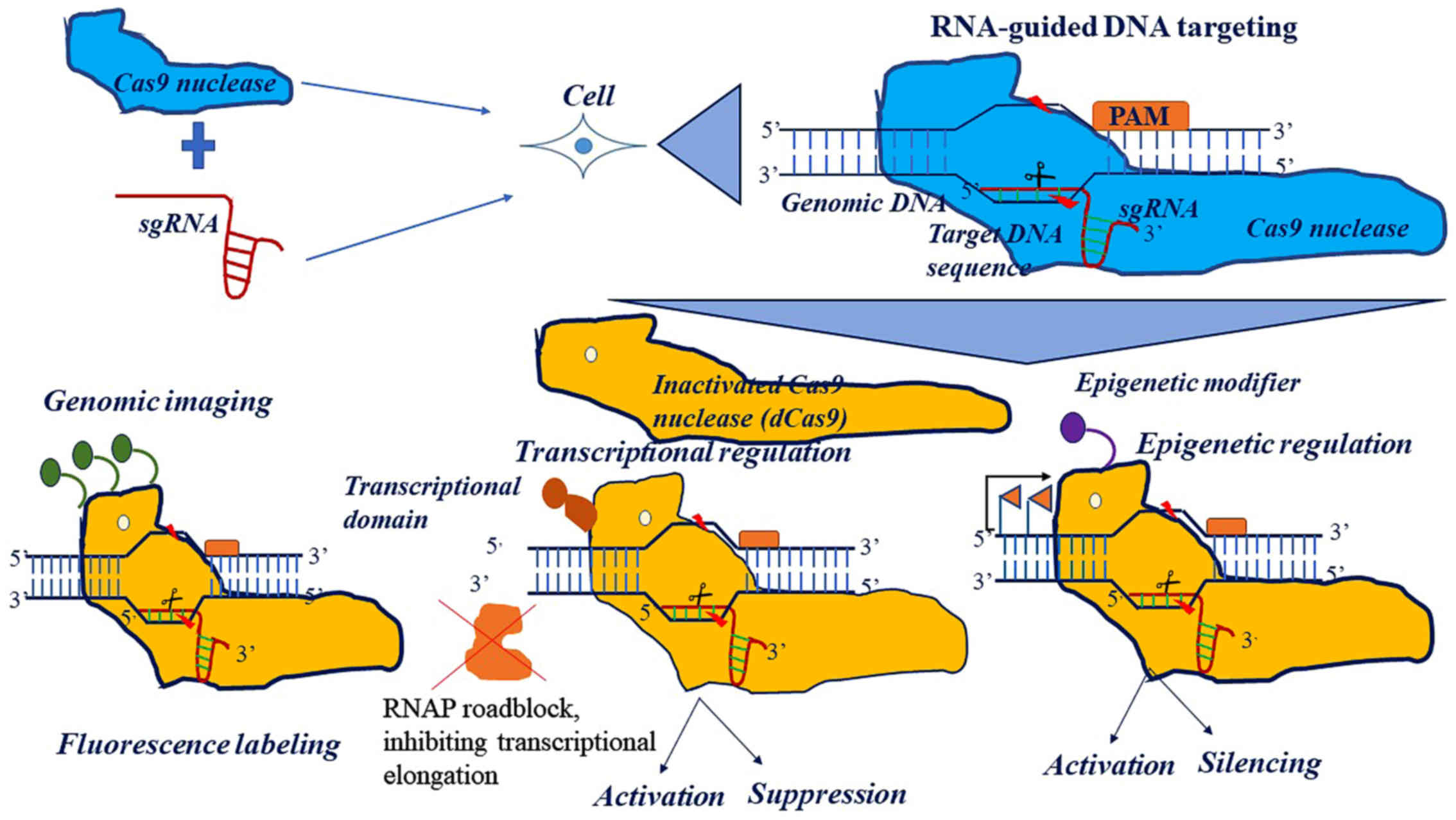
Mechanisms of action of the CRISPR
system
In general, the main function of the CRISPR system
is mediated by the CRISPR-associated protein (Cas) nuclease, which
interacts with DNA and generates double-strand breaks in DNA
sequence, matching the broken genomic region with a single guide
RNA (sgRNA). The sgRNA is a chimeric RNA which consists of
programmable CRISPR RNA and a trans-activating RNA (tracrRNA)
(1). More specifically, the
CRISPR-Cas system includes a cluster of proteins, categorized into
Class 1 (Types I, III and IV) and Class 2 (Types II, V and VI)
(2), all of which constitute
specific RNA-guided DNA endonuclease (Cas) proteins (1–4). A
distinct characteristic of Cas proteins is that they are driven by
RNAs, and not by other proteins, in recognizing the desired DNA
sequence. The CRISPR-Cas subtype that is usually selected is Class
2, which generally exploits Cas9 nuclease (1,3,4). As
regards the RNA-mediated action of the CRISPR system, it has been
reported that the 100 bp sgRNA forms complementary bonds with the
target DNA sequence of 17–20 nucleotides, via Watson-Crick
base-pairing and the tracrRNA is the component which Cas9 nuclease
binds to. Specifically, the sgRNA recognizes the target sequence
located upstream of a triplicate sequence named the protospacer
adjacent motif (PAM), given that the PAM motif recruits Cas9
nuclease at the site of DNA cleavage (5) (Fig.
1). Of note, the PAM sequence plays a determinant role in
recognizing the correct DNA sequence and preventing the direction
of RNA to self-targets and non-specific sequences (6). In the case of Cas9 that is derived
from Streptococcus pyogenes, the motif of the PAM sequence
may be composed of any base, followed by two additional guanine
bases (7). In addition, the
superiority of Cas9 nuclease has been demonstrated not only against
other nucleases, but also against the silencing mechanism, known as
RNA interference (RNAi), that has been used extensively for the
disruption of gene expression. Another important aspect is that the
CRISPR genome editing tool acts in the nucleus, whereas the RNAi
system exerts its action in the cytoplasm. RNAi seems to have
prevalent unpredictable off-target effects (8) due to the extensive modulation of
microRNAs (miRNAs or miRs) (9) or
partial silencing (10). Compared
to RNAi techniques, the CRISPR system functions at the DNA level,
resulting in the permanent inactivation of a given gene, with fewer
off-target effects. On the other hand, the main disadvantage of the
CRISPR system is its low efficiency in some cases, as its effects
are undetectable in a cell population, thus prompting researchers
to optimize the system for implementation in the clinical
setting.
As regards the challenges associated with the CRISPR
technique, the method has been referred to sustain its activity
even if the sgRNA and the target sequence do not form complementary
bonds in 3 sequential bases at the 3′ terminal end of sgRNA
(11). It has also been mentioned
that the scale of side-effects begins at 10 and exacerbates to
1,000. However, the phenomenon can be partially rescued with the
careful design of guide RNA (gRNA) (12); characteristically, the specificity
of the CRISPR system can be accelerated by reducing the sequence of
gRNA to 17–18 nucleotides (13).
Controlling gene expression through the
CRISPR system
Advances in the CRISPR engineering tool have led to
the creation of inactivated (catalytically dead) Cas9 (dCas9).
Modified Cas9 involves the following: Catalytic substitution (D10A)
in the RuvC domain, the abolishment of endonuclease activity and
catalytic replacement (H847A) in the HNH domain.
dCas9 bound to DNA elements alone may inhibit the
transcription either at the initiation or the elongation level.
Specifically, it has been demonstrated that dCas9 can impair
transcription, excluding the potential of crosstalk between
cis-acting motifs of any promoter with trans-acting
transcription factors during transcriptional initiation (14). In the process of transcriptional
elongation, the DNA is not cleaved due to the lack of dCas9
endonuclease activity and the RNA polymerase is sterically
prevented, causing transcriptional stalling in a strand-specific
manner (14). It is evident that
the association between the sgRNA-dCas9 complex and RNAP is
responsible for transcriptional elongation.
Apart from simply blocking the transcription of
target genes, it may be possible to couple the dCas9 protein with
numerous regulatory domains, modulate different biological
processes and generate functional outcomes. Importantly, the
inactivated domain of Cas9 can be combined with activator or
repressor domains, thus controlling gene expression. The
inactivated domain of Cas9 can be fused to epigenetic modifiers,
thus paving the way in the control of gene expression and
epigenetic modifications in a fast and reliable manner (15) (Fig.
1).
In the case of dCas9, the DNA is not cleaved due to
the lack of Cas9 endonuclease activity. The dCas9 can be fused to
transcription factor domains or epigenetic modifiers or fluorescent
molecules, thus determining the control of gene expression or
imaging, respectively.
Overall, the control of gene expression can be
mediated in a spatial-temporal manner, via the binding of dCas9
with a repressor domain, such as Kruppel-associated box (KRAB)
(16) or an activator domain, such
as VP16 and VP64 (17), thereby
inhibiting or triggering gene expression, respectively. On the one
hand, the interruption of transcription in target genes can be
culminated, via a combination of dCas9 with transcriptional
repressor domains (KRAB or SID) and epigenetic modifiers that are
responsible for epigenetic silencing (17,18).
In CRISPR interference (CRISPRi), the transcriptional silencing is
very high at both the prokaryotic and the eukaryotic level
(14). Of note, CRISPRi can
repress the transcription of an endogenous gene or of multiple
genes independently in bacterial and mammalian cells, as
illustrated by Mandegar et al by eliminating the factors,
octamer-binding transcription factor 4 (OCT4), NANOG and sex
determining region Y)-box 2 (SOX2), the kinases, Rho-associated,
coiled-coil-containing protein kinase 1 (ROCK1) and glycogen
synthase kinase 3 (GSK3)-β, a cardiac mesoderm transcription
factor, and cardiac disease-associated genes [BCL2-associated
athanogene 3 (BAG3), myosin binding protein C (MYBPC)3 and human
ether-a-go-go-related gene (hERG)] in induced pluripotent stem
cells (iPSCs) (19). The repressor
activity of CRISPRi has only been validated using quantitative
fluorescence assays and native elongating transcript sequencing
(20). On the other hand, Cas9
nuclease can boost gene transcription via coupling with VP16/VP64
or p65 activation domains. It is noteworthy that the level of
transcriptional upregulation depends on the presence of single
(18,21–23)
or multiple sgRNAs which have a cumulative effect on transcription
(21–23). In order to augment the activation
signal, researchers have used a SunTag array, in which dCas9 is
fused to a protein platform loaded with 24 epitopes, each of which
is recognized by a single chain variable fragment (scFv) of
antibody, which is in turn linked to activator effector domains
like VP64 (24). For example, it
has been reported that CXCR4 expression is more prominently
augmented (50 fold) with the SunTag array, as compared to a 2-fold
increase that is mediated by the action of dCas9-VP64 (25). In the same context, the significant
upregulation of gene expression can be mediated by ‘in trans’
interactions through synergistic activation mediator (SAM). The SAM
approach is reportedly based on bringing activators (e.g.,
p65-HSF1) fused with MS2 bacteriophage coat protein and Cas9 loaded
with VP64 to interact with sgRNAs containing MS2 target sites
(26). Furthermore, a method of
enhancing the activation of gene expression though dCas9-VPR (VP64,
p65 and Rat) has been presented, which specifically exploits the
synergistic power of distinct activating factors ‘in cis’
(27).
Based on the ability of RNA to function as a
scaffold, Zalatan et al (28) were inspired to design a CRISPR
sgRNA sequence which was fused to single RNA hairpin domains via a
linker of 2 bases, using viral sequences that recruit the assembly
of RNA-binding factors. In turn, the RNA-binding factors
orchestrated the landscape for transcriptional activation or
suppression of target genes, via interaction with the appropriate
transcription factors. As a result, the expression of multiple
genes was controlled simultaneously, either by activation or
suppression, whereas at the same time this unique system allowed
for large scale screening of pairwise combinations of genes, many
of which constituted important regulators of tumorigenesis
(28).
In all cases, there are certain factors that
influence the outcome of upregulated or attenuated gene expression.
The orientation of sgRNAs relative to the transcriptional start
site (TSS) of genes and the selection between amino terminal or
carboxyterminal fusion of dCas9 with effector domains may be of
fundamental importance in determining the end transcriptional
result. For example, it has been noted that the well-established
VPR system of transcriptional activation can also be exploited for
the suppression of endogenous genes when gRNAs are orientated
downstream of the TSS (29).
Despite the considerable number of studies on
tumorigenesis, there is a certain paucity of data pertinent to
various regulatory elements, including enhancers. For example, the
functional involvement of enhancers in tumorigenesis has been
validated through their capacity to orchestrate gene expression
profiles (30). Classical research
methods, such as high-throughput reporter assays, have been
employed to address enhancer properties (31); however, inadequate results have
been produced. Nowadays, the revolutionary CRISPR approach provides
us with the opportunity to monitor the distribution of regulatory
elements and determine their functional properties. Impressively,
Xie et al (32) devised a
conceptual framework (Mosaic-seq), which utilizes a CRISPR
barcoding approach in combination with single-cell RNA sequencing
by Drop-seq, to quantitatively evaluate enhancer function and
identify the elements of super enhancers (32). Specifically, K562 cells infected
with dCas9 fused to the KRAB domain, resulting in the repression of
gene expression at potential enhancers; subsequently, the cells
were infected with a labeled sgRNA library against possible
enhancers and assorted following the application of antibiotic
tests for RNA sequencing (Drop-seq). Mosaic-seq has offered insight
into enhancer penetrance, endogenous activity and a deeper
understanding of the hierarchy of the constituents that comprise
super enhancers in a genome-wide, single-cell and unbiased manner
(32).
For the study of promoter sequences, Chiba et
al used the CRISPR system to trigger cancer-related TERT
promoter deletions, in order to delineate the consequences in
neoplastic as well as normal cells, such as human embryonic stem
cells (33). In other words, the
particular genome engineering tool allows researchers to identify
the cause of aberrantly long telomeres in tumorigenesis.
Approaches for spatiotemporal control of
gene expression
The CRISPR system can be further evaluated to
function in a spatial-temporal manner and many scientific advances
have been made in this direction. One method that has garnered
significant attraction is gene manipulation according to induction
by light. For example, the heterodimerization of CRY2 and CIB1
proteins has been observed in response to 450 nm light, allowing
the assembly of VP64 to dCas9 (34). Another method supports that Cas9
nuclease is uncoupled to its gRNA in the absence of light. The
researchers have substantiated that a lysine residue (K866) is
responsible for the activation of Cas9 nuclease, given that
conformational changes are performed upon binding of sgRNA and UV
illumination (35). Another method
has employed the segregation of Cas9 nuclease and its conjunction
with photoinducible partners. The action of the CRISPR system is
performed only after irradiation with blue light in which the
distinct segments of Cas9 nuclease are assembled in a complex
(36). Last but not least, the
inducible effect of the CRISPR system on gene expression can be
mediated by a chemical stimulus (doxycycline, rapamycin and
4-hydroxytamoxifen) (37,38).
Progress made in elucidating the role of
non-coding RNAs using the CRISPR system
Despite the reported advances in generating schemes
for the insertion or elimination of genes, there appears to be a
lack of strategies that can be applied at the RNA level. The CRISPR
approach is not restricted to the modification of the coding
compartment of the cell, but it also applies to the non-coding
compartment, which usually has a significant regulatory
functions.
The advantage of CRISPRi over RNAi is that the
former is an exogenous system that does not intervene with
endogenous processes and facilitates the study of long-noncoding
RNAs (lncRNAs), as well as miRNAs. The CRISPRi system is based on
the action of catalytically dCas9 which does not exert endonuclease
activity (14,17). Importantly, dCas9 abrogates the
transcriptional machinery, thus hindering gene expression by
exploiting the sgRNA sequence only, and maps the presence of cis
regulatory element binding in the lactose regulatory network
(19). For the CRISPRi system to
function properly, sgRNAs should be designed such that the distance
between the TSS and the target should be approximately 300 bp
(39), so as to avoid mismatches
and side-effects.
In modulating gene expression, lncRNAs have been
shown to play a fundamental role in both normal developmental
processes and in abnormal cellular states such as cancer, thus
highlighting the need for further investigation (40). lncRNAs have been reported to be
modified by the CRISPR approach only (41). The CRISPR system acts by either
disrupting the target lncRNA or by inserting RNA-destabilizing
elements or by introducing a poly-adenylation stop signal, thus
leading to gene replacement or promoter disruption (40). Several research groups have
employed CRISPR strategies to disrupt the function of lncRNAs in
mouse models. The ablation of the Malat1 lncRNA constitutes an
indicative example of the above strategy (42). An alternative strategy supports the
replacement of an lncRNA sequence with a marker sequence (such as
LacZ), thus allowing the monitoring of the reporter gene expression
as driven by the promoter of the lncRNA (43). However, that method was relatively
costly and tested on mice that were specifically deficient for 18
lncRNAs (43). Notably, it was
previously demonstrated that the HOXA gene cluster can be regulated
by the lncRNA transcript known as Haunt. Using the CRISPR system,
it was found that Haunt lncRNA functioned as a roadblock, impairing
HOXA gene expression when it was assigned as aberrant (44). Therefore, the CRISPR genome
engineering tool appears to be the first method for the
identifaction of the molecular mechanisms mediated by lncRNAs in
various biological processes.
Nonetheless, the implementation of the CRISPR method
requires specific information regarding the traits of lncRNAs, such
as their effect on the ‘cis’ or ‘trans’ expression of
genes or on the expression of multiple genes. For example, only
approximately 40% of assigned lncRNAs can be subjected to the
CRISPR screen, where gene sequences near the lncRNAs may be
influenced by the action of Cas9 nuclease and the danger of no
specificity has not been circumvented.
As regards the study of miRNAs, Xiao et al
were the first group that used the CRISPR method to eliminate large
genomic regions of 1 Mb in size in zebrafish, thereby suppressing
the action of miRNAs that constitute 1 or 3% of the zebrafish
genome (dre-mir-126a or miRNA cluster Chr. 9) (45). In particular, the CRISPR method was
used to suppress the following: miR-137 in ovarian cancer (46), the UCA1 lncRNA in bladder cancer
(47), as well as the Hotair (HOX
transcript antisense RNA) (48)
and BC200 lncRNAs in breast cancer (49). Furthermore, the inverse association
between miR-24-3p and E3 ubiquitin ligase and the TRIM11 oncogene
was previously investigated by the CRISPR method, through which the
attenuation of colorectal tumor progression was facilitated and a
novel therapeutic target in colorectal cancer was highlighted
(50).
Last but not least, the Molecular Chipper approach
was previously used to investigate the functional regulatory
elements involved in the biogenesis of lncRNAs, highlighting
important cis-domains for miR-142 biogenesis (51). In the same direction, Shechner
et al demonstrated an innovative CRISPR-Display method in
studying the functional significance of regulatory molecules such
as lncRNAs, miRNAs, circular RNAs, piwi-interacting RNAs and many
others types of RNA (52). The
pioneering CRISPR-Display platform was designed to shed further
light on the molecular mechanisms exerted by lncRNAs on a spectrum
of biological processes, such as epigenetic modifications,
transcriptional regulation and chromatin remodeling. The
CRISP-Display platform has allowed researchers to evaluate the
functionality of RNA molecules, either based on their
transcriptional profile or on topological changes and alterations
in the tridimensional landscape (52). A characteristic example of
exploiting accessory RNA domains was demonstrated by Shechner et
al, who conjugated the RepA domain of Xist with gRNA.
Therefore, the repressive action of Xist, caused by its association
with the RepA domain, was unveiled. The CRISPR-Display platform
also functions in a flexible and multiplex manner, as it
facilitates fusing the gRNA to a lncRNA and monitoring of the
transcription of a reported gene with concomitant imaging of
another DNA locus, thus allowing for simultaneous analysis of
several targets based on the available RNA motifs (53).
The main characteristic of the CRISP-Display
platform is the delivery of dCas9 to predetermined DNA loci
according to the specificity of gRNA and the dCas9 fusion with any
of transcriptional effector domains, including VP4 (52). As regards the RNA cargos used by
Cas9, it was indicated that effector domains can be linked to dCas9
nuclease either directly (known as ‘direct activation) or
indirectly, using aptamers which can function as ‘accessory RNA
domains’ of 4.8 kb in size, with the potential of binding the
suitable transcriptional domain (knownas ‘bridged’ activation)
without sterical barriers, given that the RNA domains can be
positioned at the 5′ or the 3′ end of gRNA. The only restraint in
implementing the CRISP-Display approach in all the non-coding RNAs
was the restricted panel of RNA domains that can be appended to
sgRNAs (52).
Exploiting the CRISPR/Cas approach in the
field of epigenetics
The etiology of several diseases may be linked to
epigenetic alterations, which may have a significant impact on the
transcriptional level or on DNA methylation patterns or histone
modifications, thus altering diverse biological functions such as
transcription. It has been demonstrated that mutations in inactive
nucleases cause them to be fused with epigenetic modifiers and the
transcriptional landscape is reorganized. Stable transcriptional
suppression has been achieved through fusion of dCas9 with a
repression domain such as KRAB and DNA methylatransferase (53). Specifically, Cas9 nuclease has been
successfully fused with the Dnmt3 DNA methyltransferases (54) and with the G9a or SUV39H1
methyltransferase to mediate epigenetic silencing. Notably, the
synergistic effect of the dCas9 repressor domain fused to DNA
methyltransferasesDNMT3A and DNMT3L appears to be more effective
than using the dCas9-repressor domain alone (55).
By contrast, the fusion of dCas9 to other epigenetic
modulators, such as TET1 demethylase (56), LSD1 histone demethylase or the p300
histone acetyltransferase (57)
seems to upregulate endogenous genes. Nonetheless, experimental
methods need to be further investigated in order to elucidate the
crosstalk between epigenetic modifiers and endogenous proteins, as
they often cause false phenotypes (58).
Implementation of the CRISPR system in the
field of imaging
The organization of chromatin in the nucleus is
usually addressed as a critical issue. Identifying the principles
behind chromatin orientation can help in the understanding of how
the spatial organization of chromatin can affect gene expression.
For this purpose, FISH technologies that utilize the denaturation
of double-strand DNA, heating/formamide and the attachment of
fluorescent probes to denatured DNA have been extensively used. The
main drawbacks of the available techniques are disruption of the
genomic structure, restricted resolution and the prohibitive
expense of oligo probes (59).
Previous data have demonstrated an impressive
technique to monitor not only the repetitive elements in telomeres,
but also gene sequences in live cells, circumventing the classical
problems of fixation and DNA denaturation that emerge during FISH
technologies or artificial insertions of LacO or TetO cassettes
(60). Specifically, Chen et
al modified a form of nuclease Cas9 [coupled with a fluorescent
molecule, such as enhanced green fluorescent protein (EGFP)] and
optimized small guide RNA, thus managing to monitor the route of
desired genomic sequences in live cells and track the dynamic
nature of chromatin (61). In the
meantime, Cas9 nuclease appeared to be a more powerful as compared
to the transcription activator-like effector nuclease that had been
usually used for detecting telomere and satellite DNA (62), as Cas9 nuclease has the capacity to
label multiple genomic sequences in live cells. Consequently,
imaging using Cas9 nuclease proved to be a significant advancement,
not only in terms of relating chromatin state with gene function,
but also in understanding the aneuploidy that is usually seen in
cancer cells (63). In addition,
CRISPR imaging appears to be very effective in studying
interactions among genomic elements as compared to other techniques
(chromatin conformation capture-3C and derivatives such as Hi-C)
(64). Importantly, Cas9 nuclease
was recently modulated to monitor RNA in live cells instead of RNA
FISH/MeCP2, thus facilitating visualization of the endogenous RNA
distribution in live cells without exogenous tags, and even their
accumulation in stress granules of live cells (65). A particularly innovative technology
was devised for the detection of genes within undisturbed nuclei of
fixed cells and tissues; Cas9 and CRISPR repeats (clustered
regularly interspaced short palindromic repeats) were combined with
an RNA sequence to serve as probes in identifying the genes of
interest at the genome level. That approach not only preserved the
spatial relationships of the genetic elements, which appear very
important in understanding gene expression, but the process was
overall remarkably quick (15 min), convenient and amenable to
diagnostic tests. Previous approaches have used fluorescent DNA
probes to hybridize to genes of interest, which often require heat
treatment and disruptive chemicals that distort the natural
organization of the nucleus.
Furthermore, the combination of inactivated Ca9 and
EGFP has proven to be very sufficient in monitoring the
localization of any genomic sequence, apart from the telomeres in
which it was first applied (61).
In this direction, another advantageous method for enriching the
signal in live cell imaging is to use the dCas9-SunTag system, in
which fluorescent molecules are fused to individual scFv which are
located on a protein epitope tail fused to dCas9 (24). In addition, phototoxicity has been
minimized by reducing excitation laser power and subsequently by
amplifying the signal at a specific target (24). If one considers the scaffold
properties of sgRNA (50), random
RNA aptamers could be exploited for the assembly of fluorescent
molecules (66), instead of using
epitopes.
Utilizing the CRISPR system in monitoring
cell fate
Despite being essential tools in the identification
of tumorigenic genes, pooled genetic screens cannot be used to
analyze neoplasm cellular complexity and clonality, as the results
derived from screens can lead to false-negative and -positive
conclusions (67).
Based on the notion that cancer is a stochastic
procedure in which cancer cells adapt to the environment by
acquiring ‘de novo’ mutations, researchers have revealed a
new barcoding system that enables monitoring the fate of cancer
subpopulations. Notably, the tumor immune escape is attributed to
complex tumor heterogeneity, possibly pertaining to the origin of
cancer cells. In the CRSIPR-barcoding system, a homology directed
repair mechanism has been used to specifically introduce mutations
in cancer cells, along with silent mutations as a reference, thus
recapitulating the clonal history of cells. It has been
demonstrated that CRISPR-barcoding is multiplex, allowing the
remodeling of many mutations in cancer cell populations and leading
to a more comprehensive understanding of cellular networks. The
effects of distinct drug combinations (gefitinib-WZ4002, ALK
inhibitor TAE684-WZ4002, gefitinib-trametinib) in prostate cancer
cells (PC9 cells) harboring either KRAS G12D mutation or EGFR T790M
mutation or EML4-ALK rearrangement have been deciphered. In
addition, the effects caused by inactivation of the p53 gene (TP53
R273H) in breast cancer cells (MCF7 cells) and colon cancer cells
(HCT-116 cells) have been identified, whereas the expression of
adenomatous polyposis coli gene (APC) in colon cancer cells (DLD-1
cells) has been abrogated. Furthermore, CRISPR-barcoding has been
employed to repair the ALK-F1174L activating mutation in Kelly
neuroblastoma cells, resulting in correction of the mutation or in
generating truncated protein (ALK-STOP barcode). Overall, this new
CRISPR-barcoding tool has facilitated the monitoring and study of
cell fate, both in response to therapy and in the absence of
therapy. The new strategy may be used for the identification of any
type of genetic modification, is compatible with many cancer types
and functional assays, but above all it does not cause side effects
owing to silent mutations, thus paving the way for the development
of new therapeutic protocols (68).
Conclusions
The CRISPR system has been employed for the
enhancement or attenuation of gene expression in a more reliable
manner as compared to previous genome engineering techniques. The
innovative use of the CRISPR system has been implemented to
modulate gene expression through the alteration of epigenetic
modifiers or crucial noncoding RNAs. Importantly, the CRISPR system
is of utmost importance in delineating the role of non-coding RNAs
(miRNAs, lncRNAs) which are often implicated in tumor progression
and heterogeneity (Fig. 2).
Finally, the CRISPR system constitutes a flexible platform that has
been used for monitoring the localization of endogenous genes or
cell fate in general.
References
|
1
|
Wright AV, Nuñez JK and Doudna JA: Biology
and applications of CRISPR systems: Harnessing nature's toolbox for
genome engineering. Cell. 164:29–44. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wiedenheft B, Sternberg SH and Doudna JA:
RNA-guided genetic silencing systems in bacteria and archaea.
Nature. 482:331–338. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
van der Oost J, Westra ER, Jackson RN and
Wiedenheft B: Unravelling the structural and mechanistic basis of
CRISPR-cas systems. Nat Rev Microbiol. 12:479–492. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mougiakos I, Bosma EF, de Vos WM, van
Kranenburg R and van der Oost J: Next generation prokaryotic
engineering: The CRISPR-cas toolkit. Trends Biotechnol. 34:575–587.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bolotin A, Quinquis B, Sorokin A and
Ehrlich SD: Clustered regularly interspaced short palindrome
repeats (CRISPRs) have spacers of extrachromosomal origin.
Microbiology. 151:2551–2561. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Westra ER, Semenova E, Datsenko KA,
Jackson RN, Wiedenheft B, Severinov K and Brouns SJ: Type I-E
CRISPR-cas systems discriminate target from non-target DNA through
base pairing-independent PAM recognition. PLoS Genet.
9:e10037422013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mojica FJ, Díez-Villaseñor C,
García-Martínez J and Almendros C: Short motif sequences determine
the targets of the prokaryotic CRISPR defence system. Microbiology.
155:733–740. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jackson AL, Bartz SR, Schelter J,
Kobayashi SV, Burchard J, Mao M, Li B, Cavet G and Linsley PS:
Expression profiling reveals off-target gene regulation by RNAi.
Nat Biotechnol. 21:635–637. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Marine S, Bahl A, Ferrer M and Buehler E:
Common seed analysis to identify off-target effects in siRNA
screens. J Biomol Screen. 17:370–378. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Moore JD: The impact of CRISPR-Cas9 on
target identification and validation. Drug Discov Today.
20:450–457. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ran FA, Hsu PD, Wright J, Agarwala V,
Scott DA and Zhang F: Genome engineering using the CRISPR-Cas9
system. Nat Protoc. 8:2281–2308. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kuscu C, Arslan S, Singh R, Thorpe J and
Adli M: Genome-wide analysis reveals characteristics of off-target
sites bound by the Cas9 endonuclease. Nat Biotechnol. 32:677–683.
2014. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fu Y, Sander JD, Reyon D, Cascio VM and
Joung JK: Improving CRISPR-cas nuclease specificity using truncated
guide RNAs. Nat Biotechnol. 32:279–284. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qi LS, Larson MH, Gilbert LA, Doudna JA,
Weissman JS, Arkin AP and Lim WA: Repurposing CRISPR as an
RNA-guided platform for sequence-specific control of gene
expression. Cell. 152:1173–1183. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Boettcher M and McManus MT: Choosing the
right tool for the job: RNAi, TALEN, or CRISPR. Mol Cell.
58:575–585. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lawhorn IE, Ferreira JP and Wang CL:
Evaluation of sgRNA target sites for CRISPR-mediated repression of
TP53. PLoS One. 9:e1132322014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gilbert LA, Larson MH, Morsut L, Liu Z,
Brar GA, Torres SE, Stern-Ginossar N, Brandman O, Whitehead EH,
Doudna JA, et al: CRISPR-mediated modular RNA-guided regulation of
transcription in eukaryotes. Cell. 154:442–451. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Konermann S, Brigham MD, Trevino A, Hsu
PD, Heidenreich M, Cong L, Platt RJ, Scott DA, Church GM and Zhang
F: Optical control of mammalian endogenous transcription and
epigenetic states. Nature. 500:472–476. 2013.PubMed/NCBI
|
|
19
|
Mandegar MA, Huebsch N, Frolov EB, Shin E,
Truong A, Olvera MP, Chan AH, Miyaoka Y, Holmes K, Spencer CI, et
al: CRISPR interference efficiently induces specific and reversible
gene silencing in human iPSCs. Cell Stem Cell. 18:541–553. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Larson MH, Gilbert LA, Wang X, Lim WA,
Weissman JS and Qi LS: CRISPR interference (CRISPRi) for
sequence-specific control of gene expression. Nat Protoc.
8:2180–2196. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Maeder ML, Linder SJ, Cascio VM, Fu Y, Ho
QH and Joung JK: CRISPR RNA-guided activation of endogenous human
genes. Nat Methods. 10:977–979. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mali P, Aach J, Stranges PB, Esvelt KM,
Moosburner M, Kosuri S, Yang L and Church GM: CAS9 transcriptional
activators for target specificity screening and paired nickases for
cooperative genome engineering. Nat Biotechnol. 31:833–838. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Perez-Pinera P, Kocak DD, Vockley CM,
Adler AF, Kabadi AM, Polstein LR, Thakore PI, Glass KA, Ousterout
DG, Leong KW, et al: RNA-guided gene activation by
CRISPR-Cas9-based transcription factors. Nat Methods. 10:973–976.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tanenbaum ME, Gilbert LA, Qi LS, Weissman
JS and Vale RD: A protein-tagging system for signal amplification
in gene expression and fluorescence imaging. Cell. 159:635–646.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
La Russa MF and Qi LS: The new state of
the art: Cas9 for gene activation and repression. Mol Cell Biol.
35:3800–3809. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Konermann S, Brigham MD, Trevino AE, Joung
J, Abudayyeh OO, Barcena C, Hsu PD, Habib N, Gootenberg JS,
Nishimasu H, et al: Genome-scale transcriptional activation by an
engineered CRISPR-Cas9 complex. Nature. 517:583–588. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chavez A, Scheiman J, Vora S, Pruitt BW,
Tuttle MPR, Iyer E, Lin S, Kiani S, Guzman CD, Wiegand DJ, et al:
Highly efficient Cas9-mediated transcriptional programming. Nat
Methods. 12:326–328. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zalatan JG, Lee ME, Almeida R, Gilbert LA,
Whitehead EH, La Russa M, Tsai JC, Weissman JS, Dueber JE, Qi LS
and Lim WA: Engineering complex synthetic transcriptional programs
with CRISPR RNA scaffolds. Cell. 160:339–350. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Braun CJ, Bruno PM, Horlbeck MA, Gilbert
LA, Weissman JS and Hemann MT: Versatile in vivo regulation of
tumor phenotypes by dCas9-mediated transcriptional perturbation.
Proc Natl Acad Sci USA. 113:E3892–E3900. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mansour MR, Abraham BJ, Anders L,
Berezovskaya A, Gutierrez A, Durbin AD, Etchin J, Lawton L, Sallan
SE, Silverman LB, et al: Oncogene regulation. An oncogenic
super-enhancer formed through somatic mutation of a noncoding
intergenic element. Science. 346:1373–1377. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Inoue F, Kircher M, Martin B, Cooper GM,
Witten DM, McManus MT, Ahituv N and Shendure J: A systematic
comparison reveals substantial differences in chromosomal versus
episomal encoding of enhancer activity. Genome Res. 27:38–52. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xie S, Duan J, Li B, Zhou P and Hon GC:
Multiplexed engineering and analysis of combinatorial enhancer
activity in single cells. Molecular cell. 66:285–99. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chiba K, Johnson JZ, Vogan JM, Wagner T,
Boyle JM and Hockemeyer D: Cancer-associated TERT promoter
mutations abrogate telomerase silencing. eLife. 4:42015. View Article : Google Scholar
|
|
34
|
Polstein LR and Gersbach CA: A
light-inducible CRISPR-Cas9 system for control of endogenous gene
activation. Nat Chem Biol. 11:198–200. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hemphill J, Borchardt EK, Brown K, Asokan
A and Deiters A: Optical Control of CRISPR/Cas9 gene editing. J Am
Chem Soc. 137:5642–5645. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nihongaki Y, Kawano F, Nakajima T and Sato
M: Photoactivatable CRISPR-Cas9 for optogenetic genome editing. Nat
Biotechnol. 33:755–760. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu KI, Ramli MN, Woo CW, Wang Y, Zhao T,
Zhang X, Yim GR, Chong BY, Gowher A, Chua MZ, et al: A
chemical-inducible CRISPR-Cas9 system for rapid control of genome
editing. Nat Chem Biol. 12:980–987. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zetsche B, Volz SE and Zhang F: A
split-Cas9 architecture for inducible genome editing and
transcription modulation. Nat Biotechnol. 33:139–142. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gilbert LA, Horlbeck MA, Adamson B,
Villalta JE, Chen Y, Whitehead EH, Guimaraes C, Panning B, Ploegh
HL, Bassik MC, et al: Genome-scale CRISPR-mediated control of gene
repression and activation. Cell. 159:647–661. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Haemmerle M and Gutschner T: Long
non-coding RNAs in cancer and development: Where do we go from
here? Int J Mol Sci. 16:1395–1405. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hale CR, Zhao P, Olson S, Duff MO,
Graveley BR, Wells L, Terns RM and Terns MP: RNA-guided RNA
cleavage by a CRISPR RNA-Cas protein complex. Cell. 139:945–956.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gutschner T: Silencing long noncoding RNAs
with genome-editing tools. Methods Mol Biol. 1239:241–250. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sauvageau M, Goff LA, Lodato S, Bonev B,
Groff AF, Gerhardinger C, Sanchez-Gomez DB, Hacisuleyman E, Li E,
Spence M, et al: Multiple knockout mouse models reveal lincRNAs are
required for life and brain development. eLife. 2:e017492013.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yin Y, Yan P, Lu J, Song G, Zhu Y, Li Z,
Zhao Y, Shen B, Huang X, Zhu H, et al: Opposing roles for the
lncRNA haunt and its genomic locus in regulating HOXA gene
activation during embryonic stem cell differentiation. Cell Stem
Cell. 16:504–516. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xiao A, Wang Z, Hu Y, Wu Y, Luo Z, Yang Z,
Zu Y, Li W, Huang P, Tong X, et al: Chromosomal deletions and
inversions mediated by TALENs and CRISPR/Cas in zebrafish. Nucleic
Acids Res. 41:e1412013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li X, Chen W, Zeng W, Wan C, Duan S and
Jiang S: microRNA-137 promotes apoptosis in ovarian cancer cells
via the regulation of XIAP. Br J Cancer. 116:66–76. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhen S, Hua L, Liu YH, Sun XM, Jiang MM,
Chen W, Zhao L and Li X: Inhibition of long non-coding RNA UCA1 by
CRISPR/Cas9 attenuated malignant phenotypes of bladder cancer.
Oncotarget. 8:9634–9646. 2017.PubMed/NCBI
|
|
48
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Singh R, Gupta SC, Peng WX, Zhou N,
Pochampally R, Atfi A, Watabe K, Lu Z and Mo YY: Regulation of
alternative splicing of Bcl-x by BC200 contributes to breast cancer
pathogenesis. Cell Death Dis. 7:e22622016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yin Y, Zhong J, Li SW, Li JZ, Zhou M, Chen
Y, Sang Y and Liu L: TRIM11, a direct target of miR-24-3p, promotes
cell proliferation and inhibits apoptosis in colon cancer.
Oncotarget. 7:86755–86765. 2016.PubMed/NCBI
|
|
51
|
Cheng J, Roden CA, Pan W, Zhu S, Baccei A,
Pan X, Jiang T, Kluger Y, Weissman SM, Guo S, et al: A molecular
chipper technology for CRISPR sgRNA library generation and
functional mapping of noncoding regions. Nat Commun. 7:111782016.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Shechner DM, Hacisuleyman E, Younger ST
and Rinn JL: Multiplexable, locus-specific targeting of long RNAs
with CRISPR-Display. Nat Methods. 12:664–670. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Plummer RJ, Guo Y and Peng Y: A CRISPR
reimagining: New twists and turns of CRISPR beyond the
genome-engineering revolution. J Cell Biochem. 2017, https://doi.org/10.1002/jcb.26406 View Article : Google Scholar
|
|
54
|
Vojta A, Dobrinić P, Tadić V, Bočkor L,
Korać P, Julg B, Klasić M and Zoldoš V: Repurposing the CRISPR-Cas9
system for targeted DNA methylation. Nucleic Acids Res.
44:5615–5628. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Amabile A, Migliara A, Capasso P, Biffi M,
Cittaro D, Naldini L and Lombardo A: Inheritable silencing of
endogenous genes by hit-and-run targeted epigenetic editing. Cell.
167:219–232. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liu XS, Wu H, Ji X, Stelzer Y, Wu X,
Czauderna S, Shu J, Dadon D, Young RA and Jaenisch R: Editing DNA
methylation in the mammalian genome. Cell. 167:233–247. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Hilton IB, D'Ippolito AM, Vockley CM,
Thakore PI, Crawford GE, Reddy TE and Gersbach CA: Epigenome
editing by a CRISPR-Cas9-based acetyltransferase activates genes
from promoters and enhancers. Nat Biotechnol. 33:510–517. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Pineda M, Moghadam F, Ebrahimkhani MR and
Kiani S: Engineered CRISPR systems for next generation gene
therapies. ACS Synth Biol. 6:1614–1626. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Beliveau BJ, Joyce EF, Apostolopoulos N,
Yilmaz F, Fonseka CY, McCole RB, Chang Y, Li JB, Senaratne TN,
Williams BR, et al: Versatile design and synthesis platform for
visualizing genomes with oligopaint FISH probes. Proc Natl Acad Sci
USA. 109:21301–21306. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Heun P, Laroche T, Shimada K, Furrer P and
Gasser SM: Chromosome dynamics in the yeast interphase nucleus.
Science. 294:2181–2186. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chen B, Gilbert LA, Cimini BA,
Schnitzbauer J, Zhang W, Li GW, Park J, Blackburn EH, Weissman JS,
Qi LS and Huang B: Dynamic imaging of genomic loci in living human
cells by an optimized CRISPR/Cas system. Cell. 155:1479–1491. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Miyanari Y, Ziegler-Birling C and
Torres-Padilla ME: Live visualization of chromatin dynamics with
fluorescent TALEs. Nat Struct Mol Biol. 20:1321–1324. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Roukos V, Voss TC, Schmidt CK, Lee S,
Wangsa D and Misteli T: Spatial dynamics of chromosome
translocations in living cells. Science. 341:660–664. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
van Steensel B and Dekker J: Genomics
tools for unraveling chromosome architecture. Nat Biotechnol.
28:1089–1095. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Nelles DA, Fang MY, O'Connell MR, Xu JL,
Markmiller SJ, Doudna JA and Yeo GW: Programmable RNA tracking in
live cells with CRISPR/Cas9. Cell. 165:488–496. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang S, Su JH, Zhang F and Zhuang X: An
RNA-aptamer-based two-color CRISPR labeling system. Sci Rep.
6:268572016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Jaitin DA, Weiner A, Yofe I, Lara-Astiaso
D, Keren-Shaul H, David E, Salame TM, Tanay A, van Oudenaarden A
and Amit I: Dissecting immune circuits by linking CRISPR-pooled
screens with single-Cell RNA-Seq. Cell. 167:1883–1896. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Guernet A, Mungamuri SK, Cartier D,
Sachidanandam R, Jayaprakash A, Adriouch S, Vezain M, Charbonnier
F, Rohkin G, Coutant S, et al: CRISPR-barcoding for intratumor
genetic heterogeneity modeling and functional analysis of oncogenic
driver mutations. Mol Cell. 63:526–538. 2016. View Article : Google Scholar : PubMed/NCBI
|