Introduction
Hepatoblastoma (HB) is a common type of liver
malignancy in young children, which is typically diagnosed in those
<5 years (1). HB accounts for
>90% of malignant liver tumors in childhood (2). The incidence of HB is >1.2 million
children every year globally, with high incidences in Japan, China
and USA (3). The etiology of HB
remains unclear, and low birth weight, preterm birth and maternal
gestational age (<20 years and >35 years) may be associated
with the morbidity of HB (1,4). HB
is hypothesized to originate from primitive hepatic stem cells and
it is not associated with hepatic virus infection, cirrhosis or
other liver disease, which is different from malignant liver tumors
in adults. The primary treatment strategies for HB are surgical
resection and chemotherapy. However, the prognosis of HB remains
poor, as HB is a tumor of hypervascularity, the majority of
patients are asymptomatic and many cases are diagnosed at a late
stage. In addition, the treatment of HB continues to be challenging
due to drug resistance and the occurrence of metastasis, such as
lung, brain and lymph gland metastasis (5,6).
Thus, there is an urgent requirement to investigate the
pathogenesis of HB and develop novel therapeutic targets for the
treatment of patients with HB.
Malignant solid tumors must acquire sufficient blood
supply to promote their survival, proliferation and metastasis.
There are three dominant tumor microcirculation patterns, including
angiogenesis, mosaic vessels and vasculogenic mimicry (VM). VM was
first reported in melanoma by Maniotis et al (7) in 1999, which described the specific
capacity of tumor cells to form extracellular matrix (ECM)-rich
networks. It is distinct from angiogenesis and predominantly occurs
in the early stage of tumorigenesis (8). VM is lined by tumor cells, which may
provide the tumor with oxygen and nutrients, and has been observed
in different types of cancer, and is associated with high tumor
grade, invasion, metastasis and short survival (9–18).
Li et al (12) reported
that patients with VM were at a higher risk for hematogenous
metastasis and distant recurrence when compared with patients
without VM (12). Antiangiogenic
drugs have been demonstrated to be unsatisfactory in the treatment
of malignant tumors during clinical trials and animal testing over
the past decades (19–21). This may be attributed to the
particular structure of the VM, which is lined by tumor cells
rather than endothelial cells, meanwhile the tumor cells engaged in
VM are exposed directly to the blood flow and thus more easily
spread throughout the circulation. Therefore, VM may be an
important target for novel cancer therapeutic strategies to treat
tumors.
Arsenic trioxide (As2O3) has
been used to successfully treat acute promyelocytic leukemia
(22,23) and other types of cancer cell lines
(24–28). As2O3 induces
cell apoptosis, and reduces the invasive and metastatic activities
in cancer cells (24,25,29),
which involve the caspase-3 signaling pathway, reactive oxygen
species, DNA damage, oxidative stress and cell cycle arrest.
However, As2O3, as a vascular disrupting
agent, delayed tumor growth in colon cancer and gastric cancer
cells at a dose of >2.5 mg/kg in a mouse model (30,31).
However, the effectiveness of As2O3 is based
upon a high concentration or long exposure and cell toxicity, which
leads to undesirable side effects and limits the application of
this drug. A previous study reported that
As2O3 exerted its antitumor activity via
depletion of regulatory T cells in a murine model of colon cancer
at a low concentration without side effects (32). To the best of our knowledge,
whether the rapid progression of HB is associated with VM formation
and whether a low dosage of As2O3 exerts
inhibitory effects on VM formation in HB have not been reported.
The aim of the present study was to investigate the impact of a low
dosage of As2O3 on VM formation in HB and the
relative mechanisms in order to develop novel therapeutic
strategies for HB.
Materials and methods
Chemicals
The As2O3 solution was
purchased from Heilongjiang Harbin Medical Pharmaceutical Co., Ltd.
(Heilongjiang, China). Matrigel was purchased from BD Biosciences
(Franklin Lakes, NJ, USA), the CD105-antibody was purchased from
Thermo Fisher Scientific, Inc. (Waltham, MA, USA), and
anti-vascular endothelial (VE)-cadherin, anti-Bcl-2-associated X
protein (Bax) and anti-B-cell lymphoma 2 (Bcl-2) antibodies were
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Matrix metalloproteinase (MMP)-2 and MMP-9 antibodies were
purchased from Nanjing KeyGen Biotech Co., Ltd. (Nanjing, China),
and the animals were obtained from Hunan Slack Jingda Experimental
Animals Co., Ltd. (Hunan, China).
Cell culture and animal treatment
The HB-derived cell line HepG2 cells were
donated by the Department of General Surgery of the First
Affiliated Hospital of Harbin Medical University (Harbin, China).
Cells were cultured in high-glucose Dulbecco's modified Eagle's
medium (DMEM) supplemented with 10% heat-inactivated fetal bovine
serum (both from Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin. All cells were maintained at 37°C in a
humidified incubator with 5.0% carbon dioxide.
The present study was approved by the Ethics
Committee of The First Affiliated Hospital of Harbin Medical
University (Heilongjiang, China). A total of 10 healthy male nude
mice (age, 4–6 weeks; weight, 18–22 g) were used in the present
study. The animals were maintained under specific pathogen-free
conditions using a laminar airflow rack. The animals had access to
sterilized food and autoclaved water ad libitum, and
experiments commenced after 1 week of acclimatization. Mice were
injected subcutaneously in the right flank with 1×106/
ml HepG2 cells suspended in 200 µl serum-free DMEM, and
were randomly allocated into two groups following the injection of
tumor cells. Then, 2 ml As2O3 (2 mg/kg) or an
equivalent volume of phosphate-buffered saline (PBS) was
administered intraperitoneally every 2 days for a total of 20 days.
Tumor size was measured using Vernier calipers every 2 days, and
tumor volume (TV) was calculated using the formula, TV
(mm3) = (LxW2)/2, where L and W were the
longest and shortest tumor dimensions, respectively. Growth curves
of TV were drawn for the two groups. The mice were carefully
monitored for symptoms of toxicity and were weighed twice a week.
The mean mouse body weight of each group was calculated and used as
a parameter of toxicity, as previously described (33). The mice were anesthetized before
the tumors were removed, and the tumors were weighed and fixed with
10% formaldehyde for the pathological tissue sections.
Methylthiazol tetrazolium (MTT) cell
viability assay
MTT assay was conducted to assess the effect of
As2O3 on the viability of HepG2
cells. Cells were seeded in 96-well plates at a density of 5,000
cells/well, experiments were performed as follows: Varying
concentrations of As2O3 for 24 h and 2 µM
As2O3 for varying durations. A culture medium
without As2O3 served as the control.
Following treatment, the medium was aspirated and the cells were
rinsed with PBS, 15 ml MTT solution (5 mg/ml in PBS) was added to
each well. After a 4-h incubation, 100 µl dimethyl sulfoxide was
added to dissolve the formazan crystal and incubated for another 10
min at room temperature, the absorbance value was measured at 490
nm using an Epoch microplate spectrophotometer (BioTek Instruments,
Inc., Winooski, VT, USA).
Matrigel tube formation assay
Matrigel tube formation assay was performed to
determine the VM capacity of HepG2 cells and the effect
of As2O3 on VM channels. Each well of a
96-well plate was coated with 80 µl Matrigel and allowed to
solidify at 37°C for 30 min. Cells were seeded into the coated
wells and incubated in a humidified incubator at 37°C with 5%
CO2 for 24 h. The effect of As2O3
on tube formation was evaluated using varying concentrations of
As2O3 for 24 h and 2 µM
As2O3 for varying durations. Cells in the
control group were cultured in complete medium without
As2O3 for 24 h. Finally, cells were imaged
using an inverted microscope (Olympus, Tokyo, Japan; magnification,
×200) and tube numbers were counted.
Western blot analysis
Following treatment with
As2O3, cells were harvested and lysed using
RIPA buffer (Beijing Solarbio Science and Technology Co., Ltd.,
Beijing, China) containing 1% phenylmethane sulfonyl fluoride
protease inhibitors. Equivalent quantities of proteins were
separated by SDS-PAGE using a 4.5% stacking gel and an 8% running
gel, and the resolved proteins were transferred to polyvinylidene
fluoride membranes using the semi-dry transfer system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The membranes were blocked
with 5% skimmed milk for 2 h and incubated with primary antibodies
overnight at 4°C. The primary antibodies included: MMP-2 (cat. no.
KG22546, 1:500) and MMP-9 (cat. no. KG22551, 1:500) (both from
Nanjing KeyGen Biotech Co., Ltd., Nanjing, China), VE-cadherin
(cat. no. sc-9989, 1:500), Bax (cat. no. sc-6236, 1:500), Bcl-2
(cat. no. sc-783, 1:500) (all from Santa Cruz Biotechnology, Inc.)
and GAPDH (cat. no. KC-5G4, 1:1,000; Shanghai Kangcheng
Bioengineering Co., Ltd., Shanghai, China). The membranes were then
incubated with horseradish peroxidase-conjugated secondary
antibodies (cat. no. ZB-2301, 1:3,000; OriGene Technologies, Inc.,
Beijing, China) for 1 h at room temperature and the protein bands
on the membranes visualized using an ECL detection system (GE
Healthcare Life Sciences, Little Chalfont, UK).
Immunohistochemical (IHC) and
histochemical double-staining
Tissue sections (4 µm) were sliced from the
paraffin-embedded tumor samples, and were stained with hematoxylin
and eosin according to routine protocols. All slides were dewaxed
in xylenes, rinsed in graded ethanol solutions and finally
rehydrated in double-distilled water. Endogenous peroxidase
activity was blocked with 3% hydrogen peroxide in methanol at an
ambient temperature for 20 min. Antigen retrieval was performed by
heating the slides in 0.01 M sodium citrate buffer for 3 min. After
blocking with 3% normal goat serum for 30 min to reduce
non-specific binding, the slides were incubated in a humidified
chamber overnight at 4°C with the following primary antibodies:
CD105 (cat. no. MS-1290, 1:150; Thermo Fisher Scientific, Inc.),
MMP-2 (cat. no. KG22546, 1:150), MMP-9 (cat. no. KG22551, 1:150)
(both from Nanjing KeyGen Biotech Co., Ltd.), VE-cadherin (cat. no.
sc-9989, 1:150; Santa Cruz Biotechnology, Inc.), then incubated
with secondary antibodies (cat. no. PV-6000; OriGene Technologies,
Inc.) for 20 min at room temperature. The sections were stained
with freshly dispensed 3,3′-diaminobenzidine solution for
observation under a microscope followed by rinsing three times in
PBS. Following staining with anti-CD105 and visualization under an
inverted microscope, sections were re-dyed with Periodic
acid-Schiff (Beijing Reagan Biological Technology Co., Ltd.,
Beijing, China) at 37°C for 15 min in the dark according to the
instructions. CD105-positive vessels indicated blood vessels in
tissues and PAS+/CD105− channels were defined
as VM. For VE-cadherin, MMP-2 and MMP-9 staining, the percentage
and intensity of positive cells were measured as previously
described (34). More than 5
microscopic fields in one section were observed under ×200
magnification.
Statistical analysis
Statistical analysis was performed using Graph Pad
Prism 5 (GraphPad Inc., San Diego, CA, USA). The differences were
analyzed using a non-parametric Mann-Whitney t-test. All
experiments were performed in triplicate, data were expressed as
means ± standard deviation. P<0.05 was considered to indicate a
statistically significant difference.
Results
HepG2 cells form typical
tubular structures in vitro
HepG2 cells were plated on Matrigel in
96-well plates and 1 h later changes in cell morphology were
observed, including shape changes, elongation and the formation of
needle-like structures (Fig. 1A).
After 12 h, typical tubular networks emerged that represented VM
(Fig. 1B).
As2O3
significantly inhibits the tube formation of HepG2 cells
prior to cell viability decrease in vitro
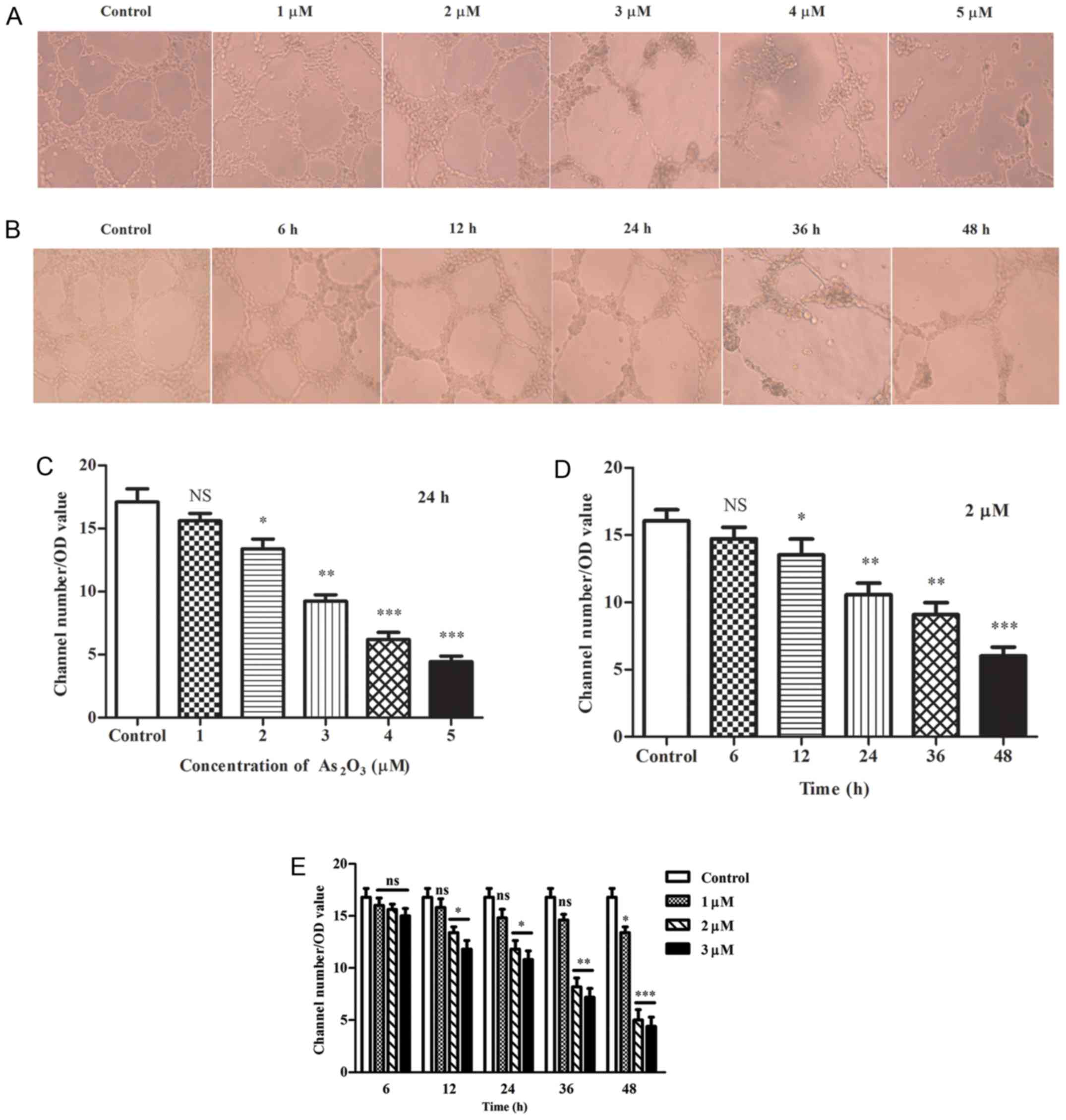
In Matrigel tube formation assay, the mean value of
tube-like structure numbers in 10 fields under an inverted
microscope was obtained, and divided by the optical density (OD)
value. The results indicated that As2O3
induced depolymerization of tube-like structures in a concentration
and time-dependent manner. Significant decreases were observed at
2, 3, 4 and 5 µM, respectively (P<0.05; Fig. 2A and C); however, the number of
tubes was only moderately reduced at 1 µM for 24 h (P>0.05;
Fig. 2A and C). When the tubes
were exposed to 2 µM As2O3 for varying
durations, the number of tubes was decreased notably in all groups
except the 6 h group when compared with the control (P<0.05;
Fig. 2B and D). Furthermore, in
the As2O3 groups, the walls of the tubes
became thinner, more fragile and irregular, particularly at
concentrations of 4 and 5 µM for 24 h. Furthermore, the tubular
structures were severely damaged and cells could not form typical
structures correctly compared with the remaining groups, whereas in
the control group the wall of the tubes became thicker (Fig. 2A). Additionally, the structures
diminished gradually with the extension of time at 2 µM
As2O3 (Fig.
2B).
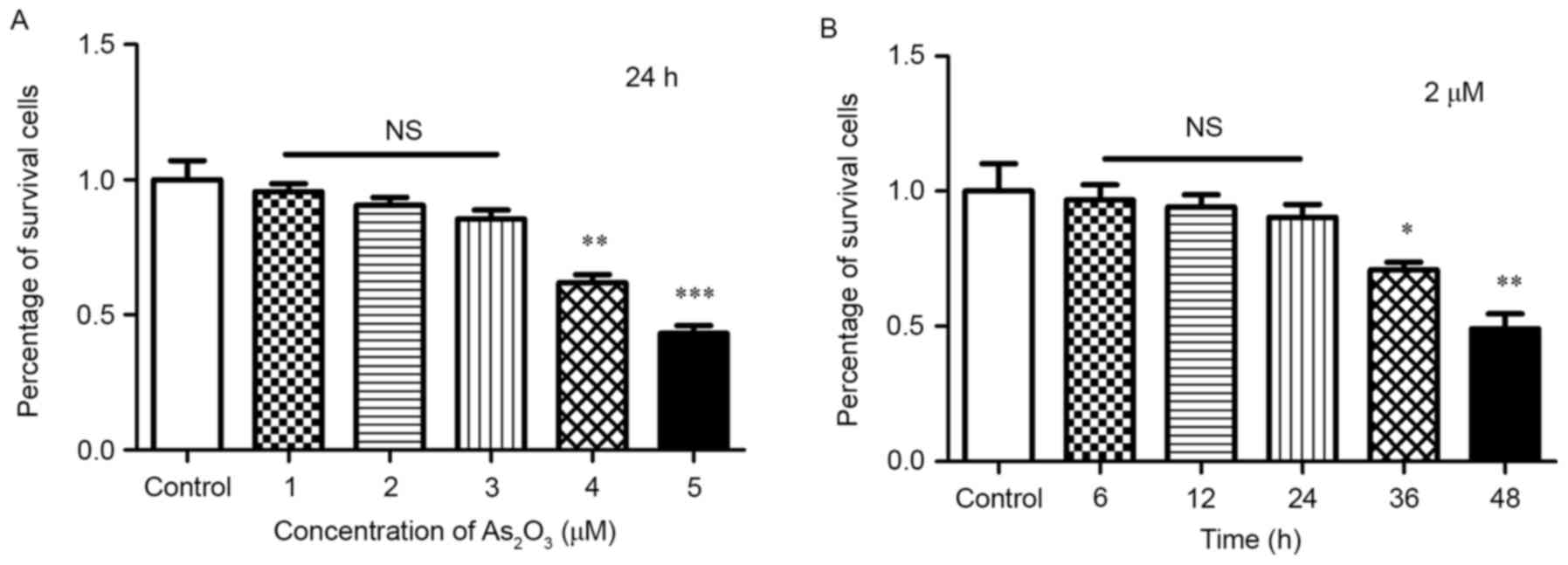
Although the viability of cells was reduced by
As2O3, the number of channels was divided by
the OD value, hence the inhibition of tube structures was not
affected by the reduction of total cell numbers. In addition, only
when the cells were exposed to 4 and 5 µM
As2O3 for 24 h and 2 µM for >36 h, the
cell viability was reduced significantly (P<0.05; Fig. 3), but did not decrease notably with
3 µM for 24 h and 2 µM within 24 h. No significant differences were
identified when compared with the control group (P>0.05;
Fig. 3). Therefore, the inhibition
of tube channels was earlier and more obvious than the reduction in
cell numbers.
As2O3 inhibits
the VM-associated molecules before apoptosis proteins emerge in
vitro
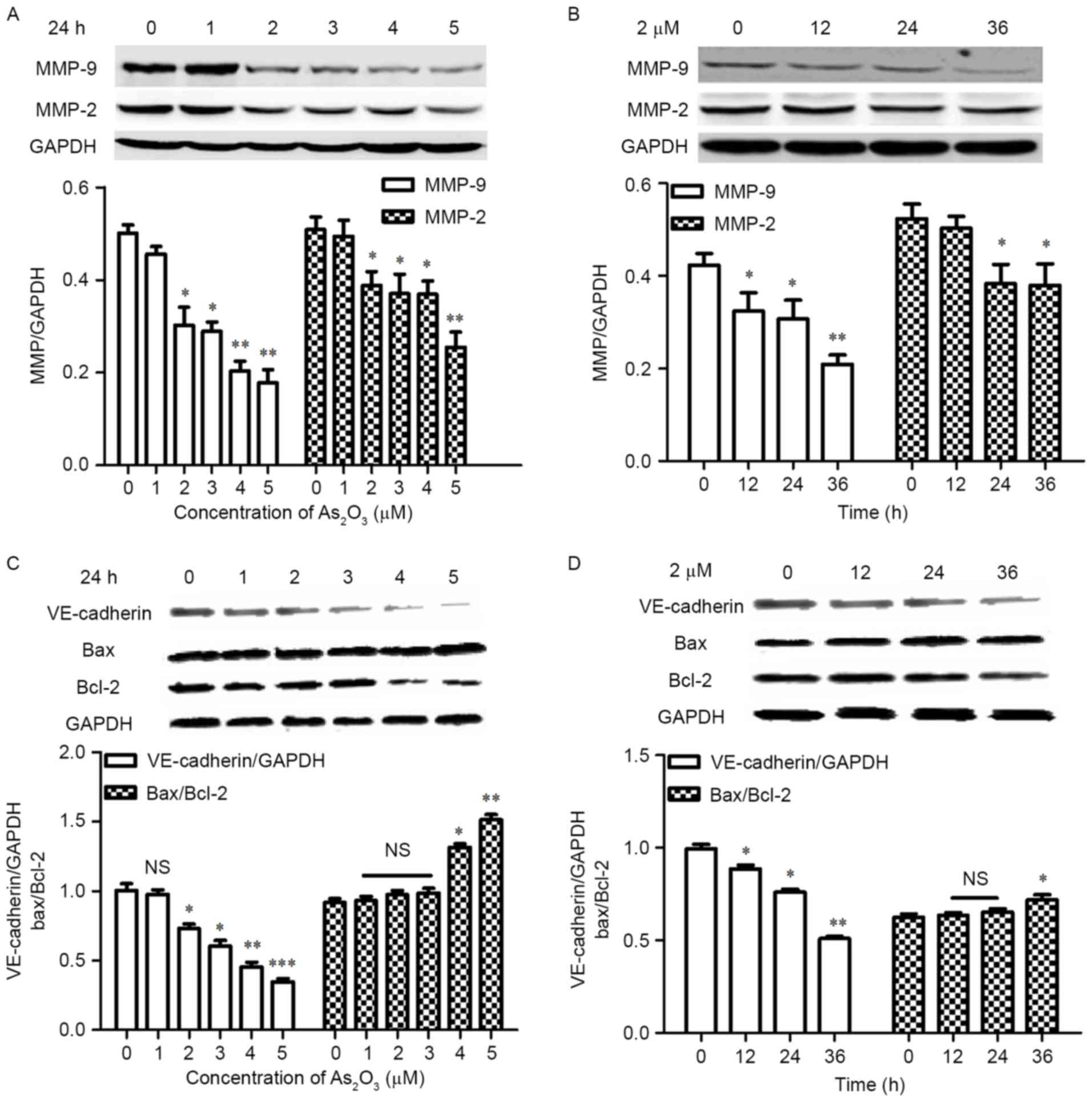
MMP-2 and MMP-9 are two important molecules in ECM
remodeling, which are vital in the formation of VM. VE-cadherin is
the key molecule in the malignant tumors capable of VM formation
and associated with tumor invasion (35,36).
The western blot results indicate that the expression levels of
MMP-2 and MMP-9 (Fig. 4A), and
VE-cadherin (Fig. 4C) decreased
significantly and in a concentration-dependent manner when cells
were treated with 2, 3, 4 and 5 µM As2O3 for
24 h (P<0.05). The ratio of Bax/Bcl-2 was upregulated
significantly at 4 and 5 µM for 24 h (P<0.05; Fig. 4C). When cells were exposed to 2 and
3 µM As2O3 for 24 h, the Bax/Bcl-2 ratio was
increased slightly, but no statistical difference was identified
when compared with the control group (P>0.05). The MMP-9
(Fig. 4B) and VE-cadherin
(Fig. 4D) expression levels
decreased significantly and in a time-dependent manner when the
cells were exposed to 2 µM As2O3 for varying
durations compared with the control (P<0.05), and MMP-2
expression levels decreased significantly at 24 and 36 h
(P<0.05; Fig. 4B). However, the
ratio of Bax/Bcl-2 was not significantly increased within 24 h
(P>0.05), although a significant difference was identified when
the duration was prolonged to 36 h (P<0.05; Fig. 4D), which indicated that the
inhibition of VM was associated with the decrease of VM-associated
proteins rather than cell apoptosis.
VM structures exist in HB xenografts
from mice models, and low-dose As2O3 inhibits
VM channels and VM-associated molecules in vivo
In vivo experiments demonstrated that VM
structures existed in the HepG2 cell tumor xenografts,
which were lined by tumor cells without participation of
endothelial cells and surrounded by a PAS+ material
(stained pink; Fig. 5A;
magnification, ×200), which was secreted by tumor cells.
PAS+/CD105− channels were validated by red
blood cells and tumor cells in the channel. This was predominantly
observed in the marginal zone of the tumor tissues.
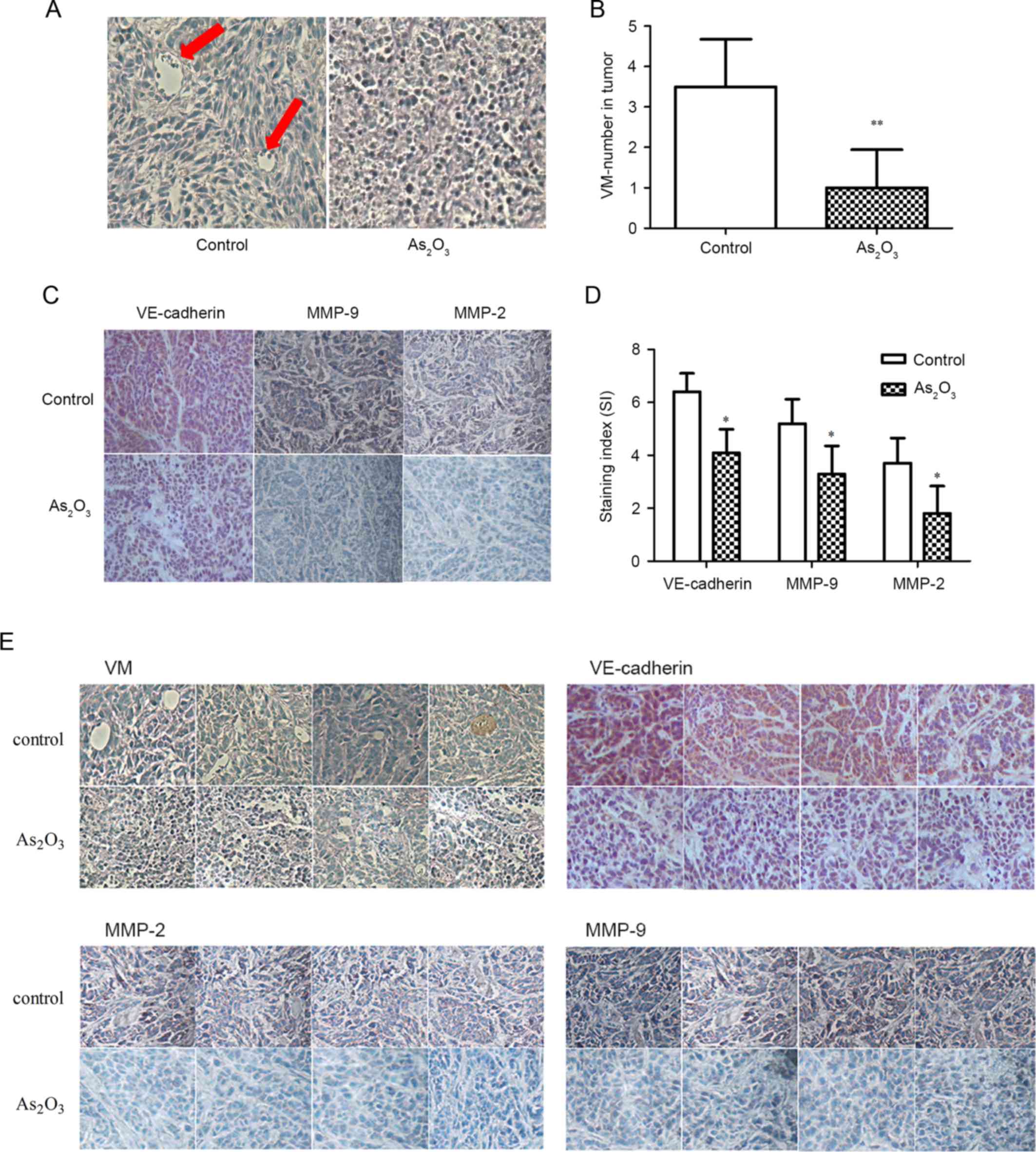 | Figure 5.Effect of As2O3
on VM and VM-associated proteins in vivo (magnification,
×200). (A) Evidence of VM in subcutaneous tumor transplants. The
channels (red arrow) lined with tumor cells were periodic
acid-Schiff+/CD105−. Red blood cells and
tumor cells are present in the channel and no hemorrhage or
necrotic cells were observed. (B) As2O3 (2
mg/kg) significantly decreased the number of VM structures. (C)
As2O3 (2 mg/kg) treatment affected the
expression level of VM-associated proteins in the xenografts. The
staining of VE-cadherin, MMP-2 and −9 in the control group is
stronger than that in the As2O3 group. (D)
The staining indices of VE-cadherin, MMP-2 and MMP-9 in the
As2O3 group were significantly lower than
those in the control. (E) PAS/CD105 and immunohistochemical
staining of the remaining four specimens. *P<0.05 and
**P<0.01 vs. control (specimen from the xenograft of mice who
had been administrated with 2 ml PBS peritoneal injection).
As2O3, arsenic trioxide; VM, vasculogenic
mimicry; VE, MMP, matrix metalloproteinase; Bax, Bcl-2-associated X
protein; Bcl-2, B-cell lymphoma 2; VE, vascular endothelial; NS,
not statistically significant (P>0.05). |
In previous studies, high-dose
As2O3 significantly inhibited tumor growth.
In the present study, low concentration of
As2O3 (2 mg/kg) was administrated
intraperitoneally to the mice to observe the effect on VM; this
dosage is markedly lower than that used in previous studies, which
may lead to cell toxicity. An equal quantity of PBS served as the
control. The results indicate that the VM structure was observed in
all the control specimens (5/5); however, in the
As2O3 group, the cells were depressed and
could not form channels correctly, and VM structures were only
observed in two tissue sections. The number of VM channels in the
As2O3 group (1.0±0.95) was decreased by 72%
compared with the control group (3.56±1.81), and a statistically
significant difference was observed (P<0.01; Fig. 5B). No obvious necrotic areas were
observed in the visual fields under the microscope.
Additionally, IHC analysis demonstrated that the
staining intensity of VM-associated markers in the control group
was brown, which was stronger than that of the
As2O3 group, and the percentage of positive
cells in the control group was markedly more than that of the
As2O3 group (Fig. 5C). The staining index in the
As2O3 group was significantly lower than that
of the control (P<0.05; Fig.
5D). These results indicate that As2O3
inhibits the VM structures by decreasing VM-associated proteins
in vivo at a low concentration.
Low-dose As2O3
inhibits tumor growth in HB mouse models
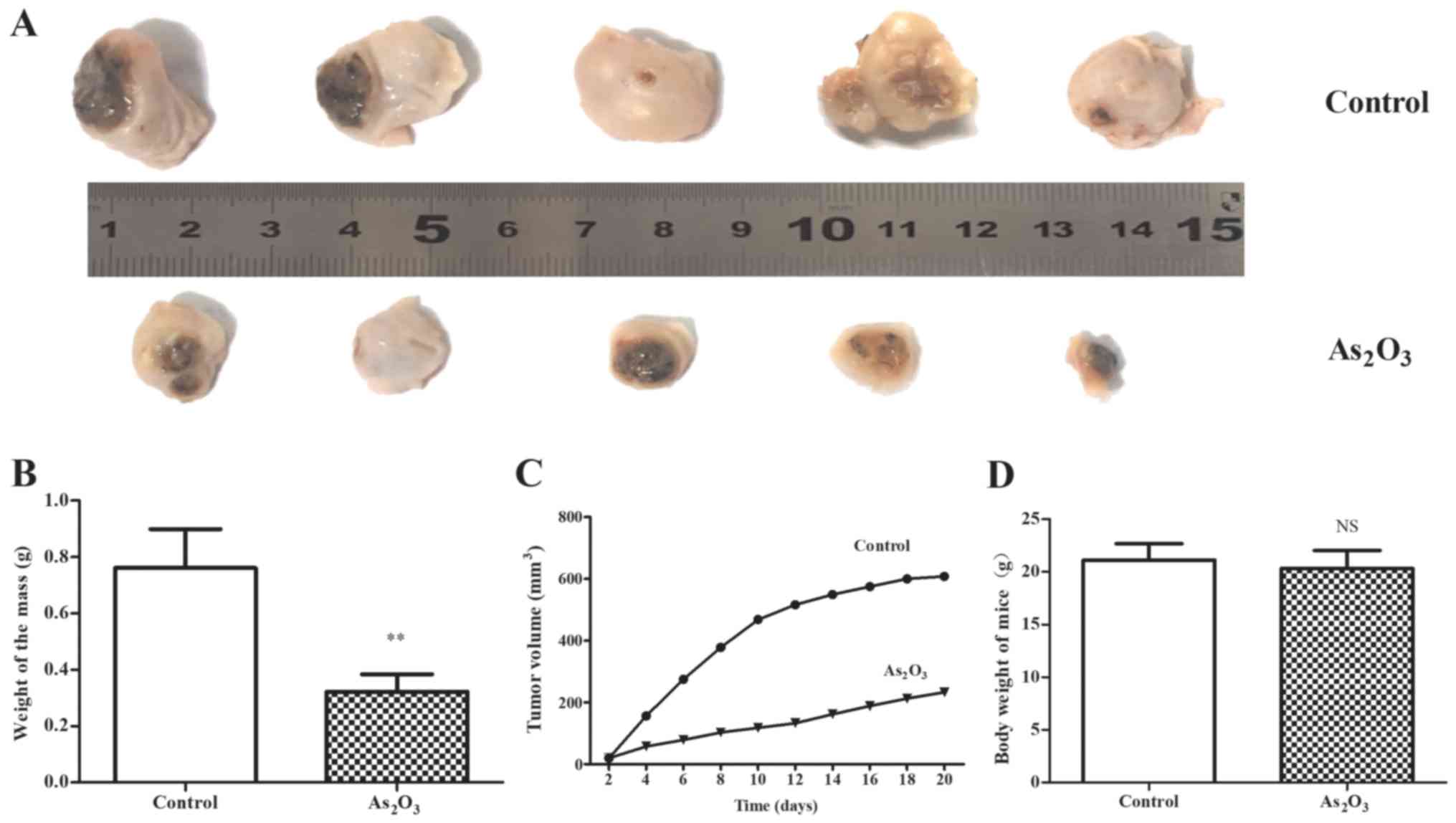
In the in vivo study, 2 mg/kg
As2O3 exhibited significant anti-tumor
efficacy on the mouse model of HepG2 cells. The results
indicated that the tumors in the
As2O3-treated group were notably smaller than
that of the control (Fig. 6A), and
the increase of tumor volume in the As2O3
group was notably slower than that of the control group. The tumor
growth curve in the As2O3 group was smooth,
while it was steep in the control group, particularly in the first
10 days (Fig. 6C). Additionally,
the weight of tumors in the As2O3 group was
statistically lower than that of the control group (P<0.01;
Fig. 6B). This may be attributed
to the inhibition of VM, which is the main blood supply of the
tumor during the early stage of tumorigenesis. In addition, the
loss of body weight in the As2O3 group was
not significant (P>0.05) and no other severe adverse effects
were observed in the nude mice treated with
As2O3.
Discussion
HB is the most common type of pediatric liver
malignancy, typically affecting children aged <5 years. HB are
divided into two histological subtypes based on the level of cell
differentiation (37), including
epithelial and mixed epithelial mesenchymal. The majority of HB is
epithelial, consisting of embryonal and fetal cells, such as
HepG2 cells. HB is a tumor characterized by
hypervascularity (38) and the
incidence of HB has increased in the past 30 years (39). The overall survival of this disease
remains poor and, due to its rarity, there is little experience of
treating HB. Therefore, there is an urgent requirement to establish
an effective treatment strategy for HB.
VM is associated with high tumor grade, invasion,
metastasis and short survival, and is a marker of poor clinical
prognosis in adult liver cancer, such as hepatocellular carcinoma
(34,40). In the present study, the epithelial
HB cell line HepG2 cells (41,42)
were found to form the typical structure of VM in vivo and
in vitro, which may indicate why HB tends to metastasize and
relapse.
Formation of VM is a complex process that involves
various mechanisms, including EPH receptor A2 (EphA2), cancer stem
cells, epithelial-mesenchymal transition, the phosphoinositide
3-kinase signaling pathway, tumor cell plasticity, remodeling of
the ECM and microenvironments (such as hypoxia). ECM degradation
and remodeling are important in the formation of VM channels, a
variety of molecular and signaling pathways, including MMPs and
VE-cadherin, participate in remodeling of the ECM (35,36,43,44).
Among MMPs, MMP-2 and MMP-9 are two important enzymes, which are
involved in the remodeling of ECM and are key mediators of
invasion, metastasis, tumor angiogenesis and facilitate VM
formation (45,46). MMP-2 overexpression may be
associated with lymph node metastasis from gastric carcinoma and
has been reported to be linked to the recurrence of breast cancer
(47,48). VE-cadherin is associated with tumor
invasion and is considered to be essential for VM network
formation. Plastic tumor cells lacking VE-cadherin are incapable of
VM tube formation (36). By
contrast, upregulation of VE-cadherin, EphA2, MMP-2 and MMP-9
promotes the formation of VM (49). Therefore, VM-targeting strategies
should include these markers to combat the recurrence and
metastasis of malignant tumors. Certain experimental evidence
indicates that curcumin, thalidomide and gene deletion techniques
may successfully inhibit VM and tumor growth. However, the 5-year
survival rate of patients with VM remains poor, and there are few
effective and feasible methods to treat tumors exhibiting VM.
To the best of our knowledge, this is the first
study discussing the effect of As2O3 on VM
formation. In the present study, a low concentration of
As2O3 was found to inhibit the formation of
VM structures in vivo and in vitro, and
As2O3-treated HepG2 cells
exhibited a significantly lower VM capacity than those of the
control group. As2O3 destroys the formation
of the tubular structures in vitro at a concentration of
<3 µM within 24 h before cell toxicity appeared, in a time- and
concentration-dependent manner. During the in vivo
experiments, the VM channels were decreased markedly following
treatment with 2 mg/kg As2O3, twice a day for
a total of 20 days, which was markedly lower than that of
clinically tolerable concentrations (50). Additionally, tumor growth was
suppressed significantly without obvious adverse reactions.
Furthermore, the expression levels of VE-cadherin,
MMP-2 and MMP-9 were decreased significantly with the treatment of
low dosage As2O3 in vivo and in
vitro. Previous studies indicated that
As2O3 upregulates the ratio of Bax/Bcl-2 and
induces cell apoptosis (29,51,52);
the present in vitro results had the same conclusion,
although the ratio of Bax/Bcl-2 increased significantly only when
exposed to higher concentrations and longer exposure of
As2O3 (in cells exposed to 2 and 3 µM
As2O3 for 24 h, the ratio of Bax/Bcl-2
increased only slightly). However, the expression levels of
VE-cadherin, MMP-2 and MMP-9 were decreased notably in cells
exposed to 2 and 3 µM As2O3 for 24 h, which
indicated that As2O3 inhibits tube structures
in HepG2 cells via inhibition of VM-associated proteins
rather than by inducing cell apoptosis <3 µM within 24 h. IHC
results from the in vivo study indicated that the staining
intensity of VE-cadherin, MMP-2 and MMP-9 were obviously weaker in
the 2 mg/kg As2O3 group when compared with
that of control group, and no necrotic areas were observed in the
tissues. These results indicate that As2O3
inhibits VE-cadherin, MMP-2 and MMP-9, and thus the formation of VM
and tumor growth at low concentrations without cell toxicity.
Complete surgical resection of the tumor is
essential for survival of patients with HB (53,54).
HB is a chemotherapy-sensitive tumor and preoperative chemotherapy
effectively improves the surgical resectability of the tumor and
prolongs the survival time of patients, even in cases of metastasis
(55). In previous decades,
As2O3 treatment has exerted therapeutic
effects on adult liver malignant tumors and induced attenuation of
invasion potential; however, the application of this drug has been
limited due to the dose-dependent heart, liver and kidney toxicity
(56), therefore the therapeutic
window of As2O3 is particularly narrow.
Establishing the appropriate dosage of As2O3
without significant adverse reactions has become a breakthrough in
the treatment of malignant tumors. Furthermore, the drug resistance
of HB often occurs after four cycles of chemotherapy (57), therefore avoiding this issue
presents a therapeutic challenge.
In conclusion, the present results demonstrate that
low-dose As2O3 is effective in reducing the
formation of VM channels and tumor growth in HB mouse models by
inhibiting VM-associated proteins without causing cytotoxicity.
Therefore, As2O3 may present as a promising
candidate drug to treat HB by targeting VM.
Acknowledgements
The present study was supported by the doctoral
program of higher education (grant no. 20122307110014).
References
|
1
|
Heck JE, Meyers TJ, Lombardi C, Park AS,
Cockburn M, Reynolds P and Ritz B: Case-control study of birth
characteristics and the risk of hepatoblastoma. Cancer Epidemiol.
37:390–395. 2013. View Article : Google Scholar :
|
|
2
|
Stiller CA, Pritchard J and
Steliarova-Foucher E: Liver cancer in European children: Incidence
and survival, 1978–1997. Report from the Automated Childhood Cancer
Information System project. Eur J Cancer. 42:2115–2123. 2006.
View Article : Google Scholar
|
|
3
|
Czaudernaa P, Haeberle B, Hiyama E,
Rangaswami A, Krailo M, Maibach R, Rinaldi E, Feng YR, Aronson D,
Malogolowkin M, et al: The Children's Hepatic tumors International
Collaboration (CHIC): Novel global rare tumor database yields new
prognostic factors in hepatoblastoma and becomes a research model.
Eur J Cancer. 52:92–101. 2016. View Article : Google Scholar
|
|
4
|
Tanimura M, Matsui I, Abe J, Ikeda H,
Kobayashi N, Ohira M, Yokoyama M and Kaneko M: Increased risk of
hepatoblastoma among immature children with a lower birth weight.
Cancer Res. 58:3032–3035. 1998.
|
|
5
|
Pateva IB, Egler RA and Stearns DS:
Hepatoblastoma in an 11-year-old. Case report and a review of the
literature. Medicine (Baltimore). 96:e58582017. View Article : Google Scholar :
|
|
6
|
Park KW, Seo CJ, Yun DY, Kim MK, Kim BS,
Han YS, Oh KH and Lee CH: A case of hepatoblastoma misdiagnosed as
combined hepatocellular carcinoma and cholangiocarcinoma in an
adult. Clin Mol Hepatol. 21:300–308. 2015. View Article : Google Scholar :
|
|
7
|
Maniotis AJ, Folberg R, Hess A, Seftor EA,
Gardner LM, Pe'er J, Trent JM, Meltzer PS and Hendrix MJ: Vascular
channel formation by human melanoma cells in vivo and in vitro:
Vasculogenic mimicry. Am J Pathol. 155:739–752. 1999. View Article : Google Scholar :
|
|
8
|
Zhang S, Guo H, Zhang D, Zhang W, Zhao X,
Ren Z and Sun B: Microcirculation patterns in different stages of
melanoma growth. Oncol Rep. 15:15–20. 2006.
|
|
9
|
Sood AK, Seftor EA, Fletcher MS, Gardner
LM, Heidger PM, Buller RE, Seftor RE and Hendrix MJ: Molecular
determinants of ovarian cancer plasticity. Am J Pathol.
158:1279–1288. 2001. View Article : Google Scholar :
|
|
10
|
Fujimoto A, Onodera H, Mori A, Nagayama S,
Yonenaga Y and Tachibana T: Tumour plasticity and extravascular
circulation in ECV304 human bladder carcinoma cells. Anticancer
Res. 26:59–69. 2006. View Article : Google Scholar
|
|
11
|
Baeten CI, Hillen F, Pauwels P, de Bruine
AP and Baeten CG: Prognostic role of vasculogenic mimicry in
colorectal cancer. Dis Colon Rectum. 52:2028–2035. 2009. View Article : Google Scholar
|
|
12
|
Li M, Gu Y, Zhang Z, Zhang S, Zhang D,
Saleem AF, Zhao X and Sun B: Vasculogenic mimicry: A new prognostic
sign of gastric adenocarcinoma. Pathol Oncol Res. 16:259–266. 2010.
View Article : Google Scholar
|
|
13
|
Wu S, Yu L, Cheng Z, Song W, Zhou L and
Tao Y: Expression of maspin in non-small cell lung cancer and its
relationship to vasculogenic mimicry. J Huazhong Univ Sci Technolog
Med Sci. 32:346–352. 2012. View Article : Google Scholar
|
|
14
|
Van der Schaft DW, Hillen F, Pauwels P,
Kirschmann DA, Castermans K, Egbrink MG, Tran MG, Sciot R, Hauben
E, Hogendoorn PC, et al: Tumor cell plasticity in Ewing sarcoma, an
alternative circulatory system stimulated by hypoxia. Cancer Res.
65:11520–11528. 2005. View Article : Google Scholar
|
|
15
|
Vartanian AA, Stepanova EV, Gutorov SL,
Solomko ESh, Grigorieva IN, Sokolova IN, Baryshnikov AY and
Lichinitser MR: Prognostic significance of periodic
acid-Schiff-positive patterns in clear cell renal cell carcinoma.
Can J Urol. 16:4726–4732. 2009.
|
|
16
|
Francescone R, Scully S, Bentley B, Yan W,
Taylor SL, Oh D, Moral L and Shao R: Glioblastoma-derived tumor
cells induce vasculogenic mimicry through Flk-1 protein activation.
J Biol Chem. 287:24821–24831. 2012. View Article : Google Scholar :
|
|
17
|
Folberg R and Maniotis AJ: Vasculogenic
mimicry. APMIS. 112:508–525. 2004. View Article : Google Scholar
|
|
18
|
Hendrix MJ, Seftor EA, Hess AR and Seftor
RE: Vasculogenic mimicry and tumour-cell plasticity: Lessons from
melanoma. Nat Rev Cancer. 3:411–421. 2003. View Article : Google Scholar
|
|
19
|
Cascone T and Heymach JV: Targeting the
angiopoietin/Tie2 pathway: Cutting tumor vessels with a
double-edged sword? J Clin Oncol. 30:441–444. 2012. View Article : Google Scholar
|
|
20
|
Jain RK, Duda DG, Clark JW and Loeffler
JS: Lessons from phase III clinical trials on anti-VEGF therapy for
cancer. Nat Clin Pract Oncol. 3:24–40. 2006. View Article : Google Scholar
|
|
21
|
Goel S, Duda DG, Xu L, Munn LL, Boucher Y,
Fukumura D and Jain RK: Normalization of the vasculature for
treatment of cancer and other diseases. Physiol Rev. 91:1071–1121.
2011. View Article : Google Scholar :
|
|
22
|
Zhang Y, Zhang Z, Li J, Li L, Han X, Han
L, Hu L, Wang S, Zhao Y, Li X, et al: Long-term efficacy and safety
of arsenic trioxide for first-line treatment of elderly patients
with newly diagnosed acute promyelocytic leukemia. Cancer.
119:115–125. 2013. View Article : Google Scholar
|
|
23
|
Breitenbach Lallemand V, Guillemin MC,
Janin A, Daniel MT, Degos L, Kogan SC, Bishop JM and de Thé H:
Retinoic acid and arsenic synergize to eradicate leukemic cells in
a mouse model of acute promyelocytic leukemia. J Exp Med.
189:1043–1052. 1999. View Article : Google Scholar :
|
|
24
|
Pettersson HM, Pietras A, Persson
Munksgaard M, Karlsson J, Johansson L, Shoshan MC and Påhlman S:
Arsenic trioxide is highly cytotoxic to small cell lung carcinoma
cells. Mol Cancer Ther. 8:160–170. 2009. View Article : Google Scholar
|
|
25
|
Yu J, Qian HL, Li Y, Wang Y, Zhang X,
Liang X, Fu M and Lin C: Arsenic trioxide
(As2O3) reduces the invasive and metastatic
properties of cervical cancer cells in vitro and in vivo. Gynecol
Oncol. 106:400–406. 2007. View Article : Google Scholar
|
|
26
|
Du J, Zhou N, Liu H, Jiang F, Wang Y, Hu
C, Qi H, Zhong C, Wang X and Li Z: Arsenic induces functional
re-expression of estrogen receptor α by demethylation of DNA in
estrogen receptor-negative human breast cancer. PLoS One.
7:e359572012. View Article : Google Scholar :
|
|
27
|
Munshi NC, Tricot G, Desikan R, Badros A,
Zangari M, Toor A, Morris C, Anaissie E and Barlogie B: Clinical
activity of arsenic trioxide for the treatment of multiple myeloma.
Leukemia. 16:1835–1837. 2002. View Article : Google Scholar
|
|
28
|
Ma ZB, Xu HY, Jiang M, Yang YL, Liu LX and
Li YH: Arsenic trioxide induces apoptosis of human gastrointestinal
cancer cells. World J Gastroenterol. 20:5505–5510. 2014. View Article : Google Scholar :
|
|
29
|
Alarifi S, Ali D, Alkahtani S, Siddiqui MA
and Ali BA: Arsenic trioxide-mediated oxidative stress and
genotoxicity in human hepatocellular carcinoma cells. Onco Targets
Ther. 6:75–84. 2013.
|
|
30
|
Lee JC, Lee HY, Moon CH, Lee SJ, Lee WH,
Cha HJ, Park SC, Lee YH, Park HJ, Song HT and Min YJ: Arsenic
trioxide as a vascular disrupting agent: Synergistic effect with
irinotecan on tumor growth delay in a CT26 allograft model. Transl
Oncol. 6:83–91. 2013. View Article : Google Scholar :
|
|
31
|
Xiao YF, Wu DD, Liu SX, Chen X and Ren LF:
Effect of arsenic trioxide on vascular endothelial cell
proliferation and expression of vascular endothelial growth factor
receptors Flt-1 and KDR in gastric cancer in nude mice. World J
Gastroenterol. 13:6498–6505. 2007. View Article : Google Scholar :
|
|
32
|
Thomas-Schoemann A, Batteux F, Mongaret C,
Nicco C, Chéreau C, Annereau M, Dauphin A, Goldwasser F, Weill B,
Lemare F and Alexandre J: Arsenic trioxide exerts antitumor
activity through regulatory T cell depletion mediated by oxidative
stress in a murine model of colon cancer. J Immunol. 189:5171–5177.
2012. View Article : Google Scholar
|
|
33
|
Verma RJ, Vasu A and Sayyed AA: Arsenic
toxicity in mice and its possible amelioration. J Environ Sci
(China). 16:447–453. 2004.
|
|
34
|
Sun B, Zhang S, Zhang D, Du J, Guo H, Zhao
X, Zhang W and Hao X: Vasculogenic mimicry is associated with high
tumor grade, invasion and metastasis, and short survival in
patients with hepatocellular carcinoma. Oncol Rep. 16:693–698.
2006.
|
|
35
|
Zhao N, Sun H, Sun B, Zhu D, Zhao X, Wang
Y, Gu Q, Dong X, Liu F, Zhang Y and Li X: miR-27a-3p suppresses
tumor metastasis and VM by down-regulating VE-cadherin expression
and inhibiting EMT: An essential role for Twist-1 in HCC. Sci Rep.
6:230912016. View Article : Google Scholar :
|
|
36
|
Hendrix MJ, Seftor EA, Meltzer PS, Gardner
LM, Hess AR, Kirschmann DA, Schatteman GC and Seftor RE: Expression
and functional significance of VE-cadherin in aggressive human
melanoma cells: Role in vasculogenic mimicry. Proc Natl Acad Sci
USA. 98:8018–8023. 2001. View Article : Google Scholar :
|
|
37
|
Bell D, Ranganathan S, Tao J and Monga SP:
Novel advances in understanding of molecular pathogenesis of
hepatoblastoma: A Wnt/β-catenin perspective. Gene Expr. 17:141–154.
2017. View Article : Google Scholar
|
|
38
|
Dong R, Liu GB, Liu BH, Chen G, Li K,
Zheng S and Dong KR: Targeting long non-coding RNA-TUG1 inhibits
tumor growth and angiogenesis in hepatoblastoma. Cell Death Dis.
7:e22782016. View Article : Google Scholar :
|
|
39
|
Czauderna P, Lopez-Terrada D, Hiyama E,
Häberle B, Malogolowkin MH and Meyers RL: Hepatoblastoma state of
the art: Pathology, genetics, risk stratifcation, and chemotherapy.
Curr Opini Pediatr. 26:19–28. 2014. View Article : Google Scholar
|
|
40
|
Sun T, Zhao N, Zhao XL, Gu Q, Zhang SW,
Che N, Wang XH, Du J, Liu YX and Sun BC: Expression and functional
significance of Twist1 in hepatocellular carcinoma: Its role in
vasculogenic mimicry. Hepatology. 51:545–556. 2010. View Article : Google Scholar
|
|
41
|
López-Terrada D, Cheung SW, Finegold MJ
and Knowles BB: HepG2 is a hepatoblastoma-derived cell
line. Hum Pathol. 40:1512–1515. 2009. View Article : Google Scholar
|
|
42
|
Xia Z, Zhang N and Ding DK: Proliferation
and migration of hepatoblastoma cells are mediated by IRS-4 via
PI3K/Akt pathways. Int J Clin Exp Med. 7:3763–3769. 2014.
|
|
43
|
Sood AK, Fletcher MS, Coffin JE, Yang M,
Seftor EA, Gruman LM, Gershenson DM and Hendrix MJ: Functional role
of matrix metalloproteinases in ovarian tumor cell plasticity. Am J
Obstet Gynecol. 190:899–909. 2004. View Article : Google Scholar
|
|
44
|
Zhang S, Zhang D and Sun B: Vasculogenic
mimicry: Current status and future prospects. Cancer Lett.
254:157–164. 2007. View Article : Google Scholar
|
|
45
|
Chang C and Werb Z: The many faces of
metalloproteases: Cell growth, invasion, angiogenesis and
metastasis. Trends Cell Biol. 11:S37–S43. 2001. View Article : Google Scholar :
|
|
46
|
Chen LX, He YJ, Zhao SZ, Wu JG, Wang JT
and Zhu LM: Inhibition of tumor growth and vasculogenic mimicry by
curcumin through downregulation of the EphA2/PI3K/MMP pathway in a
murine choroidal melanoma model. Cancer Biol Ther. 11:229–235.
2011. View Article : Google Scholar
|
|
47
|
Jinga DC, Blidaru A, Condrea I, Ardeleanu
C, Dragomir C, Szegli G, Stefanescu M and Matache C: MMP-9 and
MMP-2 gelatinases and TIMP-1 and TIMP-2 inhibitors in breast
cancer: Correlations with prognostic factors. J Cell Mol Med.
10:499–510. 2006. View Article : Google Scholar
|
|
48
|
Mönig SP, Baldus SE, Hennecken JK,
Spiecker DB, Grass G, Schneider PM, Thiele J, Dienes HP and
Hölscher AH: Expression of MMP-2 is associated with progression and
lymph node metastasis of gastric carcinoma. Histopathology.
39:597–602. 2001. View Article : Google Scholar
|
|
49
|
Kirschmann DA, Seftor EA, Hardy KM, Seftor
RE and Hendrix MJ: Molecular pathways: Vasculogenic mimicry in
tumor cells: Diagnostic and therapeutic implications. Clin Cancer
Res. 18:2726–2732. 2012. View Article : Google Scholar :
|
|
50
|
Øra I, Bondesson L, Jönsson C, Ljungberg
J, Pörn-Ares I, Garwicz S and Pâhlman S: Arsenic trioxide inhibits
neuroblastoma growth in vivo and promotes apoptotic cell death in
vitro. Biochem Biophys Res Commun. 277:179–185. 2000. View Article : Google Scholar
|
|
51
|
Siu KP, Chana JY and Fung KP: Effect of
arsenic trioxide on human hepatocellular carcinoma HepG2 cells:
Inhibition of proliferation and induction of apoptosis. Life Sci.
71:275–285. 2002. View Article : Google Scholar
|
|
52
|
Wang Y, Bai C, Guan H, Chen R, Wang X,
Wang B, Jin H and Piao F: Subchronic exposure to arsenic induces
apoptosis in the hippocampus of the mouse brains through the
Bcl-2/Bax pathway. J Occup Health. 57:212–221. 2015. View Article : Google Scholar
|
|
53
|
Venkatramani R, Furman WL, Fuchs J,
Warmann SW and Malogolowkin MH: Current and future management
strategies for relapsed or progressive hepatoblastoma. Paediatr
Drugs. 14:221–232. 2012. View Article : Google Scholar
|
|
54
|
von Schweinitz D: Management of liver
tumors in childhood. Semin Pediatr Surg. 15:17–24. 2006. View Article : Google Scholar
|
|
55
|
Zheng MH, Zhang L, Gu DN, Shi HQ, Zeng QQ
and Chen YP: Hepatoblastoma in adult: Review of the literature. J
Clin Med Res. 1:13–16. 2009.
|
|
56
|
Miller WH Jr, Schipper HM, Lee JS, Singer
J and Waxman S: Mechanisms of action of arsenic trioxide. Cancer
Res. 62:3893–3903. 2002.
|
|
57
|
Von Schweinitz D, Hecker H,
Schmidt-von-Arndt G and Harms D: Prognostic factors and staging
systems in childhood hepatoblastoma. Int J Cancer. 74:593–599.
1997. View Article : Google Scholar
|