Introduction
Scopolamine (SCO), a non-selective muscarinic
acetylcholine receptor antagonist, is known to interfere with
cholinergic transmission in the brain (1,2).
Recently, we have reported that the SCO-induced dysregulation of
cholinergic activity in the hippocampus leads to interference in
processes of learning and memory in mice (3–5).
Thus, SCO has been used as a pharmacological tool for the
development of experimental animal models of cognitive deficits;
many researchers have identified the models as a candidate for the
treatment of Alzheimer's disease (6–8).
The hippocampus or hippocampal formation, which
consists of the hippocampus proper (CA1-3 areas) and dentate gyrus,
displays critical roles in cognitive processes such as learning,
memory consolidation, and information retrieval (9–12).
Memory loss followed by cognitive decline is found to be a common
feature of various neurological disorders like Alzheimer's disease
(13). The cholinergic system has
long been known to be critically involved in hippocampal-dependent
cognitive function (14,15). Deficits in the hippocampal
cholinergic system can affect learning and memory; namely, the loss
of cholinergic function in the hippocampus is correlated with
significant cognitive impairments (16,17).
The cytoskeleton extends throughout the cytoplasm of
cell and provides a number of important functions, such as the
maintenance of cellular shape, holding cellular organelles in
place, and assisting in the transportation of communication signals
between cells (18). Abnormalities
of the neuronal cytoskeleton frequently represent a key feature of
Alzheimer's disease, in which the disruption of the normal
cytoskeletal framework of neurons typically triggers dystrophic
neurites (19).
Neurofilaments (NFs) do not only support the
morphology of neurons as cytoskeletal components but also play a
fundamental role in axonal transport (18). NFs are composed of three major
subunits, neurofilament-200 kDa (NF-H), neurofilament-165 kDa
(NF-M) and neurofilament-68 kDa (NF-L); according to observed
molecular weight (20). NF-H and
NF-M are involved in the stabilization of newly-sprouted axonal
processes (21,22), and NF-L is expressed in axonal
growth cones (23). In particular,
NF-H plays an important role in the stabilization and maturation of
pre-existing connections (24).
Previous studies have suggested that NFs are closely related to
neurodegenerative diseases (25–28).
The pathological disruption of axonal NFs results in the breakdown
and fasciculation of the filaments, and its most notable
characteristic is whorls of NFs and ring-like structures in swollen
axons (28,29).
However, few studies regarding changes in NFs
expressions in animal models of SCO-induced amnesia have been
reported. Therefore, in this study, we examined chronological
changes in NFs immunoreactivities and levels of NFs proteins in the
mouse hippocampus, as well as in memory impairments following
chronic systemic treatment with SCO.
Materials and methods
Experimental animals and SCO
treatment
Male ICR mice (B.W., 25–30 g; 8 weeks of age) were
purchased from the Orient Bio Inc. (Seongnam, South Korea). The
procedures for animal handling and care, and experimental protocols
were reviewed and approved based on ethical procedures and
scientific care by the Kangwon National University-Institutional
Animal Care and Use Committee (KW-130424-2).
Mice were intraperitoneally injected with 1 mg/kg of
SCO (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), once daily,
for 1, 2, 3 and 4 weeks (n=14 at each point in time). Dose
of SCO was selected based on previous studies (30,31).
The control mice (n=14 at each point in time) were injected
with the same volume of saline (pH 7.4). The mice were sacrificed
1, 2, 3 and 4 weeks after SCO or saline treatment.
Passive avoidance test
Short-term memory ability was evaluated by assessing
the latency of passive avoidance test. Animals (n=7) in each group
were tested 1 day before sacrifice following training for 1 day. In
short, according to our published procedure (32), the test was performed with an
apparatus that consisted of two compartments (light and dark) with
a grid floor (GEM 392; San Diego Instruments., San Diego, CA, USA).
In training session, mice were allowed to explore environments in
both light and dark compartments for 1 min and given an inescapable
foot-shock (0.3 mA for 3 sec) after entering the dark compartment.
Test session was performed 15 min after the training session using
the same paradigm, but without applying the foot-shock. The
interval between the starting of the test session and mouse's entry
into the dark compartment was defined as the latency time of
passive avoidance test. When mouse did not enter the dark room
within 180 sec, the latency was recorded as 180 sec.
Water maze performance
The animals used in the passive avoidance test were
used for spatial learning and memory test using the Morris water
maze task according to our published protocol (4). In short, a circular pool (90 cm in
diameter and 45 cm height) filled with water was conceptually
divided into four quadrants, and a platform (6 cm in diameter and
29 cm in height) was placed in one quadrant 1 cm below the water
surface. Test was conducted on the last day of every week after
saline or SCO treatment, and training was conducted before the test
for 3 consecutive days (saline and SCO were administered
continuously during training and test periods). Mice (n=7 in
each group) were allowed to swim for 120 sec to search for the
hidden platform. If they failed to locate the platform within 120
sec, escape would be assisted and escape latency was recorded as
120 sec. At the end of each trial, each mouse would stay on the
platform for 3 sec. Escape latency (the time taken to find the
platform) was recorded with Noldus Ethovision video tracking system
(EthoVision XT; Noldus Information Technology, Wageningen, The
Netherlands).
Western blot analysis
NFs levels in the hippocampal formation (n=7
at each point in time) were analyzed according to our published
procedure (32). In brief,
hippocampal tissues were homogenized and the protein level was
determined using a Micro bicinchoninic acid (BCA) protein assay kit
(Pierce Biotechnology, Inc., Rockford, IL, USA). The membranes were
incubated with rabbit anti- NF-H, NF-M and NF-L (1:1,000; Chemicon
International, Inc., Billerica, MA, USA), and exposed to peroxidase
conjugated goat anti-rabbit IgG (Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) and an enhanced luminol-based chemiluminescent
(ECL) kit (Pierce Biotechnology, Inc.).
The result of the western blot analysis was scanned,
and densitometric analysis for the quantification of the bands was
done using Scion Image software (Scion Corp., Frederick, MD, USA),
which was used to count ROD. A ratio of the ROD was calibrated as
%, with control group designated as 100%.
Immunohistochemistry
According to our published procedure (33), in brief, mice (n=7 at each
point in time) were anesthetized with pentobarbital sodium (40
mg/kg, JW Pharmaceutical Co., Ltd., Seoul, Korea) and perfused with
4% paraformaldehyde. Brain tissues were serially sectioned in a
cryostat (Leica, Wetzlar, Germany) into 30-µm coronal sections. The
sections were incubated with diluted rabbit anti-NF-H (1:1,000;
Chemicon International, Inc., Temecula, CA, USA), NF-M (1:1,000;
Chemicon International, Inc.) or NF-L (1:1,000; Chemicon
International, Inc.), and exposed to biotinylated goat anti-rabbit
IgG and streptavidin peroxidase complex (Vector Laboratories, Inc.,
Burlingame, CA, USA). Finally, they were visualized with
3,3′-diaminobenzidine tetrachloride (Sigma-Aldrich; Merck
KGaA).
Statistical analysis
All data are presented as mean ± SEM. The normality
test was performed using Kolmogorov and Smirnov test for testing
normal distributions, and Bartlett test for testing identical
standard distributions. All data passed normality test. A
multiple-sample comparison was applied to test the differences
between groups (ANOVA and the Tukey multiple range test as post hoc
test using the criterion of the least significant differences).
Statistical analysis was performed using GraphPad Instat (Instat
Statistics; GraphPad Software, Inc., La Jolla, CA, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
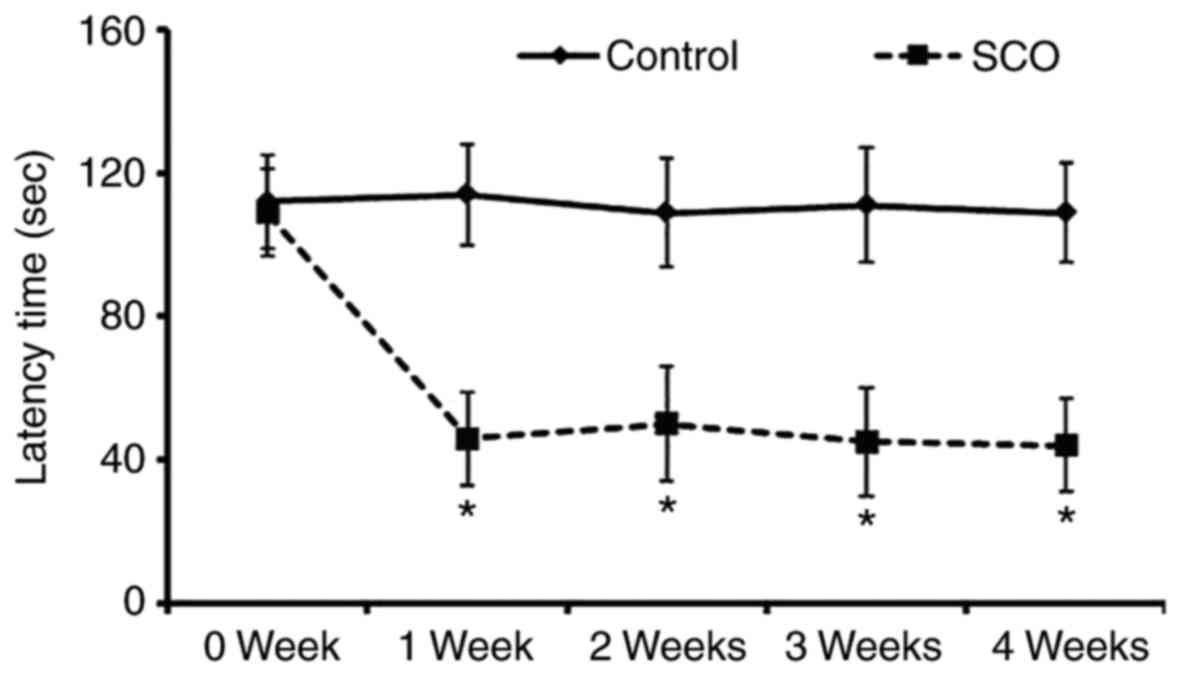
Avoidance memory defect
To investigate the SCO-induced short-term memory
impairment, exploratory preference was measured by passive
avoidance test which was reflected by decrease of the latency to
enter the dark compartment (Fig.
1). There was no significant difference in the latency time
until 4 weeks in the vehicle-treated mice (control mice). However,
the latency time of the SCO-treated mice was significantly reduced
1 week after SCO treatment compared to that in the vehicle-treated
mice, and the latency time was maintained until 4 weeks after SCO
treatment.
Spatial memory defect
Hippocampal dependent spatial learning and memory
defect induced by SCO was evaluated using the Morris water maze
test. As shown in Table I, the
vehicle-treated mice readily learned and memorized the location of
the submerged hidden platform. However, in the SCO-treated mice,
the escape latency 1 week after SCO treatment was significantly
longer than that in the vehicle-treated mice, and the increased
escape latency was maintained until 4 weeks after SCO
treatment.
 | Table I.Effect of chronic scopolamine
treatment on the Morris water maze test. |
Table I.
Effect of chronic scopolamine
treatment on the Morris water maze test.
|
| Training escape
latency (sec) | Test escape latency
(sec) |
|---|
|
|
|
|
|---|
| Group | Day 1 | Day 2 | Day 3 | Day 4 |
|---|
| Control |
| 0
weeks |
68.2±5.5 |
60.4±4.2 |
49.8±5.8 |
43.6±7.9 |
| 1
week |
66.3±3.9 |
59.2±3.2 |
46.3±4.2 |
41.2±6.2 |
| 2
weeks |
69.8±4.7 |
61.6±3.9 |
45.2±5.5 |
39.8±7.1 |
| 3
weeks |
64.1±4.6 |
57.7±8.1 |
42.6±3.1 |
35.5±8.0 |
| 4
weeks |
67.5±5.2 |
54.4±3.9 |
43.5±6.2 |
33.4±9.1 |
| SCO |
| 0
weeks |
65.7±3.5 |
55.7±4.1 |
44.6±5.5 |
40.1±8.2 |
| 1
week |
77.4±5.7a |
78.5±7.3a |
76.5±8.2a |
79.3±9.0a |
| 2
weeks |
73.5±6.3a |
75.2±5.9a |
74.9±7.8a |
75.7±10.3a |
| 3
weeks |
79.6±7.2a |
77.4±6.2a |
75.7±6.8a |
77.9±7.4a |
| 4
weeks |
80.2±9.3a |
81.8±6.6a |
77.4±5.7a |
75.8±9.2a |
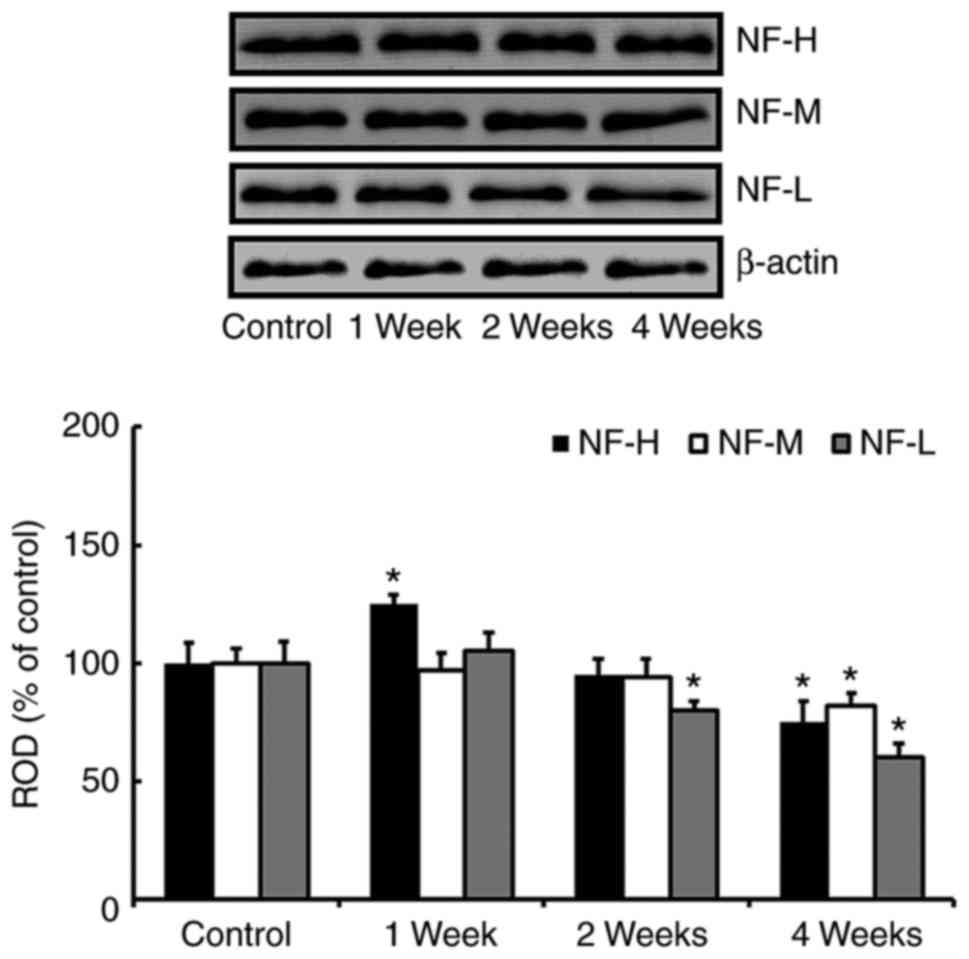
Changes in levels of NFs proteins
In the western blot study, we found that change
patterns in levels of NF-H, NF-M and NF-L proteins in the
hippocampus after SCO treatment were generally different according
kinds of the proteins (Fig. 2).
One week after SCO treatment, only NF-H level was significantly
increased compared with that of the control group. Two weeks after
SCO treatment, only NF-L level was significantly decreased compared
with that of the control group. Four weeks after SCO treatment, all
the NFs levels were significantly decreased compared with those of
the control group.
In our preliminary study, we found that there were
no significant differences in protein levels of NFs in the
hippocampus between 3 and 4 weeks SCO-treated groups. For
reference, we recently reported that significant difference in the
protein level of myelin basic protein, which is most abundant
structural protein in the myelin, was not found in the hippocampus
between 3 and 4 weeks SCO-treated groups (4).
Change in NF-H immunoreactivity
CA1 region: In the control group, NF-H
immunoreactivity was shown in the stratum pyramidale, and in cells
of stratum oriens and radiatum, which had NF-H immunoreactive
processes (Fig. 3Aa and Ab). In
the SCO-treated mice, NF-H-immunoreactive cells were increased in
the stratum oriens and radiatum 1 week after SCO treatment,
although the immunoreactivity in the stratum pyramidale was not
significantly changed compared with that in the control mice
(Fig. 3Bb). Two and 4 weeks after
SCO treatment, NF-H immunoreactive cells in the stratum oriens and
radiatum were not detected, although the immunoreactivity in the
stratum pyramidale was not significantly changed compared with that
in the control (Fig. 3Cb and
Db).
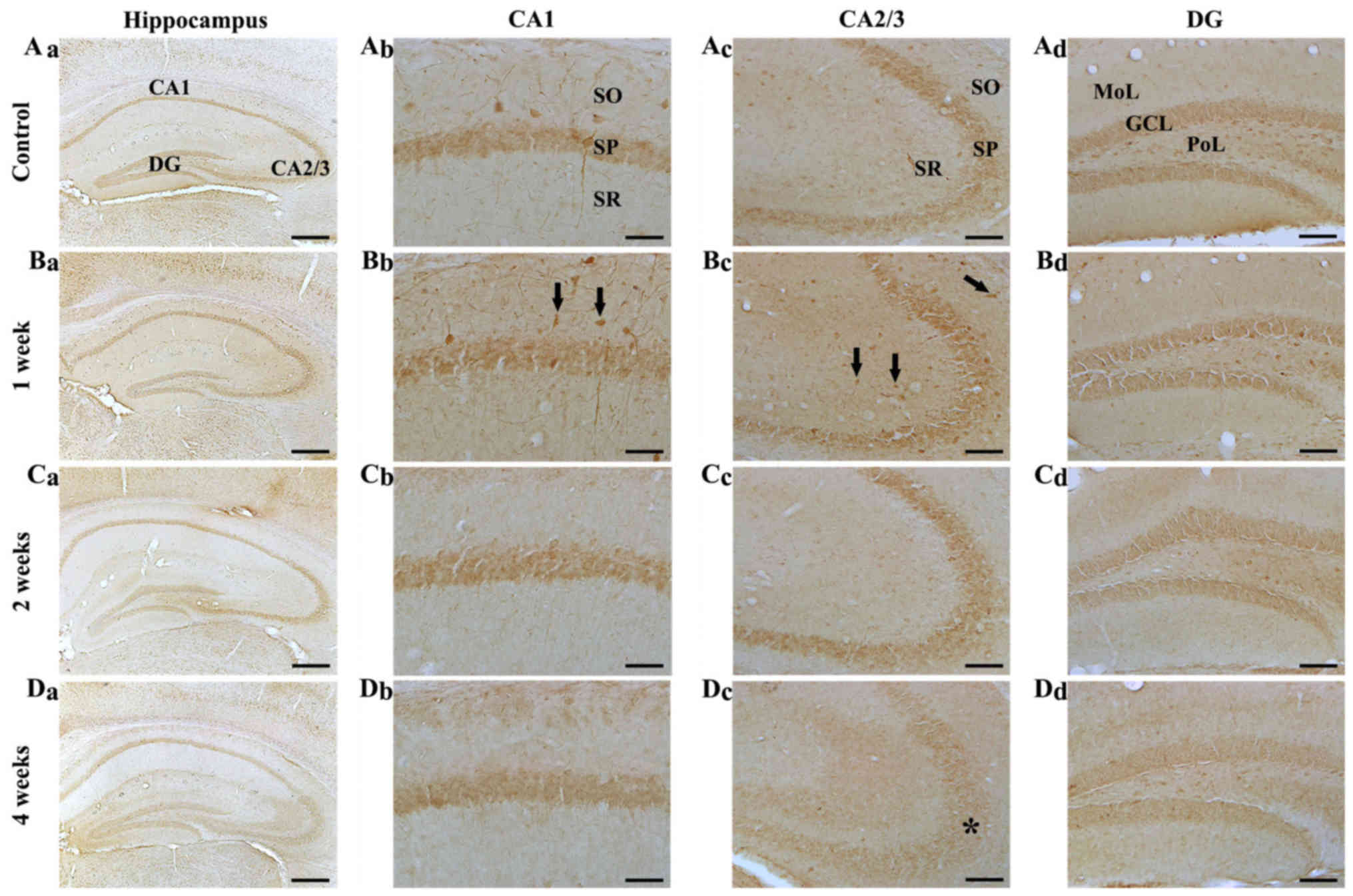 | Figure 3.NF-H immunohistochemistry in the
hippocampal subregions of the (A) control and SCO-treated mice
following (B) 1, (C) 2 and (D) 4 weeks. NF-H immunoreactive cells
(indicated by arrows) increased in the CA1-3 regions (Bb and Bc) 1
week following scopolamine treatment. A total of 2 and 4 weeks
following SCO treatment (C and D, a-d), NF-H immunoreactivity
decreased in all hippocampal subregions, in particular, in the
stratum pyramidale (indicated by SP and an asterisk) of the (Dc)
CA2/3 region 4 weeks following SCO treatment. Scale bars=800 µm
(Aa-Da), 50 µm (Ab-Db) and 100 µm (Ac-Dc and Ad-Dd). CA, cornus
ammonis; DG, dentate gyrus; GCL, granule cell layer; Mol, molecular
layer; PoL, polymorphic layer; SO, stratum oriens; SR, stratum
radiatum; SP, stratum pyramidale; NF-H, neurofilament-200 kDa; SCO,
scopolamine. |
CA2/3 region: In the control group, the distribution
pattern of NF-H immunoreactivity in the CA2/3 region was similar to
that in the CA1 region of the control (Fig. 3Aa and Ac). In the SCO-treated mice,
NF-H immunoreactive cells and NF-H immunoreactivity were
significantly increased 1 week after SCO treatment to the control
(Fig. 3Bc). Two and 4 weeks after
SCO treatment, NF-H-immunoreactive cells were rarely detected in
the stratum oriens and radiatum, and, 4 weeks after SCO treatment,
NF-H immunoreactivity in the stratum pyramidale was more
significantly decreased compared with that in the 2 weeks
SCO-treated group (Fig. 3Cc and
Dc).
Dentate gyrus: In the control group, NF-H
immunoreactivity was mainly found in the granular cell layer and
many cells of the polymorphic layer (Fig. 3Aa and Ad). In the SCO-treated mice,
NF-H immunoreactivity in the granular cell layer and many cells of
the polymorphic layer was gradually decreased with time after SCO
treatment (Fig. 3Bd-Dd).
In the preliminary study, we did not find any
difference in NF-H immunoreactivity in the hippocampus between 3
and 4 weeks SCO-treated groups.
Change in NF-M immunoreactivity
CA1 region: Strong NF-M immunoreactivity was
observed in fibers in the stratum oriens in the control mice, and
NF-M-immunoreactive fibers were scattered in the other layers
(Fig. 4Aa and Ab). In the
SCO-treated mice, NF-M immunoreactivity in the CA1 region was
gradually reduced after SCO treatment and significantly decreased 4
weeks after SCO treatment (Fig.
4Bb-Db).
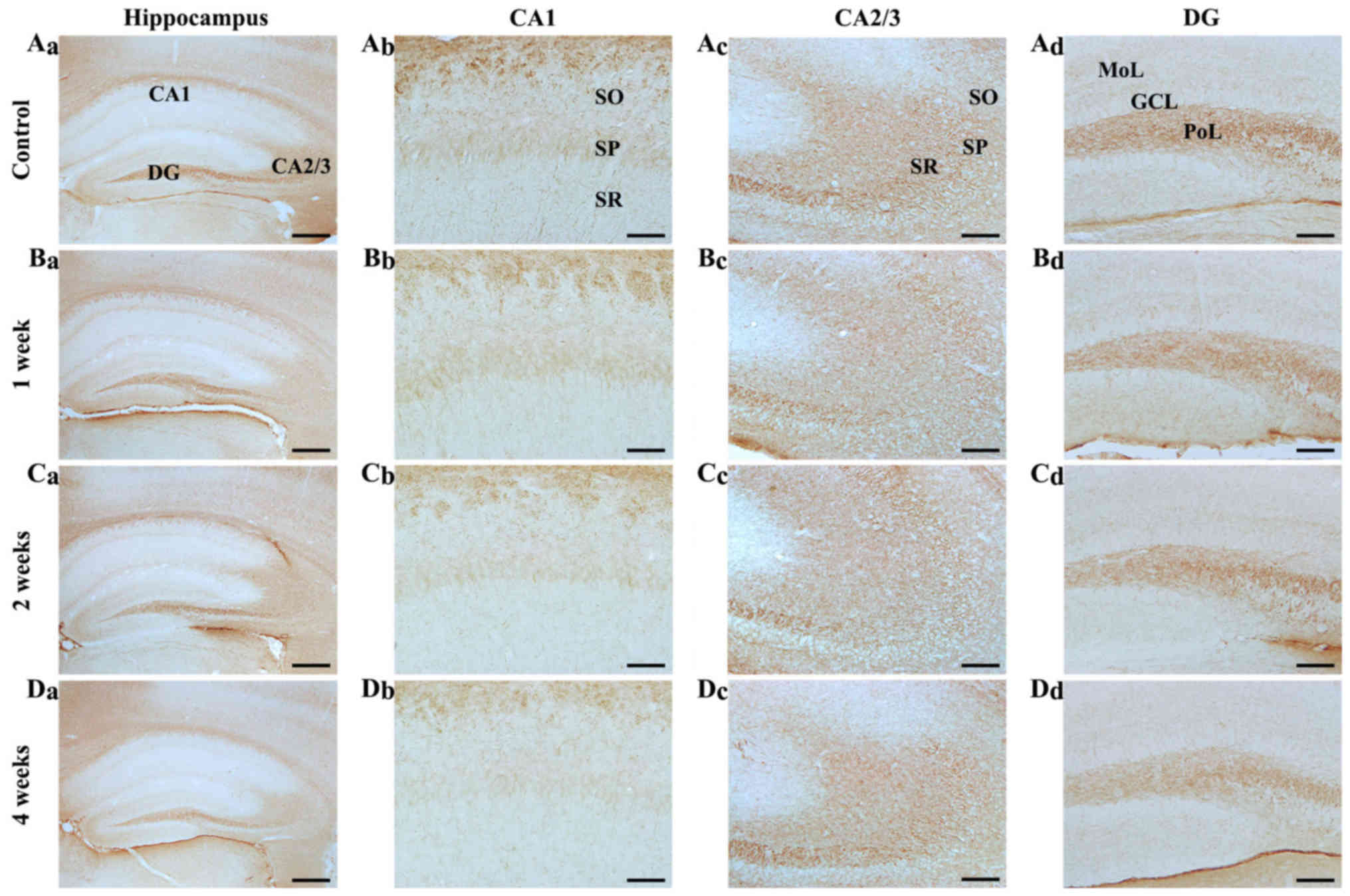 | Figure 4.NF-M immunohistochemistry in the
hippocampal subregions of the (A) control and SCO-treated mice
following (B) 1, (C) 2 and (D) 4 weeks. NF-M immunoreactivity in
all hippocampal subregions of the SCO-treated mice were not
markedly altered until 2 weeks following SCO treatment; however,
NF-M immunoreactivity was decreased (Da-d) 4 weeks following SCO
treatment. Scale bar=800 µm (Aa-Da), 50 µm (Ab-Db) and 100 µm
(Ac-Dc and Ad-Dd). CA, cornus ammonis; DG, dentate gyrus; GCL,
granule cell layer; Mol, molecular layer; PoL, polymorphic layer;
SO, stratum oriens; SP, stratum pyramidale; SR, stratum radiatum;
NF-M, neurofilament-165 kDa; SCO, scopolamine. |
CA2/3 regions: In the control mice, NF-M
immunoreactivity was strongly detected in all the layers (Fig. 4Aa and Ac). In the SCO-treated mice,
the distribution pattern of NF-M immunoreactivity was not
significantly changed in all the layers until 4 weeks after SCO
treatment (Fig. 4Bc-Dc).
Dentate gyrus: In the control mice, NF-M
immunoreactivity was mainly detected in the polymorphic layer
(Fig. 4Aa and Ad). In the
SCO-treated mice, NF-M immunoreactivity was slightly decreased 1
and 2 weeks after SCO treatment and significantly reduced 4 weeks
after SCO treatment compared with that in the control (Fig. 4Bd-Dd).
In the preliminary study, there was no significant
difference in NF-M immunoreactivity in the hippocampus between 3
and 4 weeks SCO-treated groups.
Change in NF-L immunoreactivity
CA1 region: NF-L immunoreactivity in the control
group was weakly detected in all the layers (Fig. 5Aa and Ab). In the SCO-treated
group, NF-L immunoreactivity was not significantly changed at any
time after SCO treatment (Fig.
5Bb-Db).
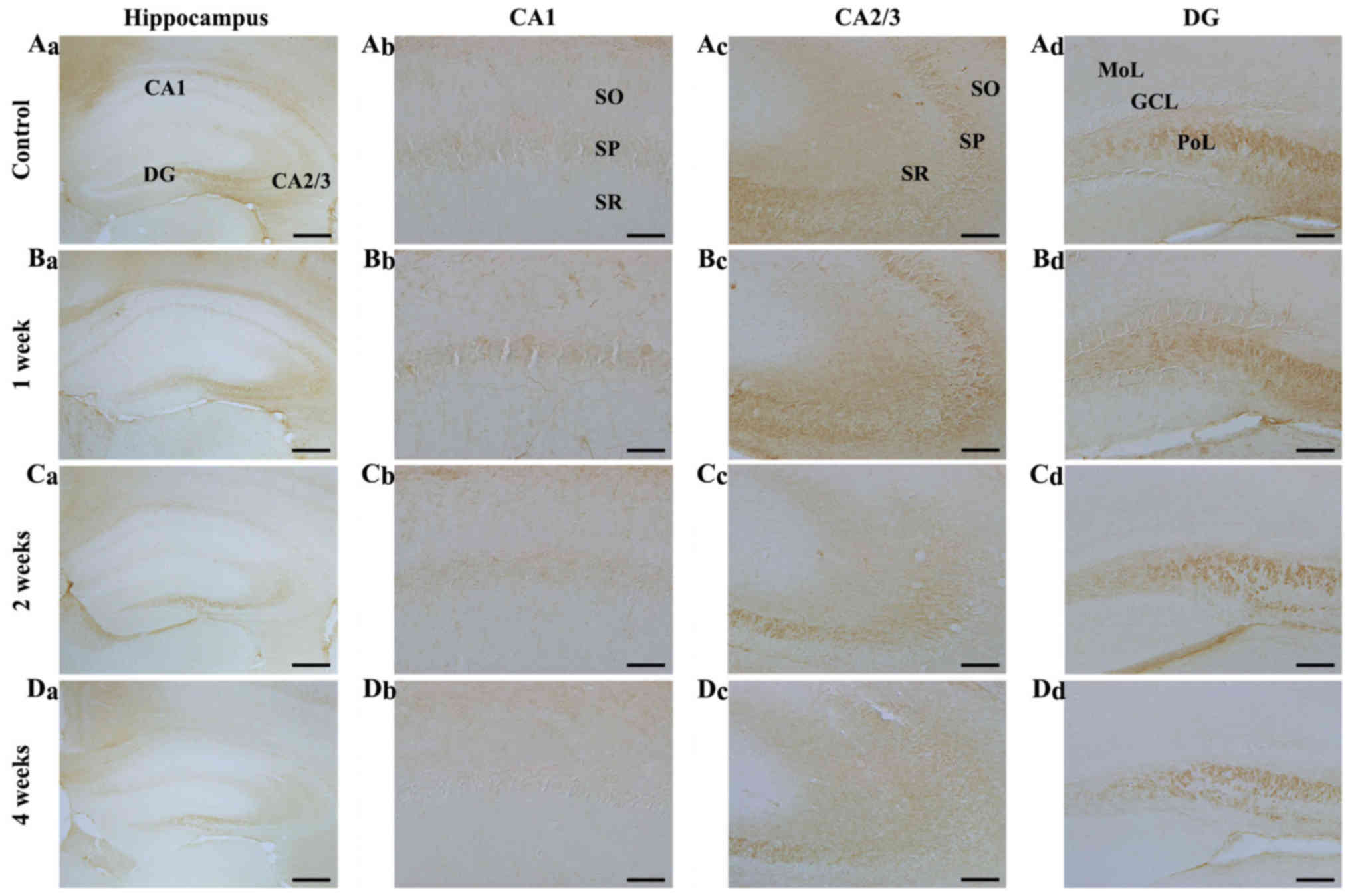 | Figure 5.NF-L immunohistochemistry in the
hippocampal subregions of the (A) control and SCO-treated mice
following (B) 1, (C) 2 and (D) 4 weeks. NF-L immunoreactivity was
increased in the (Bc) SP 1 week following SCO treatment; however,
NF-L immunoreactivity decreased in all hippocampal subregions, (C
and D) 2 and 4 weeks following SCO treatment. Scale bar=800 µm
(Aa-Da), 50 µm (Ab-Db) and 100 µm (Ac-Dc and Ad-Dd). CA, cornus
ammonis; DG, dentate gyrus; GCL, granule cell layer; Mol, molecular
layer; PoL, polymorphic layer; SO, stratum oriens; SP, stratum
pyramidale; SR, stratum radiatum; NF-L, neurofilament-68 kDa; SCO,
scopolamine. |
CA2/3 region: In the control mice, NF-L
immunoreactivity in the CA2/3 region was stronger than that in the
CA1 region (Fig. 5Aa and Ac). In
the SCO-treated mice, NF-L immunoreactivity was significantly
increased in the stratum pyramidale 1 week after SCO treatment
compared with that in the control (Fig. 5Bc), thereafter, NF-L
immunoreactivity was gradually decreased in all the layers and very
low 4 weeks after SCO treatment (Fig.
5Cc and Dc).
Dentate gyrus: In the control mice, strong NF-L
immunoreactivity was found in the polymorphic layer (Fig. 5Aa and Ad). In the SCO-treated mice,
NF-L immunoreactivity was not changed 1 week after SCO treatment
compared to that in the control (Fig.
5Bd). However, NF-L immunoreactivity significantly reduced 2
weeks after SCO treatment and more significantly decreased 4 weeks
after SCO treatment (Fig. 5Cd and
Dd).
In our preliminary study, no significant difference
in NF-L immunoreactivity was observed in the hippocampus between 3
and 4 weeks SCO-treated groups.
Discussion
Memory loss followed by cognitive decline is found
to be a common feature of various neurological disorders, which may
be due to an impairment in the neuronal circuit activity in the
hippocampus (13). Rodent models
of cognitive deficits have been established using SCO-induced
amnesia (34). In particular,
SCO-induced cognitive impairment in animals is similar to that
observed in the dysfunction of AD (6,7,35).
Therefore, the SCO-induced memory impairment model has served as a
useful tool with which to investigate learning and memory
processes.
SCO-induced cognitive deficits have been
demonstrated using various behavioral tasks (3,36).
In the present study, the SCO-induced cognitive impairment was
assessed by the passive avoidance and Morris water maze tests. The
passive avoidance test is a measurement of cognitive memory based
on avoidance of a fear-inducing context. Rodents naturally prefer
dark compartments; however, receiving an electric shock in a dark
compartment causes a conflict with this tendency (37). Furthermore, the Morris water maze
test is designed to evaluate spatial learning ability and long-term
spatial memory (38,39). We found that chronic systemic SCO
treatment reduced the step-through latency in the passive avoidance
test, and caused the mice to take a longer time to find the
platform than the control group in the Morris water-maze task.
Therefore, these findings indicate that chronic systemic treatment
with SCO can induce short-term memory deficits, as well as spatial
learning and memory impairment.
NFs are key intermediate filaments in neurons and
major components of the axonal cytoskeleton (27,40).
Alterations of NFs have been observed in neurodegenerative diseases
(25–27,41),
and disruption of their expression shows important consequences in
neuronal function (26,42). In addition, early alterations of
NFs represent the development of diffuse axonal injury, and
precedes microtubule fracture and depolymerization (43).
A long time ago, Leifer and Kowall reported that
selective cellular vulnerability in human cerebral ischemia was
related to the reduction of hippocampal pyramidal cells via
staining the pyramidal cells with SMI-32, a marker for
nonphosphorylated NFs (44).
Posmantur et al (45)
reported that progressive alterations of NF-L immunoreactivity
occurred in dendrites at 15 min after a traumatic brain injury and
that a definite loss of dendritic NFs was found 3 to 24 h after the
injury. Moreover, it has been reported that NFs are disrupted in
the hippocampus under depressive states, and the disruption is
associated with a dysfunction of hippocampal plasticity in
depressive states (46,47). These papers show that alterations
in NFs may serve as a trigger for further secondary damage to
axons, representing a key insight into a temporal aspect of
cytoskeletal degeneration (43).
In the present study, we examined chronological
changes in NFs in the mouse hippocampus after chronic systemic
treatment with SCO, using the western blot analysis and
immunohistochemistry for NFs; we found that NF-H immunoreactivity
and levels were significantly increased 1 week after SCO treatment.
Thereafter, they significantly decreased with time; the NF-M
immunoreactivity and levels gradually decreased and were
significantly decreased 4 weeks after the SCO treatment; and NF-L
immunoreactivity and levels were significantly decreased from 2
weeks after the SCO treatment. Furthermore, alteration in each
immunoreactivity was distinctively different according to the
hippocampal subregions and their layers as well as according to the
time after SCO treatment. Based on our and the above-mentioned
findings, it is obvious that dysfunction in axonal transport might
underlie alterations of NFs.
In brief, our present results show that long-term
treatment with SCO induced cognitive deficits from 1 week after SCO
treatment, although NFs levels and immunoreactivities were
significantly and diversely altered in the mouse hippocampus
according to the hippocampal subregions and the time-course after
the SCO treatment. Therefore, more research needs to be conducted
about what occurs in cognitive deficits according to an alteration
in each NF expression in the hippocampus following chronic systemic
treatment with SCO.
Acknowledgements
The authors would like to thank Mr. Seung Uk Lee for
his technical help in this study. This study was supported by
Bio-Synergy Research Project (NRF-2015M3A9C4076322) of the Ministry
of Science, ICT and Future Planning through the National Research
Foundation, by the Bio and Medical Technology Development Program
of the NRF funded by the Korean government, MSIP
(NRF-2015M3A9B6066835), and by a Priority Research Centers Program
grant (NRF-2009-0093812) through the National Research Foundation
of Korea funded by the Ministry of Science, ICT and Future
Planning.
References
|
1
|
Bartus RT, Dean RL III, Beer B and Lippa
AS: The cholinergic hypothesis of geriatric memory dysfunction.
Science. 217:408–414. 1982. View Article : Google Scholar
|
|
2
|
Zhou MM, Xue Y, Sun SH, Wen M, Li ZJ, Xu
J, Wang JF, Yanagita T, Wang YM and Xue CH: Effects of different
fatty acids composition of phosphatidylcholine on brain function of
dementia mice induced by scopolamine. Lipids Health Dis.
15:1352016. View Article : Google Scholar :
|
|
3
|
Lee B, Sur B, Shim J, Hahm DH and Lee H:
Acupuncture stimulation improves scopolamine-induced cognitive
impairment via activation of cholinergic system and regulation of
BDNF and CREB expressions in rats. BMC Complement Altern Med.
14:3382014. View Article : Google Scholar :
|
|
4
|
Park JH, Choi HY, Cho JH, Kim IH, Lee TK,
Lee JC, Won MH, Chen BH, Shin BN, Ahn JH, et al: Effects of chronic
scopolamine treatment on cognitive impairments and myelin basic
protein expression in the mouse hippocampus. J Mol Neurosci.
59:579–589. 2016. View Article : Google Scholar
|
|
5
|
Yoo DY, Choi JH, Kim W, Nam SM, Jung HY,
Kim JH, Won MH, Yoon YS and Hwang IK: Effects of luteolin on
spatial memory, cell proliferation and neuroblast differentiation
in the hippocampal dentate gyrus in a scopolamine-induced amnesia
model. Neurol Res. 35:813–820. 2013. View Article : Google Scholar
|
|
6
|
Ahmed T and Gilani AH: Inhibitory effect
of curcuminoids on acetylcholinesterase activity and attenuation of
scopolamine-induced amnesia may explain medicinal use of turmeric
in Alzheimer's disease. Pharmacol Biochem Behav. 91:554–559. 2009.
View Article : Google Scholar
|
|
7
|
Klinkenberg I and Blokland A: The validity
of scopolamine as a pharmacological model for cognitive impairment:
A review of animal behavioral studies. Neurosci Biobehav Rev.
34:1307–1350. 2010. View Article : Google Scholar
|
|
8
|
Lian W, Fang J, Xu L, Zhou W, Kang, Xiong
W, Jia H, Liu AL and Du GH: DL0410 ameliorates memory and cognitive
impairments induced by scopolamine via increasing cholinergic
neurotransmission in mice. Molecules. 22:pii: E410. 2017.
View Article : Google Scholar
|
|
9
|
Hami J, Kheradmand H and Haghir H: Sex
differences and laterality of insulin receptor distribution in
developing rat hippocampus: An immunohistochemical study. J Mol
Neurosci. 54:100–108. 2014. View Article : Google Scholar
|
|
10
|
Huang W, Cao J, Liu X, Meng F, Li M, Chen
B and Zhang J: AMPK plays a dual role in regulation of CREB/BDNF
pathway in mouse primary hippocampal cells. J Mol Neurosci.
56:782–788. 2015. View Article : Google Scholar
|
|
11
|
Kong Y, Bai PS, Sun H and Nan KJ:
Expression of the newly identified gene CAC1 in the hippocampus of
Alzheimer's disease patients. J Mol Neurosci. 47:207–218. 2012.
View Article : Google Scholar
|
|
12
|
Zhao L, Sun C, Xiong L, Yang Y, Gao Y,
Wang L, Zuo H, Xu X, Dong J, Zhou H and Peng R: MicroRNAs: Novel
mechanism involved in the pathogenesis of microwave exposure on
rats' hippocampus. J Mol Neurosci. 53:222–230. 2014. View Article : Google Scholar
|
|
13
|
Terry AV Jr, Callahan PM, Hall B and
Webster SJ: Alzheimer's disease and age-related memory decline
(preclinical). Pharmacol Biochem Behav. 99:190–210. 2011.
View Article : Google Scholar :
|
|
14
|
Gold PE: Acetylcholine modulation of
neural systems involved in learning and memory. Neurobiol Learn
Mem. 80:194–210. 2003. View Article : Google Scholar
|
|
15
|
Hasselmo ME: The role of acetylcholine in
learning and memory. Curr Opin Neurobiol. 16:710–715. 2006.
View Article : Google Scholar :
|
|
16
|
Lippa AS, Critchett DJ, Ehlert F, Yamamura
HI, Enna SJ and Bartus RT: Age-related alterations in
neurotransmitter receptors: An electrophysiological and biochemical
analysis. Neurobiol Aging. 2:3–8. 1981. View Article : Google Scholar
|
|
17
|
Vijayan VK: Cholinergic enzymes in the
cerebellum and the hippocampus of the senescent mouse. Exp
Gerontol. 12:7–11. 1977. View Article : Google Scholar
|
|
18
|
Nixon RA and Sihag RK: Neurofilament
phosphorylation: A new look at regulation and function. Trends
Neurosci. 14:501–506. 1991. View Article : Google Scholar
|
|
19
|
Serrano-Pozo A, Frosch MP, Masliah E and
Hyman BT: Neuropathological alterations in Alzheimer disease. Cold
Spring Harb Perspect Med. 1:a0061892011. View Article : Google Scholar :
|
|
20
|
Dong DL, Xu ZS, Chevrier MR, Cotter RJ,
Cleveland DW and Hart GW: Glycosylation of mammalian
neurofilaments. Localization of multiple O-linked
N-acetylglucosamine moieties on neurofilament polypeptides L and M.
J Biol Chem. 268:16679–16687. 1993.
|
|
21
|
Sánchez I, Hassinger L, Sihag RK,
Cleveland DW, Mohan P and Nixon RA: Local control of neurofilament
accumulation during radial growth of myelinating axons in vivo.
Selective role of site-specific phosphorylation. J Cell Biol.
151:1013–1024. 2000. View Article : Google Scholar :
|
|
22
|
Yabe JT, Chan WK, Chylinski TM, Lee S,
Pimenta AF and Shea TB: The predominant form in which neurofilament
subunits undergo axonal transport varies during axonal initiation,
elongation, and maturation. Cell Motil Cytoskeleton. 48:61–83.
2001. View Article : Google Scholar
|
|
23
|
Hayes RL, Yang K, Whitson JS and
Postmantur R: Cytoskeletal derangements following central nervous
system injury: Modulation by neurotrophic gene transfection. J
Neurotrauma. 12:933–941. 1995. View Article : Google Scholar
|
|
24
|
Tu PH, Elder G, Lazzarini RA, Nelson D,
Trojanowski JQ and Lee VM: Overexpression of the human NFM subunit
in transgenic mice modifies the level of endogenous NFL and the
phosphorylation state of NFH subunits. J Cell Biol. 129:1629–1640.
1995. View Article : Google Scholar :
|
|
25
|
Al-Chalabi A and Miller CC: Neurofilaments
and neurological disease. Bioessays. 25:346–355. 2003. View Article : Google Scholar
|
|
26
|
Julien JP: Neurofilament functions in
health and disease. Curr Opin Neurobiol. 9:554–560. 1999.
View Article : Google Scholar
|
|
27
|
Perrot R, Berges R, Bocquet A and Eyer J:
Review of the multiple aspects of neurofilament functions, and
their possible contribution to neurodegeneration. Mol Neurobiol.
38:27–65. 2008. View Article : Google Scholar
|
|
28
|
Vickers JC, Kirkcaldie MT, Phipps A and
King AE: Alterations in neurofilaments and the transformation of
the cytoskeleton in axons may provide insight into the aberrant
neuronal changes of Alzheimer's disease. Brain Res Bull.
126:324–333. 2016. View Article : Google Scholar
|
|
29
|
Siedler DG, Chuah MI, Kirkcaldie MT,
Vickers JC and King AE: Diffuse axonal injury in brain trauma:
Insights from alterations in neurofilaments. Front Cell Neurosci.
8:4292014. View Article : Google Scholar :
|
|
30
|
Wang X, Wang ZH, Wu YY, Tang H, Tan L,
Wang X, Gao XY, Xiong YS, Liu D, Wang JZ and Zhu LQ: Melatonin
attenuates scopolamine-induced memory/synaptic disorder by rescuing
EPACs/miR-124/Egr1 pathway. Mol Neurobiol. 47:373–381. 2013.
View Article : Google Scholar
|
|
31
|
Yan BC, Park JH, Chen BH, Cho JH, Kim IH,
Ahn JH, Lee JC, Hwang IK, Cho JH, Lee YL, et al: Long-term
administration of scopolamine interferes with nerve cell
proliferation, differentiation and migration in adult mouse
hippocampal dentate gyrus, but it does not induce cell death.
Neural Regen Res. 9:1731–1739. 2014. View Article : Google Scholar :
|
|
32
|
Lee JC, Park JH, Ahn JH, Kim IH, Cho JH,
Choi JH, Yoo KY, Lee CH, Hwang IK, Cho JH, et al: NNew GABAergic
neurogenesis in the hippocampal CA1 region of a gerbil model of
long-term survival after transient cerebral ischemic injury. Brain
Pathol. 26:581–592. 2016. View Article : Google Scholar
|
|
33
|
Park JH, Lee CH, Yoo KY, Choi JH, Hwang
IK, Lee JY, Kang IJ and Won MH: FoxO3a immunoreactivity is markedly
decreased in the dentate gyrus, not the hippocampus proper, of the
aged gerbil. Exp Gerontol. 46:836–840. 2011. View Article : Google Scholar
|
|
34
|
Blokland A: Scopolamine-induced deficits
in cognitive performance: A review of animal studies. Scopolamine
Rev. 1:1–76. 2005.
|
|
35
|
Ebert U and Kirch W: Scopolamine model of
dementia: Electroencephalogram findings and cognitive performance.
Eur J Clin Invest. 28:944–949. 1998. View Article : Google Scholar
|
|
36
|
Shi Z, Chen L, Li S, Chen S, Sun X, Sun L,
Li Y, Zeng J, He Y and Liu X: Chronic scopolamine-injection-induced
cognitive deficit on reward-directed instrumental learning in rat
is associated with CREB signaling activity in the cerebral cortex
and dorsal hippocampus. Psychopharmacology (Berl). 230:245–260.
2013. View Article : Google Scholar
|
|
37
|
Romanski LM and LeDoux JE: Information
cascade from primary auditory cortex to the amygdala:
Corticocortical and corticoamygdaloid projections of temporal
cortex in the rat. Cereb Cortex. 3:515–532. 1993. View Article : Google Scholar
|
|
38
|
D'Hooge R and De Deyn PP: Applications of
the Morris water maze in the study of learning and memory. Brain
Res Brain Res Rev. 36:60–90. 2001. View Article : Google Scholar
|
|
39
|
Morris R: Developments of a water-maze
procedure for studying spatial learning in the rat. J Neurosci
Methods. 11:47–60. 1984. View Article : Google Scholar
|
|
40
|
Fletcher DA and Mullins RD: Cell mechanics
and the cytoskeleton. Nature. 463:485–492. 2010. View Article : Google Scholar :
|
|
41
|
Norgren N, Rosengren L and Stigbrand T:
Elevated neurofilament levels in neurological diseases. Brain Res.
987:25–31. 2003. View Article : Google Scholar
|
|
42
|
Gotow T: Neurofilaments in health and
disease. Med Electron Microsc. 33:173–199. 2000. View Article : Google Scholar
|
|
43
|
Fournier AJ, Rajbhandari L, Shrestha S,
Venkatesan A and Ramesh KT: In vitro and in situ visualization of
cytoskeletal deformation under load: traumatic axonal injury. FASEB
J. 28:5277–5287. 2014. View Article : Google Scholar
|
|
44
|
Leifer D and Kowall NW:
Immunohistochemical patterns of selective cellular vulnerability in
human cerebral ischemia. J Neurol Sci. 119:217–228. 1993.
View Article : Google Scholar
|
|
45
|
Posmantur RM, Newcomb JK, Kampfl A and
Hayes RL: Light and confocal microscopic studies of evolutionary
changes in neurofilament proteins following cortical impact injury
in the rat. Exp Neurol. 161:15–26. 2000. View Article : Google Scholar
|
|
46
|
Reinés A, Cereseto M, Ferrero A, Bonavita
C and Wikinski S: Neuronal cytoskeletal alterations in an
experimental model of depression. Neuroscience. 129:529–538. 2004.
View Article : Google Scholar
|
|
47
|
Sanna MD, Ghelardini C and Galeotti N:
Effect of amitriptyline treatment on neurofilament-H protein in an
experimental model of depression. Brain Res Bull. 128:1–6. 2017.
View Article : Google Scholar
|