Introduction
Adipose tissue, as a highly active endocrine organ,
produces and releases a range of bioactive substances, such as
adipokines, inflammatory factors and hormones (1,2).
Among these, adiponectin (APN) has been reported as a major
anti-inflammatory mediator inhibiting the development of
cardiovascular disease (3,4). There is enough evidence indicating
that adipocyte-derived APN can exert protective effects in
different animal models such as hypertension and obesity (5–7).
However, APN attenuates cardiac inflammation and remodeling in the
classical angiotensin II (Ang II)-induced hypertension model
(8). However, the potential
protective role of APN on the Ang II-induced vascular remodeling
has not been fully elucidated.
Adiponectin, as a fat-derived hormone, fulfills a
critical role as an important messenger between adipocytes and
other organs through the adiponectin receptor 1 and/or receptor 2
(AdipoR1/R2). These receptors are ubiquitously expressed in the
adipose tissue, skeletal muscle and cardiovascular system (9,10).
Previous studies have demonstrated that APN attenuates vascular
remodeling through inhibiting vascular smooth muscle cell (VSMC)
proliferation and phenotype trans differentiation (11,12).
However, little is known about the endogenous APN and its receptor
expression and function in the vasculatures of the Ang II-induced
hypertensive model. In the present study, APN and AdipoR expression
in vascular cells was demonstrated and the effects of APN on VSMCs
migration were investigated. The distribution of these molecules in
the vasculature of Ang-II induced hypertension were detected in
vivo, and whether the APN receptor agonist AdipoRon could
attenuate hypertension-associated vascular injury was
determined.
Materials and methods
Cell culture
Vascular cells, such as human umbilical vein
endothelial cells (HUVECs; American Type Culture Collection,
Manassas, VA, USA) and VSMCs (PromoCell GmbH, Heidelberg, Germany),
were cultured in high glucose Dulbecco's modified Eagles medium
(DMEM) with 10% fetal bovine serum (both from Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) as previously described
(13). Cells at 4–6 passages were
used in subsequent experiments. 3T3-L1 adipocytes were
differentiated from 3T3-L1 fibroblasts (American Type Culture
Collection) as previously described (14). Briefly, 3T3-L1 fibroblasts were
cultured to reach confluency in a 12-well plate and were
differentiated into adipocytes in the specific differentiation
medium, consisting of DMEM supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.) and
3-isobutyl-1-methylxanthine (5 µM), dexamethasone (0.25 µM) (both
from Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and insulin (5
µg/ml; Gibco; Thermo Fisher Scientific, Inc.) for 2 days, followed
by treatment with DMEM supplemented with 10% FBS and insulin (5
µg/ml).
Animal models
Male, 8–12 week-old C57BL/6J background mice were
used in the present study. To induce hypertension, mice (n=5) were
subcutaneously infused with Ang II (Sigma-Aldrich; Merck KGaA) at a
1,500 ng/kg/min dose for 14 days with ALZET mini osmotic pumps
(DURECT Corporation, Cupertino, CA, USA), and in the sham group
mice were treated with saline (n=5). Blood pressure was measured
non-invasively every day by the tail-cuff method (Softron BP-98A;
Softron Co., Ltd., Tokyo, Japan). To determine the role of APN
receptor in hypertensive vascular injury, Ang II-infused mice
received APN receptor agonist AdipoRon (n=5, 30 mg/kg/day; Selleck
Chemicals, Houston, TX, USA) via a gavage tube. The experiments
were performed in adherence with the National Institutes of Health
guidelines on the Use of Laboratory Animals and were approved by
the Animal Care and Use Committee of Shanghai University of
Traditional Chinese Medicine (Shanghai, China).
Histological analysis
Thoracic aortas of Ang II-induced hypertensive mice
were obtained as previously described (15), and then were fixed in 4%
paraformaldehyde at 4°C for 24 h, embedded in paraffin, and 5 µm
sections were prepared for hematoxylin (15 min at room temperature)
and eosin (5 min at room temperature) (H&E) and picrosirius red
staining (30 min at room temperature) to assess the vascular
hypertrophy and fibrosis according to the manufacturers (Wuhan
Servicebio Technology Co., Ltd., Wuhan, China). Morphometric
analysis was performed using Image-Pro Plus 6.0 software to assess
vascular hypertrophy by measuring the medium and lumen area.
Fibrotic staining was expressed as a percentage of stained areas
(red) to the total areas examined. For immunofluorescence staining,
paraffin sections were first deparaffinized and rehydrated in
xylene (30 min), 100% ethanol (5 min), 90% ethanol (5 min) and 80%
ethanol (5 min). Following boiling in 10 mM sodium citrate buffer
for 10 min to unmask antigens, the slides were blocked in buffer
containing 5% normal goat serum (Invitrogen; Thermo Fisher
Scientific, Inc.) at room temperature for 1 h, incubated with
primary antibody at 4°C overnight, followed by incubation with the
fluorochrome-conjugated secondary antibody (1:200, for 1 h at room
temperature; Invitrogen; Thermo Fisher Scientific, Inc.). Primary
antibodies included perilin (PLIN1, 1:100, cat no. ab3526; Abcam,
Cambridge, UK), APN (1:200, cat no. AF1119; R&D Systems, Inc.,
Minneapolis, MN, USA), AdipoR1 (1:150, cat no. ab70362), AdipoR2
(1:150, cat no. ab77612) (both from Abcam), α smooth muscle actin
(α-SMA, 1:500, cat no. A5228; Sigma-Aldrich; Merch KGaA) and p-p38
(1:100, cat no. 4511; CST Biological Reagents Co., Ltd., Shanghai,
China). Images were captured by a laser scanner confocal microscopy
system (LSM 710; Zeiss GmbH, Jena, Germany).
Western blot analysis
Adipocytes, VSMCs and HUVECs were lysed in a
radioimmune precipitation assay buffer (0.1% sodium dodecyl
sulfate, 1.0% Triton X-100, 2 µg/ml aprotonin and 2 ng/ml
leupeptin) supplemented with protease inhibitor cocktail (Roche
Diagnostics, Basel, Switzerland) on ice for 10 min and sonicated.
Cell lysates were centrifuged (12,000 × g, 15 min at 4°C) and the
supernatant was preserved at 20°C for western blot analysis.
Protein concentrations were determined using the Bradford method.
Aliquots of protein (20 µg) were run on a 10% sodium dodecyl
sulfate-polyacrylamide gel followed by blotting onto polyvinylidene
fluoride membranes by wet transfer. Membranes were blocked in
blocking buffer (5% nonfat milk in TBS buffer with 0.1% Tween-20)
and were incubated with primary antibody at 4°C overnight. The
primary antibodies included APN (1:2,000, cat no. AF1119, R&D
Systems, Inc.), AdipoR1 (1:1,000, cat no. ab70362), AdipoR2
(1:1,000, cat no. ab77612) (both from Abcam), α-SMA (1:5,000, cat
no. A5228, Sigma-Aldrich; Merch KGaA), p-p38 (1:1,000, cat no.
4511), t-p38 (1:1,000, cat no. 9212) (both from CST Biological
Reagents Co., Ltd.) and β-actin (1:5,000, cat no. ab8226; Abcam).
The membranes were washed with TBS with Tween-20 for 3×10 min
followed by incubation with the horseradish peroxidase-conjugated
secondary antibodies [rabbit anti-mouse IgG (1:5,000, cat no.
ab6728); goat anti-rabbit IgG (1:5,000, cat no. ab6721); rabbit
anti-goat IgG (1:5,000, cat no. ab6741) (all from Abcam)] for 1 h
at room temperature. Protein bands were detected by
chemiluminescence (EMD Millipore, Billerica, MA, USA).
Migration assay
VSMCs migration was measured using a Transwell
chamber with a polycarbonate membrane (6.5 mm diameter and 8.0 µm
pore size; Corning Incorporated, Corning, NY, USA). Briefly,
serum-starved VSMCs were trypsinized and resuspended in DMEM with
0.5% bovine serum albumin (Sigma Sigma-Aldrich; Merch KGaA). Cell
suspension (100 µl, 1.0×106 cells/ml) was loaded in the
upper compartment of the chamber. DMEM with APN (10 µg/ml) were
added to lower compartment and incubated at 37°C. Following 2 h
incubation, Ang II (100 nM) was added. After 6 h, the cells
remaining on the upper surface of the chamber were removed with a
cotton swap, migrated cells on the lower surface of the chamber
were fixed in 4% paraformaldehyde at room temperature for 1 h and
stained with hematoxylin (15 min at room temperature) and eosin (5
min at room temperature) and three random microscopic fields
(magnification, ×200) per well were quantified by a fluorescence
microscope (DM3000; Leica, Wetzlar, Germany).
Statistical analysis
Statistical analysis was performed using SPSS 19.0
(SPSS; IBM Corp., Armonk, NY, USA). Data are expressed as the mean
± standard deviation. Comparisons of experimental groups were
analyzed by Student's t-test (between two groups) or one-way
analysis of variance, followed by the post-hoc Dunnett's test for
data with more than two groups (Levene's tests for equal variance
and Welch's and Brown-Forsythe's test for unequal variances).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Expression of APN and AdipoRs in
vascular cells
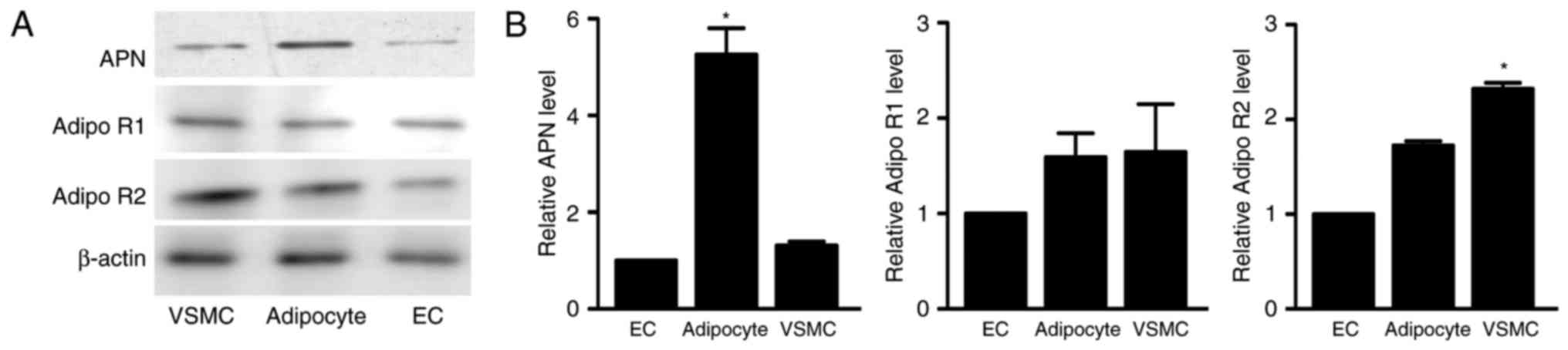
APN and AdipoR expression were detected in
adipocytes, VSMCs and endothelial cells (ECs) by western blot
analysis. As demonstrated in Fig.
1, APN expression was highest in the adipocytes, with the
expression of APN in the VSMCs and ECs less than in the adipocytes.
Additionally, AdipoR1 and AdipoR2 were expressed in the VSMCs, ECs
and adipocytes.
Expression of APN and AdipoR in the
vasculature
To detect APN and AdipoR expression in the blood
vessels, immunostaining of cross-sections of thoracic aorta in Sham
and Ang II-induced hypertensive mice was performed. Ang II infusion
resulted in a dramatic increased blood pressure including systolic
and diastolic blood pressure (Fig.
2A). In accordance with the in vitro results, APN was
mainly distributed in the perivascular adipocytes (Fig. 2B and C, perilipin-positive cells),
while AdipoR1 and AdipoR2 were expressed in both VSMCs (α smooth
muscle actin-positive cells) and adipocytes (Fig. 2D and E). Ang II-induced
hypertension resulted in decreased APN and AdipoR expression in the
vasculature.
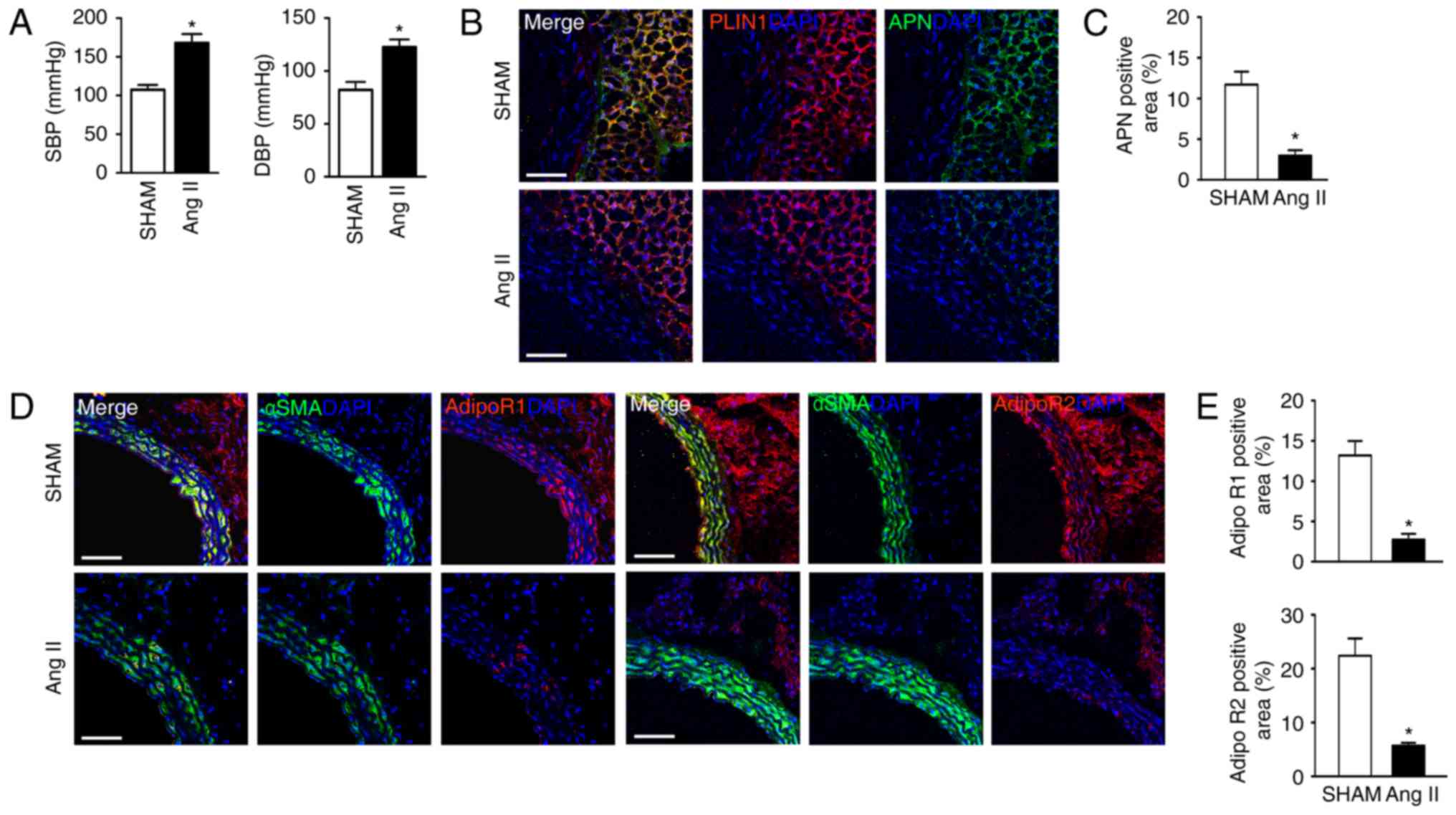 | Figure 2.Expression and distribution of APN and
its receptors in the vasculature. (A) Blood pressure measurements
of SHAM and Ang II-infused mice. Representative immunofluorescence
images of (B) APN+ and PLIN+ cells and (C)
quantification of their protein levels and representative
immunofluorescence images of (D) αSMA+,
AdipoR1+ and AdipoR2+ cells and (E)
quantification of their protein levels, in the perivascular
adipocytes and VSMCs from sham and Ang II mice. DAPI counterstained
for the nucleus. Scale bar, 100 µm. Data are expressed as the mean
± standard deviation (n=5). *P<0.05 vs. sham. APN, adiponectin;
Ang, angiotensin; VSMC, vascular smooth muscle cell; AdipoR,
adiponectin receptor; SBP, systolic blood pressure; DBP, diastolic
blood pressure; αSMA, α smooth muscle actin; PLIN, perilipin. |
Effects of Ang II on APN and AdipoR
expression
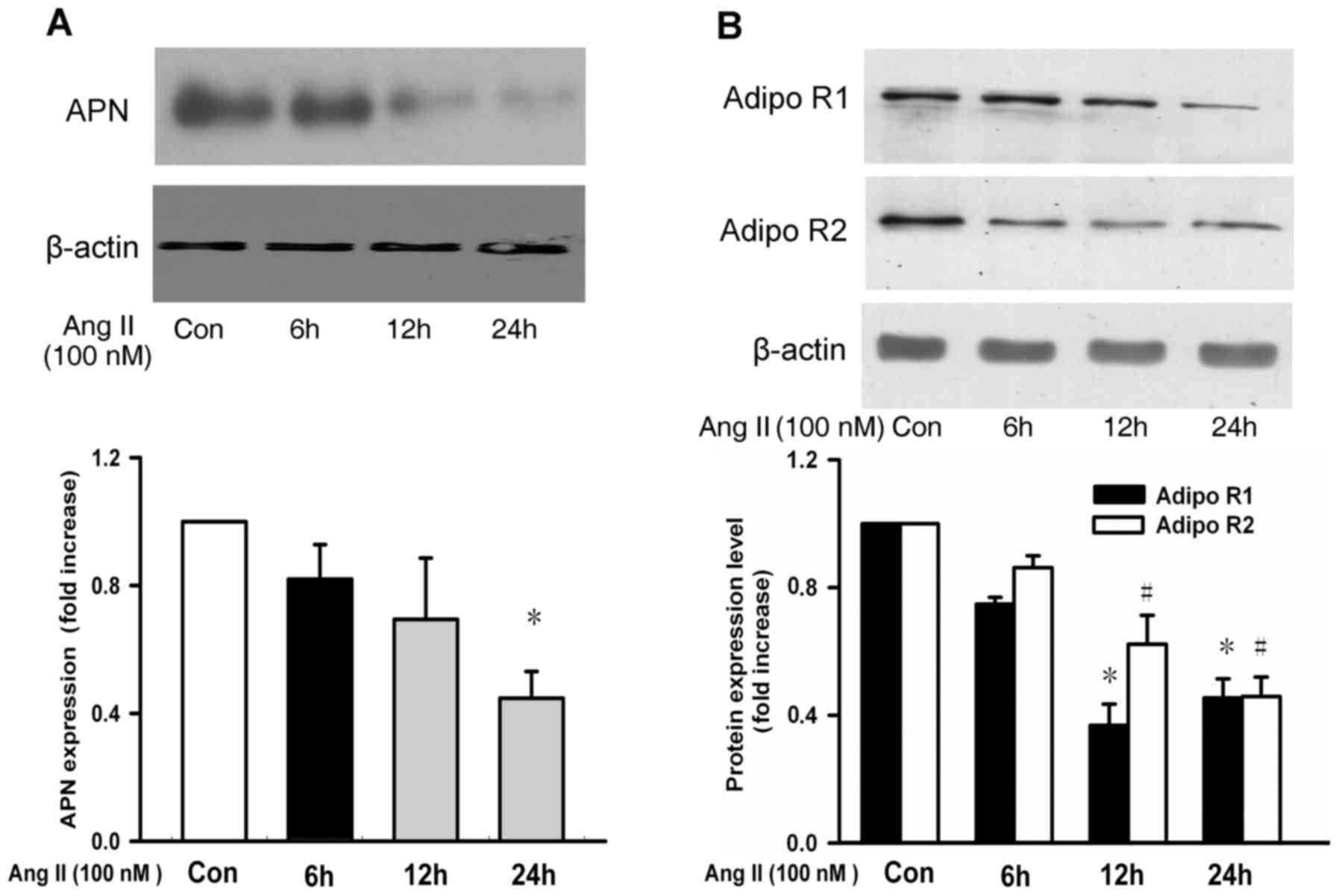
To detect the direct effects of Ang II on APN, APN
levels in the 3T3-L1 adipocytes pretreated with Ang II were
measured. The result demonstrated that APN expression was decreased
in a time-dependent manner following Ang II treatment (Fig. 3A). In addition, it was also
illustrated that AdipoR1 and AdipoR2 expression was decreased in a
time-dependent manner following Ang II treatment in the VSMCs
(Fig. 3B).
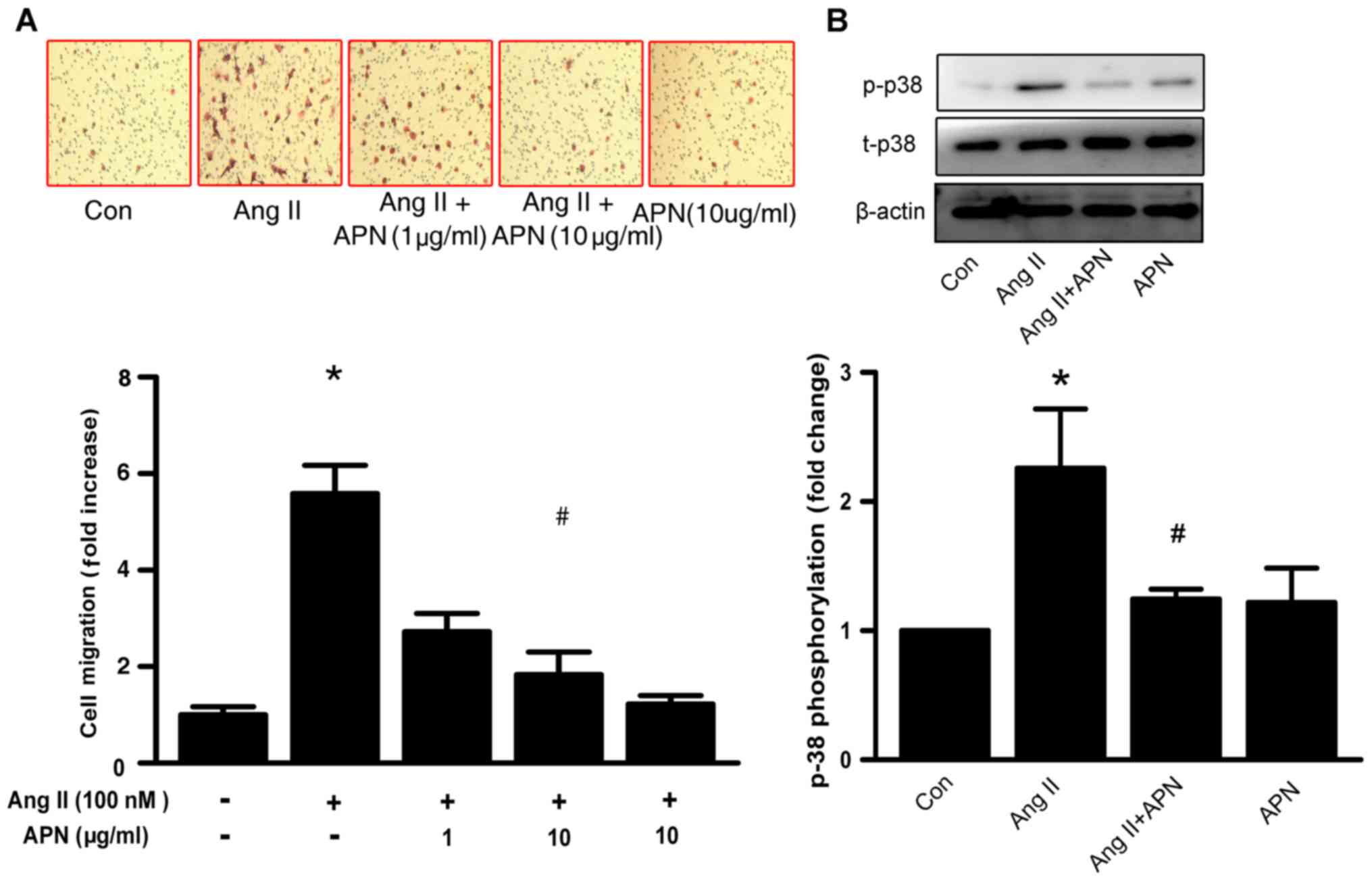
APN attenuates VSMCs migration
Vascular active substance-induced VSMCs migration
serves a pivotal role in hypertension-associated vascular
remodeling (16). Whether APN
affects Ang II-induced VSMCs migration was examined by utilizing
the Transwell chamber assay. As depicted in Fig. 4A, APN (10 µg/ml) significantly
attenuated Ang II-induced VSMCs migration. The p38 signaling
pathway is known to be involved in the regulation of VSMC migration
(17) In the present study, the
phosphorylation of p38 mitogen-activated protein kinase (MAPK) was
reduced following APN treatment in Ang II-stimulated VSMCs
(Fig. 4B).
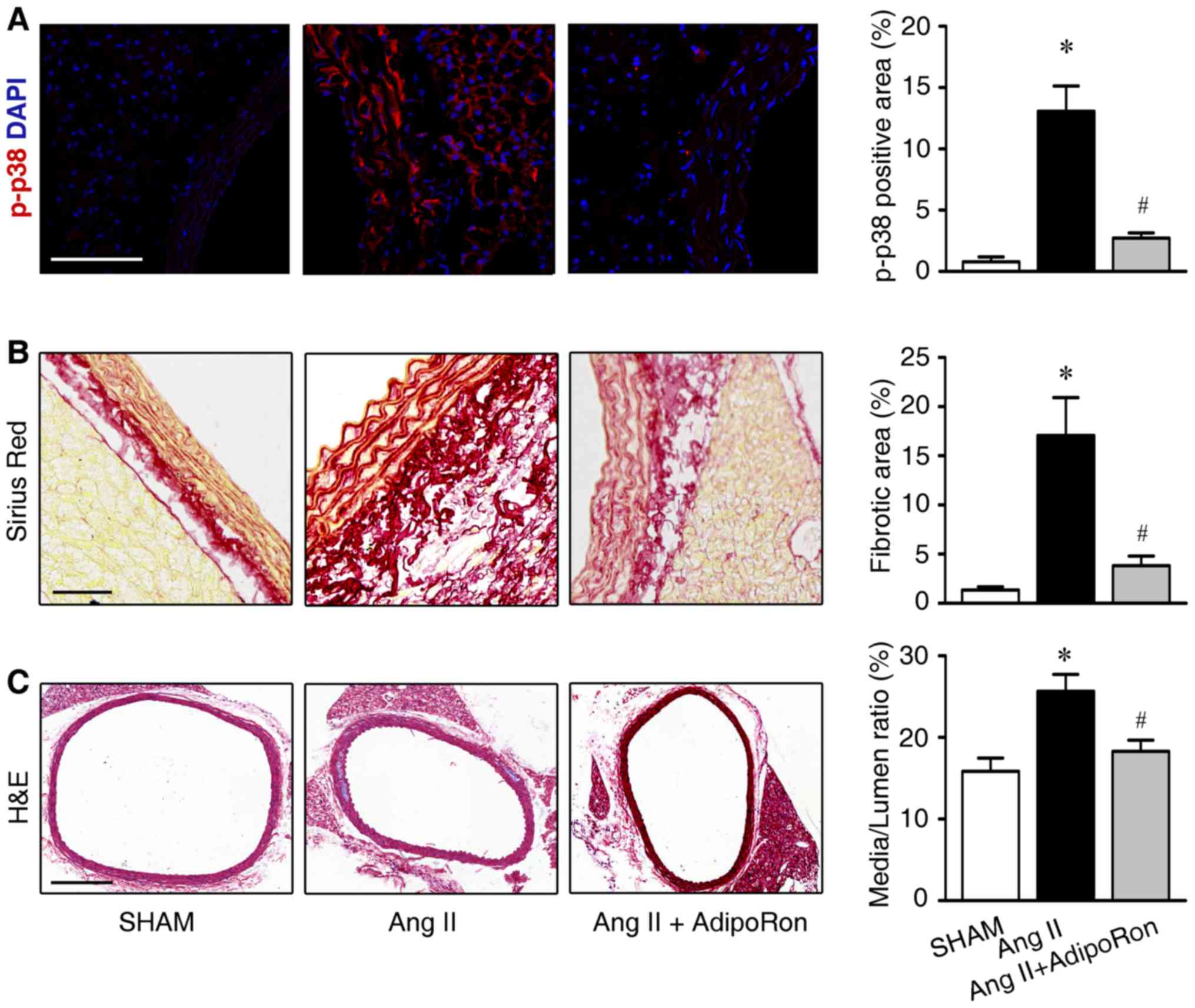
AdipoRon attenuates hypertensive
vascular remodeling
AdipoRon is an orally active APN receptor activator,
which potentially exerts protective effects against cardiac
remodeling (18,19). In order to detect the effects of
AdipoRon in hypertension-associated vascular remodeling, the Ang
II-infused mice were orally treated with AdipoRon (30 mg/kg/day).
As demonstrated in Fig. 5A,
AdipoRon treatment resulted in decreased p38 phosphorylation. In
addition, AdipoRon attenuated Ang II-induced perivascular collagen
deposition and vascular hypertrophy as detected by Sirious Red and
H&E stainings (Fig. 5B and C,
respectively).
Discussion
In the present study, the distribution of APN and
AdipoR in the vasculature was detected and it was demonstrated that
APN was predominantly expressed in the perivascular adipocytes,
while AdipoR1 and R2 were ubiquitously expressed in the blood
vessels. APN attenuated Ang II-induced VSMCs migration and p38
phosphorylation. Furthermore, APN receptor agonist, AdipoRon,
appeared to attenuate Ang II-induced hypertensive vascular
hypertrophy and fibrosis.
Perivascular adipocytes are an important source of
adipokines and their role in regulating vascular function is well
established (20–22). APN, an abundant adipocyte-derived
factor with a variety of biological functions, was first described
in 1995 (23). Over the past two
decades, numerous studies have elucidated its physiological
functions, which may improve insulin sensitivity in insulin target
tissues, modulate inflammatory responses, and serve a crucial role
in the regulation of energy metabolism (24,25).
In the cardiovascular system, APN ameliorates Ang II-induced
cardiac remodeling by attenuating inflammation, it inhibits
connective tissue growth factor-induced VSMCs proliferation, and
promotes endothelial nitric oxide synthase expression and nitric
oxide release from endothelial cells (24,26).
Previous studies have demonstrated that hypertensive mice express
decreased APN plasma levels compared with the controls (27). To the best of our knowledge, the
present study was the first to demonstrate that hypertension
induced APN downregulation in local perivascular adipocytes. VSMC
dysfunction, including migration, proliferation and phenotype
transdifferentiation, has an important role in hypertension-induced
vascular remodeling. VSMC migration is associated with vascular
remodeling when exposed to high stress, such as hypertension. It
has been demonstrated that Ang II significantly increased the VSMC
migration (28). APN receptors,
AdipoR1 and AdipoR2, were originally identified by screening a
skeletal muscle expression library for cDNAs that encoded proteins
that to the globular domain of APN (29). Both receptors contain seven
transmembrane domains and belong to the progestin and adipoQ
receptor family, which has the opposite transmembrane topology to
the G-protein coupled receptors, and have the N-terminus in the
cytoplasm and the C-terminus facing the extracellular space.
Although ubiquitously expressed, AdipoR1 is most abundant in
skeletal muscle whereas AdipoR2 is mainly restricted to the liver
(30,31). In the present study, ubiquitous
expression of AdipoR1 and AdipoR2 in the vasculature was
demonstrated, including ECs, VSMCs and perivascular adipocytes. In
an obesity and diabetic mouse model, AdipoR1 expression was reduced
in the target tissue (32,33). However, in a rat model of chronic
kidney disease, AdipoR1 and AdipoR2 expression was significantly
increased in renal cells (34). In
the present study, AdipoR1 and AdipoR2 expression was decreased in
the cultured VSMCs treated with Ang II. In addition, Ang II-induced
hypertension resulted in a significant decrease of AdipoR1 and
AdipoR2 expression in the perivascular adipocytes and VSMCs in
vivo. AdipoR1 or AdipoR2 mediate the effects of APN by
regulating multiple signaling pathways, including the peroxisome
proliferator-activated receptor-α, the AMP-activated protein
kinase, and the p38 MAPK (12,35,36).
Previous studies have demonstrated that APN could attenuate lipid
deposition and apoptosis through activating p38 MAPK pathway
(37,38). In the present study, it was
demonstrated that APN alone could induce p38 phosphorylation, which
is in accordance with previous studies (39,40).
Furthermore, Ang II promoted p38 phosphorylation. It could be
speculated that the APN/p38 axis may have an inhibitory feedback
effect on p38 phosphorylation, with p38 phosphorylation level is
decreased following Ang II treatment. APN receptor agonist,
AdipoRon, inhibited p38 phosphorylation in the vasculature of Ang
II-induced hypertensive mice.
In conclusion, the present study demonstrated that
Ang II-induced hypertensive mice had decreased expression of APN
and AdipoR in the perivascular adipocytes and VSMCs. APN attenuated
Ang II-induced VSMCs migration and p38 phosphorylation. APN
receptor agonist AdipoRon attenuated Ang II-induced hypertensive
vascular remodeling. These results suggest that APN and APN
receptors may be potential therapeutic targets for inhibiting
hypertensive vascular injury.
Acknowledgements
The present study was supported by the Putuo
Hospital Level Subject (AASI index in old hypertension patients
complicated with metabolism diseases; grant no. PT65487).
References
|
1
|
Thalmann S and Meier CA: Local adipose
tissue depots as cardiovascular risk factors. Cardiovasc Res.
75:690–701. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Molica F, Morel S, Kwak BR,
Rohner-Jeanrenaud F and Steffens S: Adipokines at the crossroad
between obesity and cardiovascular disease. Thromb Haemost.
113:553–566. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li FY, Cheng KK, Lam KS, Vanhoutte PM and
Xu A: Cross-talk between adipose tissue and vasculature: Role of
adiponectin. Acta Physiol (Oxf). 203:167–180. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cai X, Li X, Li L, Huang XZ, Liu YS, Chen
L, Zhang K, Wang L, Li X, Song J, et al: Adiponectin reduces
carotid atherosclerotic plaque formation in ApoE−/−mice: Roles of
oxidative and nitrosative stress and inducible nitric oxide
synthase. Mol Med Rep. 11:1715–1721. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Withers SB, Bussey CE, Saxton SN, Melrose
HM, Watkins AE and Heagerty AM: Mechanisms of
adiponectin-associated perivascular function in vascular disease.
Arterioscler Thromb Vasc Biol. 34:1637–1642. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu Y, Palanivel R, Rai E, Park M, Gabor
TV, Scheid MP, Xu A and Sweeney G: Adiponectin stimulates autophagy
and reduces oxidative stress to enhance insulin sensitivity during
high-fat diet feeding in mice. Diabetes. 64:36–48. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li C, Wang Z, Wang C, Ma Q and Zhao Y:
Perivascular adipose tissue-derived adiponectin inhibits
collar-induced carotid atherosclerosis by promoting macrophage
autophagy. PLoS One. 10:e01240312015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Qi GM, Jia LX, Li YL, Li HH and Du J:
Adiponectin suppresses angiotensin II-induced inflammation and
cardiac fibrosis through activation of macrophage autophagy.
Endocrinology. 155:2254–2265. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yamauchi T, Kamon J, Ito Y, Tsuchida A,
Yokomizo T, Kita S, Sugiyama T, Miyagishi M, Hara K, Tsunoda M, et
al: Cloning of adiponectin receptors that mediate antidiabetic
metabolic effects. Nature. 423:762–769. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang ZV and Scherer PE: Adiponectin, the
past two decades. J Mol Cell Biol. 8:93–100. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhan JK, Wang YJ, Wang Y, Tang ZY, Tan P,
Huang W and Liu YS: Adiponectin attenuates the osteoblastic
differentiation of vascular smooth muscle cells through the
AMPK/mTOR pathway. Exp Cell Res. 323:352–358. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang W, Shu C, Li Q, Li M and Li X:
Adiponectin affects vascular smooth muscle cell proliferation and
apoptosis through modulation of the mitofusin-2-mediated
Ras-Raf-Erk1/2 signaling pathway. Mol Med Rep. 12:4703–4707. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Alexis JD, Wang N, Che W, Lerner-Marmarosh
N, Sahni A, Korshunov VA, Zou Y, Ding B, Yan C, Berk BC and Abe J:
Bcr kinase activation by angiotensin II inhibits
peroxisome-proliferator-activated receptor gamma transcriptional
activity in vascular smooth muscle cells. Circ Res. 104:69–78.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang Z, Hong LK, Follett J, Wabitsch M,
Hamilton NA, Collins BM, Bugarcic A and Teasdale RD: Functional
characterization of retromer in GLUT4 storage vesicle formation and
adipocyte differentiation. Faseb J. 30:1037–1050. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao Y, Zhang C, Wei X, Li P, Cui Y, Qin
Y, Wei X, Jin M, Kohama K and Gao Y: Heat shock protein 60
stimulates the migration of vascular smooth muscle cells via
Toll-like receptor 4 and ERK MAPK activation. Sci Rep. 5:153522015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Montezano AC, Cat Nguyen Dinh A, Rios FJ
and Touyz RM: Angiotensin II and vascular injury. Curr Hypertens
Rep. 16:4312014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu L, Huang X, Huang K, Gui C, Huang Q and
Wei B: Ligustrazine attenuates the platelet-derived growth
factor-BB-induced proliferation and migration of vascular smooth
muscle cells by interrupting extracellular signal-regulated kinase
and P38 mitogen-activated protein kinase pathways. Mol Med Rep.
12:705–711. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Okada-Iwabu M, Yamauchi T, Iwabu M, Honma
T, Hamagami K, Matsuda K, Yamaguchi M, Tanabe H, Kimura-Someya T,
Shirouzu M, et al: A small-molecule AdipoR agonist for type 2
diabetes and short life in obesity. Nature. 503:493–499. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fisman EZ and Tenenbaum A: Adiponectin: A
manifold therapeutic target for metabolic syndrome, diabetes and
coronary disease? Cardiovasc Diabetol. 13:1032014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Takaoka M, Nagata D, Kihara S, Shimomura
I, Kimura Y, Tabata Y, Saito Y, Nagai R and Sata M: Periadventitial
adipose tissue plays a critical role in vascular remodeling. Circ
Res. 105:906–911. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sheng LJ, Ruan CC, Ma Y, Chen DR, Kong LR,
Zhu DL and Gao PJ: Beta3 adrenergic receptor is involved in
vascular injury in deoxycorticosterone acetate-salt hypertensive
mice. FEBS lett. 590:769–778. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Takaoka M, Suzuki H, Shioda S, Sekikawa K,
Saito Y, Nagai R and Sata M: Endovascular injury induces rapid
phenotypic changes in perivascular adipose tissue. Arterioscler
Thromb Vasc Biol. 30:1576–1582. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Scherer PE, Williams S, Fogliano M,
Baldini G and Lodish HF: A novel serum protein similar to C1q,
produced exclusively in adipocytes. J Biol Chem. 270:26746–26749.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee S and Kwak HB: Role of adiponectin in
metabolic and cardiovascular disease. J Exerc Rehabil. 10:54–59.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chakraborti CK: Role of adiponectin and
some other factors linking type 2 diabetes mellitus and obesity.
World J Diabetes. 6:1296–1308. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lima Freitas LC, Braga VA, do Socorro de
Franca Silva M, Cruz JC, Sousa Santos SH, de Oliveira Monteiro MM
and Balarini CM: Adipokines, diabetes and atherosclerosis: An
inflammatory association. Front Physiol. 6:3042015.PubMed/NCBI
|
|
27
|
Ndisang JF and Jadhav A: Heme arginate
therapy enhanced adiponectin and atrial natriuretic peptide, but
abated endothelin-1 with attenuation of kidney histopathological
lesions in mineralocorticoid-induced hypertension. J Pharmacol Exp
Ther. 334:87–98. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang F, Ren X, Zhao M, Zhou B and Han Y:
Angiotensin-(1–7) abrogates angiotensin II-induced proliferation,
migration and inflammation in VSMCs through inactivation of
ROS-mediated PI3 K/Akt and MAPK/ERK signaling pathways. Sci Rep.
6:346212016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kadowaki T and Yamauchi T: Adiponectin and
adiponectin receptors. Endocr Rev. 26:439–451. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yamauchi T and Kadowaki T: Adiponectin
receptor as a key player in healthy longevity and obesity-related
diseases. Cell Metab. 17:185–196. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lustig Y, Hemi R and Kanety H: Regulation
and function of adiponectin receptors in skeletal muscle. Vitam
Horm. 90:95–123. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wade TE, Mathur A, Lu D, Swartz-Basile DA,
Pitt HA and Zyromski NJ: Adiponectin receptor-1 expression is
decreased in the pancreas of obese mice. J Surg Res. 154:78–84.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lin T, Qiu Y, Liu Y, Mohan R, Li Q and Lei
B: Expression of adiponectin and its receptors in type 1 diabetes
mellitus in human and mouse retinas. Mol Vis. 19:1769–1778.
2013.PubMed/NCBI
|
|
34
|
Yu Y, Bao BJ, Fan YP, Shi L and Li SQ:
Changes of adiponectin and its receptors in rats following chronic
renal failure. Ren Fail. 36:92–97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Thundyil J, Pavlovski D, Sobey CG and
Arumugam TV: Adiponectin receptor signalling in the brain. Br J
Pharmacol. 165:313–327. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Almabrouk TA, Ewart MA, Salt IP and
Kennedy S: Perivascular fat, AMP-activated protein kinase and
vascular diseases. Br J Pharmacol. 171:595–617. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yan J, Gan L, Qi R and Sun C: Adiponectin
decreases lipids deposition by p38 MAPK/ATF2 signaling pathway in
muscle of broilers. Mol Biol Rep. 40:7017–7025. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang Y, Zhang J, Zhang L, Gao P and Wu X:
Adiponectin attenuates high glucose-induced apoptosis through the
AMPK/p38 MAPK signaling pathway in NRK-52E cells. PLoS One.
12:e01782152017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pu Y, Wang M, Hong Y, Wu Y and Tang Z:
Adiponectin promotes human jaw bone marrow mesenchymal stem cell
chemotaxis via CXCL1 and CXCL8. J Cell Mol Med. 21:1411–1419. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Huang CY, Chang AC, Chen HT, Wang SW, Lo
YS and Tang CH: Adiponectin promotes VEGF-C-dependent
lymphangiogenesis by inhibiting miR-27b through a CaMKII/AMPK/p38
signaling pathway in human chondrosarcoma cells. Clin Sci (Lond).
130:1523–1533. 2016. View Article : Google Scholar : PubMed/NCBI
|