Introduction
For centuries, Aconitum plants have been extensively
applied to treat various diseases, including inflammation, pain,
neurologic and cardiovascular diseases in China and some other
countries (1,2). Aconitine, one of the major bioactive
alkaloids derived from Aconitum plants, belongs to genus Aconitum
of Ranunculaceae family and is frequently employed to treat
rheumatoid arthritis, cardiologic disorders and tumors (3,4). The
ester combining with C8 and C14 is mainly responsible for its high
toxicity, while hydrolysis of esters can reduce the toxicity
effectively (5). Aconitine
poisoning incidents have happened occasionally caused by misuse,
suicide, or homicide and toxic symptoms include dizziness,
numbness, nausea, vomiting, ventricular arrhythmias, or even death
(6–8). Thus, the application of aconitine in
clinics has been severely limited due to its toxic effects.
In recent years, accumulating evidences have
demonstrated the pharmacological and toxicological characteristics
of aconitine (9–11). Previous studies confirmed that
aconitine could block the inactivation of voltage-dependent sodium
channels causing persistent Na+ influx at resting
potential, thereby leading to arrhythmia (12,13).
Moreover, aconitine diminishes the amplitude of delayed rectifier
K+ current in Jurkat T-lymphocytes, which probably
affects the function of immune cells (14). Ca2+ is well known to
play crucial roles in the pathogenesis of heart dysfunctions and
disruption of intracellular Ca2+ homeostasis may cause
arrhythmia (15,16). It is well established that
aconitine increases intracellular Ca2+, triggers
arrhythmia, and finally induces apoptosis through activation and
phosphorylation of p38 MAPK signaling pathway in rats (17). In addition, recent study revealed
aconitine inhibits tumor growth and induces cell apoptosis via
NF-κB signaling pathway in human pancreatic cancer (18). Based on above findings, it is
convinced that aconitine could induce abnormal electrical
activities in various cell and animal models. However, the impact
of aconitine on cardiomyocytes apoptosis remains unclear.
Mitochondria are well known to be involved in
various physiological and pathologic processes, including
generation of reactive oxygen species, ATP production by oxidative
phosphorylation, and metabolic pathways (19,20).
The peroxisome proliferator activated receptor γ co-activator 1α
(PGC-1α) is a master mitochondrial transcriptional regulator that
promotes mitochondrial biogenesis and energy metabolism in various
cells (21). It has been
demonstrated that PGC-1α expression is significantly decreased in
cardiac hypertrophy and heart failure (22–24).
Considering that PGC-1α plays a vital role in multiple
cardiomyopathies, extensive efforts should be made to explore the
potential effects of aconitine on PGC-1α expression and
mitochondrial function. Meanwhile, recent evidences have also
suggested that the mitochondrial pathway is one of classic
apoptosis pathways (25).
Cytochrome c, as a member of electron transport chain in
mitochondria, is known to act important roles in the electron
transfer and cell respiration (26). Furthermore, the release of
Cytochrome c from mitochondria might subsequently activate the
executioner caspases, resulting in cell apoptosis (27). Therefore, we turned our attention
to aconitine-induced cell apoptosis.
The multiple effects of aconitine have been
reported, however, whether aconitine could induce H9c2 cells
apoptosis through mitochondria-dependent apoptotic pathway has been
little studied. In our present research, H9c2 cell line was used as
a model in vitro exposed to aconitine and further
investigated its possible apoptosis mechanisms.
Materials and methods
Materials
Aconitine was purchased from the National Institute
for the Control of Pharmaceutical and Biological Products (Beijing,
China). The molecular weight of aconitine is 645.74. HPLC analysis
showed the purity was >98%. Aconitine was dissolved in the ethyl
alcohol and diluted to corresponding concentration.
Cell culture and cell viability
assay
Rat embryonic ventricular myocardial H9c2 cells were
obtained from the American Type Culture Collection (ATCC, Manassas,
VA, USA). Cells were cultured in Dulbecco's modified Eagle's medium
(DMEM; Gibco; Invitrogen, Carlsbad, CA) containing 10% fetal bovine
serum (FBS; Gibco; Invitrogen), at 37°C in a humidified incubator
of 5% CO2. Cell viability was measured using Cell
Counting kit-8 (CCK-8; Dojindo, Shanghai, China), according to the
suppliers' instructions. H9c2 cells were seeded at densities of
2×104 cells/ml into 96-well plates and then treated with
different concentrations of aconitine (0–250 µM) for 24 h. CCK-8
was added to each well and then incubated for 4 h. The optical
density was read at 450 nm with ELx808 Absorbance Microplate Reader
(BioTek Instruments, Inc., Winooski, VT, USA) and cell viability
was calculated. The experiments were repeated at least three
times.
Assessments of nuclear morphology by
DAPI staining
Cells were seeded onto 24-well plates and treated
with aconitine (0, 100, 200 µM) for 24 h. Cells were washed with
PBS thrice, fixed in 4% paraformaldehyde for 30 min, then nuclei
were stained with DAPI staining solution for 10 min at room
temperature in the dark. Morphologic changes were observed and
images were acquired by an inverted fluorescence microscope (Nikon
Corporation, Tokyo, Japan).
Detection of apoptotic rate of H9c2
cells by flow cytometry
The Annexin V-FITC/PI Apoptosis Detection kit
(Beyotime Institute of Biotechnology, Haimen, China) was used to
determine apoptosis of cells. In brief, cells were seeded into
6-well plates. After 24 h exposure to aconitine (0–200 µM), cells
were trypsinized, washed twice with cold PBS and re-suspended in
195 µl binding buffer. The cells were then incubated with 5 µl
Annexin V-FITC and 10 µl PI working solution for 15 min in the dark
at room temperature, according to the manufacturer's instructions.
Cellular fluorescence was measured by flow cytometry (FC500;
Beckman Coulter, Inc., Brea, CA, USA). Each experiment was repeated
at least three times.
Reactive oxygen species assay
Intracellular reactive oxygen species (ROS) in H9c2
cells was assessed using Reactive Oxygen Species Assay kit (S0033;
Beyotime Beyotime Institute of Biotechnology, Haimen, China.
DCFH-DA, a non-fluorescent probe, can be hydrolyzed to DCFH, and
further oxidized to the highly fluorescent compound
dichlorofluorescein (DCF) in the presence of ROS. H9c2 cells were
incubated with aconitine for 4 h, subsequently, treated with 10 µM
DCFH-DA for 20 min at 37°C. The cells were washed with PBS three
times and changes of green fluorescence were observed by
fluorescence microscope (excitation 488 nm, emission 525 nm). In
addition, the cells were also harvested and washed twice with PBS,
then analyzed using flow cytometry (FC500; Beckman Coulter,
Inc.).
ATP contents assay
Cellular ATP contents were assessed by Enhanced ATP
Assay kit (S0027; Beyotime) according to the manufacturer's
instructions. Briefly, cells were seeded at 4×105
cells/well in six-well plates. After incubated with aconitine for
24 h, cells were rinsed and lysed using ATP lysis buffer on ice.
Samples were collected and centrifuged at 12,000 rpm for 10 min at
4°C to acquire supernatant for further determination. Samples and
ATP detection working dilution were added and luminescence activity
was measured immediately using luminometer (GloMax 20/20; Promega
Corporation, Madison, WI, USA). Standard curve of ATP measure was
made in each assay. Subsequently, the intracellular ATP contents
were normalized by the protein contents in each sample.
Detection of mitochondrial
transmembrane potential
Mitochondrial Membrane Potential Assay kit (JC-1;
Beyotime Institute of Biotechnology) was used to evaluate
mitochondrial membrane potential (∆Ψm) of cells following the
manufacturer's instructions. Briefly, cells were seeded onto
24-well plates and then treated with aconitine (0–200 µM) for 4 h.
Cells were incubated with the medium containing JC-1 (molecular
probes) for 20 min at 37°C. CCCP was used as a positive control to
induce the decrease of mitochondrial membrane potential. After
washing with ice-cold JC-1 (1x) for two times, images were observed
by a fluorescent microscope (Nikon Corporation). Additionally,
cells were collected and rinsed with PBS, then analyzed using flow
cytometry (FC500; Beckman Coulter, Inc.). The ratios of red
fluorescence intensity over green fluorescence intensity
represented the levels of ΔΨm.
Western blot analysis
Cells treated with different concentrations of
aconitine were lysed with the RIPA Lysis Buffer (Beyotime Institute
of Biotechnology) and protein concentrations were measured by BCA
Protein assay kit (Beyotime Institute of Biotechnology). Equal
amount of protein extracts (80 µg) from each sample was separated
by 10 or 12% SDS-PAGE and transferred to polyvinylidene fluoride
(PVDF) membrane. The membranes were blocked with 5% BSA and then
incubated overnight at 4°C with primary antibodies against Bax
(1:1,000; 2772S; Cell Signaling Technology, Inc., Danvers, MA,
USA), Bcl-2 (1:1,000; ab13285; Abcam, Cambridge, UK), PGC-1α
(1:1,000; ab191838; Abcam), Caspase-3 (1:1,000; ab179517; Abcam),
Cytochrome c (1:1,000; 136F3; Cell Signaling Technology) and
β-Actin (1:5,000; AT0001; CMCTAG). Membranes were washed three
times and incubated with secondary antibody at a dilution of
1:1,000 in the same buffer for 2 h at room temperature. After
washed three times, membranes were visualized using the ECL
chemiluminescence system (ChemiScope, Shanghai). The intensity of
specific brands was quantified and normalized to a loading control.
All the experiments were conducted at least three times.
Statistical analysis
All experiments were repeated at least three times.
Data were expressed as means ± SD. All statistical analyses were
performed using GraphPad Prism software version 5.0. The
statistical significance of differences were evaluated using
one-way ANOVA followed by Bonferron's post hoc correction.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Aconitine suppresses cell viability in
cultured H9c2 cells
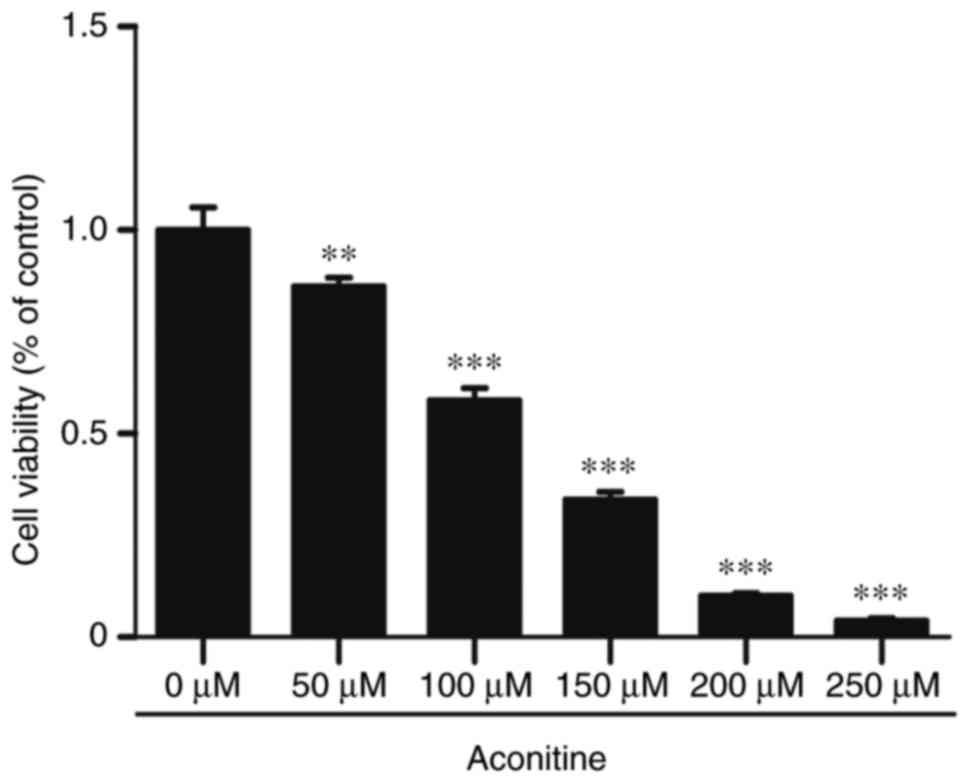
The cytotoxic effect of aconitine on H9c2 cells was
detected by CCK-8 kit. As shown in Fig. 1, aconitine treatment for 24 h
potently suppressed cell viability in a concentration-dependent
manner (0–250 µM). Compared with the control group, cells were
treated with 100, 200 µM of aconitine, cell viability were
significantly decreased to 58±2.91 and 10±0.5%, respectively. The
results indicated that aconitine could remarkably inhibit cell
viability in a concentration-dependent manner in H9c2 cells.
Assessments of nuclear morphology by
DAPI staining
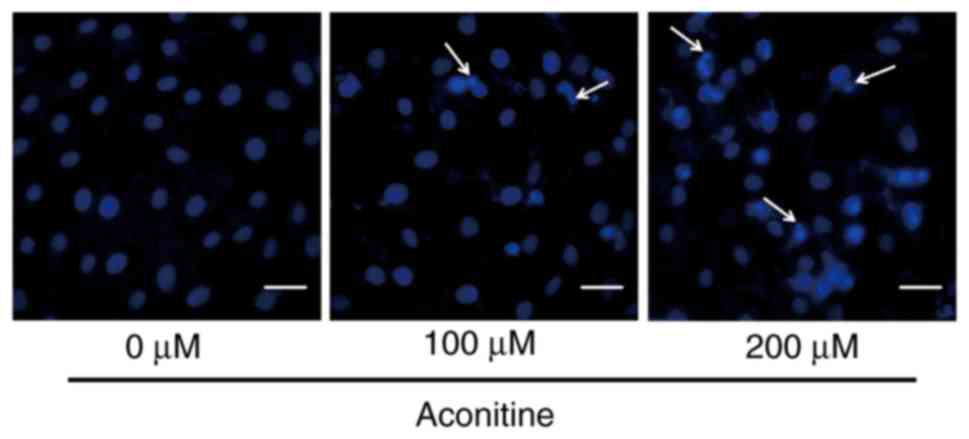
To verify the effects of aconitine on apoptosis in
H9c2 cells, DAPI staining was used to observe the morphological
changes of aconitine-treated H9c2 cells. The fragmentation of cell
nuclei exposed to aconitine for 24 h was observed, and cell nuclei
shrinkage and chromatin condensation were increased slightly with
the concentration of aconitine in H9c2 cells as compared to the
control (Fig. 2).
Detection of aconitine-induced cell
apoptosis by flow cytometry
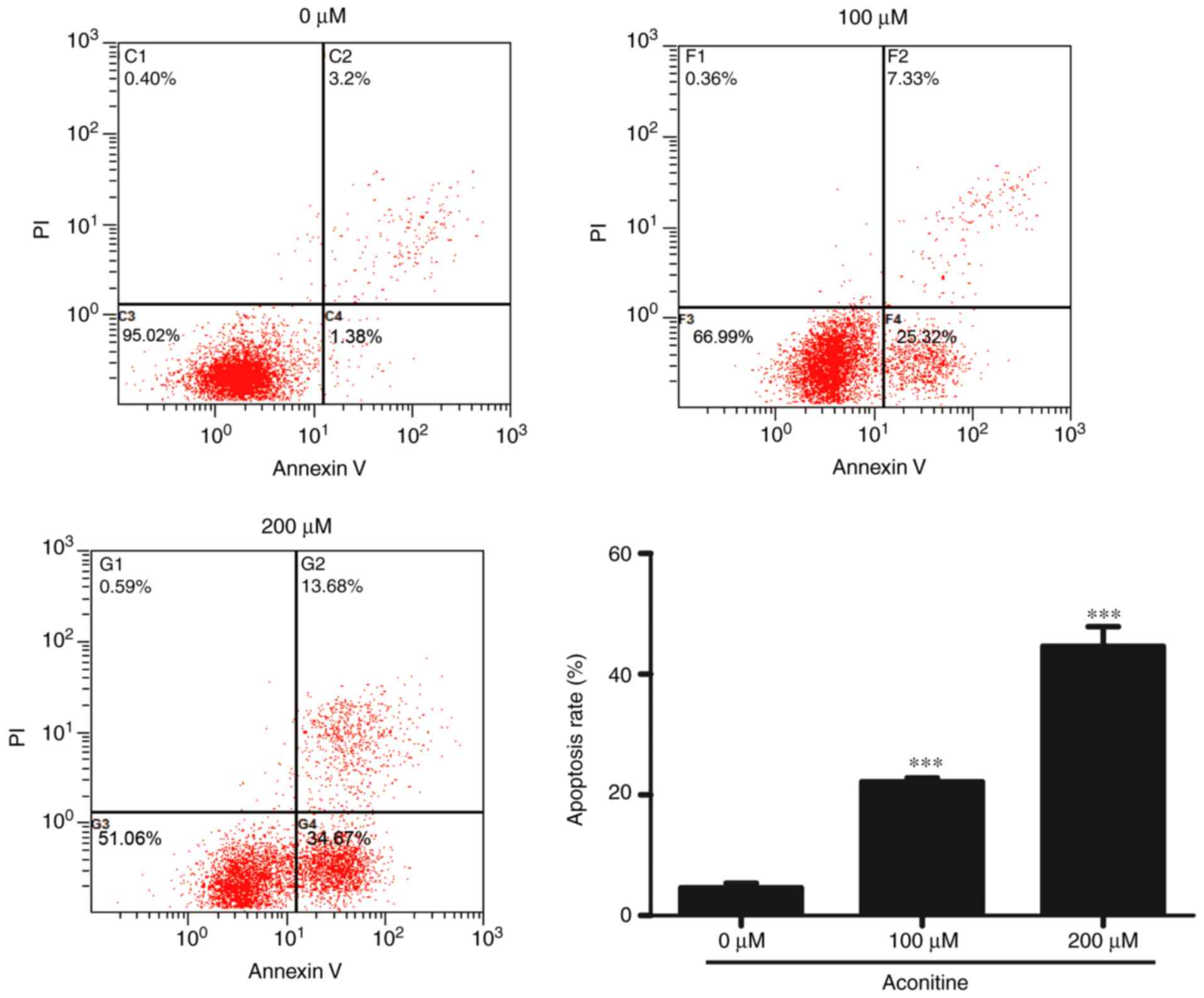
In attempt to illustrate whether the decreased cell
viability induced by aconitine was associated with cell apoptosis,
Annexin V-FITC/PI double staining was further performed. As shown
in Fig. 3, after exposed to 0,
100, and 200 µM aconitine for 24 h, cell apoptosis rates of both
early and late apoptosis were 4.61±0.76, 22.16±0.64, and
44.64±3.23%, respectively, and the cell necrosis rate significantly
increased. Our data indicated that treatment with aconitine not
only reduced the numbers of surviving cells but also increased the
cell numbers in early and late apoptosis.
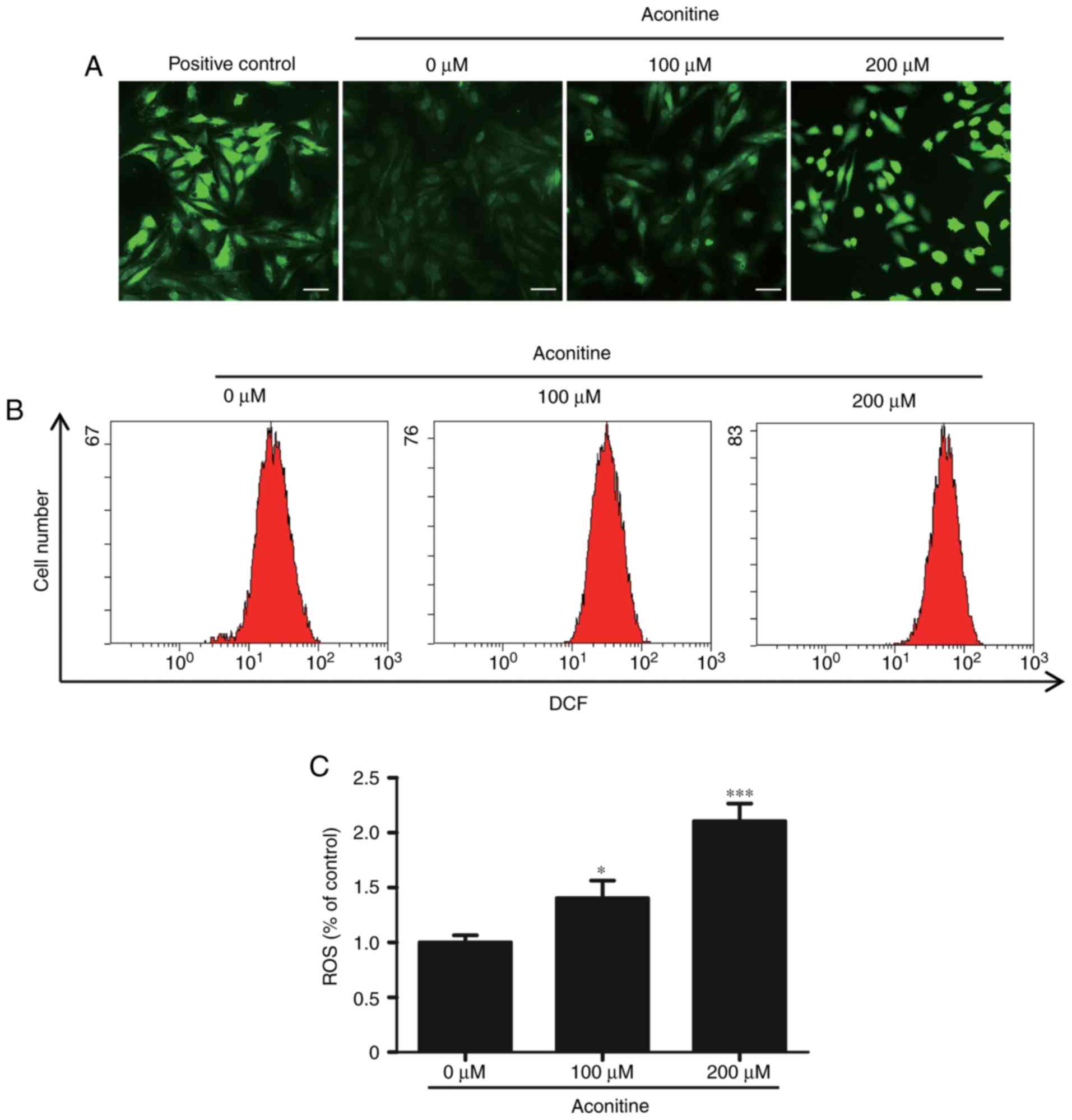
Effects of aconitine on reactive
oxygen species
Reactive oxygen species is mainly produced by
mitochondria, which is associated with the process of cells
apoptosis. Meanwhile, excessive ROS can influence cell viability
and alter mitochondrial function. In order to clarify whether
aconitine could increase cellular ROS levels, H9c2 cells were
treated with aconitine (0–200 µM) for 4 h. As seen in Fig. 4A, green fluorescence distinctly
increased after treatment with aconitine. Flow cytometry analysis
revealed that the levels of ROS after treated with aconitine (200
µM) were obviously increased to approximately two folds compared
with the control cells (Fig. 4B and
C). The results of flow cytometry were consistent with the
fluorescent images, indicating that aconitine could effectively
promote the production of ROS in H9c2 cells compared with the
control group.
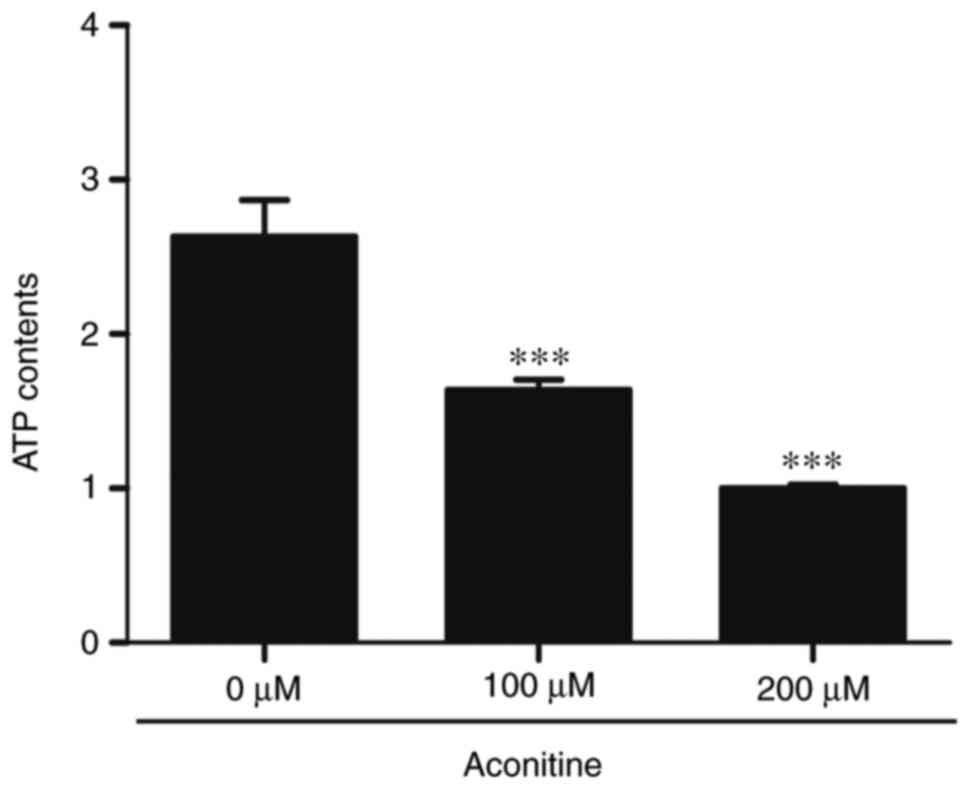
Inhibition the generation of
intracellular ATP induced by aconitine
To evaluate the changes of ATP contents during
apoptosis of H9c2 cells induced by aconitine, ATP contents were
detected by enhanced ATP assay kit. As depicted in Fig. 5, the cellular ATP contents were
decreased in a concentration-dependent manner after exposure to
aconitine for 24 h compared with the control. Aconitine (200 µM)
notably decreased ATP contents, as compared to control.
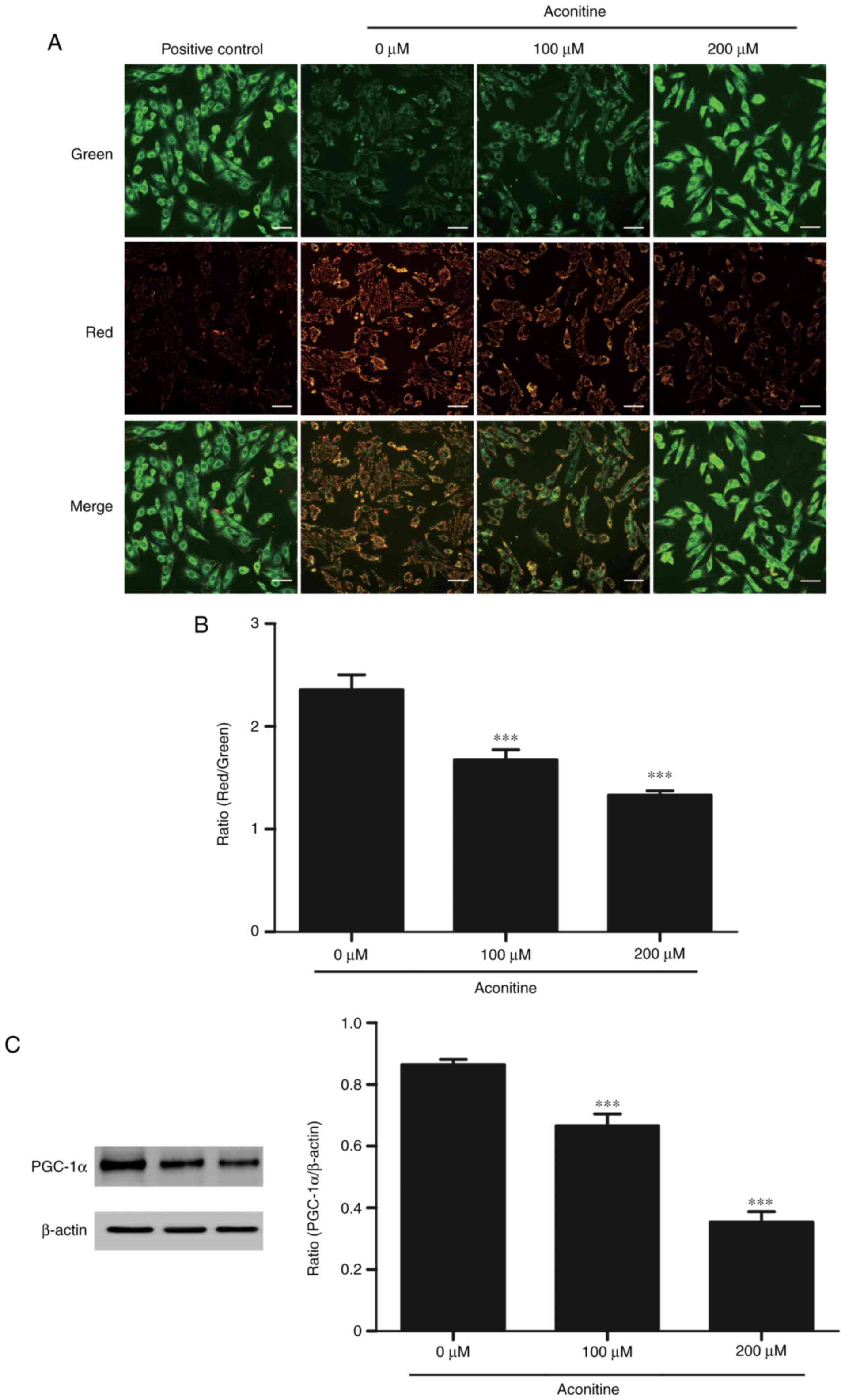
Decreased mitochondrial transmembrane
potential and PGC-1α expression induced by aconitine
To investigate the effects of aconitine on
mitochondrial functions in H9c2 cells, mitochondrial transmembrane
potential was measured by JC-1 staining. As shown in Fig. 6, fluorescent images of H9c2 cells
stained with JC-1 showed that cells emitted red fluorescence with a
little green fluorescence in control group, suggesting that
mitochondrial membrane potential was normal. When ∆Ψm is low, the
JC-1 aggregates becomes monomer form green fluorescence with a
little red fluorescence in cells treated with aconitine (100, 200
µM) for 4 h. The ratio of red and green fluorescence indicates the
relative level of ∆Ψm. The results showed that the ∆Ψm was
significantly lower compared with the control group. Moreover,
aconitine decreased PGC-1α expression remarkably.
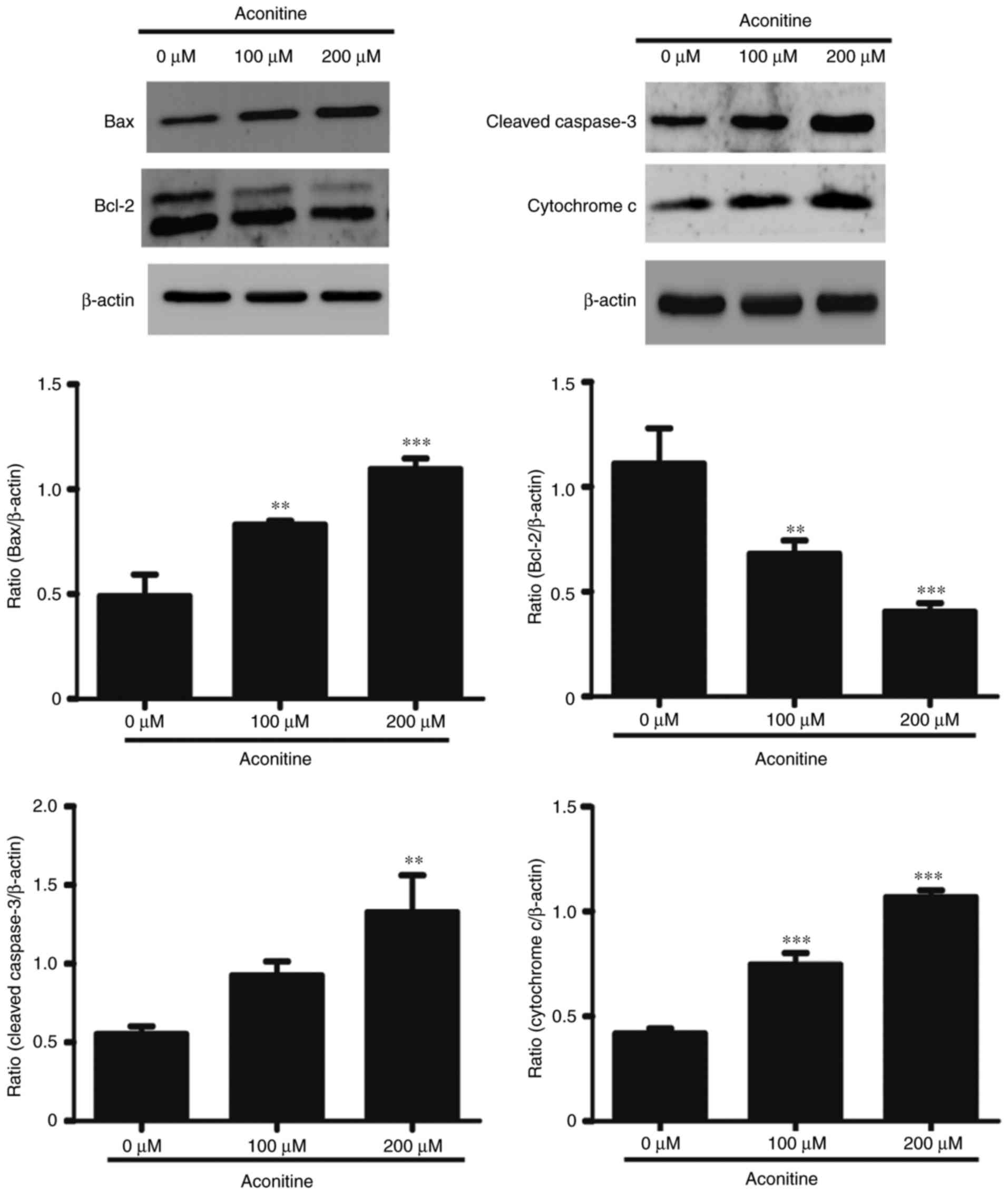
Aconitine induced apoptosis-related
proteins expression in H9c2 cells
To confirm the signaling pathway involved cell
apoptosis, western blotting was further performed. We examined the
expression of Caspase-3, Bcl-2, Bax, Cytochrome c (Fig. 7). These results demonstrates
aconitine could upregulate Bax and cleaved Caspase-3, as well as
downregulate Bcl-2. In addition, aconitine significantly increased
the expression level of Cytochrome c.
Discussion
Aconitum alkaloids are mainly used in China and
other Asian countries to treat rheumatoid arthritis and
cardiovascular disorders (28).
However, the high toxicity restricts its clinical application
(29). Previous studies have
mainly focused on the aconitine-induced arrhythmia. However, little
information is available about the impacts of aconitine on
mitochondria. Mitochondria are known to have critical roles in
regulating energy production and metabolism especially in high
energy demanding cells, such as cardiomyocytes and neuronal cells
(30). Emerging researches have
demonstrated that the integrity of mitochondrial structure and
function is essential for maintaining normal cardiac function
(31,32). Therefore, we paid our attention to
investigate the effects of aconitine on mitochondria and explore
the relevant apoptosis signaling pathways in H9c2 cells.
To investigate the effects of aconitine on H9c2
cells, CCK-8 assay was firstly used to assess the cytotoxicity. It
is intriguing that cell viability was potently inhibited after
exposed to aconitine for 24 h. Nevertheless, the results of Annexin
V-FITC/PI double staining indicated that aconitine could markedly
induce cell apoptosis in early and late apoptosis. Consistent with
the above results, DAPI staining showed that aconitine-exposed H9c2
cells exhibited obvious morphological changes of apoptosis
including cell nuclei shrinkage and chromatin condensation. In
conclusion, the results demonstrated aconitine could induce
apoptosis in H9c2 cells.
Mitochondrial membrane potential determined by the
difference of mitochondrial inner and outer membrane acts a vital
role in maintaining mitochondrial function (33). ROS, a product of aerobic metabolism
in mitochondria, is involved in early stages of apoptosis, and
triggers the loss of the ∆Ψm (34,35).
PGC-1α, as a crucial coactivator of nuclear receptors, stimulates
mitochondrial biogenesis and energy metabolism. Ectopic expression
of PGC-1α could result in mitochondrial ultrastructural
abnormalities and mitochondrial dysfunction (36,37).
Previous studies have demonstrated that the dissipation of ∆Ψm
could impact the generation of ATP and ROS, initiate the release of
pro-apoptotic factors, thereby leading to mitochondrial dysfunction
and cell apoptosis (38). In the
study, we confirmed that aconitine enhanced ROS generation,
decreased ATP contents and ∆Ψm, and downregulated PGC-1α
expression, indicating that mitochondrial damage might be concerned
with the development of aconitine-induced apoptosis in H9c2
cells.
Apoptosis is a programmed cell death process, which
is generally modulated via three major pathways:
Mitochondrial-dependent pathway, endoplasmic reticulum pathway, and
death receptor pathway (39–41).
It seems increasingly evident that mitochondrial-dependent pathway
has been considered as the one of the major signaling pathways in
apoptosis of various cells (42).
More importantly, mitochondrial transmembrane potential, reactive
oxygen species, Bcl-2 family members, and caspases are associated
with mitochondrial-mediated apoptosis pathway (43). We further analyzed the expression
of apoptosis associated proteins including Bcl-2 family proteins
(Bcl-2, Bax), Caspase-3, and Cytochrome c by western blotting to
confirm the possible mechanisms connected with aconitine-induced
apoptosis. Anti-apoptotic protein Bcl-2 decreased while Bax
increased after treatment with aconitine. Thus, the ratio of
Bcl-2/Bax, which plays a crucial role for the activation of the
mitochondrial apoptotic pathway, was notably decreased in cells
exposed to aconitine. Bcl-2 family proteins are involved in the
process of apoptosis and play central role in regulating
mitochondrial-dependent apoptotic pathway (44,45).
It is worth mentioning that the release of Cytochrome c is
essential to initiate mitochondrial-mediated cells apoptotic
pathway (46,47). The levels of Cytochrome c,
Caspase-3 were significantly increased at the concentrations of
100, 200 µM aconitine, which are in line with the severity of cell
apoptosis. Increasing evidences also suggest that mitochondria play
a crucial role in the release of Cytochrome c and pro-apoptotic
proteins, which subsequently active caspases and induce apoptosis
(48,49). In addition, previous studies have
demonstrated PGC-1α was downregulated in human breast cancer, colon
cancer (50,51), indicating that PGC-1α may be
associated with the prognosis of cancers. More recently, the
expression of PGC-1α decreased was also found in human epithelial
ovarian cancer and PGC-1α overexpression could induce Ho-8910 cell
apoptosis by the PPARγ-dependent pathway (52). Contrary with the result, we found
decreased PGC-1α was associated with the process of cell apoptosis,
which is probably due to cancer cells differ from normal cells in
many ways that allow them to grow out of control, accompany gene
mutations and become invasive. More importantly, it has been
demonstrated doxorubicin decreased significantly the expression of
PPARα and PGC-1α in primary cardiomyocytes in vitro,
indicating that PGC-1α could involve energy metabolism remodeling
and induce cells apoptosis (53,54).
We speculate decreased PGC-1α may correlate with apoptosis in H9c2
cells exposed to aconitine, however, the functional mechanisms of
PGC-1α in aconitine-induced cells apoptosis still need to be
further investigated. Our results suggested that
mitochondrial-dependent pathway is partially involved in
aconitine-induced apoptosis in H9c2 cells.
In summary, the present study demonstrated that
aconitine induced apoptosis of H9c2 cells at least in part via
mitochondria-dependent apoptotic pathway. As described already, the
apoptotic pathway was triggered by decreased PGC-1α expression,
induced mitochondrial dysfunction, upregulated Cytochrome c, Bax,
cleaved Caspase-3, downregulated Bcl-2, ultimately leading to
apoptosis of H9c2 cells. Although H9c2 cells provided a unique
model in vitro to investigate the mechanisms of cells
apoptosis in our preliminary study, there is still some limitations
including single cell line. Thus, animal model and primary
cardiomyocytes will be used to further validate our conclusion in
the following research. Therefore, our findings may provide a
possible mechanism of the aconitine-induced apoptosis partially
through mitochondria-mediated pathway in H9c2 cells.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81571848) and the
Priority Academic Program Development of Jiangsu Higher Education
Institutions.
References
|
1
|
Chen JH, Lee CY, Liau BC, Lee MR, Jong TT
and Chiang ST: Determination of aconitine-type alkaloids as markers
in fuzi (Aconitum carmichaeli) by LC/(+)ESI/MS(3). J Pharm Biomed
Anal. 48:1105–1111. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hikino H, Murakami M, Konno C and Watanabe
H: Determination of aconitine alkaloids in aconitum roots. Planta
Med. 48:67–71. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li X, Gu L, Yang L, Zhang D and Shen J:
Aconitine: A potential novel treatment for systemic lupus
erythematosus. J Pharmacol Sci. 133:115–121. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang X, Wang H, Zhang A, Lu X, Sun H, Dong
H and Wang P: Metabolomics study on the toxicity of aconite root
and its processed products using ultraperformance
liquid-chromatography/electrospray-ionization synapt
high-definition mass spectrometry coupled with pattern recognition
approach and ingenuity pathways analysis. J Proteome Res.
11:1284–1301. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li TF, Gong N and Wang YX: Ester
hydrolysis differentially reduces aconitine-induced
anti-hypersensitivity and acute neurotoxicity: Involvement of
spinal microglial dynorphin expression and implications for
aconitum processing. Front Pharmacol. 7:3672016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fujita Y, Terui K, Fujita M, Kakizaki A,
Sato N, Oikawa K, Aoki H, Takahashi K and Endo S: Five cases of
aconite poisoning: Toxicokinetics of aconitines. J Anal Toxicol.
31:132–137. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pullela R, Young L, Gallagher B, Avis SP
and Randell EW: A case of fatal aconitine poisoning by Monkshood
ingestion. J Forensic Sci. 53:491–494. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Arlt EM, Keller T, Wittmann H and
Monticelli F: Fatal aconitine intoxication or thyroid storm? A case
report. Leg Med (Tokyo). 14:154–156. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu XX, Jian XX, Cai XF, Chao RB, Chen QH,
Chen DL, Wang XL and Wang FP: Cardioactive C19-diterpenoid
alkaloids from the lateral roots of Aconitum carmichaeli ‘Fu Zi’.
Chem Pharm Bull (Tokyo). 60:144–149. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhu L, Wu J, Zhao M, Song W, Qi X, Wang Y,
Lu L and Liu Z: Mdr1a plays a crucial role in regulating the
analgesic effect and toxicity of aconitine by altering its
pharmacokinetic characteristics. Toxicol Appl Pharmacol. 320:32–39.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ono T, Hayashida M, Tezuka A, Hayakawa H
and Ohno Y: Antagonistic effects of tetrodotoxin on
aconitine-induced cardiac toxicity. J Nippon Med Sch. 80:350–361.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wright SN: Comparison of
aconitine-modified human heart (hH1) and rat skeletal (mu1) muscle
Na+ channels: An important role for external Na+ ions. J Physiol.
538:759–771. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang SY and Wang GK: Voltage-gated sodium
channels as primary targets of diverse lipid-soluble neurotoxins.
Cell Signal. 15:151–159. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu SN, Chen BS and Lo YC: Evidence for
aconitine-induced inhibition of delayed rectifier K(+) current in
Jurkat T-lymphocytes. Toxicology. 289:11–18. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fu M, Wu M, Wang JF, Qiao YJ and Wang Z:
Disruption of the intracellular Ca2+ homeostasis in the cardiac
excitation-contraction coupling is a crucial mechanism of
arrhythmic toxicity in aconitine-induced cardiomyocytes. Biochem
Biophys Res Commun. 354:929–936. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou YH, Piao XM, Liu X, Liang HH, Wang
LM, Xiong XH, Wang L, Lu YJ and Shan HL: Arrhythmogenesis toxicity
of aconitine is related to intracellular ca(2+) signals. Int J Med
Sci. 10:1242–1249. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun GB, Sun H, Meng XB, Hu J, Zhang Q, Liu
B, Wang M, Xu HB and Sun XB: Aconitine-induced Ca2+ overload causes
arrhythmia and triggers apoptosis through p38 MAPK signaling
pathway in rats. Toxicol Appl Pharmacol. 279:8–22. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ji BL, Xia LP, Zhou FX, Mao GZ and Xu LX:
Aconitine induces cell apoptosis in human pancreatic cancer via
NF-κB signaling pathway. Eur Rev Med Pharmacol Sci. 20:4955–4964.
2016.PubMed/NCBI
|
|
19
|
Kakkar P and Singh BK: Mitochondria: A hub
of redox activities and cellular distress control. Mol Cell
Biochem. 305:235–253. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kroemer G, Dallaporta B and Resche-Rigon
M: The mitochondrial death/life regulator in apoptosis and
necrosis. Annu Rev Physiol. 60:619–642. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Won JC, Park JY, Kim YM, Koh EH, Seol S,
Jeon BH, Han J, Kim JR, Park TS, Choi CS, et al: Peroxisome
proliferator-activated receptor-gamma coactivator 1-alpha
overexpression prevents endothelial apoptosis by increasing ATP/ADP
translocase activity. Arterioscler Thromb Vasc Biol. 30:290–297.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Garnier A, Fortin D, Deloménie C, Momken
I, Veksler V and Ventura-Clapier R: Depressed mitochondrial
transcription factors and oxidative capacity in rat failing cardiac
and skeletal muscles. J Physiol. 551:491–501. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Patten IS and Arany Z: PGC-1 coactivators
in the cardiovascular system. Trends Endocrinol Metab. 23:90–97.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sihag S, Cresci S, Li AY, Sucharov CC and
Lehman JJ: PGC-1alpha and ERRalpha target gene downregulation is a
signature of the failing human heart. J Mol Cell Cardiol.
46:201–212. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ding F, Shao ZW, Yang SH, Wu Q, Gao F and
Xiong LM: Role of mitochondrial pathway in compression-induced
apoptosis of nucleus pulposus cells. Apoptosis. 17:579–590. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Babbitt SE, Sutherland MC, Francisco San
B, Mendez DL and Kranz RG: Mitochondrial cytochrome c biogenesis:
No longer an enigma. Ternds Biochem Sci. 40:446–455. 2015.
View Article : Google Scholar
|
|
27
|
Wei H, Li Z, Hu S, Chen X and Cong X:
Apoptosis of mesenchymal stem cells induced by hydrogen peroxide
concerns both endoplasmic reticulum stress and mitochondrial death
pathway through regulation of caspases, p38 and JNK. J Cell
Biochem. 111:967–978. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang P, Zhang F, Wang Z, Jiang Y and Lu
Y: Simultaneous determination of four trace aconitum alkaloids in
urine using ultra performance liquid chromatography-mass
spectrometry. Se Pu. 31:211–217. 2013.(In Chinese). PubMed/NCBI
|
|
29
|
Lin CC, Chan TY and Deng JF: Clinical
features and management of herb-induced aconitine poisoning. Ann
Emerg Med. 43:574–579. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Riba A, Deres L, Eros K, Szabo A, Magyar
K, Sumegi B, Toth K, Halmosi R and Szabados E: Doxycycline protects
against ROS-induced mitochondrial fragmentation and ISO-induced
heart failure. PLoS One. 12:e01751952017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schilling JD: The mitochondria in diabetic
heart failure: From pathogenesis to therapeutic promise. Antioxid
Redox Signal. 22:1515–1526. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ong SB, Subrayan S, Lim SY, Yellon DM,
Davidson SM and Hausenloy DJ: Inhibiting mitochondrial fission
protects the heart against ischemia/reperfusion injury.
Circulation. 121:2012–2022. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jayaraman S: Flow cytometric determination
of mitochondrial membrane potential changes during apoptosis of T
lymphocytic and pancreatic beta cell lines: Comparison of
tetramethylrhodamineethylester (TMRE), chloromethyl-X-rosamine
(H2-CMX-Ros) and MitoTracker Red 580 (MTR580). J Immunol Methods.
306:68–79. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pourahmad J and O'Brien PJ: A comparison
of hepatocyte cytotoxic mechanisms for Cu2+ and Cd2+. Toxicology.
143:263–273. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shih CM, Ko WC, Wu JS, Wei YH, Wang LF,
Chang EE, Lo TY, Cheng HH and Chen CT: Mediating of
caspase-independent apoptosis by cadmium through the
mitochondria-ROS pathway in MRC-5 fibroblasts. J Cell Biochem.
91:384–397. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Huss JM and Kelly DP: Nuclear receptor
signaling and cardiac energetics. Circ Res. 95:568–578. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Russell LK, Mansfield CM, Lehman JJ,
Kovacs A, Courtois M, Saffitz JE, Medeiros DM, Valencik ML,
McDonald JA and Kelly DP: Cardiac-specific induction of the
transcriptional coactivator peroxisome proliferator-activated
receptor gamma coactivator-1 alpha promotes mitochondrial
biogenesis and reversible cardiomyopathy in a developmental
stage-dependent manner. Circ Res. 94:525–533. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Luo G, Xu X, Guo W, Luo C, Wang H, Meng X,
Zhu S and Wei Y: Neuropeptide Y damages the integrity of
mitochondrial structure and disrupts energy metabolism in cultured
neonatal rat cardiomyocytes. Peptides. 71:162–169. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hou Q, Cymbalyuk E, Hsu SC, Xu M and Hsu
YT: Apoptosis modulatory activities of transiently expressed Bcl-2:
Roles in cytochrome C release and Bax regulation. Apoptosis.
8:617–629. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Won SJ, Chung KS, Ki YS, Choi JH, Cho WJ
and Lee KT: CWJ-081, a novel 3-arylisoquinoline derivative, induces
apoptosis in human leukemia HL-60 cells partially involves reactive
oxygen species through c-Jun NH2-terminal kinase pathway. Bioorg
Med Chem Lett. 20:6447–6451. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao YY, Shen X, Chao X, Ho CC, Cheng XL,
Zhang Y, Lin RC, Du KJ, Luo WJ, Chen JY and Sun WJ:
Ergosta-4,6,8(14),22-tetraen-3-one induces G2/M cell cycle arrest
and apoptosis in human hepatocellular carcinoma HepG2 cells.
Biochim Biophys Acta. 1810:384–390. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Olson M and Kornbluth S: Mitochondria in
apoptosis and human disease. Curr Mol Med. 1:91–122. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Khazaei S, Ramachandran V, Hamid Abdul R,
Mohd Esa N, Etemad A, Moradipoor S and Ismail P: Flower extract of
Allium atroviolaceum triggered apoptosis, activated caspase-3 and
down-regulated antiapoptotic Bcl-2 gene in HeLa cancer cell line.
Biomed Pharmacother. 89:1216–1226. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hardwick JM, Chen YB and Jonas EA:
Multipolar functions of BCL-2 proteins link energetics to
apoptosis. Trends Cell Biol. 22:318–328. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Rao SR, Sundararajan S, Subbarayan R and
Girija Murugan D: Cyclosporine-A induces endoplasmic reticulum
stress and influences pro-apoptotic factors in human gingival
fibroblasts. Mol Cell Biochem. 429:179–185. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Saelens X, Festjens N, Vande Walle L, van
Gurp M, van Loo G and Vandenabeele P: Toxic proteins released from
mitochondria in cell death. Oncogene. 23:2861–2874. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huang QR, Li Q, Chen YH, Li L, Liu LL, Lei
SH, Chen HP, Peng WJ and He M: Involvement of anion exchanger-2 in
apoptosis of endothelial cells induced by high glucose through an
mPTP-ROS-Caspase-3 dependent pathway. Apoptosis. 15:693–704. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ravindran J, Gupta N, Agrawal M, Bhaskar
Bala AS and Rao Lakshmana PV: Modulation of ROS/MAPK signaling
pathways by okadaic acid leads to cell death via, mitochondrial
mediated caspase-dependent mechanism. Apoptosis. 16:145–161. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Feilchenfeldt J, Bründler MA, Soravia C,
Tötsch M and Meier CA: Peroxisome proliferator-activated receptors
(PPARs) and associated transcription factors in colon cancer:
Reduced expression of PPARgamma-coactivator 1 (PGC-1). Cancer Lett.
203:25–33. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jiang WG, Douglas-Jones A and Mansel RE:
Expression of peroxisome-proliferator activated receptor-gamma
(PPARgamma) and the PPARgamma co-activator, PGC-1, in human breast
cancer correlates with clinical outcomes. Int J Cancer.
106:752–757. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang Y, Ba Y, Liu C, Sun G, Ding L, Gao
S, Hao J, Yu Z, Zhang J, Zen K, et al: PGC-1alpha induces apoptosis
in human epithelial ovarian cancer cells through a
PPARgamma-dependent pathway. Cell Res. 17:363–373. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yang Y, Zhang H, Li X, Yang T and Jiang Q:
Effects of PPARα/PGC-1α on the myocardial energy metabolism during
heart failure in the doxorubicin induced dilated cardiomyopathy in
mice. Int J Clin Exp Med. 7:2435–2442. 2014.PubMed/NCBI
|
|
54
|
Yang Y, Zhang H, Li X, Yang T and Jiang Q:
Effects of PPARα/PGC-1α on the energy metabolism remodeling and
apoptosis in the doxorubicin induced mice cardiomyocytes in vitro.
Int J Clin Exp Pathol. 8:12216–12224. 2015.PubMed/NCBI
|