Introduction
The morbidity and prevalence associated with
allergic diseases has increased markedly over the past few decades
(1,2), and allergic diseases are considered
the most common type of human disease worldwide (3). A survey investigating the
epidemiology of allergic diseases within 30 countries revealed that
22% of individuals had immunoglobulin E (IgE)-mediated allergic
diseases, including asthma, rhinitis and conjunctivitis; severe
allergic diseases associated with trees or plants have been
reported to affect the quality of life of children and adults
(4,5). Among 1,500 million patients, ~50% of
adults and ≥80% of children who suffer from asthma were reported to
be induced by allergic factors; the World Health Organization have
estimated that >180,000 annual cases of mortality are associated
with asthma (6).
Platanus acerifolia pollen has been described
to cause airway-associated allergies worldwide, particularly during
early spring (7). Three major
allergens have been identified within P. acerifolia pollen,
including Pla a 1, Pla a 2 and Pla a 3. The nonglycosylated protein
Pla a 1 has a prevalence of 84% among Platanus-allergic
patients (8). Pla a 2 is a
glycoprotein associated with the allergic responses of 84% of
patients with Platanus-induced pollinosis (9,10).
Pla a 3 is a non-specific lipid transfer protein (9). Evidence has indicated that 45% of
patients allergic to P. acerifolia pollen are sensitive to
natural Pla a 3 (11). In the
present study, the Pla a 2 allergen was expressed and purified from
an Escherichia coli system, which may provide a foundation
for the future study of the diagnosis and treatment of P.
acerifolia pollen-associated allergies.
Allergen-specific immunotherapy (AIT) has been
reported to reduce the severity of symptoms in people with allergic
asthma (12,13), allergic rhinitis (14) and conjunctivitis (15). The efficacy and safety of AIT has
been demonstrated in numerous clinical trials (16,17).
In addition to the reduction of allergy-associated symptoms, AIT
serves a key role in preventing the emergence of novel allergies
and demonstrates long-term effects following the termination of AIT
treatment. Increasing evidence regarding the
immunological-associated effects of AIT, in particular the use of
allergen-derived B and T cell epitopes (18), has provided novel concepts for the
improvement of safe and effective vaccination (19). However, to the best of our
knowledge, the epitopes of Pla a 2 have not been investigated. In
the present study, B and T cell-associated epitopes of Pla a 2 were
identified using numerous immunoinformatics tools; these epitopes
may aid the development of potential peptide-based vaccine design
for pollen allergy.
Materials and methods
Sequence retrieval
The complete amino acid sequence of Pla a 2 was
acquired from the Nucleotide database of the National Center for
Biotechnology Information (http://www.ncbi.nlm.nih.gov/nucleotide/), accession
number Q6H9K0.
Patients and samples
A total of 10 adult individuals comprised two
groups: i) Five allergic subjects (aged between 20 and 43; 2 males
and 3 females; recruited in May 2016; diagnosis was established
based on clinical symptoms associated with allergic rhinitis during
the pollination season, a positive skin prick test result and a
seropositive IgE test to P. acerifolia pollen extract; and
ii) five healthy controls (age, 19–45; 2 males and 3 females;
recruited in May of 2016). Written informed consent for the use of
blood samples was obtained from the fingertips of all participants
prior to study entry according to The Declaration of Helsinki. The
study protocol was approved by the ethics committee of the First
Affiliated Hospital of Nanjing Medical University (Nanjing,
China).
Expression and purification of Pla a 2
allergen in E coli
The nucleotide sequence corresponding to the mature
Pla a 2 allergen (signal peptides were removed, which cannot be
excised by cell recognition and they may affect the proper folding
of proteins.) was synthesized by GenScript (Nanjing, China) and was
sub-cloned into a pET28a vector (Novagen, Madison, WI, USA)
using BamHI and XhoI sites, and was verified by
Sanger (20). Specifically, the
full-length Pla a 2 gene was amplified by reverse
transcription-quantitative polymerase chain reaction with a pair of
specific primers and the PCR product was cloned into the
pET28a vector using Tag enzymes (Takara Biotechnology Co.,
Ltd., Tokyo, Japan). The recombinant pET28a-Pla a 2 plasmid
was transformed into the ArcticExpress™ (DE3) RP Escherichia
coli host strain (21,22). Firstly, 1 µl pET28a-Pla a 2
plasmid was transformed into 100 µl ArcticExpress™ (DE3)
Escherichia coli cells and placed on ice for 30 min and then
heated in a water bath at 42°C for 45 sec, followed by an ice bath
for 2 min. A total of 500 µl LB medium without antibiotics was
added to the tube with shaking at 200 rpm for 1 h at 37°C.
Subsequently, 200 µl transformed cells were added to the LB agarose
solid state medium and placed at 37°C for 1 h. Finally, the tube
was cultured overnight at 37°C.
The positive clones were cultivated overnight in 3
ml lysogeny-broth kanamycin (concentration 0.05 g/ml) at 37°C until
an absorbance of 0.6–0.8 at 600 nm was attained. The culture was
induced with 1 mM isopropyl-b-D-thiogalactopyranoside and harvested
following incubation for 4 h at 37°C. Uninduced culture (subject to
the same conditions without IPTG) was used as a control. Expression
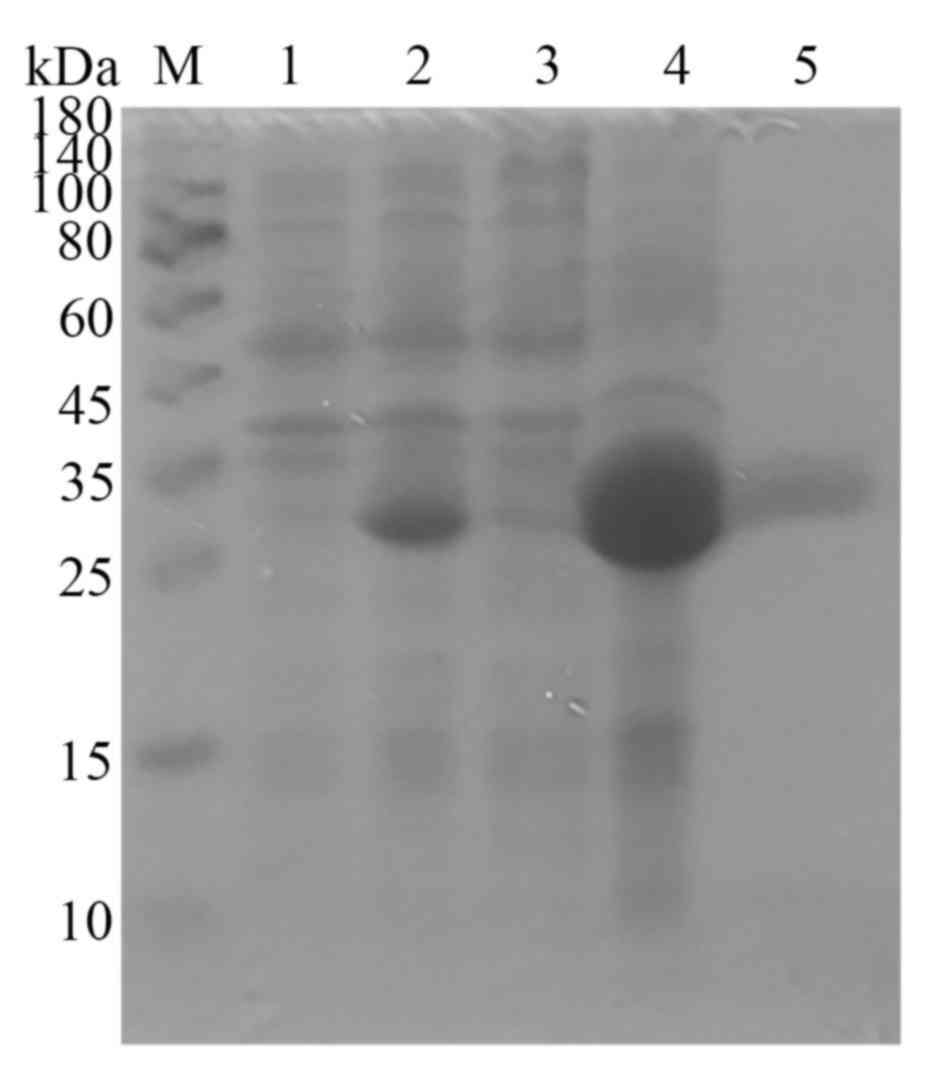
of recombinant Pla a 2 was confirmed using 12% SDS-PAGE. The cell
mass from 200 ml induced culture was resuspended in 20 mM Tris-HCl
buffer (pH 8.0). The cell suspension was sonicated at 40 kHz in 10
cycles of 4 sec pulse on and 8 sec pulse off at 4°C, and was
subsequently centrifuged at 10,000 × g for 30 min. Analysis
revealed that recombinant Pla a 2 was mainly located in the
supernatant (Fig. 1); purification
was conducted via Nickel affinity chromatography (GenScript)
(23,24). The washing buffer contained 100 mM
NaH2PO4, 20 mM Tris-HCl and 100 mM imidazole,
pH 8.0; eluting buffer contained 100 mM
NaH2PO4, 20 mM Tris-HCl and 250 mM imidazole,
pH 8.0 (25).
IgE binding activity of recombinant
Pla a 2 allergen
Purified Pla a 2 (5 µg) protein was separated by 12%
SDS-PAGE. The proteins were then transferred to polyvinylidene
difluoride (PVDF) membranes and were blocked with 5% skim milk at
room temperature for 2 h. Subsequently, PVDF membranes were
incubated with a serum mixture (1:40 in PBS) from six patients with
P. acerifolia pollen allergy, overnight at 4°C. The serum
mixture served as the primary antibody. Subsequently, IgE-allergen
complexes were detected using horseradish peroxidase-conjugated
goat anti-human IgE monoclonal antibody (cat no. A9667; diluted
1:3,000; Sigma-Aldrich, Merck KGaA, Darmstadt, Germany) at room
temperature; an ImageQuant LAS 4000 mini detection system (GE
Healthcare, Chicago, IL, USA) was employed for the detection of
these complexes. An enhanced chemiluminescence substrate including
luminol/enhancer solution and peroxide solution was used (1:1
mixed; Bio-Rad Laboratories, Inc., Hercules, CA, USA). Serum
mixture from six healthy individuals was applied as a negative
control and the experiment was repeated three times.
Homology modeling
SWISS-MODEL Repository is a database of
three-dimensional (3D) protein structure models generated by the
SWISS-MODEL homology modeling pipeline (26). The homologous templates suitable
for Pla a 2 were selected using the SWISS-MODEL server (http://swissmodel.expasy.org/) based on its complete
amino acid sequence (27). The
most appropriate template was retrieved from results of previous
analyses and was utilized for homology modeling.
B cell epitope prediction
Three immunoinformatics tools, including DNAStar
protean system (https://www.dnastar.com/t-allservices.aspx), BepiPred
1.0 server (http://www.cbs.dtu.dk/services/BepiPred/) and
Bioinformatics Predicted Antigenic Peptides (BPAP) system
(http://imed.med.ucm.es/Tools/antigenic.pl) were used
to predict the B cell-associated epitopes of Pla a 2 (28). The results obtained from the three
immunoinformatics tools were combined to produce the most
informative results (29). Four
properties (hydrophilicity, flexibility, accessibility and
antigenicity) of the amino acid sequence were selected as
parameters for epitope prediction in the DNAStar protean system
(30). The BepiPred 1.0 server and
the BPAP system required the amino acid sequence of Pla a 2.
Prediction of T cell epitopes
T cell epitope prediction was based on the
identification of bound peptide fragments on major
histocompatibility complex (MHC) complexes. The binding
significance of each peptide to the given MHC molecule is based on
the estimated strength of binding exhibited by a predicted nested
core peptide at a set threshold level. NetMHCII-2.2 (http://www.cbs.dtu.dk/services/NetMHCII/) (31) was employed to investigate human
leukocyte antigen (HLA)-DQ alleles. NetMHCIIpan-3.0 (http://www.cbs.dtu.dk/services/NetMHCIIpan/) was
applied to predict HLA-DR-based T cell epitopes (32). In the present study,
HLA-DQA10101-DQB10501, HLADQA-10501-DQB10201, HLA-DQA10501-DQB10301
and HLA-DQA10102-DQB10602 were used to predict HLA-DQ-based T cell
epitopes. Combining the four results indicated the most informative
HLA-DQ-associated T cell epitope result. Providing three epitopes
were revealed, the consensus would suggest an epitope. This method
was also applied to HLA-DR-based T cell epitope prediction.
HLA-DRB-10101, HLA-DRB30101, HLA-DRB40101 and HLA-DRB50101 were
used to predict HLA-DR-based T cell epitopes. The most informative
results were obtained by combining the results of the HLA-DQ- and
HLA-DR-based T cell epitope predictions. B and T cell epitopes
identified by computational tools were mapped into a linear
sequence and onto a 3D model of Pla a 2 to determine their position
(33).
Results
Expression and purification of Pla a 2
in E coli
P. acerifolia pollen Pla a 2 was subcloned
into a pET28a vector and transformed into BL21 (DE3) E.
coli host strain. It was demonstrated that Pla a 2 was
expressed mainly in the supernatant fraction (Fig. 1). The dissolved inclusion body of
Pla a 2 was purified via Nickel affinity chromatography. The purity
of the purified Pla a 2 was identified by SDS-PAGE. A single band
with an apparent molecular weight of ~25 kDa was observed (Fig. 1).
Immunoreactivity of Pla a 2 to
IgE
The ability of Pla a 2 to bind IgE within the sera
of patients associated with P. acerifolia pollen allergy was
determined by western blotting. Pla a 2 demonstrated positive IgE
reactivity to mixed serum of patients associated with P.
acerifolia pollen allergy; reactivity to healthy controls was
not observed, as presented in Fig.
2.
Homology modeling
Protein Data Bank accession no. 4c2I revealed a
marked sequence homology (25.22%) to the Pla a 2 allergen and was
used for homology modeling. The predicted B and T cell-associated
epitopes were superimposed on the surface of the Pla a 2 allergen
as presented in Fig. 3.
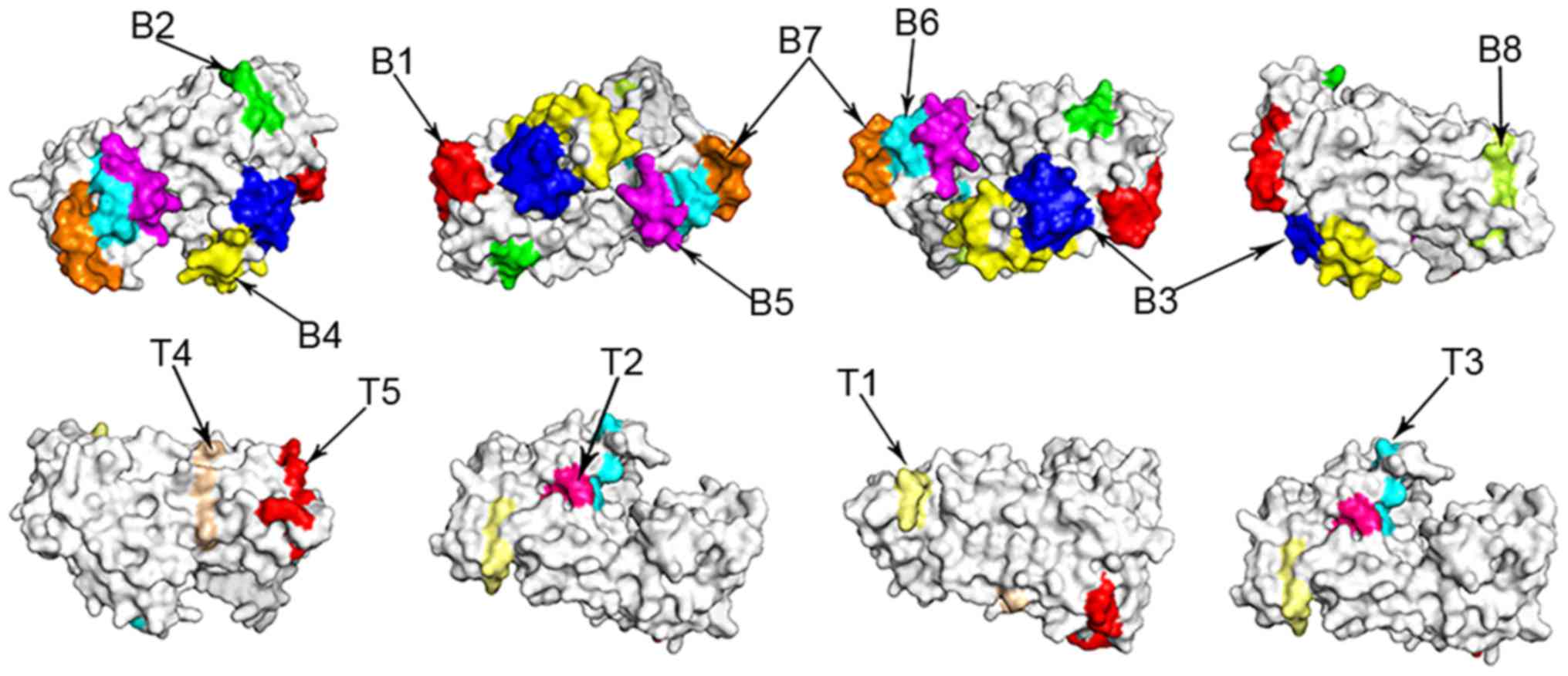 | Figure 3.Predicted B and T cell epitopes
identified on the surface of the 3-dimensional structure of the Pla
a 2 allergen. Specifically, B1 refers to B cell epitope 15–24, B2
refers to B cell epitope 60–66, B3 refers to B cell epitope 78–86,
B4 refers to B cell epitope 109–124, B5 refers to B cell epitope
232–240, B6 refers to B cell epitope 260–269, B7 refers to B cell
epitope 298–306, B8 refers to B cell epitope 315–322, T1 refers to
T cell epitope 62–67, T2 refers to T cell epitope 86–91, T3 refers
to T cell epitope 125–132, T4 refers to T cell epitope 217–222 and
T5 refers to T cell epitope 343–350. |
Prediction of B cell epitopes
The probability of epitope formation within the Pla
a 2 sequence is indicated by antigenic index. Regions of high
hydrophobicity are also associated with epitope identification. In
addition, surface accessibility and fragment flexibility are
important features for antigenic epitope prediction. From these
sequence properties, the final predicted epitope regions of Pla a 2
were 15–24, 60–66, 78–86, 105–111, 117–125, 232–240, 260–268,
298–306 and 315–322 (Table I)
using DNAstar. The predicted results of BepiPred 1.0 server were
2–9, 15–24, 36–46, 58–66, 74–87, 100–126, 170–179, 202–213,
219–226, 233–240, 260–273, 294–306, 317–323 and 350–373.
Additionally, the predicted results of BPAP system were 5–12,
23–30, 32–38, 40–47, 67–80, 88–100, 133–144, 153–162, 164–172,
180–189, 190–207, 210–220, 224–230, 244–250, 283–301, 303–309,
322–335, 356–365 and 368–374. Potential B cell epitopes of Pla a 2
were then selected from the results of these three tools. The most
informative result of the three immunoinformatics analyses led to
the prediction of eight peptides (15–24, 60–66, 78–86, 109–124,
232–240, 260–269, 298–306 and 315–322), which are presented in
Table II.
 | Table I.Predictions of B and T cell
epitopes. |
Table I.
Predictions of B and T cell
epitopes.
| Prediction | Tools | Location of the
prediction results |
|---|
| B cell epitope | DNAStar
protean | 15–24, 60–66,
78–86, 105–111, 117–125, 232–240, 260–268, 298–306, 315–322 |
|
| BepiPred | 2–9, 15–24, 36–46,
58–66, 74–87, 100–126, 170–179, 202–213, 219–226, 233–240, 260–273,
294–306, 317–323, 350–373 |
|
| BPAP | 5–12, 23–30, 32–38,
40–47, 67–80, 88–100, 133–144, 153–162, 164–172, 180–189, 190–207,
210–220, 224–230, 244–250, 283–301, 303–309, 322–335, 356–365,
368–374 |
| T cell epitope
(HLA-DQ) |
HLA-DQA10101-DQB10501 | 84–90, 146–151 |
|
|
HLA-DQA10102-DQB10602 | 2–4, 17–29, 46–52,
90–92, 121–125, 131–137, 141–152, 160–163, 182–192, 204–212,
234–240, 248–253, 277–281, 317–324 |
|
|
HLA-DQA10501-DQB10201 | 149–154,
279–283 |
|
|
HLA-DQA10501-DQB10301 | 2–5, 9–20, 23–35,
45–52, 63–74, 87–108, 161–169, 173–188, 198–206, 212–227, 243–252,
256–265, 291–303, 311–325, 334–353, 360–364 |
| T cell epitope
(HLA-DR) | DRB101:01 | 86–90, 127–133,
309–311 |
|
| DRB301:01 | 62–67, 77–83,
86–91, 125–130, 134–139 |
|
| DRB401:01 | 127–132 |
|
| DRB501:01 | 127–132, 295 |
| Table II.Predicted B and T cell epitopes of
Pla a 2. |
Table II.
Predicted B and T cell epitopes of
Pla a 2.
| Peptide | Type of
epitope | Position | Primary
sequence |
|---|
| P1 | B1 | 15–24 | DYGAKGAGDI |
| P2 | B2 | 60–66 | GPCKGSK |
| P3 | B3 | 78–86 | PADPSKFKS |
| P4 | B4 | 109–124 |
QGQTAWAKNNCDKNPN |
| P5 | B5 | 232–240 | GRYNNEKEV |
| P6 | B6 | 260–269 | KTWPNSPPGA |
| P7 | B7 | 298–306 | QCSRQAPSR |
| P8 | B8 | 315–322 | NNIRGTST |
| P9 | T1 | 62–67 | CKGSKI |
| P10 | T2 | 86–91 | SDGWVS |
| P11 | T3 | 125–132 | CKHAAMNL |
| P12 | T4 | 217–222 | QVNCGP |
| P13 | T5 | 343–350 | GEINLSYR |
Prediction of T cell epitopes
NetMHCII-2.2 and NetMHCIIpan-3.0 were used to
identify the T cell epitopes of Pla a 2. For HLA-DQ alleles, the
results of HLADQA10101-DQB10501, HLA-DQA10501-DQB10201,
HLADQA10501-DQB10301 and HLA-DQA10102-DQB10602 are presented in
Table I. For HLA-DR-based T cell
epitopes, the results of HLA- DRB1*01:01, HLA- DRB3*01:01, HLA-
DRB4*01:01, and HLA- DRB5*01:01 are also presented in Table I. As a result, Pla a 2 was
predicted to have five T cell epitope sequences, 62–67, 86–91,
125–132, 217–222 and 343–350, as presented in Table II.
Discussion
Despite improved understanding of the
pathophysiology of allergic diseases and advances in associated
pharmacological treatment, the prevalence has continued to increase
over the past several decades (34). P. acerifolia
pollen-associated allergies constitute ~50% of cases of allergy and
are frequently associated with severe allergic diseases, including
rhinitis and asthma. Characterization and identification of pollen
allergens will be beneficial to the diagnosis and treatment of
pollen-induced allergic illnesses (35).
P. acerifolia is an important cause of
pollinosis in many cities. The quantification of major allergens
has become a significant goal for the standardization of allergen
products intended for clinical use (36). Therefore, determination of the Pla
a 2 allergen is crucial for the development of P. acerifolia
pollen allergen-associated vaccines. The first objective of the
present study was to express and purify the Pla a 2 allergen. The
second objective was to predict the B and T cell epitopes of Pla a
2. It was demonstrated that Pla a 2 expression in the soluble form
was successful within E. coli. In addition, purified Pla a 2
allergen was analyzed by western blot analysis and was demonstrated
to possess the ability to bind IgE within the sera of patients with
P. acerifolia pollen allergy (8,37,38).
To further understand the structure and function of
Pla a 2, the sequence properties and 3D structure of Pla a 2 were
analyzed. Homology modeling was applied to investigate the 3D
structure of Pla a 2; the structure of 4C2I was the most
appropriate template with marked identity to Pla a 2. Prediction
using informatics tools is a familiar and useful method for
selecting epitopes from immunologically relevant proteins. In the
present study, three tools were employed (DNAStar protean system,
BepiPred 1.0 server and BPAP) to predict B cell epitopes. As a
result, eight peptides (15–24, 60–66, 78–86, 109–124, 232–240,
260–269, 298–306 and 315–322) were predicted as B cell epitopes;
however, further investigation in clinical samples is required. In
addition, NetMHCIIpan-3.0 and NetMHCII-2.2 were used to predict T
cell epitopes of Pla a 2-associated allergies; five potential T
cell epitope sequences were identified: 62–67, 86–91, 125–132,
217–222 and 343–350.
In conclusion, the predicted B and T cell epitopes
of Pla a 2 allergen may be used to benefit allergen immunotherapies
and reduce the frequency of allergic reactions. However, the
accuracy associated with such immunotherapy must be confirmed in
future experiments.
Acknowledgements
The present study was supported by grants from the
Special Fund for Forest-scientific Research in the Public Welfare
(grant. no. 201304103); the National Natural Science Foundation of
China (grant nos. 81571568, 31340073 and 81273274); Jiangsu
Province's Key Provincial Talents Program (grant. no. RC201170) and
the Priority Academic Program Development of Jiangsu Higher
Education Institutions.
References
|
1
|
Malmstrom K, Kaila M, Kajosaari M, Syvanen
P and Juntunen-Backman K: Fatal asthma in Finnish children and
adolescents 1976–1998: validity of death certificates and a
clinical description. Pediatr Pulmonol. 42:210–215. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sullivan PW, Smith KL, Ghushchyan VH,
Globe DR, Lin SL and Globe G: Asthma in USA: Its impact on
health-related quality of life. J Asthma. 50:891–899. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Moorman JE, Rudd RA, Johnson CA, King M,
Minor P, Bailey C, Scalia MR and Akinbami LJ: National surveillance
for asthma-United States, 1980–2004. MMWR Surveill Summ. 56:1–54.
2007.PubMed/NCBI
|
|
4
|
Snijders BE, Thijs C, Dagnelie PC, Stelma
FF, Mommers M, Kummeling I, Penders J, van Ree R and van den Brandt
PA: Breast-feeding duration and infant atopic manifestations, by
maternal allergic status, in the first 2 years of life (KOALA
study). J Pediatr. 151(347–351): 351. e341–342. 2007.
|
|
5
|
Hendaus MA, Jomha FA and Ehlayel M:
Allergic diseases among children: nutritional prevention and
intervention. Ther Clin Risk Manag. 12:361–372. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Warner JO, Kaliner MA, Crisci CD, Del
Giacco S, Frew AJ, Liu GH, Maspero J, Moon HB, Nakagawa T, Potter
PC, et al: Allergy practice worldwide: A report by the world
allergy organization specialty and training council. Int Arch
Allergy Immunol. 139:166–174. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Alcazar P, Garcia-Mozo H, Mdel Trigo M,
Ruiz L, Gonzalez-Minero FJ, Hidalgo P, de la Guardia Diaz C and
Galan C: Platanus pollen season in Andalusia (southern
Spain): Trends and modeling. J Environ Monit. 13:2502–2510. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fernandez-Gonzalez D, Gonzalez-Parrado Z,
Vega-Maray AM, Valencia-Barrera RM, Camazon-Izquierdo B, De Nuntiis
P and Mandrioli P: Platanus pollen allergen, Pla a 1:
Quantification in the atmosphere and influence on a sensitizing
population. Clinical and experimental allergy. J Br Soc Allergy
Clin Immunol. 40:1701–1708. 2010. View Article : Google Scholar
|
|
9
|
Lauer I, Miguel-Moncin MS, Abel T,
Foetisch K, Hartz C, Fortunato D, Cistero-Bahima A, Vieths S and
Scheurer S: Identification of a plane pollen lipid transfer protein
(Pla a 3) and its immunological relation to the peach
lipid-transfer protein, Pru p 3. Clin Exp Allergy. 37:261–269.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ibarrola I, Arilla MC, Martinez A and
Asturias JA: Identification of a polygalacturonase as a major
allergen (Pla a 2) from Platanus acerifolia pollen. J
Allergy Clin Immunol. 113:1185–1191. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wangorsch A, Larsson H, Messmer M,
Garcia-Moral A, Lauer I, Wolfheimer S, Schulke S, Bartra J, Vieths
S, Lidholm J, et al: Molecular cloning of plane pollen allergen Pla
a 3 and its utility as diagnostic marker for peach associated plane
pollen allergy. Clin Exp Allergy. 46:764–774. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Abramson MJ, Puy RM and Weiner JM:
Allergen immunotherapy for asthma. Cochrane Database Syst Rev
Cd001186. 2003. View Article : Google Scholar
|
|
13
|
Vadlamudi A and Shaker M: New developments
in allergen immunotherapy. Curr Opin Pediatr. 27:649–655. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Calderon MA, Alves B, Jacobson M, Hurwitz
B, Sheikh A and Durham S: Allergen injection immunotherapy for
seasonal allergic rhinitis. Cochrane Database Syst Rev Cd001936.
2007. View Article : Google Scholar
|
|
15
|
Dahl R, Kapp A, Colombo G, de Monchy JG,
Rak S, Emminger W, Rivas MF, Ribel M and Durham SR: Efficacy and
safety of sublingual immunotherapy with grass allergen tablets for
seasonal allergic rhinoconjunctivitis. J Allergy Clin Immunol.
118:434–440. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Madonini E and Musarra A: Allergen
immunotherapy in asthma: Current evidence and future requirements.
Eur Ann Allergy Clin Immunol. 43:103–110. 2011.PubMed/NCBI
|
|
17
|
Kim JM, Lin SY, Suarez-Cuervo C,
Chelladurai Y, Ramanathan M, Segal JB and Erekosima N:
Allergen-specific immunotherapy for pediatric asthma and
rhinoconjunctivitis: A systematic review. Pediatrics. 131:1155–67.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Thomas WR: The advent of recombinant
allergens and allergen cloning. J Allergy Clin Immunol.
127:855–859. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jutel M, Kosowska A and Smolinska S:
Allergen immunotherapy: Past, Present, and Future. Allergy Asthma
Immunol Res. 8:191–197. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shendure JA, Porreca GJ, Church GM,
Gardner AF, Hendrickson CL, Kieleczawa J and Slatko BE: Overview of
DNA sequencing strategies. Curr Protoc Mol Biol Chapter.
7:Unit7.12011.
|
|
21
|
Minato Y, Ueda T, Machiyama A, Shimada I
and Iwai H: Segmental isotopic labeling of a 140 kDa dimeric
multi-domain protein CheA from Escherichia coli by expressed
protein ligation and protein trans-splicing. J Biomol NMR.
53:191–207. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kumar S, Jain KK, Singh A, Panda AK and
Kuhad RC: Characterization of recombinant pectate lyase refolded
from inclusion bodies generated in E. coli BL21(DE3). Protein Expr
Purif. 110:43–51. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu Y, Sun X, Wang G, Tao A, Wu Y, Li M,
Shi H and Xie M: Expression, purification and identification of Pla
a1 in a codon-optimized Platanus pollen allergen. Mol Med
Rep. 12:2197–2202. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Y, Du WX, Fregevu C, Kothary MH,
Harden L and McHugh TH: Expression, purification, and
characterization of almond (Prunus dulcis) allergen Pru du 4. J
Agric Food Chem. 62:12695–12700. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tong X, Guo M, Jin M, Chen H, Li Y and Wei
JF: In silico epitope prediction, expression and functional
analysis of Per a 10 allergen from the American cockroach. Int J
Mol Med. 38:1806–1814. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kiefer F, Arnold K, Kunzli M, Bordoli L
and Schwede T: The SWISS-MODEL Repository and associated resources.
Nucleic Acids Res. 37:D387–392. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Arnold K, Bordoli L, Kopp J and Schwede T:
The SWISS-MODEL workspace: A web-based environment for protein
structure homology modelling. Bioinformatics. 22:195–201. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang H, Chen H, Jin M, Xie H, He S and Wei
JF: Molecular cloning, expression, IgE binding activities and in
silico epitope prediction of Per a 9 allergens of the American
cockroach. Int J Mol Med. 38:1795–1805. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang X and Yu X: An introduction to
epitope prediction methods and software. Rev Med Virol. 19:77–96.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zheng LN, Lin H, Pawar R, Li ZX and Li MH:
Mapping IgE binding epitopes of major shrimp (Penaeus
monodon) allergen with immunoinformatics tools. Food Chem
Toxicol. 49:2954–2960. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nielsen M and Lund O: NN-align. An
artificial neural network-based alignment algorithm for MHC class
II peptide binding prediction. BMC Bioinformatics. 10:2962009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Karosiene E, Rasmussen M, Blicher T, Lund
O, Buus S and Nielsen M: NetMHCIIpan-3.0, a common pan-specific MHC
class II prediction method including all three human MHC class II
isotypes, HLA-DR, HLA-DP and HLA-DQ. Immunogenetics. 65:711–724.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen H, Yang HW, Wei JF and Tao AL: In
silico prediction of the T-cell and IgE-binding epitopes of Per
a 6 and Bla g 6 allergens in cockroaches. Mol Med Rep.
10:2130–2136. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Goldblum RM, Ning B, Judy BM, Holthauzen
LM, van Bavel J, Kamijo A and Midoro-Horiuti T: A single mouse
monoclonal antibody, E58 modulates multiple IgE epitopes on group 1
cedar pollen allergens. Mol Immunol. 74:106–112. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li X, Yang HW, Chen H, Wu J, Liu Y and Wei
JF: In silico prediction of T and B cell epitopes of Der f
25 in Dermatophagoides farinae. Int J Genom. 2014:4839052014.
|
|
36
|
van Ree R: The CREATE project: EU support
for the improvement of allergen standardization in Europe. Allergy.
59:571–574. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Asero R, Mistrello G, Amato S and Villalta
D: Monosensitization to a novel plane pollen allergen. Eur Ann
Allergy Clin Immunol. 44:167–169. 2012.PubMed/NCBI
|
|
38
|
Enrique E, Alonso R, Bartolome B, San
Miguel-Moncin M, Bartra J, Fernandez-Parra B, Tella R, Asturias JA,
Ibarrola I, Martínez A, et al: IgE reactivity to profilin in
Platanus acerifolia pollen-sensitized subjects with
plant-derived food allergy. J Investig Allergol Clin Immunol.
14:335–342. 2004.PubMed/NCBI
|