Cysteine-rich angiogenic inducer 61 (CCN1/Cyr61) was
originally revealed to act as a growth factor-inducible
immediate-early gene, present in serum and platelet-derived growth
factor-stimulated mouse BALB/c 3T3 cells (1). As a member of CCN family, it was
first cloned by O'Brien in 1990 via differential hybridization
screening of a cDNA library from BALB/c 3T3 clone A31 cells
(2). The CCN family is composed of
six members including CCN1/Cyr61, connective tissue growth factor,
nephroblastoma overexpressed, Wnt-1-induced secreted protein
(WISP)-1, WISP-2 and WISP-3 (3).
CCN1/Cyr61 is a basic secreted protein localized on the cell
surface, cell interior and the extracellular matrix (ECM).
CCN1/Cyr61 targets cells including fibroblasts, epithelial cells,
endothelial cells, smooth muscle cells and neurons through
integrins and heparin sulfate proteoglycan (HSPG) receptors.
CCN1/Cyr61 exhibits a variety of functions in differing target
cells. The biological properties of CCN1/Cyr61 in the regulation of
cell survival, proliferation, differentiation, migration, adhesion
and synthesis of ECM have been demonstrated to be important in the
progression of embryogenesis and tumorigenesis in the female
reproductive system.
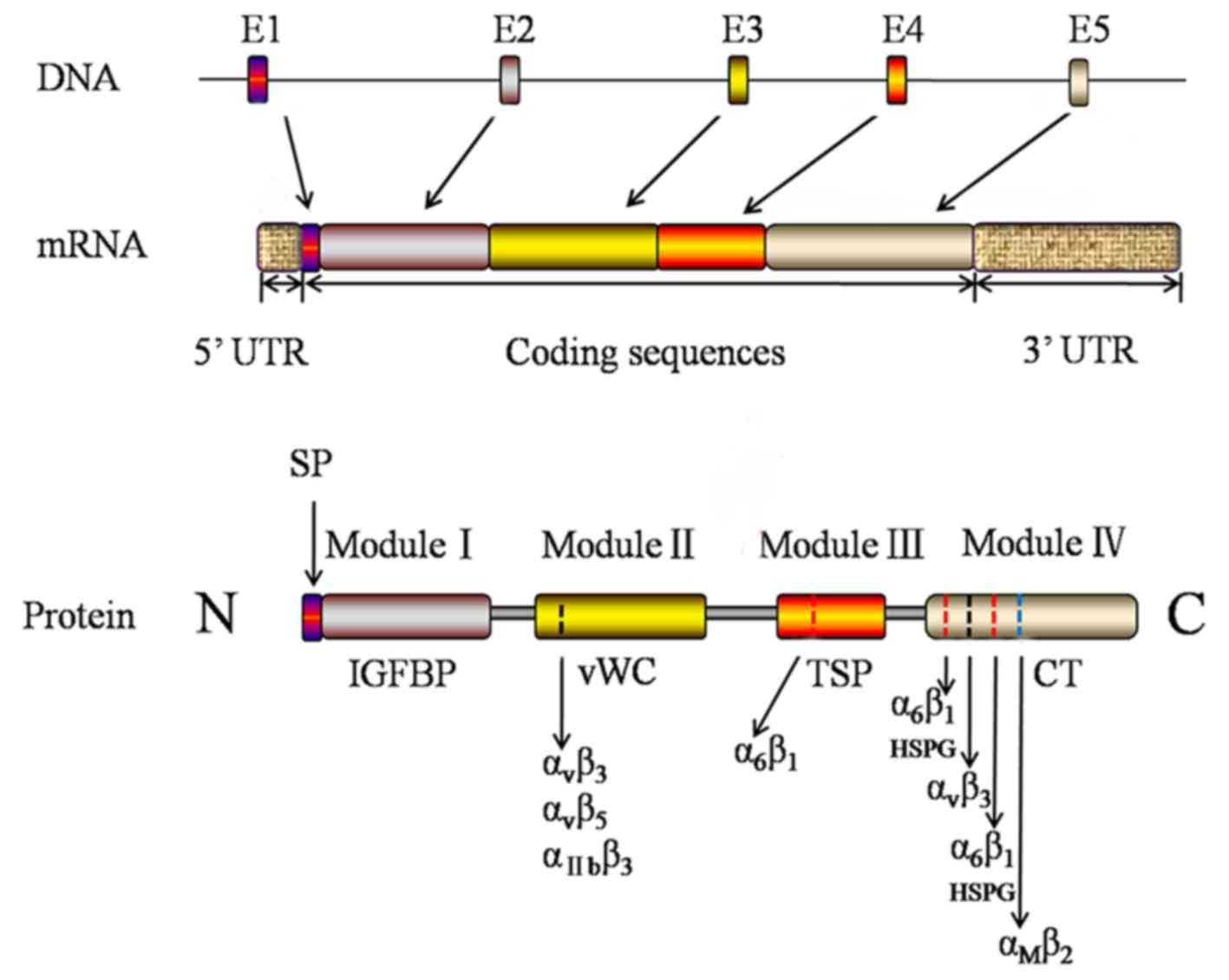
Sequence analysis revealed the presence of a mosaic
structure to the CCN1/Cyr61 protein, including four highly
conserved modules (3). As
presented in Fig. 1, from the N-
to the C-terminal, the four structurally distinct modules of
CCN1/Cyr61 are insulin-like growth factor binding protein
(IGFBP)-like domain, von Willebrand factor type C repeats
(vWC) domain, thrombospondin type 1 (TSP-1) repeat homology domain,
and the C-terminal domain that contains a cysteine-knot (8). Currently, the four structurally
distinct modules of CCN1/Cyr61 have been important targets to
explore the biological function of the protein. Module I is the
binding domain of IGFBP which consists of 68 amino acids and shares
homology with the cysteine-rich N-terminal region of traditional
IGFBPs. A previous study suggested that CCN1/Cyr61 demonstrates a
low affinity to IGFs (9). Module
II consists of 68 amino acids and includes a vWC domain which
exists in vWC factor, mucoprotein, thrombospondin and collagenous
fiber. Module II may additionally have a role in oligomerization
(3). Module III contains a motif
firstly identified in TSP-1 repeat homology and subsequently
observed in various extracellular proteins. TSP-1 homology is
involved in binding to soluble and matrix sulfated glycoconjugates
(10). A previous study revealed
that Module III contains cell attachment sites (11). Module IV, the C-terminal domain, is
present in PDGF, nerve growth factor and TGF-β. It has been
hypothesized that Module IV may be involved with cell surface
receptors (12). The modular
structural organization of the CCN1/Cyr61 protein suggests that its
diverse biological functions may result from the combined actions
of individual modules, either acting independently or in
conjugation. The interaction of the module and different factors
endows CCN1/Cyr61 with complex biological effects in the organism
tissues, cell environment and at all developmental stages.
CCN1/Cyr61 coordinates the essential activities of
life and demonstrates a diverse functional pattern in various
pathological and physiological conditions, in addition to its role
in various tissues and cells. CCN1/Cyr61 primarily binds to
integrins and HSPGs in a cell-type specific manner, and thus
mediates various biological activities via these interactions.
CCN1/Cyr61 has been reported to mediate numerous
cellular activities, including cell adhesion, migration,
proliferation and apoptosis. CCN1/Cyr61 is a cell adhesion protein,
which is associated with the adhesion activities of endothelial
cells, fibroblasts cells, smooth muscle cells, monocytes and
platelets, through binding to particular integrins (Table I) (13–18).
Integrin αvβ3, αvβ5,
αIIbβ3, α6β1, and
αMβ2 serve as adhesion receptors for
CCN1/Cyr61 on different cell types which are summarized in Table I. It has previously been
demonstrated that cell adhesive properties of CCN1/Cyr61 in human
umbilical vein endothelial cells are mediated through interaction
with integrin αvβ3 (14). In addition, it was demonstrated
that CCN1/Cyr61 mediates endothelial cell adhesion through
interaction with α6β1 (13). CCN1/Cyr61 promotes cell adhesion
indirectly by altering the structure of the ECM protein. Kireeva
et al (19) reported that
CCN1/Cyr61 mediates cell and ECM and cell-cell adhesion by
connecting with ECM and the cell membrane.
Cell migration is a complex biological process which
involves a series of stimulating factors, signaling pathways and
molecular mechanisms. It has been demonstrated that CCN1/Cyr61
regulates cell migration of fibroblasts and endothelial cells.
Zhang et al (20) firstly
verified that PDGF-induced CCN1/Cyr61 exhibits a key role in
regulating smooth muscle cell migration by connecting intracellular
PDGF-extracellular signal-regulated kinase and jun N-terminal
kinase signals with integrin/focal adhesion kinase (FAK) signaling.
Fibroblasts may suppress the inflammatory response, and may be
considered as a novel target for the therapy of inflammation. Huang
et al (21) demonstrated
that CCN1/Cyr61 markedly facilitates rheumatoid arthritis (RA)
fibroblast-like synoviocyte invasion and migration and may act as a
promising therapeutic target for RA treatment. Wei et al
(22) suggested that CCN1/Cyr61
expression is significantly associated with the metastasis of
gastric cardia adenocarcinoma (GCA). It has therefore been
hypothesized that CCN1/Cyr61 may act as a metastatic biomarker for
evaluating the prognosis of GCA and a potential molecular target
for anti-metastatic therapy of GCA.
In addition to promoting cell adhesion and
migration, CCN1/Cyr61 regulates cell survival and apoptosis. The
overexpression of CCN1/Cyr61 activates the nuclear factor (NF)-κB
pathway and induces hepatic stellate cell apoptosis through
endoplasmic reticulum stress and initiation of the unfolded protein
response (23). Conversely,
CCN1/Cyr61 expression promotes kidney tubular epithelial cell
proliferation and cell apoptosis is inhibited (21). The functions of CCN1/Cyr61 in
promoting cell survival or inducing apoptosis in different
cell-types is important to elucidate its role in tissue
reconstruction.
Previous studies have suggested CCN1/Cyr61 is a
primary physiological mediator of angiogenesis. CCN1/Cyr61
modulates vascular formation in vitro directly through
binding to αvβ3 to enhance the endothelial
cell adhesion, migration, proliferation and microtubule formation,
or indirectly by accommodating the activity of VEGF/VEGF receptor.
You et al (24) suggested
that CCN1/Cyr61 induces the expression of monocyte chemoattractant
protein-1 to mediate angiogenesis via the
ανβ3 integrin, FAK,
phosphoinositide-3-kinase/Akt Serine/Threonine Kinase 1 (Akt) and
the IKB kinase signaling pathways in monkey chorioretinal vessel
endothelial cells RF/6A (24). A
previous study investigating pancreatic neuroendocrine tumors
revealed a positive correlation between CCN1/Cyr61 expression
levels and tumor angiogenesis, which suggests an ancillary function
of CCN1/Cyr61 in vascular development (25). CCN1/Cyr61 was revealed to be
expressed in the placenta, cardiovascular, skeletal and nervous
systems during embryonic development. CCN1/Cyr61 expression levels
are additionally significantly decreased in pre-eclamptic placentas
(26). This suggests that
CCN1/Cyr61 is involved in placental angiogenesis and may be
important in the pathogenesis of pre-eclampsia.
CCN1/Cyr61-deficient mice lead to a placental vasculogenesis
deficiency with incomplete vascular nets. A previous study reported
~30% of mice died from the chorioallantoic junction defect in the
early stage of pregnancy and the others died in the middle of
pregnancy due to placental vascular insufficiency and extensive
loss in the vasculature integrity (27). Overall, these results demonstrate
that CCN1/Cyr61 is critically involved in developmental
angiogenesis.
CCNl/Cyr61 is important in cartilage cell
differentiation and chondrogenesis. The expression of CCNl/Cyr61 is
closely associated with angiogenesis and the process of the
differentiation from marrow stromal cells (MSC) to chondrocytes
during embryogenesis (28). In
adult cartilage, Chijiiwa et al (29) demonstrated that CCNl/Cyr61 is
involved in chondrocyte cloning in osteoarthritic cartilage by
inhibiting the activity of ADAM metallopeptidase with
thrombospondin type 1 motif 4 (ADAMTS; aggrecanase-1). Conversely,
the emergence of CCNl/Cyr61 may lead to the replacement of bone
tissue by cartilage through inducing angiogenesis. A previous study
revealed that CCNl/Cyr61 may be associated with the induction of a
passage-dependent decrease in chondrogenic differentiation and a
passage-dependent increase in osteogenic potential of
tonsil-derived MSC (30).
Mammalian reproduction is a complex process
involving ovulation, fertilization, embryo development and
implantation and the failure of any process may result in
sterility, malformations or adverse pregnancy outcomes. Previous
studies demonstrated that CCNl/Cyr61 is expressed on the corpus
luteum (CL) (31), conceptus and
endometrium in early pregnancy (32), trophoblastic origin of the placenta
(33), midterm fetal smooth muscle
vessel walls of the arterial circulatory system (34), endothelial cells of vessels
surrounding the cloaca, embryonic hippocampal neurons (35) and mesodermal and ectodermal
mesenchymal cells (28). It
contributes to the process of embryogenesis including the
implantation (36), fetal
neovascularization (34), fetal
brain development (35), fetal
cartilaginous development (28)
and fetal-maternal cross-talk (32). The abnormal expression of
CCNl/Cyr61 may result in pre-eclampsia, epispadias, embryonic death
resulting from disability of allantoic fusion and the disturbance
of vascular dysplasia (34,37).
CL, which is a glandular-like structure rich in
blood vessels and exhibits a key role in mammalian reproduction.
The development of the follicle into the CL involves
neovascularization, and previous studies have demonstrated that
CCNl/Cyr61 may mediate this process (Fig. 2). Zhang et al (31) suggested that CCNl/Cyr61 is
expressed in the CL of cows and the expression is upregulated at
days 4, 6, 10 and 16 of CL. It was therefore hypothesized that the
upregulation may be associated with the angiogenesis of the CL. A
further study conducted by Romereim et al (38) demonstrated that the CCNl/Cyr61 gene
is highly expressed in small luteal cells isolated from cows. These
findings support the hypothesis that CCNl/Cyr61 is important in the
development of the CL. However, the expression pattern, regulation
and function of CCNl/Cyr61 in human CLs still remains unclear and
requires further investigation.
Implantation is an important process during the
early development of embryos. CCNl/Cyr61 has previously been
suggested to act as a mediator for embryo implantation. It has
previously been detected to be upregulated in the invasive
extravillous trophoblasts and unclosed endometrial luminal
epithelium surrounding the embryo (39). Pre-eclampsia is one of the common
complications of pregnancy, which originates in the placenta and
results in complication of maternal and fetal health (40). The hypothesis of the etiology has
been recognized as insufficient invasion of trophoblast cells into
the decidua and the obstruction of maternal spiral artery
remodeling (41). This leads to an
inadequate blood flow in the placenta and may result in the
inhibition of placental development and intrauterine growth
restrictions (42). Recombinant
CCNl/Cyr61 protein-stimulated trophoblast cells demonstrate a
stronger invasion ability compared with cells without the stimulus
(43). Chen et al (33) revealed that CCNl/Cyr61 mRNA
expression levels in placentas of pre-eclampsia cases are decreased
compared with than those in normal pregnancies. Furthermore,
Kipkeew et al (36)
indicated that CCNl/Cyr61 and CCN3 promote the migration capability
of trophoblast cells by activating FAK and Akt kinase. This
evidence verified the hypothesis of a negative correlation between
the expression of CCNl/Cyr61 and pre-eclampsia. These findings may
act as a research foundation for the early diagnosis and prediction
of pre-eclampsia through screening for expression of CCNl/Cyr61.
Zhang et al (44) suggested
that that micro (mi)RNA 155 contributes to pre-eclampsia
development by downregulating CCNl/Cyr61. MiRNAs are relatively
stable and the identification of circulating miRNAs involved in
promoting the development of pre-eclampsia by regulating the
expression of CCNl/Cyr61 in the plasma of pregnant woman, may aid
in the clinical prediction of pre-eclampsia.
CCNl/Cyr61 is additionally important in the
maintenance of the pregnancy and the embryonic development. Klein
et al (32) demonstrated
that CCNl/Cyr61 is co-expressed on the conceptus and endometrium in
the early stage of pregnancy and involved the maintenance of the
early pregnancy. It contributes to the cross-talk of the conceptus
and endometrium through affecting the proliferation and
differentiation of endometrial and trophectoderm cells and
stimulation of endometrial-angiogenesis. Furthermore, previous
studies have demonstrated that CCNl/Cyr61 regulates
neovascularization and matrix remodeling in the process of
embryogenesis. CCNl/Cyr61 gene knock-out mice often suffer from
vascular anomalies of placenta and embryonic fatalities (34). The expression of CCNl/Cyr61 may be
detected in endothelial cells of vessels surrounding the cloaca and
the umbilical cord on gestational days 10 and 11.5 in mice, which
suggests it may be involved in the development of human epispadias
(37). In addition, the expression
of CCNl/Cyr61 in fetal hippocampal neurons has been demonstrated to
be increased compared with mature hippocampal neurons, which
suggests it may exhibit an important role during development of the
embryonic brain (35). Overall,
CCNl/Cyr61 is important role in the female reproductive system.
It was previously demonstrated that CCNl/Cyr61 acts
as a tumor-suppressor or promoter in different types of tumors
depending on the complex external environment (45–47).
CCNl/Cyr61 is involved in the occurrence and development of tumors
through its role in tumor angiogenesis, tumor cell proliferation
and apoptosis and tumor metastasis (25,47–49).
An overview of the expression and function of CCNl/Cyr61 in female
reproductive tumors is presented in Table II (50–61).
Further research and understanding of the function of CCNl/Cyr61 in
different types of cancer may provide a promising basis for cancer
prognosis and therapy.
Uterine leiomyoma (UL) is the most common benign
tumor of the female reproductive system, which is hormone dependent
and results from smooth muscle cell proliferation (62). Molecular genetic studies have
provided compelling evidence that abnormally expressed genes have
an important influence on UL occurrence (63,64).
CCNl/Cyr61 expression in UL is downregulated when compared with
matched normal uterine smooth muscle cells, suggesting it may
exhibit a tumor-suppressor role (50). Di Tommaso et al (51) revealed that estrogen upregulates
the expression of the CCNl/Cyr61 gene in myometrium tissue, however
has no influence on the expression of the CCNl/Cyr61 gene in
fibroid tissue, detected via gene expression analysis. The results
indicated that the expression of CCNl/Cyr61 may not be regulated by
the estrogen receptor in uterine fibroids. Fundamental research was
conducted by Wallace et al (52) to determine if hypoxia induced
CCNl/Cyr61 secretion through the Endothelin-A (ETA)
receptor in UL. However, the results identified that blockade of
the ETA receptor demonstrated no significant influence on the
increase of CCNl/Cyr61 expression under normoxic or hypoxic
conditions and the secretion of CCNl/Cyr61 may not be regulated by
the ETA receptor in UL. The exact mechanism still remains unclear
and further investigation is required in the future.
Endometriosis (EMT) is one of the most common
diseases of obstetrics and gynecology, and commonly associates with
infertility in women of child-bearing age (65–67).
The specific pathogenesis of EMT remains to be fully elucidated.
Although EMT is a frequently occurring benign disease, it is
characterized by malignant behaviors including recurrence and
metastasis (68,69). At present, various
pharmacotherapies and other methods have been proposed to treat EMT
(70,71). However, their usage has been
limited due to strong and obvious adverse effects (65,72).
It was previously demonstrated that the expression of the
CCNl/Cyr61 gene is controlled by estrogen and is enhanced in
endometria of women suffering from EMT and in endometriotic
lesions, suggesting that CCNl/Cyr61 may act as a useful maker gene
for EMT (53). A further study
identified that estrogen promotes the expression of the CCNl/Cyr61
gene during the early stages of EMT-like establishment and the
CCNl/Cyr61 gene contributes to regulation of cell proliferation and
the generation of vasculature in the development of lesions, using
the conditional knockout mice lacking uterine CCNl/Cyr61 (54). Furthermore, Gashaw et al
(73) demonstrated that the
expression level of CCNl/Cyr61 is upregulated by hypoxia-inducible
factor 1-α, through use of the benign endometrial cell line. The
study suggests that targeting CCNl/Cyr61 signaling during the early
stages of lesion establishment may provide a novel treatment method
for EMT.
Endometrial cancer (EC) is a common malignant
gynecological tumor, however non-invasive diagnostic tools for
early and accurate diagnosis are still limited (74). Hormonal dysregulation is believed
to contribute to the etiology of EC (75). Previous studies demonstrated that
long-term replacement estrogen therapy is associated with a type I
EC and progestational hormone is not sufficient to prevent this
(56). Estrogen may upregulate the
expression of CCNl/Cyr61 and current data indicate that there is a
significant increase of CCNl/Cyr61 expression in the endometrium of
women with polycystic ovarian syndrome and premalignant lesions.
Hence, measures to detect the expression of CCNl/Cyr61 may result
in its use as an early diagnostic marker of type I EC cancer
However, Chien et al (55)
demonstrated that the expression level of Cyr61 is decreased in
endometrial tumors compared with normal endometrium. At present,
little is known regarding the antithetical function of CCNl/Cyr61
in EC. Previous findings from Witek et al (76) revealed that CCNl/Cyr61 genes
expressed in all cancer grades and their expressions levels are
potentially correlated with the survival of patients. Therefore,
detecting the expression level of CCNl/Cyr61 may act as a novel
non-invasive diagnostic tool for early and accurate diagnosis of
EC.
Cervical cancer is one of the most common
gynecological tumors occurring in women worldwide, with >190,000
newly diagnosed cases per year (77). The human papillomavirus infection
is an important factor in the development of cervical cancer, and
vaccination against it provides an effective way to prevent
development of cervical cancer. However, the pathogenesis
underlying cervical cancer remains unknown, and its incidence is
increasing each year. Xie et al (58) demonstrated that CCNl/Cyr61 gene
expression is downregulated in cervical cancer compared with normal
tissues and its expression level is inversely correlated with
miR-205 expression. miR-205 has been reported to contribute to the
development of cervical cancer and regulate cell proliferation and
migration in human cervical cancer cells. The identified inverse
expression pattern between CCNl/Cyr61 and miR-205 suggests that
CCNl/Cyr61 may be one of the target genes of miR-205. In addition,
by performing genome-wide expression analysis of miRNAs, in
addition to investigation of mRNAs in cervical cancer, Joy et
al (57) demonstrated that
CCNl/Cyr61 is a key gene involved in vascularization of tumors and
suggests its expression level is negatively associated with the
level of hsa-miR-221. The roles of CCNl/Cyr61 in human cervical
cancer may provide novel insights into carcinogenesis and the
clinical diagnosis.
Ovarian cancer is the leading cause of gynecological
malignancy-associated mortalities worldwide and the symptoms are
insidious in onset (78,79). The search for novel biomarkers for
early diagnosis in ovarian carcinoma may provide effective medical
support. Lee et al (60)
demonstrated that CCNl/Cyr61 promotes ovarian cancer cell
proliferation and inhibits apoptosis by regulating p53 and NF-κB
expression through the phosphoinositide-3-kinase/AKT
Serine/Threonine kinase/mechanistic target of rapamycin signaling
pathways. Bartel et al (59) revealed that CCNl/Cyr61 is
preferentially expressed in high grade serous carcinomas. It was
demonstrated that CCNl/Cyr61 expression is significantly
upregulated in ovarian epithelial carcinoma tissue compared with
benign ovarian tumor tissue samples and the levels of CCNl/Cyr61
are associated with lymph node metastases (61). CCNl/Cyr61 may therefore be useful
in targeted diagnosis and therapy in ovarian epithelial
carcinoma.
The present review systematically clarified the
biological functions of CCN1/Cyr61 and its associated roles in the
female reproductive system, including embryogenesis and
tumorigenesis. CCNl/Cyr61 may regulate numerous cellular
activities, angiogenesis and chondrogenesis. It is important in the
reproduction and the occurrence and development of tumors in the
reproductive system. However, the underlying mechanism that results
in CCNl/Cyr61 acting as a tumor-suppressor or tumor-promoter in
different types reproductive system tumors, remains to be fully
elucidated, due to the complicated external environment, including
complex crosstalk of intracellular molecules and multi-module
activated signal transduction pathways.
The present review was supported by the National
Natural Science Foundation of China (grant nos. 81671473 and
81602728), the Youth Talent's Project of Jiangsu Province (grant
no. QNRC2016164) and Provincial Foundation of Jiangsu Province
(grant nos. BK20131151 and BE2016633).
|
1
|
Lau LF and Nathans D: Identification of a
set of genes expressed during the G0/G1 transition of cultured
mouse cells. EMBO J. 4:3145–3151. 1985.PubMed/NCBI
|
|
2
|
O'Brien TP, Yang GP, Sanders L and Lau LF:
Expression of cyr61, a growth factor-inducible immediate-early
gene. Mol Cell Biol. 10:3569–3577. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bork P: The modular architecture of a new
family of growth regulators related to connective tissue growth
factor. FEBS Lett. 327:125–130. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jay P, Bergé-Lefranc JL, Marsollier C,
Méjean C, Taviaux S and Berta P: The human growth factor-inducible
immediate early gene, CYR61, maps to chromosome 1p. Oncogene.
14:1753–1757. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Borkham-Kamphorst E, Schaffrath C, Van de
Leur E, Haas U, Tihaa L, Meurer SK, Nevzorova YA, Liedtke C and
Weiskirchen R: The anti-fibrotic effects of CCN1/CYR61 in primary
portal myofibroblasts are mediated through induction of reactive
oxygen species resulting in cellular senescence, apoptosis and
attenuated TGF-β signaling. Biochim Biophys Acta. 1843:902–914.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen Y and Du XY: Functional properties
and intracellular signaling of CCN1/Cyr61. J Cell Biochem.
100:1337–1345. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chin LH, Hsu SP, Zhong WB and Liang YC:
Involvement of cysteine-rich protein 61 in the epidermal growth
factor-induced migration of human anaplastic thyroid cancer cells.
Mol Carcinog. 55:622–632. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rachfal AW and Brigstock DR: Structural
and functional properties of CCN proteins. Vitam Horm. 70:69–103.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Berschneider B and Königshoff M: WNT1
inducible signaling pathway protein 1 (WISP1): A novel mediator
linking development and disease. Int J Biochem Cell Biol.
43:306–309. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Choi J, Lin A, Shrier E, Lau LF, Grant MB
and Chaqour B: Degradome products of the matricellular protein CCN1
as modulators of pathological angiogenesis in the retina. J Biol
Chem. 288:23075–23089. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin J, Huo R, Wang L, Zhou Z, Sun Y, Shen
B, Wang R and Li N: A novel anti-Cyr61 antibody inhibits breast
cancer growth and metastasis in vivo. Cancer Immunol Immunother.
61:677–687. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qin Z, Fisher GJ and Quan T: Cysteine-rich
protein 61 (CCN1) domain-specific stimulation of matrix
metalloproteinase-1 expression through aVb3 integrin in human skin
fibroblasts. J Biol Chem. 288:12386–12394. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Heng EC, Huang Y, Black SA Jr and Trackman
PC: CCN2, connective tissue growth factor, stimulates collagen
deposition by gingival fibroblasts via module 3 and alpha6- and
beta1 integrins. J Cell Biochem. 98:409–420. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kireeva ML, Lam SC and Lau LF: Adhesion of
human umbilical vein endothelial cells to the immediate-early gene
product Cyr61 is mediated through integrin alphavbeta3. J Biol
Chem. 273:3090–3096. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen N, Chen CC and Lau LF: Adhesion of
human skin fibroblasts to Cyr61 is mediated through integrin alpha
6beta 1 and cell surface heparan sulfate proteoglycans. J Biol
Chem. 275:24953–24961. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Grzeszkiewicz TM, Lindner V, Chen N, Lam
SC and Lau LF: The angiogenic factor cysteine-rich 61 (CYR61, CCN1)
supports vascular smooth muscle cell adhesion and stimulates
chemotaxis through integrin alpha(6)beta(1) and cell surface
heparan sulfate proteoglycans. Endocrinology. 143:1441–1450. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schober JM, Chen N, Grzeszkiewicz TM,
Jovanovic I, Emeson EE, Ugarova TP, Ye RD, Lau LF and Lam SC:
Identification of integrin alpha(M)beta(2) as an adhesion receptor
on peripheral blood monocytes for Cyr61 (CCN1) and connective
tissue growth factor (CCN2): Immediate-early gene products
expressed in atherosclerotic lesions. Blood. 99:4457–4465. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jedsadayanmata A, Chen CC, Kireeva ML, Lau
LF and Lam SC: Activation-dependent adhesion of human platelets to
Cyr61 and Fisp12/mouse connective tissue growth factor is mediated
through integrin alpha(IIb)beta(3). J Biol Chem. 274:24321–24327.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kireeva ML, Mo FE, Yang GP and Lau LF:
Cyr61, a product of a growth factor-inducible immediate-early gene,
promotes cell proliferation, migration, and adhesion. Mol Cell
Biol. 16:1326–1334. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang F, Hao F, An D, Zeng L, Wang Y, Xu X
and Cui MZ: The matricellular protein Cyr61 is a key mediator of
platelet-derived growth factor-induced cell migration. J Biol Chem.
290:8232–8242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang TL, Mu N, Gu JT, Shu Z, Zhang K,
Zhao JK, Zhang C, Hao Q, Li WN, Zhang WQ, et al: DDR2-CYR61-MMP1
signaling pathway promotes bone erosion in rheumatoid arthritis
through regulating migration and invasion of fibroblast-like
synoviocytes. J Bone Miner Res. 32:407–418. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wei J, Yu G, Shao G, Sun A, Chen M, Yang W
and Lin Q: CYR61 (CCN1) is a metastatic biomarker of gastric cardia
adenocarcinoma. Oncotarget. 7:31067–31078. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Borkham-Kamphorst E, Steffen BT, Van de
Leur E, Haas U, Tihaa L, Friedman SL and Weiskirchen R: CCN1/CYR61
overexpression in hepatic stellate cells induces ER stress-related
apoptosis. Cell Signal. 28:34–42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
You JJ, Yang CH, Yang CM and Chen MS:
Cyr61 induces the expression of monocyte chemoattractant protein-1
via the integrin avb3, FAK, PI3K/Akt, and NF-kB pathways in retinal
vascular endothelial cells. Cell Signal. 26:133–140. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang YT, Lan Q, Ponsonnet L, Blanquet M,
Christofori G, Zaric J and Rüegg C: The matricellular protein CYR61
interferes with normal pancreatic islets architecture and promotes
pancreatic neuroendocrine tumor progression. Oncotarget.
7:1663–1674. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li X, Li C, Dong X and Gou W: MicroRNA-155
inhibits migration of trophoblast cells and contributes to the
pathogenesis of severe preeclampsia by regulating endothelial
nitric oxide synthase. Mol Med Rep. 10:550–554. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lau LF: CCN1 and CCN2: Blood brothers in
angiogenic action. J Cell Commun Signal. 6:121–123. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
O'Brien TP and Lau LF: Expression of the
growth factor-inducible immediate early gene cyr61 correlates with
chondrogenesis during mouse embryonic development. Cell Growth
Differ. 3:645–654. 1992.PubMed/NCBI
|
|
29
|
Chijiiwa M, Mochizuki S, Kimura T, Abe H,
Tanaka Y, Fujii Y, Shimizu H, Enomoto H, Toyama Y and Okada Y: CCN1
(Cyr61) is overexpressed in human osteoarthritic cartilage and
inhibits ADAMTS-4 (Aggrecanase 1) activity. Arthritis Rheumatol.
67:1557–1567. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yu Y, Park YS, Kim HS, Kim HY, Jin YM,
Jung SC, Ryu KH and Jo I: Characterization of long-term in vitro
culture-related alterations of human tonsil-derived mesenchymal
stem cells: Role for CCN1 in replicative senescence-associated
increase in osteogenic differentiation. J Anat. 225:510–518. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang B, Tsang PC, Pate JL and Moses MA: A
role for cysteine-rich 61 in the angiogenic switch during the
estrous cycle in cows: Regulation by prostaglandin F2alpha. Biol
Reprod. 85:261–268. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Klein C: Novel equine
conceptus?endometrial interactions on Day 16 of pregnancy based on
RNA sequencing. Reprod Fertil Dev. May 5–2015.(Epub ahead of
print). PubMed/NCBI
|
|
33
|
Chen X, Liu Y, Xu X and Chen H:
Correlation of Cyr61 and CTGF in placentas from the late
pre-eclamptic pregnancy. J Perinat Med. 40:199–200. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mo FE, Muntean AG, Chen CC, Stolz DB,
Watkins SC and Lau LF: CYR61 (CCN1) is essential for placental
development and vascular integrity. Mol Cell Biol. 22:8709–8720.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Malik AR, Urbanska M, Gozdz A, Swiech LJ,
Nagalski A, Perycz M, Blazejczyk M and Jaworski J: Cyr61, a
matricellular protein, is needed for dendritic arborization of
hippocampal neurons. J Biol Chem. 288:8544–8559. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kipkeew F, Kirsch M, Klein D, Wuelling M,
Winterhager E and Gellhaus A: CCN1 (CYR61) and CCN3 (NOV) signaling
drives human trophoblast cells into senescence and stimulates
migration properties. Cell Adh Migr. 10:163–178. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Draaken M, Proske J, Schramm C, Wittler L,
Bartels E, Nöthen MM, Reutter H and Ludwig M: Embryonic expression
of the cysteine rich protein 61 (CYR61) gene: A candidate for the
development of human epispadias. Birth Defects Res A Clin Mol
Teratol. 88:546–550. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Romereim SM, Summers AF, Pohlmeier WE,
Zhang P, Hou X, Talbott HA, Cushman RA, Wood JR, Davis JS and Cupp
AS: Gene expression profiling of bovine ovarian follicular and
luteal cells provides insight into cellular identities and
functions. Mol Cell Endocrinol. 439:379–394. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen Y, Ni H, Ma XH, Hu SJ, Luan LM, Ren
G, Zhao YC, Li SJ, Diao HL, Xu X, et al: Global analysis of
differential luminal epithelial gene expression at mouse
implantation sites. J Mol Endocrinol. 37:147–161. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu P, Kwok CS, Haththotuwa R, Kotronias
RA, Babu A, Fryer AA, Myint PK, Chew-Graham CA and Mamas MA:
Pre-eclampsia is associated with a twofold increase in diabetes: A
systematic review and meta-analysis. Diabetologia. 59:2518–2526.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hodgins S: Pre-eclampsia as underlying
cause for perinatal deaths: Time for action. Glob Health Sci Pract.
3:525–527. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chaiworapongsa T, Chaemsaithong P, Yeo L
and Romero R: Pre-eclampsia part 1: Current understanding of its
pathophysiology. Nat Rev Nephrol. 10:466–480. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang X, Yan G, Diao Z, Sun H and Hu Y:
NUR77 inhibits the expression of TIMP2 and increases the migration
and invasion of HTR-8/SVneo cells induced by CYR61. Placenta.
33:561–567. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang Y, Diao Z, Su L, Sun H, Li R, Cui H
and Hu Y: MicroRNA-155 contributes to preeclampsia by
down-regulating CYR61. Am J Obstet Gynecol. 202(466): e1–e7.
2010.PubMed/NCBI
|
|
45
|
Huang J, Gao K, Lin J and Wang Q:
MicroRNA-100 inhibits osteosarcoma cell proliferation by targeting
Cyr61. Tumour Biol. 35:1095–1100. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sarkissyan S, Sarkissyan M, Wu Y, Cardenas
J, Koeffler HP and Vadgama JV: IGF-1 regulates Cyr61 induced breast
cancer cell proliferation and invasion. PLoS One. 9:e1035342014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Johnson SK, Stewart JP, Bam R, Qu P,
Barlogie B, van Rhee F, Shaughnessy JD Jr, Epstein J and Yaccoby S:
CYR61/CCN1 overexpression in the myeloma microenvironment is
associated with superior survival and reduced bone disease. Blood.
124:2051–2060. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Shigeoka M, Urakawa N, Nishio M, Takase N,
Utsunomiya S, Akiyama H, Kakeji Y, Komori T, Koma Y and Yokozaki H:
Cyr61 promotes CD204 expression and the migration of macrophages
via MEK/ERK pathway in esophageal squamous cell carcinoma. Cancer
Med. 4:437–446. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Maity G, Mehta S, Haque I, Dhar K, Sarkar
S, Banerjee SK and Banerjee S: Pancreatic tumor cell secreted
CCN1/Cyr61 promotes endothelial cell migration and aberrant
neovascularization. Sci Rep. 4:49952014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Arslan AA, Gold LI, Mittal K, Suen TC,
Belitskaya-Levy I, Tang MS and Toniolo P: Gene expression studies
provide clues to the pathogenesis of uterine leiomyoma: New
evidence and a systematic review. Hum Reprod. 20:852–863. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Di Tommaso S, Massari S, Malvasi A,
Bozzetti MP and Tinelli A: Gene expression analysis reveals an
angiogenic profile in uterine leiomyoma pseudocapsule. Mol Hum
Reprod. 19:380–387. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wallace K, Chatman K, Spencer SK, Johnson
V and LaMarca B: ‘Special research presentation’ endothelin
regulation of cyr61 in uterine leiomyomas. Fertil Steril. 102
Suppl:e1062014. View Article : Google Scholar
|
|
53
|
Zhao Y, Li Q, Katzenellenbogen BS, Lau LF,
Taylor RN, Bagchi IC and Bagchi MK: Estrogen-induced CCN1 is
critical for establishment of endometriosis-like lesions in mice.
Mol Endocrinol. 28:1934–1947. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Klein R, Stiller S and Gashaw I: Epidermal
growth factor upregulates endometrial CYR61 expression via
activation of the JAK2/STAT3 pathway. Reprod Fertil Dev.
24:482–489. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chien W, Kumagai T, Miller CW, Desmond JC,
Frank JM, Said JW and Koeffler HP: Cyr61 suppresses growth of human
endometrial cancer cells. J Biol Chem. 279:53087–53096. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Fournier A, Dossus L, Mesrine S, Vilier A,
Boutron-Ruault MC, Clavel-Chapelon F and Chabbert-Buffet N: Risks
of endometrial cancer associated with different hormone replacement
therapies in the E3 N cohort, 1992–2008. Am J Epidemiol.
180:508–517. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Joy J: Epigenetic regulation of key genes
involved in cervical malignancy. 44:2014.
|
|
58
|
Xie H, Zhao Y, Caramuta S, Larsson C and
Lui WO: miR-205 expression promotes cell proliferation and
migration of human cervical cancer cells. PLoS One. 7:e469902012.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Bartel F, Balschun K, Gradhand E, Strauss
HG, Dittmer J and Hauptmann S: Inverse expression of cystein-rich
61 (Cyr61/CCN1) and connective tissue growth factor (CTGF/CCN2) in
borderline tumors and carcinomas of the ovary. Int J Gynecol
Pathol. 31:405–415. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Lee KB, Byun HJ, Park SH, Park CY, Lee SH
and Rho SB: CYR61 controls p53 and NF-kB expression through
PI3K/Akt/mTOR pathways in carboplatin-induced ovarian cancer cells.
Cancer Lett. 315:86–95. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Lin Y, Xu T, Tian G and Cui M:
Cysteine-rich, angiogenic inducer, 61 expression in patients with
ovarian epithelial carcinoma. J Int Med Res. 42:300–306. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Po LK and Liu GY: Development of uterine
smooth muscle tumour of uncertain malignant potential (STUMP) after
laparoscopic myomectomy of an atypical leiomyoma. J Minim Invasive
Gynecol. 22:S2312015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Joseph NM, Solomon DA, Frizzell N, Rabban
JT, Zaloudek C and Garg K: Morphology and immunohistochemistry for
2SC and FH aid in detection of fumarate hydratase gene aberrations
in uterine leiomyomas from young patients. Am J Surg Pathol.
39:1529–1539. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kubínová K, Mára M, Horák P and Kuzel D:
Genetic factors in etiology of uterine fibroids. Ceska Gynekol.
77:58–60. 2012.In Czech. PubMed/NCBI
|
|
65
|
Hashim Abu H: Aromatase inhibitors for
endometriosis-associated infertility; Do we have sufficient
evidence? Int J Fertil Steril. 10:270–277. 2016.PubMed/NCBI
|
|
66
|
Bhatti M, Arnold A, Ketheeswaran A,
Nesbitt-Hawes E, Deans R and Abbott J: A comparison of examination
and surgical findings in women with endometriosis. J Minim Invasive
Gynecol. 22:S55–S56. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Modotte WP, Modotti CC, Dias DS,
Bueloni-Dias FN and Rodrigues NP: Endometriosis and infertility
setup during hysterossalpingogram. J Minim Invasive Gynecol.
22:S1782015. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Cozzolino M, Nasioudis D, Sisti G and
Coccia ME: Malignant transformation of vaginal endometriosis-a
review of literature. Gynecol Obstet Invest. 82:105–112. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Tobiume T, Kotani Y, Takaya H, Nakai H,
Tsuji I, Suzuki A and Mandai M: Determinant factors of
postoperative recurrence of endometriosis: Difference between
endometrioma and pain. Eur J Obstet Gynecol Reprod Biol. 205:54–59.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Lin K, Zhan H, Ma J, Xu K, Wu R, Zhou C
and Lin J: Silencing of SRA1 regulates ER expression and attenuates
the growth of stromal cells in ovarian endometriosis. Reprod Sci.
24:836–843. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Ohara F, Abdala-Ribeiro HS, Rodrigues FC,
Aldrighi JM and Ribeiro PA: Outcomes of laparoscopic treatment of
rectosigmoid endometriosis: The linear nodulectomy and the
segmental ressection. J Minim Invasive Gynecol. 22:S952015.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Seyhan A, Ata B and Uncu G: The impact of
endometriosis and its treatment on ovarian reserve. Semin Reprod
Med. 33:422–428. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Gashaw I, Stiller S, Böing C, Kimmig R and
Winterhager E: Premenstrual regulation of the pro-angiogenic factor
CYR61 in human endometrium. Endocrinology. 149:2261–2269. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Yanokura M, Banno K, Iida M, Irie H, Umene
K, Masuda K, Kobayashi Y, Tominaga E and Aoki D: MicroRNAS in
endometrial cancer: Recent advances and potential clinical
applications. EXCLI J. 14:190–198. 2015.PubMed/NCBI
|
|
75
|
Djati Widodo MS and Rifa'i M: Role of
MicroRNAs in carcinogenesis that potential for biomarker of
endometrial cancer. Ann Med Surg (Lond). 7:9–13. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Witek Ł, Janikowski T, Bodzek P, Olejek A
and Mazurek U: Expression of tumor suppressor genes related to the
cell cycle in endometrial cancer patients. Adv Med Sci. 61:317–324.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Estimating the world cancer burden: Globocan 2000. Int J Cancer.
94:153–156. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Long Roche K, Angarita AM, Cristello A,
Lippitt M, Haider AH, Bowie JV, Fader AN and Tergas AI: ‘Little big
things’: A qualitative study of ovarian cancer survivors and their
experiences with the health care system. J Oncol Pract.
12:e974–e980. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Yokoyama Y, Futagami M, Watanabe J, Sato
N, Terada Y, Miura F, Sugiyama T, Takano T, Yaegashi N, Kojimahara
T, et al: Redistribution of resistance and sensitivity to platinum
during the observation period following treatment of epithelial
ovarian cancer. Mol Clin Oncol. 2:212–218. 2014.PubMed/NCBI
|